

Abstract
The human endometrium undergoes repetitive regeneration cycles in order to recover the functional layer, shed during menses. The basal layer, which remains in charge of endometrial regeneration in every cycle, contains adult stem or progenitor cells of epithelial and mesenchymal lineage. Some pathologies such as adenomyosis, in which endometrial tissue develops within the myometrium, originate from this layer. It is well known that the balance between adenosine triphosphate (ATP) and adenosine plays a crucial role in stem/progenitor cell physiology, influencing proliferation, differentiation, and migration. The extracellular levels of nucleotides and nucleosides are regulated by the ectonucleotidases, such as the nucleoside triphosphate diphosphohydrolase 2 (NTPDase2). NTPDase2 is a membrane-expressed enzyme found in cells of mesenchymal origin such as perivascular cells of different tissues and the stem cells of adult neurogenic regions. The aim of this study was to characterize the expression of NTPDase2 in human nonpathological cyclic and postmenopausic endometria and in adenomyosis. We examined proliferative, secretory, and atrophic endometria from women without endometrial pathology and also adenomyotic lesions. Importantly, we identified NTPDase2 as the first marker of basal endometrium since other stromal cell markers such as CD10 label the entire stroma. As expected, NTPDase2 was also found in adenomyotic stroma, thus becoming a convenient tracer of these lesions. We did not record any changes in the expression levels or the localization of NTPDase2 along the cycle, thus suggesting that the enzyme is not influenced by the female sex hormones like other previously studied ectoenzymes. Remarkably, NTPDase2 was expressed by the Sushi Domain containing 2 (SUSD2)+ endometrial mesenchymal stem cells (eMSCs) found perivascularly, rendering it useful as a cell marker to improve the isolation of eMSCs needed for regenerative medicine therapies.
Electronic supplementary material
The online version of this article (10.1007/s11302-019-09656-3) contains supplementary material, which is available to authorized users.
Keywords: Purinergic signaling, Adenomyosis, SUSD2, MSC, Ectonucleotidases
Introduction
The human endometrium, the innermost layer of the uterus, is a dynamic tissue undergoing repetitive regeneration cycles in order to recover the tissue shed during menses. The endometrium consists of two layers: the upper layer, known as functionalis, which contains the surface epithelium, the glandular epithelium, and a vascularized stroma; and the deeper basal layer (the basalis), formed by the basal region of the glands, a dense stroma, and blood vessels. The functionalis is lost during the menstrual phase while the basalis remains in charge of endometrial regeneration in every cycle, throughout the woman’s reproductive life. There is no marker of endometrial basal stroma although it would be of interest to study pathologies that arise from this zone such as adenomyosis, which consists of the presence of endometrial tissue within the myometrium. As in other highly proliferative tissues, stem/progenitor cells are responsible for this tissue regeneration. Adult stem or progenitor cells of endometrium reside in the basalis and are present in cyclic and in postmenopausal endometria [1]. The N-cadherin has recently been identified as a marker of human endometrial epithelial progenitors (eEPs) [2]. N-cadherin+ eEPs reside in the bases of endometrial glands in the basalis, adjacent to the myometrium, where the stage-specific embryonic antigen-1 (SSEA-1) is also expressed [3].
Endometrial mesenchymal stem cells (eMSCs) have been identified in both functionalis and basalis of human endometrium, using a combination of two pericyte cell markers: platelet-derived growth factor receptor beta (PDGFRβ)/CD140b and CD146 [4]. In 2012, Masuda et al. identified the perivascular marker Sushi Domain containing 2 (SUSD2) (previously known as W5C5) as an individual marker of another population of eMSCs [5]. So to date, the eMSC niche has been identified perivascularly, as in other organs [6, 7], with PDGFRβ+CD146+ cells (α-SMA+ pericytes) wrapping the endothelial cells, and the SUSD2+ cells surrounding the pericytes.
Recently, numerous studies have been carried out on the role of the purinergic signaling in MSC function [8, 9]. Extracellular nucleotides and their metabolites, the effectors of the purinergic signaling, have been included in the list of molecular signals produced by MSCs. In fact, adenosine triphosphate (ATP) and its derivatives, such as adenosine, are involved in the regulation of the proliferation, differentiation, motility, migration, death, and immunomodulatory capacity of MSCs [8, 10–21]. All of these effects are possible through the presence of purinergic receptors, specific for adenosine (P1 receptors) and nucleotides (P2 receptors), in the plasma membrane of MSCs. In addition, nucleotide and nucleoside levels in the extracellular milieu of MSC are controlled by the presence of nucleotide-hydrolyzing enzymes called ectonucleotidases, acting alone or sequentially, on the MSC cell surface. Four ectonucleotidase families have been identified: (i) ectonucleoside triphosphate diphosphohydrolase family (E-NTPDase), with NTPDase1 (CD39), 2, 3, and 8 as plasma membrane-bound members, which hydrolyses extracellular ATP to adenosine diphosphate (ADP), and ADP to adenosine monophosphate (AMP); (ii) ectonucleotide pyrophosphatase/phosphodiesterase family (E-NPP), which hydrolyses ATP to AMP; (iii) ecto-5′-nucleotidase (CD73), which converts AMP to adenosine; and (iv) alkaline phosphatase family (ALP), able to hydrolyze adenine nucleotides and pyrophosphate [22, 23].
Besides CD73, which is a cell surface marker defining MSCs, other ectonucleotidases have been identified in relation to MSCs, such as NTPDase1 in gums and NPP1 and NPP3 in bone marrow. In addition, NTPDase2, 3, 5, and 6 mRNAs have been detected in dermal and limbal MSCs [20].
NTPDase2 is a cell surface-located enzyme that mainly catalyzes the dephosphorylation of ATP with minimal ADPase activity [23]. This enzyme has been identified in different tissues, in cells of mesenchymal origin, including microvascular pericytes and adventitial cells of muscularized vessels [24], as well as in adventitia of glomerular arterioles of the kidney [25] and portal fibroblasts of the liver [26]. Moreover, NTPDase2 is highly expressed in stem/progenitor cells in adult neurogenic regions, such as the type B cells of the subventricular zone and the rostral migratory stream, and the residual radial glial cells of the hippocampal dental gyrus in mice [27, 28], where NTPDase2 is probably involved in the progenitor cell homeostasis, limiting progenitor cell proliferation and expansion under baseline conditions [29]. In murine uterus, NTPDase2 is immunolocalized at the connective tissue cells lining endometrial glands and surrounding the myometrial fibers [30]. To date, although the presence of other ectonucleotidases has been well characterized in human endometrium, with variations in the expression along the cycle, studies on NTPDase2 in the human endometrium have yet to be performed. Moreover, P2Y receptors, activated by ADP (and UDP), which are the products of the hydrolytic activity of NTPDase2, are present in human endometrium and are also described in MSCs [8, 21, 31–33].
So, the eMSCs are pericytes and perivascular cells present in endometrium, and the ectoenzyme NTPDase2 has been identified in cells with perivascular localizations, such as micropericytes and adventitial cells in several different types of tissue. Knowing the role of purinergic signaling in MSCs and, specifically, the role of NTPDase2 in the control of stem/progenitor cell proliferation in neurogenic niches, it is evident that more studies of NTPDase2 in other tissues are required.
Based on these observations, we aimed to study NTPDase2 expression in human endometrium in relation to endometrial stromal cells as well as adenomyosis.
Materials and methods
Samples
Twenty-nine normal human endometrial samples from 29 women (mean age 56, standard deviation 12.32, and range 39–86 years) were obtained from hysterectomy specimens without endometrial malignancy at the Service of Gynecology of Bellvitge Hospital between February 2017 and June 2018. Three cases of adenomyosis (mean age 48, standard deviation 6.24, and range 43–55 years) were also included in the study. Informed consent was obtained from all individual participants included in the study. Six proliferative endometria, 4 secretory endometria, and 19 atrophic endometria were used in this study. The endometrial dating was done at the Service of Pathology of Bellvitge Hospital.
Tissue samples were cut and fixed with 4% paraformaldehyde, then introduced in 30% (w/v) sucrose solution at 4 °C for 24 h for their cryoprotection and embedded in O.C.T freezing media (Tissue-Tek®; Sakura Finetk, Zoeterwoude, Netherlands). Fifteen-micrometer sections were obtained using a Cryostat Leica CM1950 (Leica, Wetzlar, Germany). Sections were put onto poly-l-lysine coated glass slides and stored at − 20 °C until use. Routine hematoxylin and eosin staining was performed.
Antibodies
Primary antibodies used in this study are listed in Table 1. Secondary antibodies used for immunohistochemistry were as follows: horseradish peroxidase (HRP)-conjugated goat anti-mouse (EnVision™ + System; DAKO, Carpinteria, CA, USA) and HRP-conjugated goat anti-rabbit (EnVision™ + System). Secondary antibodies used for immunofluorescence assays were as follows: Alexa Fluor 488-goat anti-mouse IgG (Thermo Fisher Scientific, Waltham, MA, USA), Alexa Fluor 488-goat anti-mouse IgG2b (Thermo Fisher Scientific), Alexa Fluor 555-goat anti-mouse IgG1 (Thermo Fisher Scientific), Alexa Fluor 555-goat anti-mouse IgG (Thermo Fisher Scientific), Alexa Fluor 488-goat anti-rabbit (Thermo Fisher Scientific), Alexa Fluor 555-goat anti-rabbit IgG (Thermo Fisher Scientific), and Alexa Fluor 647-goat anti-rabbit (Thermo Fisher Scientific). Secondary antibodies were used at 1:500 and dilutions were made in PBS.
Table 1.
List of primary antibodies used for immunolabeling experiments
| Antibody specificity | Name/clone | Source | Supplier | Dilution |
|---|---|---|---|---|
| NTPDase1 (CD39) | BU-61 | Mouse (IgG1) | Ancell (188-820) | 1:500 |
| NTPDase2 (CD39L1) | – | Rabbit | Enzo (ALX-215-045) | 1:100 |
| NTPDase2 (CD39L1) | H9s | Mouse (IgG2b) | http://ectonucleotidases-ab.com | 1 μg/mL |
| CD10 | SP67 | Rabbit | Ventana (790-4506) | 4.5 μg/mL |
| α-SMA | – | Rabbit | Abcam (ab5694) | 1:200 |
| SUSD2 | – | Rabbit | Abcam (ab121214) | 1:400 |
| CD146 | P1H12 | Mouse (IgG1) | Abcam (ab24577) | 1:100 |
| PDGFRβ (CD140b) | – | Rabbit | Assay Biotechnology (B0544) | 1:100 |
| CD31 | – | Rabbit | Abcam (ab28364) | 1:50 |
Immunolabeling experiments
Sections were washed twice with PBS to remove the O.C.T freezing media. The slices were pre-incubated for 1 h at room temperature (RT) with PBS containing 20% normal goat serum (NGS; Gibco, Paisley, UK), 0.2% Triton, and 0.2% gelatin (Merck, Darmstadt, Germany). For immunohistochemistry experiments, a previous blocking of endogenous peroxidase activity was performed with 10% methanol (v/v) and 2% H2O2 (v/v) in PBS for 30 min. Slices were incubated overnight at 4 °C with the primary antibodies (listed in Table 1) diluted in PBS. After three washes with PBS, tissue sections were incubated with the appropriate secondary antibodies at RT for 1 h in the case of fluorescence and for 30 min in the case of HRP-conjugated antibodies. Secondary antibodies alone were routinely included as controls for the experiments.
For immunohistochemistry, the peroxidase reaction was performed in a solution containing 0.6 mg/mL 3,3′-diaminobenzidine substrate (DAB; D-5637, Sigma-Aldrich, Saint Louis, MO, USA), and 0.5 μL/mL H2O2 in PBS for 10 min, and stopped with PBS. Nuclei were counterstained with hematoxylin and slides were then dehydrated and mounted with DPX mounting medium (VWP International Ltd., Radnor, PA, USA). Samples were observed under light Nikon Eclipse E200 and photographed under light Leica DMD 108 microscope. In fluorescence assays, for nuclei labeling, slides were mounted with aqueous mounting medium with DAPI (ProLong™ Gold antifade reagent with DAPI, Life Technologies, Paisley, UK). Samples were then observed and photographed under a Zeiss LSM 880 Confocal Laser Scanning Microscope. Fluorescence images were processed with the software ZEN 2.3 SP1 (Zeiss, Oberkochen, Germany).
Immunohistochemical staining was independently evaluated by two observers. Labeling distribution was recorded. Label intensity was scored as negative (−), weak (+), intermediate (++), or strongly positive (+++).
In situ ATPase activity experiments
A protocol based on the Wachstein/Meisel lead phosphate method was used [32, 34–36]. The sections were washed twice with 50 mM Tris-maleate buffer pH 7.4 and were pre-incubated for 30 min at RT with 50 mM Tris-maleate buffer pH 7.4 containing 2 mM MgCl2 and 0.25 M sucrose. The enzymatic reaction was carried out by incubating tissue sections for 1 h at 37 °C with 50 mM Tris-maleate buffer pH 7.4 supplemented with 0.25 M sucrose, 2 mM MgCl2, 5 mM MnCl2, 3% Dextran, 2 mM Pb(NO3)2, and 2 mM CaCl2. Experiments were performed in the presence of 2.5 mM levamisole, as an inhibitor of the alkaline phosphatase activity, and in the presence of 1 mM ATP as a substrate. Control assays were performed in the absence of nucleotide. For inhibition experiments, 1 mM POM 1 (Tocris Bioscience, Bristol, UK) was added to both pre-incubation and enzymatic reaction buffers. Reactions were revealed by incubation with 1% (NH4)2S (v/v) for exactly 1 min. Nuclei were counterstained with hematoxylin. Samples were mounted with aqueous mounting medium (FluoromountTM, Sigma-Aldrich), observed under light Nikon Eclipse E200 microscope, and photographed under light Leica DMD 108 microscope.
Combined immunofluorescence staining and in situ ATPase activity experiments
A protocol based on the combination of immunolabeling experiments and the Wachstein/Meisel lead phosphate method was performed, as previously described by Villamonte et al. [36]. Briefly, sample sections were washed twice with PBS and blocked in PBS containing 20% NGS (Gibco), 0.2% Triton, and 0.2% gelatin (Merck) at RT for 1 h. The samples were incubated overnight at 4 °C with the primary antibodies diluted with PBS. The sections were then washed three times with PBS and twice with 50 mM Tris-maleate buffer. In situ ATPase activity experiment was performed in the same sections as indicated above, using 1 mM of ATP as substrate. Subsequently, the tissues were washed three times in PBS before appropriate secondary antibody (Alexa Fluor) was added. After three final washes with PBS, samples were mounted on a glass slide with Prolong Gold antifade reagent with DAPI mounting medium (Thermo Fisher Scientific). The sections were observed and photographed under a light and fluorescence Nikon Eclipse E800 microscope. Immunofluorescence and activity images were merged using Adobe Photoshop CC (vs 20.0).
Statistical analyses
The predictive analytics software IBM SPSS Statistics v22 (IBM Corp., Armonk, NY, USA) was used for the creation of frequency tables with the distribution of NTPDase2 in each endometrial component and the label intensity in each case. Data are compiled in Table 2.
Table 2.
Summary of NTPDase2 expression in proliferative, secretory and atrophic endometria
| NTPDase2 | |
|---|---|
| Proliferative and secretory | |
| Surface epithelium | +++ a |
| Glandular epithelium | +++ a |
| Stromal cells | |
| Functional layer | − |
| Basal layer | +++ |
| Vessels | |
| Endothelial cells | − |
| Perivascular cells | +++ |
| Atrophic | |
| Surface epithelium | +++ a |
| Glandular epithelium | +++ a |
| Endometrial stromal cells | +++ |
| Vessels | |
| Endothelial cells | − |
| Perivascular cells | +++ |
aOnly cilia from ciliated cells
Results
NTPDase2 was detected in the entire length of the cilia of epithelial ciliated cells, in some perivascular cells of both functional and basal layers, and in the stroma only from the basal layer (Fig. 1). ATPase activity coincided with these locations. Combined immunofluorescence and in situ ATPase activity experiments allowed us to simultaneously identify the protein and its enzyme activity (Fig. 2a). Moreover, ATPase activity was inhibited by the E-NTPDase inhibitor POM 1, confirming that a member of the E-NTPDase family is responsible for the hydrolysis of ATP in these structures (Fig. 2b). Distribution and label intensity of NTPDase2 did not show differences at any point in the cycle or with atrophic endometrium (Suppl. Fig. 1).
Fig. 1.

Immunolocalization of NTPDase2 in a human secretory endometrium (Endo) and myometrium (Myo) by immunohistochemistry. Image A shows the immunolabeling of NTPDase 2 at the endometrial stroma but only of the basal layer. Image Aa shows the presence of NTPDase2 in the cilia of ciliated epithelial cells of the surface epithelium. Ab is a detail of the NTPDase2 immunolabeling in cells in a perivascular location. Ac shows a detail of the basal layer. NTPDase2 antibody used was H9s from http://ectonucleotidases-ab.com. Scale bars 2 mm (A), 50 μm (Aa), 25 μm (Ab), and 100 μm (Ac)
Fig. 2.

a Immunofluorescence of NTPDase2 (A, D) and in situ ATPase histochemistry (B, E) in cryosections of a human proliferative endometrium. NTPDase2 was detected by immunofluorescence in the cilia of ciliated epithelial cells (A; arrows) and in stromal cells of basal layer (A, D). Higher NTPDase2 expression and ATPase activity was found in periglandular stromal cells (C, F). Merged images (C, F) show that ATPase activity correlates with the immunolabeling of NTPDase2 protein. NTPDase2 antibody used was ALX-215-045 from Enzo. Scale bars 25 (C) and 150 μm (F). b In situ ATPase activity in functional layer of a proliferative endometrium. ATPase activity was found in endothelial cells and perivascular cells (A). Image B corresponds to the activity experiment of a serial section in the presence of POM 1 inhibitor and shows a huge inhibition of the ATPase activity. Image C shows an activity experiment performed in the absence of substrate. Scale bar 100 μm
In functional layer, NTPDase2 label was limited to perivascular locations. Only occasionally in the proliferative phase, periglandular labeling was seen in the functional layer, but always at the deepest part of the layer. NTPDase2 immunolabeling was strong in the stroma of the basal layer with stronger periglandular label also coinciding with ATPase activity. NTPDase2+ cells of stroma, including those periglandularly located, were also positive for CD10, a marker of endometrial stroma (Fig. 3). In postmenopausic endometrium, NTPDase2 was present in the stromal cells throughout the endometrium, but strongest periglandularly (Suppl. Fig. 1). To further study NTPDase2+ cells in relation to the already identified eMSCs, double and triple immunolabelings were performed. Perivascular NTPDase2+ cells were negative for α-smooth muscle actin (α-SMA), a marker of pericytes and vascular smooth muscle cells (Fig. 3). Thus, we ruled out the possibility of NTPDase2+ cells being pericytes. Moreover, triple immunolabeling for NTPDase2 with the pericyte markers CD146 and PDGFRβ, which when merged identify an endometrial mesenchymal stem cell population (eMSC), did not show colocalization of NTPDase2 and the other two proteins. In fact, CD146- and PDGFRβ-labeled cells directly wrapped endothelial cells while NTPDase2+ cells were located more at the periphery (Suppl. Fig. 2). This result confirms that NTPDase2+ cells are neither pericytes nor PDGFRβ+/CD146+ eMSCs. In contrast, we found complete colocalization with the eMSC marker SUSD2, detected perivascularly in both the functional and basal layers (Fig. 4a). Figure 4b shows this external position of the two proteins in relation to the vessel using antibodies against another member of the E-NTPDase family, NTPDase1, which was expressed by the endothelial cells (Suppl. Fig. 3). These results were obtained in cyclic and postmenopausic endometrium. NTPDase2+/SUSD2+ perivascular cells were also detected in the myometrium (Fig. 4a).
Fig. 3.

Confocal fluorescence images of the basal layer of a secretory endometrium with antibodies against NTPDase2 (A, E), CD10 (B), and α-SMA (F). Nuclei were labeled with DAPI (C, G). Merged images show that NTPDase2 colocalizes with CD10 in the stromal cells of basal layer (D) but does not co-occur with the α-SMA (H) vascular smooth muscle cells marker. NTPDase2 antibody used was H9s from http://ectonucleotidases-ab.com. Scale bar 50 μm (D, H)
Fig. 4.

a Confocal fluorescence images of a secretory (A–D) and a proliferative (E–H) endometrium and myometrium (I–L) labeled with the antibodies against NTPDase2 (A, E, I) and the eMSC marker SUSD2 (B, F, J). Nuclei were labeled with DAPI (C, G, K). The images A–D show part of the basal layer of a secretory endometrium. NTPDase2 label is present in cilia of ciliated cells of glands, in stromal cells, especially periglandularly, and in perivascular cells. Images E–H show a detail of an endometrial vessel localized in the functional layer of a proliferative endometrium and images I–L show a detail of the vessels in myometrium. NTPDase2 is only present in perivascular cells (E, I). The eMSC marker SUSD2 is present in perivascular cells (B, F, J). Merge images (D, H, L) show the colocalization of NTPDase2 and SUSD2 in the perivascular cells. Scale bars 50 μm (D, H) and 35 μm (L). b Immunolocalization of NTPDase2 (A), SUSD2 (B), and NTPDase1 (C) in the vessel of an atrophic endometrium. Nuclei were labeled with DAPI. NTPDase2 (A) and SUSD2 (B) were present in perivascular cells while NTPDase1 was detected in endothelial cells (C). Merged image shows the colocalization of NTPDase2 and SUSD2 in perivascular cells (D). Scale bar 50 μm (D) NTPDase2 antibody used in all the experiments shown in this figure was H9s from http://ectonucleotidases-ab.com
We analyzed adenomyotic lesions, where endometrial tissue is present within the myometrium, and found that the stroma was made up of NTPDase2+ cells (Fig. 5). Perivascular NTPDase2+ population was also found in these lesions (data not shown) although the small area of the lesions with minor presence of blood vessels hampered this analysis.
Fig. 5.

Immunolocalization of NTPDase2 in a human uterus with adenomyosis. Image A shows the immunolabeling of NTPDase2 at the stroma of the basal layer of the endometrium and at the stroma of adenomyosis lesion. B shows a detail of adenomyosis lesion with the presence of NTPDase2. NTPDase2 antibody used was H9s from http://ectonucleotidases-ab.com. Scale bars 1 mm (A) and 100 μm (B)
Table 2 is a compilation of the results of distribution and expression levels of NTPDase2 in human endometrium and Fig. 6 puts the NTPDase2 expression in endometrium into context.
Fig. 6.

Schematic representation of human endometrium showing the main distribution findings for NTPDase2 and other proteins. NTPDase3 is only present in spiral arteries (asterisk) [32]
Discussion
Purinergic ligands, such as ATP and adenosine, are signaling molecules that play important roles in reproduction [37], including the regulation of the endometrial functions; they also influence MSC function, contributing to the maintenance of a harmonious stem cell niche [8, 9]. The eMSC are promising therapeutic targets due to their possible involvement in the pathogenesis of gynecological diseases, such as endometriosis, adenomyosis, and cancer, and in other tissues for their immunomodulatory, differentiation, and tissue regeneration properties [4, 8, 38, 39]. More studies of the purinome in the MSCs are required in order to increase knowledge of MSC physiology and to identify new therapeutic targets. In the present work, we have characterized, the ectoenzyme NTPDase2 in the human cyclic and postmenopausic endometrium and analyzed its expression in adenomyosis samples.
We found high expression and activity of NTPDase2 in both cyclic and postmenopausic endometria without any change along the cycle or in the atrophic endometrium. Thus, NTPDase2 expression does not seem to be regulated by the female sex hormones. This is a differential feature in comparison to other ectonucleotidases whose expression fluctuates at points along the cycle [32]. The sustained expression of NTPDase2 throughout the cycle points to the constant need for regulation of the ATP levels in stroma. NTPDase2, in combination with NTPDase1, also present in the stroma, can generate AMP which in turn is converted into adenosine by the action of stromal ecto-5′nucleotidase (CD73). Considering their abundance, NTPDase2 and CD73 [32] would be the two main ectonucleotidases acting sequentially to generate adenosine in endometrial stroma. We demonstrate NTPDase2 expression, with two different antibodies, in two main locations of endometrium: in cilia from ciliated epithelial cells, especially of the luminal epithelium, which is in accordance with our previous finding of NTPDase2 expression in cilia of Fallopian tubes [36], and, in stromal cells. A striking finding is that only the stroma of the basalis is labeled; in functionalis, labeling was only detected perivascularly. CD10, a marker of stromal cells, labeled all the stroma and clearly colocalized with NTPDase2+ cells of the basal stroma; the NTPDase2 label was more intense periglandularly. To our knowledge, this is the first cell marker of basal stroma, a finding that is of relevance in the tracing of basal stroma in pathologies, such as adenomyosis, that originate in the basalis. Adenomyosis is a gynecological disease defined by the growth of endometrial cells and their migration into the myometrium. We analyzed adenomyosis samples and found NTPDase2 in the stromal cells of the lesion. Despite the diagnosis of adenomyosis made by histopathology, NTPDase2 represents a suitable marker to separate the limits of ectopic endometrial tissue from the rest of myometrium.
Since the basalis physiologically regenerates the endometrium in every cycle, it is assumed that both epithelial and mesenchymal progenitor cells are located in this basal layer of human endometrium [2, 3, 38–42]. The epithelial progenitor cells are postulated to be contained by the basalis glands [1–3]. N-cadherin+ epithelial progenitor cells were recently identified in the deep basalis at the bases of endometrial glands [2]. The combination of N-cadherin and the basal glandular epithelial marker SSEA-1 seems to be a useful tool to determine the differentiation hierarchy of epithelial cells in the basal zone of the gland [2]. Although N-cadherin+ cells might have some progenitor activity, functional studies confirming this point have not been yet performed. The presence of NTPDase2 surrounding basal glands, and not in functional glands, could be related to the possible action of NTPDase2 in the maintenance of epithelial progenitor cells. In fact, in adult murine neurogenic regions, NTPDase2 is highly expressed in progenitor cells; it is the main ectoenzyme regulating the nucleotide levels of the progenitor cell milieu [27, 28]. Gampe et al. showed an increase in the number of bromodeoxyuridine (BrdU)-positive cells in the neurogenic niches, subventricular zone, and subgranular layer of dentate gyrus, in Ntpd2 KO mice, but Ntpd2 deletion does not affect the number of neurons [29]. In addition, Ntpd2 deletion also leads to increased progenitor cell proliferation in vitro, while the addition of the ATP/ADP-hydrolyzing enzyme apyrase reduced the number of neurosphere cells derived from mice deficient in NTPDase2 expression [29]. These observations suggest that the deletion of NTPDase2 results in an increase in neural progenitor cells. So NTPDase2 may play a role in the proliferation and expansion of neural progenitor cells [29]. The role of NTPDase2 identified in neurogenic regions might well also be found in other stem cell niches, such as the endometrial basal layer. Other proteins such as Musashi-1, an RNA-binding protein identified in neural stem cells and also an epithelial progenitor cell marker, were also detected in human endometrium, mainly in the basalis in the proliferative phase [43], suggesting their possible progenitor cell function. Some stromal Musashi-1 positive cells were found in the periglandular region, where some stromal label-retaining cells were found in mouse endometrium [39, 43, 44]. It is likely that the periglandular expression of NTPDase2 matches with the periglandular clusters of stromal Musashi-1+ cells. The ATP-adenosine balance is a key regulator of progenitor/stem cell pathophysiology, involved in the differentiation and self-renewal of stem cells [21]. Further work is needed to determine the action of NTPDase2 in the progenitor/stem cell function in endometrium.
A notable result of this work is the identification of NTPDase2 as a mesenchymal progenitor marker since it is expressed in SUSD2+ eMSCs, which constitute the most abundant population of MSCs in human endometrium, representing 4.2% of the stromal cell population [5], almost three times more abundant than the PDGFβ+CD146+ cells [45]. The presence and activity of NTPDase2 in the perivascular SUSD2+ eMSCs in cyclic and atrophic endometria is in line with previous findings suggesting that human endometrium is a source of MSCs independently of the woman’s age [47]. Our results point to a role for NTPDase2 in the biological properties of the endometrial stem cells throughout the woman’s life.
The presence of NTPDase2 in eMSCs entails the hydrolysis of ATP and a sustained ADP accumulation in the stem cell niche. Different studies performed in vivo and in vitro in recent years have demonstrated that MSCs release ATP and nucleosides constitutively or in response to mechanical or chemical stimulation [21]. Moreover, MSCs are sensitive to these purinergic signaling effectors due to their broad expression of purinergic receptors on their plasma membranes, including ATP and ADP receptors [10, 13, 16, 17, 48, 49]. In fact, both ATP and adenosine are involved in the immunomodulatory capacity and in the lineage commitment of the MSCs. Nucleotides have been associated with the proliferation [8, 10, 11, 13, 21, 50] and differentiation of MSC’s [16, 17, 48], while adenosine is involved in the survival of MSCs [8], proliferation (with an opposite ATP effect) [21], and lineage commitment [15, 19, 48]. Given the implications of nucleotides and nucleosides in the maintenance and behavior of MSCs, ectonucleotidases play a necessary role in the maintenance of the homeostasis of the perivascular stem cell niche. Indeed, the remarkable machinery of ATP hydrolysis is present in this vascular environment, where, in addition to the expression of NTPDase2 in eMSCs, other members of the NTPDase family are present: NTPDase1 is expressed by endothelial cells and NTPDase3 is found in α-SMA+ pericytes of spiral arteries [32].
We also find NTPDase2+SUSD2+ perivascular cells in human myometrium, pointing to the presence of an equivalent number of MSCs as found in the endometrium, which is consistent with the shared embryonic origin of the two tissues. In fact, Patterson et al. identified PGFRβ+CD146+ perivascular, possibly progenitor, cells in murine myometrium [50] although functional studies need to be done. They found that this population of cells was label-retaining and not sensitive to female hormones, two features that indicate their possible role as progenitor cells in the myometrium. Functional studies with myometrial are needed with this NTPDase2+SUSD2+ perivascular cell population.
In conclusion, NTPDase2 is a marker of the endometrial stromal basal layer of utility in tracing adenomyosis. Moreover, the NTPDase2 antibodies label SUSD2+ perivascular cells, pointing to their usefulness in improving isolation of uterine mesenchymal progenitor cells to be used in regenerative medicine. The findings from our study provide new information about the purinergic signaling in the healthy endometrium and its physiological regeneration. Further investigation into the specific involvement of NTPDase2 in the physiology of eMSCs, human fertility, and endometrial pathology is essential for the development of new therapeutic strategies in regenerative medicine.
Electronic supplementary material
{kind=link}
Immunolocalization of NTPDase2 in human proliferative (A), secretory (B), and atrophic (C) endometrium. Stroma is labelled in the three cases although labelling is restricted to basal layer in cyclic endometria (A, B). NTPDase2 antibodies used were ALX-215-045 from Enzo (A, C) and H9s from http://ectonucleotidases-ab.com (B). Scale bars 400 μm (A), 300 μm (B), and 100 μm (C) (PNG 2943 kb)
High Resolution Image (TIF 8501 kb) (PNG 1128 kb)
{kind=link}
Confocal fluorescence images of some vessels of human atrophic endometrium labeled with NTPDase2 (A), CD146 (B), and PDGFRβ (C). Merged image shows a more external position of NTPDase2+ cells than CD146- and PDGFRβ-positive cells in the perivascular region (D). NTPDase2 antibody used was H9s from http://ectonucleotidases-ab.com. Scale bar 25 μm (D) (PNG 1128 kb) (JPG 216 kb)
High Resolution Image (TIF 6.40 mb) (PNG 1.10 mb)
{kind=link}
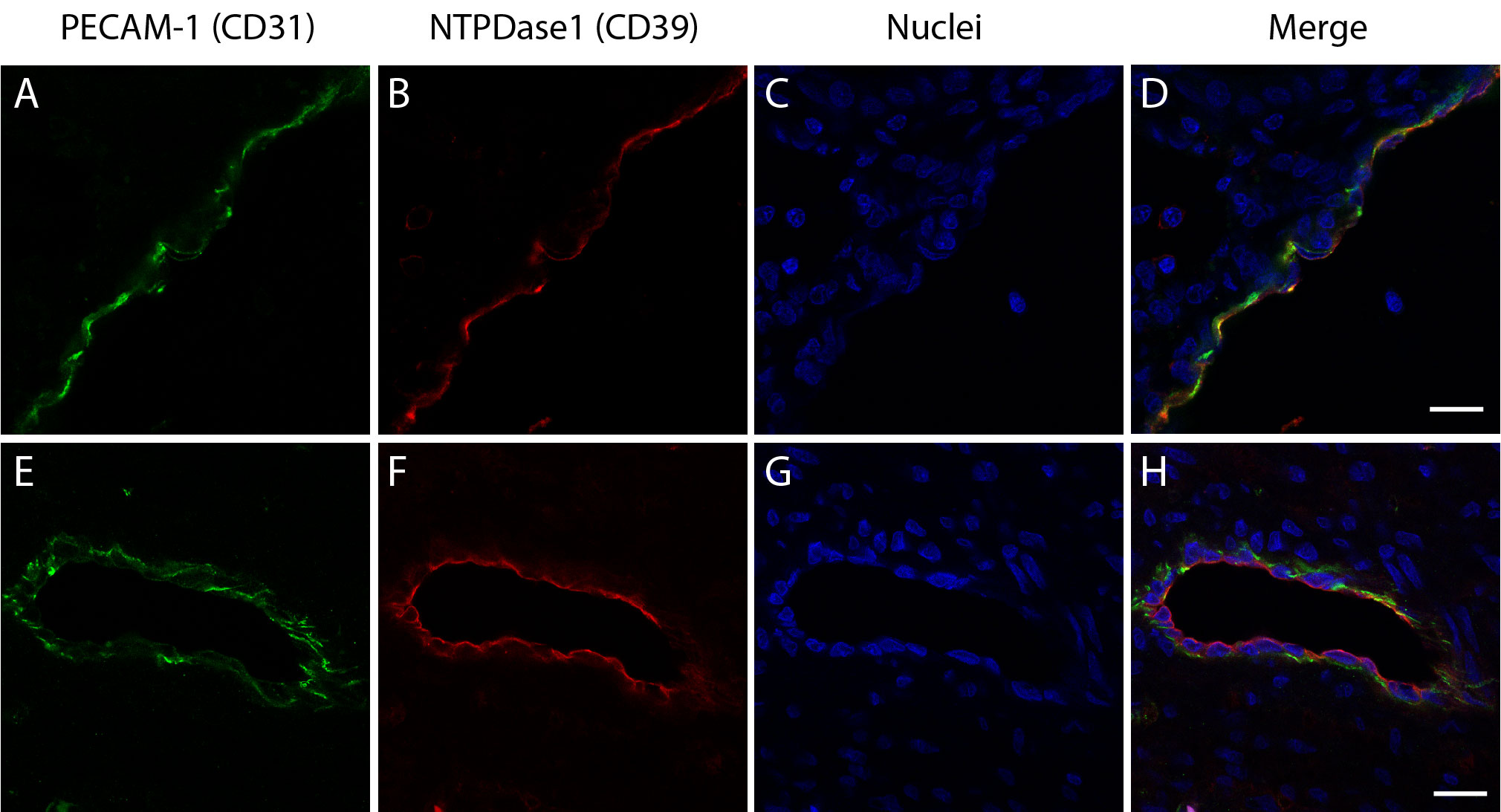
Confocal fluorescence images of endometrial blood vessels labeled with PECAM-1 (CD31) and NTPDase1 (CD39). Endothelial cells labelled with CD31 (A, E) are also positive for NTPDase1 (B, F) as shown in merge images (D, H). Scale bars 20 μm (PNG 1.01 mb)
Acknowledgments
This study was supported by a grant from the Instituto de Salud Carlos III (FIS PI15/00036), co-funded by FEDER funds/European Regional Development Fund (ERDF)-“a Way to Build Europe”-//FONDOS FEDER “una manera de hacer Europa,” and a grant from the Fundación Merck Salud (Ayuda Merck de Investigación 2016-Fertilidad). ARM was awarded a fellowship from the Asociación Española Contra el Cáncer (AECC). JS received support from the Canadian Institutes of Health Research (CIHR) and was the recipient of a “Chercheur National” research award from the Fonds de recherche du Québec – Santé (FRQS). We thank CERCA Programme (Generalitat de Catalunya) for institutional support. We are grateful to Inmaculada Gómez de Aranda for technical support and to Benjamín Torrejón of Serveis Científics I Tecnològics (Campus Bellvitge, Universitat de Barcelona). The authors thank Tom Yohannan for language editing.
Funding
This study was funded by Instituto de Salud Carlos III (grant number FIS PI15/00036); FEDER funds/European Regional Development Fund (ERDF)-“a Way to Build Europe”; Fundación Merck Salud (Ayuda Merck de Investigación 2016-Fertilidad).
Compliance with ethical standards
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments, or comparable ethical standards.
Informed consent
Informed consent was obtained from all individual participants in the study.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Gargett CE, Schwab KE, Deane JA. Endometrial stem/progenitor cells: the first 10 years. Hum Reprod Update. 2016;22(2):137–163. doi: 10.1093/humupd/dmv051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nguyen HTP, Xiao L, Deane JA, Tan K, Cousins FL, Masuda H, Sprung CN, Rosamilia A, Gargett CE. N-cadherin identifies human endometrial epithelial progenitor cells by in vitro stem cell assays. Hum Reprod. 2017;32(11):2254–2268. doi: 10.1093/humrep/dex289. [DOI] [PubMed] [Google Scholar]
- 3.Valentijn AJ, Palial K, Al-lamee H, Tempest N, Drury J, Von Zglinicki T, Saratzki G, Murray P, Gargett CE, Hapangama DK. SSEA-1 isolates human endometrial basal glandular epithelial cells: phenotypic and functional characterization and implications in the pathogenesis of endometriosis. Hum Reprod. 2013;28(10):2695–2708. doi: 10.1093/humrep/det285. [DOI] [PubMed] [Google Scholar]
- 4.Schwab KE, Gargett CE. Co-expression of two perivascular cell markers isolates mesenchymal stem-like cells from human endometrium. Hum Reprod. 2007;22(11):2903–2911. doi: 10.1093/humrep/dem265. [DOI] [PubMed] [Google Scholar]
- 5.Masuda H, Anwar SS, Bühring HJ, Rao JR, Gargett CE. A novel marker of human endometrial mesenchymal stem-like cells. Cell Transplant. 2012;21(10):2201–2214. doi: 10.3727/096368911X637362. [DOI] [PubMed] [Google Scholar]
- 6.Crisan M, Yap S, Casteilla L, Chen CW, Corselli M, Park TS, Andriolo C, Sun B, Zheng B, Zheng L, Norotte C, Teng PN, Traas J, Schugar R, Deasy BM, Badylak S, Bühring HJ, Giacobino JP, Lazzari L, Huard J, Péault B. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell. 2008;3(3):301–313. doi: 10.1016/j.stem.2008.07.003. [DOI] [PubMed] [Google Scholar]
- 7.Vezzani B, Pierantozzi E, Sorrentino V (2018) Mesenchymal stem cells: from the perivascular environment to clinical applications. Histol Histopathol. 10.14670/HH-11-998 [DOI] [PubMed]
- 8.Scarfi S. Purinergic receptors and nucleotide processing ectoenzymes: their roles in regulating mesenchymal stem cell functions. World J Stem Cells. 2014;6(2):153–162. doi: 10.4252/wjsc.v6.i2.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cavaliere F, Donno C, D’Ambrosi N (2015) Purinergic signaling: a common pathway for neural and mesenchymal stem cell maintenance and differentiation. Front Cell Neurosci 9(211). 10.3389/fncel.2015.00211 [DOI] [PMC free article] [PubMed]
- 10.Coppi E, Pugliese AM, Urbani S, Melani A, Cerbai E, Mazzanti B, Bosi A, Saccardi R, Pedata F. ATP modulates cell proliferation and elicits two different electrophysiological responses in human mesenchymal stem cells. Stem Cells. 2007;25(7):1840–1849. doi: 10.1634/stemcells.2006-0669. [DOI] [PubMed] [Google Scholar]
- 11.Ichikawa J, Gemba H. Cell density-dependent changes in intracellular Ca2+ mobilization via the P2Y2 receptor in rat bone marrow stromal cells. J Cell Physiol. 2009;219(2):372–381. doi: 10.1002/jcp.21680. [DOI] [PubMed] [Google Scholar]
- 12.Katebi M, Soleimani M, Cronstein BN. Adenosine A2A receptors play an active role in mouse bone marrow-derived mesenchymal stem cell development. J Leukoc Biol. 2009;85(3):438–444. doi: 10.1189/jlb.0908520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ferrari D, Gulinelli S, Salvestrini V, Lucchetti G, Zini R, Manfredini R, Caione L, Piacibello W, Ciciarello M, Rossi L, Idzki M, Ferrari S, Di Virgilio F, Lemoli RM. Purinergic stimulation of human mesenchymal stem cells potentiates their chemotactic response to CXCL12 and increases the homing capacity and production of proinflammatory cytokines. Exp Hematol. 2011;39(3):360–374. doi: 10.1016/j.exphem.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 14.Fruscione F, Scarfi S, Ferraris C, Bruzzone S, Benvenuto F, Guida L, Uccelli A, Salis A, Usai C, Jacchetti E, Ilengo C, Scaglione S, Quarto R, Zocchi E, De Flora A. Regulation of human mesenchymal stem cell functions by an autocrine loop involving NAD+ release and P2Y11-mediated signaling. Stem Cells Dev. 2011;20(7):1183–1198. doi: 10.1089/scd.2010.0295. [DOI] [PubMed] [Google Scholar]
- 15.Carroll SH, Wigner NA, Kulkarni N, Johnston-Cox H, Gerstenfeld LC, Ravid K. A2B adenosine receptor promotes mesenchymal stem cell differentiation to osteoblasts and bone formation in vivo. J Biol Chem. 2012;287(19):15718–15727. doi: 10.1074/jbc.M112.344994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zippel N, Limbach CA, Ratajski N, Urban C, Luparello C, Pansky A, Kassack MU, Tobiasch E. Purinergic receptors influence the differentiation of human mesenchymal stem cells. Stem Cells Dev. 2012;21(6):884–900. doi: 10.1089/scd.2010.0576. [DOI] [PubMed] [Google Scholar]
- 17.Sun D, Junger WG, Yuan C, Zhang W, Bao Y, Qin D, Wang C, Tan L, Qi B, Zhu D, Zhang X, Yu T. Shockwaves induce osteogenic differentiation of human mesenchymal stem cells through ATP release and activation of P2X7 receptors. Stem Cells. 2013;31(6):1170–1180. doi: 10.1002/stem.1356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chen M, Su W, Lin X, Guo Z, Wang J, Zhang Q, Bramd D, Ryffel B, Huang J, Liu Z, He X, Le AD, Zheng SG. Adoptive transfer of human gingiva-derived mesenchymal stem cells ameliorates collagen-induced arthritis via suppression of Th1 and Th17 cells and enhancement of regulatory T cell differentiation. Arthritis Rheum. 2013;65(5):1181–1193. doi: 10.1002/art.37894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ode A, Schoon J, Kurtz A, Gaetien M, Ode JE, Geissier S, Duda GN. CD73/5′-ecto-nucleotidase acts as a regulatory factor in osteo-/chondrogenic differentiation of mechanically stimulated mesenchymal stromal cells. Eur Cell Mater. 2013;25:37–47. doi: 10.22203/eCM.v025a03. [DOI] [PubMed] [Google Scholar]
- 20.Naasani LIS, Rodrigues C, de Campos RP, Beckenkamp LR, Iser IC, Bertoni APS, Wink MR. Extracellular nucleotide hydrolysis in dermal and limbal mesenchymal stem cells: a source of adenosine production. J Cell Biochem. 2017;118:2430–2442. doi: 10.1002/jcb.25909. [DOI] [PubMed] [Google Scholar]
- 21.Roszek K, Wujak M (2018) How to influence the mesenchymal stem cells fate? Emerging role of ectoenzymes metabolizing nucleotides. J Cell Physiol. 10.1002/jcp.26904 [DOI] [PubMed]
- 22.Zimmerman H, Zebisch M, Sträter N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal. 2012;8(3):437–502. doi: 10.1007/s11302-012-9309-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yegutkin GG. Enzymes involved in metabolism of extracellular nucleotides and nucleosides: functional implications and measurement of activities. Crit Rev Biochem Mol Biol. 2014;49(6):473–497. doi: 10.3109/10409238.2014.953627. [DOI] [PubMed] [Google Scholar]
- 24.Sévigny J, Sundberg C, Braun N, Guckelberger O, Csizmadia E, Qawi I, Imai M, Simmermann H, Robson SC. Differential catalytic properties and vascular topography of murine nucleoside triphosphate diphosphohydrolase 1 (NTPDase1) and NTPDase2 have implication for thromboregulation. Blood. 2002;99(8):2801–2809. doi: 10.1182/blood.V99.8.2801. [DOI] [PubMed] [Google Scholar]
- 25.Kishore BK, Isaac J, Fausther M, Tripp SR, Shi H, Gill PS, Braun N, Zimmermann H, Sévigny J, Robson SC. Expression of NTPDase1 and NTPDase2 in murine kidney: relevance to regulation of P2 receptor signaling. Am J Physiol Ren Physiol. 2005;288(5):F1032–F1043. doi: 10.1152/ajprenal.00108.2004. [DOI] [PubMed] [Google Scholar]
- 26.Feldbrügge L, Jiang ZG, Csizmadia E, Mitsuhashi S, Tran S, Yee EU, Rothwelier S, Vaid KA, Sévigny J, Schmelzle M, Popov YV, Robson SC. Distinct roles of ecto-nucleoside triphosphate diphosphohydrolase-2 (NTPDase2) in liver regeneration and fibrosis. Purinergic Signal. 2017;14(1):37–46. doi: 10.1007/s11302-017-9590-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shukla V, Zimmermann H, Wang L, Kettenmann H, Raab S, Hammer K, Sévigny J, Robson SC, Braun N. Functional expression of the ecto-ATPase NTPDase2 and of nucleotide receptors by neuronal progenitor cells in the adult murine hippocampus. J Neurosci Res. 2005;80(5):600–610. doi: 10.1002/jnr.20508. [DOI] [PubMed] [Google Scholar]
- 28.Mishra SK, Braun N, Shukla V, Füllgrabe M, Schomerus C, Korf HW, Gachet C, Ikehara Y, Sévigny J, Robson SC, Zimmermann H. Extracellular nucleotide signaling in adult neural stem cells: synergism with growth factor-mediated cellular proliferation. Development. 2006;133(4):675–684. doi: 10.1242/dev.02233. [DOI] [PubMed] [Google Scholar]
- 29.Gampe K, Stefani J, Hammer K, Brendel P, Pötzsch A, Enikolopov G, Enjyoji K, Acker-Palmer A, Robson SC, Zimmermann H. NTPDase2 and purinergic signaling control progenitor cell proliferation in neurogenic niches of the adult mouse brain. Stem Cells. 2015;33(1):253–264. doi: 10.1002/stem.1846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Martín-Satué M, Lavoie ÉG, Pelletier J, Fauster M, Csizmadia E, Guckelberger O, Robson SC, Sévigny J. Localization of plasma membrane bound NTPDases in the murine reproductive tract. Histochem Cell Biol. 2009;131:615–628. doi: 10.1007/s00418-008-0551-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Arase T, Uchida H, Kajitani T, Ono M, Tamaki K, Oda H, Nishikawa S, Kagami M, Nagashima T, Masuda H, Asada H, Yoshimura Y, Maruyama T. The UDP-glucose receptor P2R14 triggers innate mucosal immunity in the female reproductive tract by inducing IL-8. J Immunol. 2009;182(11):7074–7084. doi: 10.4049/jimmunol.0900001. [DOI] [PubMed] [Google Scholar]
- 32.Aliagas E, Vidal E, Torrejón-Escribano B, Taco MR, Ponce J, de Aranda IG, Sévigny J, Condom E, Martín-Satué M. Ecto-nucleotidases distribution in human cyclic and postmenopausic endometrium. Purinergic Signal. 2013;9(2):227–237. doi: 10.1007/s11302-012-9345-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Von Kügelgen I, Hoffmann K. Pharmacology and structure of P2Y receptors. Neuropharmacology. 2016;106:50–61. doi: 10.1016/j.neuropharm.2015.10.030. [DOI] [PubMed] [Google Scholar]
- 34.Wachstein M, Meisel E, Niedzwiedz A. Histochemical demonstration of mitochondrial adenosine triphosphatase with the lead-adenosine triphosphate technique. J Histochem Cytochem. 1960;8:387–388. doi: 10.1177/8.5.387. [DOI] [PubMed] [Google Scholar]
- 35.Aliagas E, Vidal A, Teixidó L, Ponce J, Condom E, Martín-Satué M. High expression of ecto-nucleotidases CD39 and CD73 in human endometrial tumors. Mediat Inflamm. 2014;2014(509027):1–8. doi: 10.1155/2014/509027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Villamonte ML, Torrejón-Escribano B, Rodríguez-Martínez A, Trapero C, Vidal A, de Aranda IG, Sévigny J, Matías-Guiu M-SM. Characterization of ecto-nucleotidases in human oviducts with an improved approach simultaneously identifying protein expression and in situ enzyme activity. Histochem Cell Biol. 2018;149(3):269–276. doi: 10.1007/s00418-017-1627-8. [DOI] [PubMed] [Google Scholar]
- 37.Burnstock G. Purinergic signalling in the reproductive system in health and disease. Purinergic Signal. 2013;10(1):157–187. doi: 10.1007/s11302-013-9399-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cousins FL, O DF, Gargett CE. Endometrial stem/progenitor cells and their role in the pathogenesis of endometriosis. Best Pract Res Clin Obstet Gynaecol. 2018;50:27–38. doi: 10.1016/j.bpobgyn.2018.01.011. [DOI] [PubMed] [Google Scholar]
- 39.Tempest N, Maclean A, Hapangama DK. Endometrial stem cell markers: current concepts and unresolved questions. Int J Mol Sci. 2018;19(10):E3240. doi: 10.3390/ijms19103240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gargett CE. Uterine stem cells: what is the evidence? Hum Reprod Update. 2007;13(1):87–101. doi: 10.1093/humupd/dml045. [DOI] [PubMed] [Google Scholar]
- 41.Cervelló I, Mirantes C, Santamaria X, Dolcet X, Matias-Guiu X, Simón C. Stem cells in human endometrium and endometrial carcinoma. Int J Gynecol Pathol. 2011;30(4):317–327. doi: 10.1097/PGP.0b013e3182102754. [DOI] [PubMed] [Google Scholar]
- 42.Gargett CE, Masuda H. Adult stem cells in the endometrium. Mol Hum Reprod. 2010;16(11):818–834. doi: 10.1093/molehr/gaq061. [DOI] [PubMed] [Google Scholar]
- 43.Gotte M, Wolf M, Staebler A, Buchweitz O, Kelsch R, Schuring AN, Kiesel L. Increased expression of the adult stem cell marker mushashi-1 in endometriosis and endometrial carcinoma. J Pathol. 2008;215(3):317–329. doi: 10.1002/path.2364. [DOI] [PubMed] [Google Scholar]
- 44.Chan RW, Gargett CE. Identification of label-retaining cells in mouse endometrium. Stem Cells. 2006;24(6):1529–1538. doi: 10.1634/stemcells.2005-0411. [DOI] [PubMed] [Google Scholar]
- 45.Darzi S, Werkmeister JA, Deane JA, Ce G. Identification and characterization of human endometrial mesenchymal stem/stromal cells and their potential for cellular therapy. Stem Cells Transl Med. 2016;5(9):1127–1132. doi: 10.5966/sctm.2015-0190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ulrich D, Tan KS, Deane J, Schwab K, Cheong A, Rosamilia A, Gargett CE. Mesenchymal stem/stromal cells in post-menopausal endometrium. Hum Reprod. 2014;29(9):1895–1905. doi: 10.1093/humrep/deu159. [DOI] [PubMed] [Google Scholar]
- 47.Ciciarello M, Zini R, Rossi L, Salvestrini V, Ferrari D, Manfredini R, Lemoli RM. Extracellular purines promote the differentiation of human bone marrow-derived mesenchymal stem cells to the osteogenic and adipogenic lineages. Stem Cells Dev. 2013;22(7):1097–1111. doi: 10.1089/scd.2012.0432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.D’Alimonte I, Nargi E, Lannutti A, Marchisio M, Pierdomenico L, Costanzo G, Di Iorio P, Ballerini P, Giuliani F, Ciccarelli R. Adenosine A1 receptor stimulation enhances osteogenic differentiation of human dental pup-derived mesenchymal stem cells via WNT signalling. Stem Cell Res. 2013;11(1):611–624. doi: 10.1016/j.scr.2013.04.002. [DOI] [PubMed] [Google Scholar]
- 49.Czarnecka J, Porowińska D, Bajek A, Holysz M, Roszek K. Neurogenic differentiation of mesenchymal stem cells induces alterations in extracellular nucleotides metabolism. J Cell Biochem. 2017;118(3):478–486. doi: 10.1002/jcb.25664. [DOI] [PubMed] [Google Scholar]
- 50.Patterson AL, George J, Chatterjee A, Carpenter T, Wolfrum E, Pru JK, Teixeira JM. Label-retaining, putative mesenchymal stem cells contribute to repair of the myometrium during uterine involution. Stem Cells Dev. 2018;27:1715–1728. doi: 10.1089/scd.2018.0146. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Immunolocalization of NTPDase2 in human proliferative (A), secretory (B), and atrophic (C) endometrium. Stroma is labelled in the three cases although labelling is restricted to basal layer in cyclic endometria (A, B). NTPDase2 antibodies used were ALX-215-045 from Enzo (A, C) and H9s from http://ectonucleotidases-ab.com (B). Scale bars 400 μm (A), 300 μm (B), and 100 μm (C) (PNG 2943 kb)
High Resolution Image (TIF 8501 kb) (PNG 1128 kb)
Confocal fluorescence images of some vessels of human atrophic endometrium labeled with NTPDase2 (A), CD146 (B), and PDGFRβ (C). Merged image shows a more external position of NTPDase2+ cells than CD146- and PDGFRβ-positive cells in the perivascular region (D). NTPDase2 antibody used was H9s from http://ectonucleotidases-ab.com. Scale bar 25 μm (D) (PNG 1128 kb) (JPG 216 kb)
High Resolution Image (TIF 6.40 mb) (PNG 1.10 mb)
Confocal fluorescence images of endometrial blood vessels labeled with PECAM-1 (CD31) and NTPDase1 (CD39). Endothelial cells labelled with CD31 (A, E) are also positive for NTPDase1 (B, F) as shown in merge images (D, H). Scale bars 20 μm (PNG 1.01 mb)