

Abstract
High-throughput sequencing of chloroplast genomes has been used to gain insight into the evolutionary relationships of plant species. In this study, we sequenced the complete chloroplast genomes of four species in the Meconopsis genus: M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea. These plants grow in the wild and are recognized as having important medicinal and ornamental applications. The sequencing results showed that the size of the Meconopsis chloroplast genome ranges from 151864 to 153816 bp. A total of 127 genes comprising 90 protein-coding genes, 37 tRNA genes and 8 rRNA genes were observed in all four chloroplast genomes. Comparative analysis of the four chloroplast genomes revealed five hotspot regions (matK, rpoC2, petA, ndhF, and ycf1), which could potentially be used as unique molecular markers for species identification. In addition, the ycf1 gene may also be used as an effective molecular marker to distinguish Papaveraceae and determine the evolutionary relationships among plant species in the Papaveraceae family. Futhermore, these four genomes can provide valuable genetic information for other related studies.
Subject terms: Plant genetics, Medical genomics
Introduction
The genus Meconopsis belongs to the Papaveraceae family of herb angiosperms and comprises approximately 49 species, 38 of which are found in China1. These plants are mainly distributed in the Himalayan foothills at an elevation of 2500–5500 m and are widely used in Tibetan folk medicine in China2. Detailed records of the medicinal usage of these plants have been written in the famous classic works on traditional Tibetan medicine, such as Jingzhu Materia Medica, Yue Wang Yao Zhen, and Four Medical Codes3. Recently, many kinds of isoquinoline alkaloids have been isolated from plants of the Meconopsis genus, and some have shown bioactivity, such as anti-inflammatory and analgesic activities4. Plants in this genus are also well known for their ornamental flowers and are widely used in horticultural gardening, with names such as fairy grass and Himalayan poppy. These plants are iconic in Tibet and Yunnan and play a significant role in the local Tibetan economy, as they are among the top ten ornamental flowering plants in the region2. Howere, overexploitation and anthropogenic habitat destruction are increasingly threatening the survival of many wild Meconopsis species. Meconopsis punicea has been listed as an endangered species on the China Species Red List5.
To understand the evolutionary relationships of plant species in the Meconopsis genus and in the Papaveraceae family, it is important to obtain genetic information or molecular markers of individual species. This “barcode” can also aid in medicinal usage, for which the accurate identification of species is required, as the regions and sources of species are often complex or unknown6–8 and can affect the efficacy of the final medicinal product.
Recent chloroplast genomic research has provided large quantities of data that are useful for selecting pertinent markers to resolve obscure phylogenetic relationships in seed plants9. At present, nearly 3000 complete chloroplast genomes are available in the NCBI database (https://www.ncbi.nlm.nih.gov/genomes/GenomesGroup.cgi?taxid=2759&opt=plastid)10. However, there is only one sequence from the chloroplast DNA of Meconopsis species in GenBank11.
In this study, we sequenced and assembled the chloroplast genomes of four Meconopsis species using a next-generation sequencing platform. We report the assembly, annotation and analysis of the chloroplast genomes of Meconopsis racemosa, Meconopsis integrifolia (Maxim.) Franch, Meconopsis horridula and Meconopsis punicea. We also constructed phylogenetic trees to perform comparisons among chloroplast genomes published for other plant species in related families. This study expands our understanding of the diversity of chloroplast genomes of Meconopsis species and their evolutionary relationships and provides fundamental data for the genetic engineering of Meconopsis chloroplasts.
Results and Discussion
Chloroplast genome sequencing, assembly and validation
Using the Illumina HiSeq 2000 system, we sequenced the complete chloroplast genomes of four Meconopsis species, M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea. Raw data were generated with an average read length of 150 bp. The complete sequences of the four chloroplast genomes were assembled by both de novo and reference-based assembly. Gaps were validated using PCR-based sequencing with one primer pair (Supplementary Table 1). The final high-quality chloroplast genome sequences were submitted to GenBank (Accession Numbers: M. racemosa, MK533649; M. integrifolia (Maxim.) Franch, MK533647; M. horridula, MK533646; M. punicea, MK533648), and the corresponding genome maps are shown in Fig. 1.
Figure 1.

Chloroplast genome maps of M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea. Genes inside circles are transcribed clockwise, genes outside circles are transcribed counterclockwise. The light gray inner circle corresponds to the AT content, and the dark gray circle corresponds to the GC content. Genes belonging to different functional groups are shown in different colors.
Chloroplast genome structural features and gene content
It was previously reported that the chloroplast genomes of angiosperms are conserved in their genomic structure in terms of gene number and order, although IR expansion or contraction occur frequently12,13. The Meconopsis chloroplast genomes are in accordance with this observation, and their genome structures are similar to those of other Papaveraceae species14. All of the Meconopsis chloroplast genomes display the typical quadripartite structure of angiosperm cpDNA, which consists of a pair of IR regions (51306–51988 bp) separated by an LSC region (82809–83982 bp) and an SSC region (17729–17898 bp). These four chloroplast genomes are highly conserved in gene content, gene order, and intron number. The Meconopsis chloroplast genomes harbor 127 genes, 90 coding proteins, 37 coding tRNAs and 8 coding rRNAs. Some genes are duplicated in the IR region, among which ten are protein-coding genes (rpl2, rpl12, rps12, rps15, rps16, rps19, ndhB, ycf1, ycf15 and ycf2), four are ribosomal RNA genes (rrn4.5, rrn5, rrn16, rrn23) and six are transfer RNA genes (trnL-CAA, trnN-GUU, trnR-ACG, trnA-UGC, trnI-GAU and trnV-GAC) (Table 1). Fifteen protein-coding genes (petB, petD, ndhA, ndhB, atpF, rps12, rps15, rps16, rps19, rpl2, rpl12, rpl16, rpoC1, clpP, and ycf3) contain one or more introns. The A content ranged from 30.4 to 30.5%, the C content ranged from 19.7 to 19.8%, the G content ranged from 18.8 to 19%, the T content ranged from 30.8 to 31%, and the GC content ranged from 38.5 to 38.8%, indicating nearly identical levels among the four Meconopsis chloroplast genomes (Table 2).
Table 1.
Summary of assembly data for the Meconopsis chloroplast genome.
| Species | Meconopsis racemosa | Meconopsis integrifolia (Maxim.) Franch | Meconopsis horridula | Meconopsis punicea |
|---|---|---|---|---|
| Genome size (bp) | 153816 | 151864 | 153785 | 153259 |
| IR (bp) | 51988 | 51306 | 51988 | 51548 |
| LSC (bp) | 83930 | 82809 | 83899 | 83982 |
| SSC (bp) | 17898 | 17749 | 17898 | 17729 |
| Total number of genes | 127 | 127 | 127 | 127 |
| rRNA | 8 | 8 | 8 | 8 |
| tRNA | 37 | 37 | 37 | 37 |
| Protein-coding genes | 90 | 90 | 90 | 90 |
| A % | 30.4 | 30.4 | 30.4 | 30.5 |
| C % | 19.8 | 19.8 | 19.8 | 19.7 |
| G % | 18.9 | 19 | 18.9 | 18.8 |
| T % | 30.9 | 30.8 | 30.9 | 31 |
| G C% | 38.7 | 38.8 | 38.8 | 38.5 |
Table 2.
Chloroplast genome gene content and functional classification in M.
| Category | Group | Genes |
|---|---|---|
| Self-replication | Large subunit of ribosome (LSU) | rpl14, rpl16a, rpl2a,b, rpl2a,b, rpl20, rpl22, rpl23b, rpl23b, rpl32, rpl33, rpl36 |
| Small subunit of ribosome (SSU) |
rps11, rps12a,b, rps14, rps15a,b, rps16a, rps18, rps19a,b, rps2, rps3, rps4, rps7b, rps8 |
|
| DNA dependent RNA polymerase | rpoA, rpoB, rpoC1a, rpoC2 | |
| Ribosome RNA | rrn16b, rrn23b, rrn4.5b, rrn5b | |
| Transfer RNAs (tRNA) |
trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC, trnH-GUG, trnI-CAU, trnI-CAU, trnL-CAAb, trnL-UAG, trnM-CAU, trnN-GUUb, trnP-UGG, trnQ-UUG, trnR-ACGb, trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GACb, trnW-CCA, trnY-GUA trnK-UUUa, trnG-UCCa, trnV-UACa, trnA-UGCa, trnL-UAAa, trnI-GAUa |
|
| Photosynthesis | Photosystem I | psaA, psaB, psaC, psaI, psaJ |
| Photosystem II |
psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ |
|
| NADH dehydrogenase |
ndhAa, ndhBa,b, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK |
|
| Cytochrome b/f complex | petA, petBa, petDa, petG, petL, petN | |
| Subunits of ATP synthase | atpA, atpB, atpE, atpFa, atpH, atpI | |
| Large subunit of rubisco | rbcL | |
| Other genes | Translational initiation factor | infA |
| ATP-dependent protease subunit p gene | clpP a | |
| Maturase | matK | |
| Envelop membrane protein | cemA | |
| Unknown function | Subunit of acetyl-CoA-carboxylase | accD |
| C-type cytochrome synthesis gene | ccsA | |
| Hypothetical chloroplast reading frames | ycf1b, ycf15b, ycf2b, ycf3a, ycf4 |
racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea. aGenes containing introns; bTwo gene copies in IR.
Amino acid abundance and codon usage
Codon usage plays an important role in shaping chloroplast genome evolution. Mutational bias has been reported to have an essential role in this process15. As shown in Supplementary Tables 2–5, the 90 protein-coding genes are encoded by 26338, 26365, 26342 and 26337 codons in the chloroplast genomes of M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea, respectively. Leucine (11.1–9.5%) was the most abundant amino acid among the proteins encoded by the chloroplast genes. Cysteine (1.2–1.7%) was the least abundant amino acid in the proteins encoded by chloroplast genes in the M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea chloroplast genomes. Leucine and isoleucine are the most commonly observed amino acids in the proteins of chloroplast genomes of angioperms16.
We calculated and summarized the codon usage of the chloroplast genomes in these four plants (Fig. 2). The codon UUA, for leucine, occurred at the highest proportion in all four species (27.1–30.3%). There were a total of 711 codons encoding tRNA genes in the M. racemosa, M. integrifolia (Maxim.) Franch and M. horridula chloroplast genomes, but only 704 codons in the tRNA-encoding genes in M. punicea (Supplementary Tables 2–5), indicating that codons ending in U and A were common; perhaps the variation in the tRNA-encoding genes is related to species evolution.
Figure 2.

Codon content of twenty amino acids and stop codons in all protein-coding genes of the chloroplast genomes of four Meconopsis species.
We also calculated the relative synonymous codon usage (RSCU) in the chloroplast genomes of the four species. Usage of the start codon methionine AUG and tryptophan UGG had no bias (RSCU = 1). All preferred relative synonymous codons (RSCU >1) ended with an A or a U, except for UUG (all 4 species), UCC (M. integrifolia (Maxim.) Franch, M. horridula and M. punicea) and UAG (M. integrifolia (Maxim.) Franch and M. punicea) (Supplementary Tables 2–5).
Plastid RNA editing prediction
RNA editing is a generic term comprising a variety of processes that alter the DNA-encoded sequence of a transcribed RNA by inserting, deleting or modifying nucleotides in a transcript17. Chloroplast RNA editing was first discovered in 1991. Nearly 30 years after the discovery of C-to-U editing in plant chloroplasts, the field has recently expanded tremendously in several research directions18. RNA editing provides a way to create transcript and protein diversity19. In higher plants, some chloroplast RNA editing sites are conserved20.
To gain insight into the RNA editing sites in Meconopsis plants, we predicted 92, 78, 84 and 94 RNA editing sites out of 27, 26, 28 and 28 plastid genes in the chloroplast genomes of M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea, respectively, with PREP (Supplementary Tables 6–9). In these four species, the amino acid conversion from S to L was the most frequent type of conversion. As previously reported, with increased amino acids, the conversion from S to L becomes more frequent21. This finding indicated that the evolutionary conservation of RNA editing is essential22,23.
Simple sequence repeats and repetitive sequence analysis
Tandem repeat sequences consisting of 1–6 nucleotide repeat units are known as simple sequence repeats (SSRs), or microsatellites24. SSRs are valuable molecular markers with a high degree of variation within species and have been used in many population genetics and polymorphism investigations. Using the MISA software tool, we analyzed the occurrences and types of SSRs in the four Meconopsis chloroplast genomes. These genomes all have SSRs, and the majority of which are mono- and dinucleotide repeats, which were identified 88 and 29 times, respectively. The mononucleotide repeats were A/T repeats, and 82.8% of the dinucleotide repeats were AT/AT repeats (Table 3). Although the AT richness in the SSRs of the four chloroplast genomes of Meconopsis species was similar to that identified in previous studies, which suggested that SSRs found in the chloroplast genome are generally composed of polythymine (T) or polyadenine (A) repeats25, the number of SSRs differs among the different species (40 in M. racemosa, 33 in M. integrifolia (Maxim.) Franch, 38 in M. horridula and 34 in M. punicea; Table 3). These findings indicate that SSRs can be used as molecular markers to identify these plant species.
Table 3.
Types and numbers of SSRs in the chloroplast genomes of M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea.
| SSR type | Repeat unit | Species | |||
|---|---|---|---|---|---|
| Meconopsis racemosa | Meconopsis integrifolia (Maxim.) Franch | Meconopsis horridula | Meconopsis punicea | ||
| Mono | A/T | 24 | 22 | 23 | 19 |
| Di | AG/CT | 1 | 1 | 1 | 1 |
| AC/GT | 0 | 0 | 0 | 1 | |
| AT/AT | 7 | 4 | 7 | 6 | |
| Tri | AAT/ATT | 2 | 2 | 2 | 2 |
| Tetra | AAAT/ATTT | 3 | 2 | 3 | 2 |
| AACC/GGTT | 1 | 1 | 1 | 1 | |
| AGAT/ATCT | 1 | 1 | 1 | 0 | |
| ATCC/ATGG | 0 | 0 | 0 | 1 | |
| Hexa | AATGAT/ATCATT | 0 | 0 | 0 | 1 |
| AAAAT/ATTTT | 1 | 0 | 0 | 0 | |
More complex and longer repeat sequences may play an important roles in sequence divergence and genomes26. In these four Meconopsis chloroplast genomes, we found that the length of repeated sequences ranged mainly from 30 to 90 bp, similar to the lengths reported in other angiosperm plants25,27,28. The numbers of repeats with at least 30 base pairs (bp) per repeat unit in the M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula, and M. punicea chloroplast genomes are 35, 49, 34 and 29, respectively. The M. racemosa chloroplast genome contains 27 repeats of 30–50 bp, 5 repeats of 51–70 bp, and 3 repeats longer than 90 bp. The M. integrifolia (Maxim.) Franch chloroplast genome contains 16 repeats of 30–50 bp, 12 repeats of 51–70 bp, 2 repeats of 71–90 bp and 19 repeats longer than 90 bp. The M. horridula chloroplast genome contains 25 repeats of 30–50 bp, 6 repeats of 51–70 bp, 1 repeat of 71–90 bp and 2 repeats longer than 90 bp. The M. punicea chloroplast genome contains 26 repeats of 30–50 bp, 1 repeat of 51–70 bp, and 2 repeats longer than 90 bp (Fig. 3).
Figure 3.
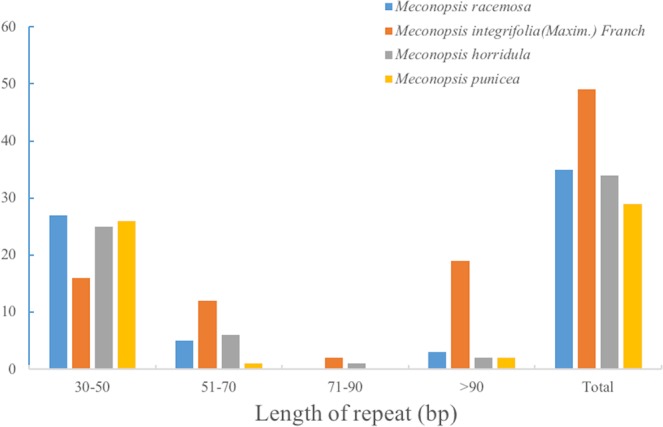
Frequency of repeat sequences of the M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea chloroplast genomes determined by REPuter.
Divergent hotspots in the Meconopsis chloroplast genome
Molecular markers with nucleotide diversity over 1.5% have been reported as highly variable regions that can be used for phylogenetic analysis and species identification in seed plants29,30. Currently, there are few molecular biology-based studies of Meconopsis plants, and there is no uniform molecular marker for species identification31–35.
A SNP (single nucleotide polymorphism) marker is a single base change in a DNA sequence, typically with two possible nucleotide alternatives at a given position36. A total of 176, 2459, 36, 2982 SNPs were found in M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea, respectively. To reveal the sequence divergence levels, the nucleotide variability values within 800 bp in all four chloroplast genomes were calculated with DnaSP 6.10.03 software. The values ranged from 0 to 0.07, revealing slight differences among the genomes. For example, the p-distance between M. racemosa and each of M. integrifolia (Maxim.) Franch, M. horridula and M. punicea is 0.016, 0.001 and 0.018, respectively. These divergence hotspot regions can provide information for marker development for phylogenetic analyses of Meconopsis species. Overall, the results reveal higher divergence in noncoding regions than in coding regions. Using whole chloroplast genomes, we found that some regions differ among the four species, such as rps16, trnC-GCA, trnD-GCU, trnT-GGU, rps15, accD-PsaI and petA (Fig. 4a). The coding regions with marked differences include the matK, rpoC2, petA, ndhF and ycf genes (Fig. 4b). These genes could be utilized as potential phylogenetic markers to reconstruct the phylogeny in this genus. Qu Yan et al. reported that the ndhF gene could not be used to distinguish M. racemosa from M. horridula37. However, our present study shows that the sequence of the ndhF gene in the chloroplast genome differs between these two species is distinct.
Figure 4.

Nucleotide variability (%) values between pairs of the four Meconopsis species. (a) Using four species whole genomes; (b) Using four species coding regions.
Divergent hotspots of chloroplast genomes have been used to identify species in other plants of the Papaveraceae family. Jianguo Zhou et al. used ycf1, rpoB-trnC, trnD-trnT, petA-psbJ, psbE-petL and ccsA-ndhD sequences in the chloroplast genome to distinguish Papaver orientale and Papaver rhoeas14. Zhe Zhang et al.38 analyzed the phylogeny of 15 species from the Papaveraceae family based on the nuclear gene ITS sequence, the chloroplast gene rbcL sequence, and the combined sequences of these genes.
Comparisons of the chloroplast genomes among nine species in the Papaveraceae family
We compared the 9 known chloroplast genome sequences of species in the Papaveraceae family (M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula, M. punicea, Macleaya microcarpa (MH394383.1), Coreanomecon hylomeconoides (KT274030.1), Papaver somniferum (KU204905.1), Papaver rhoeas (MF943221.1) and Papaver orientale (MF943222.1)). The results indicated that species with the largest chloroplast genome is the M. microcarpa (161118 bp) and that with the smallest is M. integrifolia (Maxim.) Franch genome (151864 bp) (Table 1). The M. microcarpa (161118 bp) genome was used as the reference genome.
Next, we used the online program mVISTA to analyze gene order and content in the chloroplast genome. We found that the gene order and contents of the Meconopsis plants are similar to those of other members of the Papaveraceae family (Fig. 5). Similar to other plant species, all Meconopsis species have conserved chloroplast genomes, their coding regions are more conserved than their noncoding regions, and their IR regions are more conserved than their LSC and SSC regions16,39,40.
Figure 5.
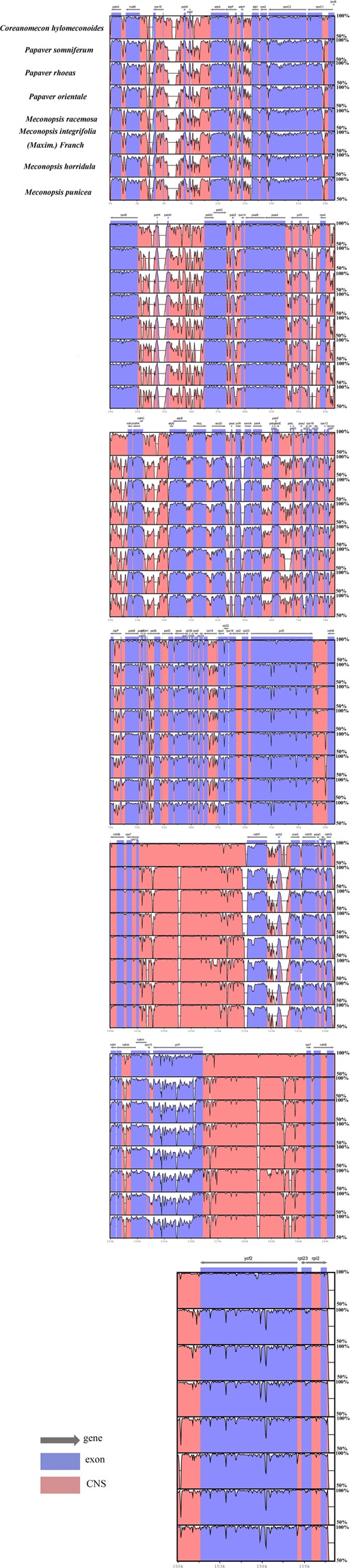
Sequence identity plot comparing the eight chloroplast genomes with Macleaya microcarpa as a reference by using mVista. Pink bars represent noncoding sequences (CNS), and white peaks represent genomic differences. The y-axis represents the percentage identity (shown: 50–100%).
Altitude and plant distribution
Altitude influences ecological factors such as water and temperature, which affects plant genetic variation and population differentiation41. In this study, the plant materials of M. racemosa and M. integrifolia (Maxim.) Franch were mainly collected from the Bayan Har mountains, Qinghai Province. This region has a cold continental climate with an average altitude of over 5000 m. The plant materials of M. horridula were collected from Matuo Country, Guoluo Tibetan Autonomous Prefecture, Qinghai Province. This region has an alpine grassland climate with an average annual temperature of −4 °C and an average altitude of over 4000 m. The plant materials of M. punicea were mainly collected in Chindu Country, Qinghai Province. This region has an average altitude of over 4000 m. Studies have shown that the evolutionary relationships of plants are affected by altitude42,43. The plant materials used in this study were collected in the same area but at different altitudes: M. racemosa 4232 m; M. integrifolia (Maxim.) Franch, 4695 m; M. horridula, 4289 m; and M. punicea, 4639 m. According to traditional plant morphology taxonomy, M. racemosa is more closely related to M. horridula than to other Meconopsis species and is more distantly related to M. integrifolia (Maxim.) Franch and M. punicea44, which is consistent with both the phylogenetic results of this study and the altitudes of their distributions. Although they are distributed in the same region, there is evident genetic isolation among them. We speculate that altitude may be an important ecological factor that affects the evolution of Meconopsis plants.
Phylogenetic analysis
With improvements and advancements in techniques, increasing numbers of chloroplast genome sequences have been used to reconstruct plant phylogenies45. To identify the phylogenetic positions of M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea within the Meconopsis genus, Bayesian inference (BI) and maximum likelihood (ML) methods of phylogenetic analysis were performed based on 90 protein-coding gene datasets from 40 plant taxa, with Sabia yunnanensis and Nelumbo nucifera used as outgroups. Both the BI and ML trees have similar phylogenetic topologies, and most nodal support values were high (Fig. 6). Using this reconstructions, M. racemosa, M. racemosa (MH394401)11 and M. horridula were grouped together, as were M. integrifolia (Maxim.) Franch and M. punicea. These species are closely related to the Papaver genus within the Papaveraceae family.
Figure 6.

Phylogenetic tree reconstruction of the 42 species inferred from Bayesian inference (BI) and maximum likelihood (ML) based on 90 protein-coding genes. Numbers above the lines represent BI/ML posterior probabilities.
In addition, we found that M. racemosa, M. horridula and M. racemosa (MH394401)11 were grouped together. For several years, the delimitation of M. racemosa and M. horridula in the genus has been highly controversial46. Fedd, Kingdon-Ward and Prain et al. considered M. racemosa and M. horridula to be the same species46. However, in Tibetan Flora, M. racemosa is described as a variant of M. horridula. M. racemosa and M. racemosa (MH394401)11 were distributed on different branches but are the same species. Incomplete lineage sorting, insufficient informative characters, hybridization or plastid capture could be responsible for the incongruent phylogenetic positions of this species47,48.
We used the five gene markers (matK, rpoC2, petA, ndhF and ycf1 genes), screened by divergent hotspots in the Meconopsis chloroplast genomes, to construct five phylogenetic trees of these four Meconopsis plants and five other plants from the Papaveraceae family (P. somniferum, P. rhoeas, P. orientale, Macleaya microcarpa and Coreanomecon hylomeconoides) using Decaisnea insignis, Euptelea pleiosperma and Nuphar advena as outgroups (Fig. 7 and Supplementary Figs 1–4). The results showed that M. racemosa, M. racemosa (MH394401)11 and M. horridula are grouped together and that M. integrifolia (Maxim.) Franch and M. punicea are grouped together. Among the five genes, the rpoC2 gene is not a suitable for potential DNA barcoding of Meconopsis plants, and the ycf1 gene has the highest node support value in the phylogenetic tree, which is consistent with previous reports that have used ycf1 to distinguish unknown Papaveraceae plants14,49. In Tibetan Flora, M. racemosa is described as a variant of M. horridula on account of the similar morphological characterization of these taxa and the consistent ITS sequence. However, Dou et al.35, using the ITS2 sequence, and Ni et al.34, using the psbA-trnH sequence, constructed an evolutionary trees and found that these taxa clustered in different branches.
Figure 7.

Phylogenetic tree reconstruction of the 13 species inferred from maximum likelihood (ML), based on the ycf1 genes.
The chloroplast genome usually contains uniparentally inherited DNA, which is well suited for studying the evolutionary history of plants, such as dating a common ancestor50. Yuan et al. used the chloroplast genome sequence of trnL-trnF and found that M. punicea is the mother of the hybrid species Meconopsis × cookei (Papaveraceae) and that M. quintuplinervia is the father33.
Conclusions
In this study, we used the Illumina HiSeq 2000 system to sequence the complete chloroplast genomes of four Meconopsis species: M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea. We demonstrate that these four Meconopsis species are divided into two groups, with M. racemosa and M. horridula in one group and M. integrifolia (Maxim.) Franch and M. punicea in the other. By comparing the chloroplast genome sequences, we were able to retrieve all genetic resources, including SNPs, SSRs, repetitive sequence, codon usage, RNA editing prediction, ‘hotspot’ regions and phylogenomic analysis. These resources will provide chloroplast genome molecular markers for the identification of these Meconopsis species. We also used four hotspot genes (matK, petA, ndhF and ycf1) to construct phylogenetic trees and clearly distinguish these species.
With the development of plant science, plastid transformation is becoming an important tool. The limited availability of complete chloroplast genomic information is one of the major factors preventing the extension of this technology to valuable plants. The Meconopsis chloroplast genome data obtained in this study could be applied in biotechnology and provide useful information for designing transformation vectors in the future.
Materials and Methods
Plant material and DNA extraction
The plant materials used in this study were seeds collected from M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea in Qinghai Province. All samples were identified by Professor Junhua Du, who is affiliated with Qinghai Normal University. Total genomic DNA was isolated from seeds using the Mag-MK Plant Genomic DNA extraction kit (Sangon Biotech, Shanghai, China), and DNA quality was assessed based on spectrophotometry and electrophoresis in a 1% (w/v) agarose gel. Total DNA samples were chosen for Illumina 2000 sequencing.
Chloroplast genome assemblage and annotation
For these four species, the high-throughput sequencing data were qualitatively assessed and assembled using NOVOPlasty 2.6.3. Gaps in the cpDNA sequences were filled by PCR amplification and Sanger sequencing. The annotations of the chloroplast genomes were performed with Geneious 8.0.4, DOGMA51, CPGAVAS52 and CPGAVAS253 followed by manual correction. The tRNAs were verified by the online tRNAscan-SE 1.21 search server. All the annotations were manually checked against the references (NC_029434.1 and NC_031446.1). The genome maps were drawn by OGDRAW. The entire chloroplast genome sequences of M.racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea, along with the gene annotations, were submitted to GenBank (Accession Numbers: M. racemosa, MK533649; M. integrifolia (Maxim.) Franch, MK533647; M. horridula, MK533646; M. punicea, MK533648).
Codon usage
Codon usage was determined for all protein-coding genes. The relative synonymous codon usage (RSCU) values and codon usage were determined with MEGA7, which was used to reveal the characteristics of the variation in synonymous codon usage54.
Simple sequence repeats and repetitive sequence analysis
Chloroplast microsatellites were identified in a high-quality sequence of clusterbean by using the MISA Perl script55. The minimum numbers for the SSR motifs were 10, 5, 4, 3, 3 and 3 for mono-,di-,tri-,tetra-,penta-,and hexanucleotide repeats, respectively. REPuter was used to identify forward repeats, reserve sequences, complementary and palindromic sequences, with a minimum repeat size of 30 bp and 90% sequence identity56.
Prediction of RNA editing sites
Twenty-eight protein-coding genes of M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea were used to predict potential RNA editing sites using the Predictive RNA Editor for Plants (PERP) suite (http://prep.unl.edu) with a cutoff value of 0.8.
Genome comparison
MAFFT was used to align the chloroplast genomes57. The complete chloroplast genomes of M. racemosa, M. integrifolia (Maxim.) Franch, M. horridula and M. punicea were compared using mVISTA58.
Divergent hotspots identification
The M. racemosa, M. integrifolia(Maxim.) Franch, M. horridula and M.punicea chloroplast genome sequences were aligned using MAFFT and were manually adjusted using Geneious 8.0.4. To analyze nucleotide diversity, we conducted a sliding window analysis using DnaSP version 6.10.03. software59. The window length was set to 800 bp, and the step size was 200 bp.
Phylogenetic analysis
The chloroplast genome sequences of M. racemosa, M. integrifolia(Maxim.) Franch, M. horridula, M. punicea and those of 38 other species were collected from NCBI (Supplementary Table 10) were used for phylogenetic analysis. All of the coding sequences from the 42 species were aligned with the MAFFT method based on codons by Geneious 8.0.4. The best nucleotide substitution model (GTR + G + I) was tested, and a maximum likelihood (ML) tree (1000 bootstrap replicates) was constructed with RAxML software60. BI analyses were conducted using GPU MrBayes. The GTR + I + G substitution model was used for BI. In the BI analyses, two simultaneous runs of 10000000 generations were conducted for the matrix. Each set was sampled every 1000 generations with a burn-in of 25%. The matK, rpoC2, petA, ndhF and ycf1 gene sequences of M. racemosa, M. integrifolia(Maxim.) Franch, M. horridula, M. punicea and 9 other species were collected from NCBI. Maximum likelihood (ML) analyses were conducted using RAxML software with the GTR model61.
Supplementary information
Acknowledgements
This work was supported by grants from the Natural Science Foundation of Tianjin (No. 18JCQNJC14000), the Tianjin City High School Science & Technology Fund Planning Project (No. 20130203), Qinghai Science and Technology Project (No. 2014-HZ-815) and the Ph.D. Candidate Research Innovation Fund of Nankai University. We thank the Guangzhou Gene Denovo Biotechnology Company for assisting with the sequencing analysis.
Author Contributions
X.-X.L., B.-B.X. and Y.W. designed the experiment and drafted and revised the manuscript. W.T., C.-G.Z. and X.-X.T. analyzed the data. J.-Q.S., J.-H.D. and M.Z. prepared the plant materials and collected the samples. All authors reviewed the manuscript.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Xiaoxue Li and Wei Tan contributed equally.
Change history
10/17/2019
An amendment to this paper has been published and can be accessed via a link at the top of the paper.
Contributor Information
Beibei Xiang, Email: xiangbeibei03230@163.com.
Yong Wang, Email: wangyong@nankai.edu.cn.
Supplementary information
Supplementary information accompanies this paper at 10.1038/s41598-019-47008-8.
References
- 1.Wang B, Song XH, Cheng CM. Advance in Ethnobotanical Investigation on Meconopsis. Chinese Academic Medical Magazine of Organisms. 2003;01:39–45. [Google Scholar]
- 2.Guo Z, et al. Chemical constituents from a Tibetan medicine Meconopsis horridula. China Journal of Chinese Materia Medica. 2014;39:1152–1156. [PubMed] [Google Scholar]
- 3.Zhao Z, et al. Peng. Advances in studies on the classification, chemical composition and pharmacological action of Meconopsis as Tibetan medicines. China. Pharmacy. 2016;27:4391–4394. [Google Scholar]
- 4.Chang Y, Wang XL, Tang XY, Yuan LY, Chen LH. A New Alkaloid from Meconopsis horridula. Natural Product Research and Development. 2017;29:731–734. [Google Scholar]
- 5.Qu, Y. & Ou, Z. The research advancement on the genus Meconpsis. Northern Horticulture 191–194 (2012).
- 6.Wang B, Song XH, Cheng CM, Yang JS. Studies on species of Meconopsis as Tibetan medicines. Chinese Wild Plant Resources. 2003;22:45–48. [Google Scholar]
- 7.Fan Y, et al. Effect of Meconopsis racemosa alcohol extract on proliferation of K562 cells and its mechanism. Journal of Chinese Medicinal Materials. 2013;36:1143–1146. [Google Scholar]
- 8.Guo, Z. Q. Study on the anti-myocardial ischemic effect and chemical composition of Meconopsis horridula as Tibetan medicine, Beijing University Of Chinese Medicine (2014).
- 9.Luo J, et al. Comparative chloroplast genomes of photosynthetic orchids: insights into evolution of the Orchidaceae and development of molecular markers for phylogenetic applications. Plos One. 2014;9:e99016. doi: 10.1371/journal.pone.0099016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ni LH, Zhao ZL, Xu HX, Chen SL, Dorje G. Chloroplast genome structures in Gentiana (Gentianaceae), based on three medicinal alpine plants used in Tibetan herbal medicine. Current Genetics. 2016;63:1–12. doi: 10.1007/s00294-016-0631-1. [DOI] [PubMed] [Google Scholar]
- 11.Zeng CX, et al. Genome skimming herbarium specimens for DNA barcoding and phylogenomics. Plant Methods. 2018;14:43. doi: 10.1186/s13007-018-0300-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chen HM, et al. Sequencing and analysis of Strobilanthes cusia (Nees) Kuntze chloroplast Genome revealed the rare simultaneous contraction and expansion of the inverted repeat region in Angiosperm. Frontiers in Plant Science. 2018;9:324. doi: 10.3389/fpls.2018.00324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chang CC, et al. The chloroplast genome of Phalaenopsis aphrodite (Orchidaceae): Comparative analysis of evolutionary rate with that of grasses and its phylogenetic implications. Molecular Biology Evolution. 2006;23:279. doi: 10.1093/molbev/msj029. [DOI] [PubMed] [Google Scholar]
- 14.Zhou JG, et al. Complete chloroplast Genomes of Papaver rhoeas and Papaver orientale: molecular structures, comparative analysis, and phylogenetic analysis. Molecules. 2018;23:437. doi: 10.3390/molecules23020437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Li B, Lin FR, Huang P, Guo WY, Zheng YQ. Complete chloroplast Genome sequence of Decaisnea insignis: Genome organization, Genomic resources and comparative analysis. Sci Rep. 2017;7:10073. doi: 10.1038/s41598-017-10409-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu X, Li Y, Yang H, Zhou B. Chloroplast Genome of the folk medicine and vegetable plant Talinum paniculatum (Jacq.) Gaertn.: gene organization, comparative and phylogenetic analysis. Molecules. 2018;23:857. doi: 10.3390/molecules23040857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mower JP. The PREP suite: predictive RNA editors for plant mitochondrial genes, chloroplast genes and user-defined alignments. Nucleic Acids Res. 2009;37:W253–259. doi: 10.1093/nar/gkp337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lenz H, Hein A, Knoop V. Plant organelle RNA editing and its specificity factors: enhancements of analyses and new database features in PREPACT 3.0. BMC Bioinformatics. 2018;19:255. doi: 10.1186/s12859-018-2244-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bundschuh R, Altmuller J, Becker C, Nurnberg P, Gott JM. Complete characterization of the edited transcriptome of the mitochondrion of Physarum polycephalum using deep sequencing of RNA. Nucleic Acids Res. 2011;39:6044–6055. doi: 10.1093/nar/gkr180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zeng WH, Liao SC, Chang CC. Identification of RNA editing sites in chloroplast transcripts of Phalaenopsis aphrodite and comparative analysis with those of other seed plants. Plant Cell Physiol. 2007;48:362–368. doi: 10.1093/pcp/pcl058. [DOI] [PubMed] [Google Scholar]
- 21.Luo J, et al. Comparative chloroplast genomes of photosynthetic orchids: insights into evolution of the Orchidaceae and development of molecular markers for phylogenetic applications. PLoS One. 2014;9:e99016. doi: 10.1371/journal.pone.0099016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Magdalena GN, Ewa F, Wojciech P. Cucumber, melon, pumpkin, and squash: Are rules of editing in flowering plants chloroplast genes so well known indeed? Gene. 2009;434:0–8. doi: 10.1016/j.gene.2008.12.017. [DOI] [PubMed] [Google Scholar]
- 23.Huang, Y. Y., Antonius, J. M. M. & Matzke, M. Complete sequence and comparative analysis of the chloroplast Genome of Coconut Palm (Cocos nucifera). Plos One8, e74736 (2013). [DOI] [PMC free article] [PubMed]
- 24.Kaila T, et al. Chloroplast Genome sequence of Clusterbean (Cyamopsis tetragonoloba L.): Genome structure and comparative analysis. Genes. 2017;8:212. doi: 10.3390/genes8090212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Li YG, Xu WQ, Zou WT, Jiang DY, Liu XH. Complete chloroplast genome sequences of two endangered Phoebe (Lauraceae) species. Bot Stud. 2017;58:37. doi: 10.1186/s40529-017-0192-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Smith TC. Chloroplast evolution:secondary dispatch symbiogenesis and multiple losses. Current Biology. 2002;12:0–0. doi: 10.1016/s0960-9822(01)00675-3. [DOI] [PubMed] [Google Scholar]
- 27.Greiner S, et al. The complete nucleotide sequences of the five genetically distinct plastid genomes of Oenothera, subsection Oenothera: I. sequence evaluation and plastome evolution. Nucleic Acids Res. 2008;36:2366–2378. doi: 10.1093/nar/gkn081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Song Y, et al. Chloroplast Genomic Resource of Paris for Species Discrimination. Sci Rep. 2017;7:3427. doi: 10.1038/s41598-017-02083-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sarkinen T, George M. Predicting plastid marker variation: can complete plastid genomes from closely related species help? PLoS One. 2013;8:e82266. doi: 10.1371/journal.pone.0082266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Korotkova N, Nauheimer L, Hasmik TV, Allgaier M, Borsch T. Variability among the most rapidly evolving plastid genomic regions is lineage-specific: implications of pairwise genome comparisons in Pyrus (Rosaceae) and other angiosperms for marker choice. Plos One. 2014;9:e112998. doi: 10.1371/journal.pone.0112998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kim K, et al. Complete chloroplast and ribosomal sequences for 30 accessions elucidate evolution of Oryza AA genome species. Sci Rep. 2015;5:15655. doi: 10.1038/srep15655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yuan CC, Li PX, Wang YF, Shi SH. The confirmation of putative natural hybrid species Meconopsis × cooei G. Taylor (Papaveraceae) based on nuclear ribosomal DNA ITS region sequence. Acta Agrestia Sinica. 2004;31:901–907. [PubMed] [Google Scholar]
- 33.Yuan CC, He XB, Yuan QM, Shi SH. Genetic relationship between a natural hybrid Meconopsis × cookei (Papaveraceae) and its parents based on cpDNA trnL-trnF region sequence. Acta Botanica Yunnanica. 2007;29:103–108. [Google Scholar]
- 34.Ni LH, Zhao Zl, Meng QW, GAAWE D. & MI, M. Identification of Tibetan medicinal plants of Meconopsis Vig. using ITS and psbA-trnH sequence. Chinese Traditional and Herbal. Drugs. 2014;45:541–545. [Google Scholar]
- 35.Dou RK, et al. Identification and analysis of Corydalis boweri, Meconopsis horridula and their close related species of the same genus by using ITS2 DNA barcode. China. Journal of Chinese Materia Medica. 2015;40:1453. [PubMed] [Google Scholar]
- 36.Vignal A, Milan D, SanCristobal M, Eggen A. A review on SNP and other types of molecular markers and their use in animal genetics. Genet Sel Evol. 2002;34:275–305. doi: 10.1051/gse:2002009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Qu Y, Zhao WY, Ou Z, Leng QS, Xiong J. Analysis of chloroplast gene ndhF and rbcL sequences of Tibetan medicine plants of Meconopsis. Journal of Central South University of Forestry Technology. 2018;38:90–95. [Google Scholar]
- 38.Zhang Z, Kong Y, Li Y, Wang XY, Liu B. Phylogeny of some Papaveraceae plants in Xinjiang based on DNA barcoding technology. Arid Zone Research. 2014;31:322–328. [Google Scholar]
- 39.Olga K, Ralph B. Elimination of deleterious mutations in plastid genomes by gene conversion. The Plant Journal. 2006;46:85–94. doi: 10.1111/j.1365-313X.2006.02673.x. [DOI] [PubMed] [Google Scholar]
- 40.Ni L, Zhao Z, Xu H, Chen S, Dorje G. The complete chloroplast genome of Gentiana straminea (Gentianaceae), an endemic species to the Sino-Himalayan subregion. Gene. 2016;577:281–288. doi: 10.1016/j.gene.2015.12.005. [DOI] [PubMed] [Google Scholar]
- 41.Zhao C. The plasticity of altitudes to the morphological characteristics of salicornia. Acta Agrestia Sinica. 2015;23:897–904. [Google Scholar]
- 42.Winkworth, R. C., Wagstaff, S. J., Glenny, D., Lockhart, P. J. J. O. D. & Evolution. Evolution of the New Zealand mountain flora: Origins, diversification and dispersal. 5, 237–247 (2005).
- 43.Wei L, Wei C. Effects of phytogenetic structure and environmental factors on plant community in changbai mountain. Journal of Arid Land Resources and Environment. 2013;27:63–68. [Google Scholar]
- 44.yi WZ, Zhuang X. Study on the classification system of Meconopsis. Plant Diversity. 1980;2:371–381. [Google Scholar]
- 45.Li B, Zheng Yq. Dynamic evolution and phylogenomic analysis of the chloroplast genome in Schisandraceae. Scientific Reports. 2018;8:9285. doi: 10.1038/s41598-018-27453-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Xie, S. j., Yang, J. w., Xu, W. y. & Yuan, C. c. In 2006 Chinese symposium on physiological ecology and molecular biology of plant stress.
- 47.Maddison WP, Knowles LL. Inferring phylogeny despite incomplete lineage sorting. Syst Biol. 2006;55:21–30. doi: 10.1080/10635150500354928. [DOI] [PubMed] [Google Scholar]
- 48.Yang HM, Zhang YX, Yang JB, Li DZ. The monophyly of Chimonocalamus and conflicting gene trees in Arundinarieae (Poaceae: Bambusoideae) inferred from four plastid and two nuclear markers. Mol Phylogenet Evol. 2013;68:340–356. doi: 10.1016/j.ympev.2013.04.002. [DOI] [PubMed] [Google Scholar]
- 49.Jeon, J. H. & Kim, S. C. Comparative analysis of the complete Chloroplast Genome sequences of three closely related East-Asian wild roses (Rosa sect. Synstylae; Rosaceae). Genes10 (2019). [DOI] [PMC free article] [PubMed]
- 50.Jheng CF, et al. The comparative chloroplast genomic analysis of photosynthetic orchids and developing DNA markers to distinguish Phalaenopsis orchids. Plant Science. 2012;190:62–73. doi: 10.1016/j.plantsci.2012.04.001. [DOI] [PubMed] [Google Scholar]
- 51.Wyman SK, Jansen RK, Boore JL. Automatic annotation of organellar genomes with DOGMA. Bioinformatics. 2004;20:3252–3255. doi: 10.1093/bioinformatics/bth352. [DOI] [PubMed] [Google Scholar]
- 52.Liu C, et al. CpGAVAS, an integrated web server for the annotation, visualization, analysis, and GenBank submission of completely sequenced chloroplast genome sequences. Bmc Genomics. 2012;13:715. doi: 10.1186/1471-2164-13-715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Shi LC, et al. CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res. 2019;1:1–9. doi: 10.1093/nar/gkz345/5486746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology Evolution. 2016;33:1870. doi: 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Thiel T, Michalek W, Varshney R, Graner A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.) Theoretical Applied Genetics. 2003;106:411–422. doi: 10.1007/s00122-002-1031-0. [DOI] [PubMed] [Google Scholar]
- 56.Kurtz S, et al. REPuter: the manifold applications of repeat analysis on a genomic scale. Nucleic Acids Research. 2001;29:4633–4642. doi: 10.1093/nar/29.22.4633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kazutaka K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology Evolution. 2013;30:772–780. doi: 10.1093/molbev/mst010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Dubchak I, Ryaboy DV. VISTA Family of Computational Tools for Comparative Analysis of DNA Sequences and Whole Genomes. Methods in Molecular Biology. 2006;338:69–89. doi: 10.1385/1-59745-097-9:69. [DOI] [PubMed] [Google Scholar]
- 59.Rozas, J. et al. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Molecular Biology Evolution34 (2017). [DOI] [PubMed]
- 60.Alexandros S. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30:1312–1313. doi: 10.1093/bioinformatics/btu033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yang H, Li T, Dang K, Bu W. Compositional and mutational rate heterogeneity in mitochondrial genomes and its effect on the phylogenetic inferences of Cimicomorpha (Hemiptera: Heteroptera) Bmc Genomics. 2018;19:264. doi: 10.1186/s12864-018-4650-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.