

Abstract
Streptomyces rochei 7434AN4 produces two structurally unrelated polyketide antibiotics, lankacidin and lankamycin, and carries three linear plasmids, pSLA2-L (211 kb), -M (113 kb), and -S (18 kb), whose nucleotide sequences were previously reported. The complete nucleotide sequence of the S. rochei chromosome has now been determined using the long-read PacBio RS-II sequencing together with short-read Illumina Genome Analyzer IIx sequencing and Roche 454 pyrosequencing techniques. The assembled sequence revealed an 8,364,802-bp linear chromosome with a high G + C content of 71.7% and 7,568 protein-coding ORFs. Thus, the gross genome size of S. rochei 7434AN4 was confirmed to be 8,706,406 bp including the three linear plasmids. Consistent with our previous study, a tap-tpg gene pair, which is essential for the maintenance of a linear topology of Streptomyces genomes, was not found on the chromosome. Remarkably, the S. rochei chromosome contains seven ribosomal RNA (rrn) operons (16S-23S-5S), although Streptomyces species generally contain six rrn operons. Based on 2ndFind and antiSMASH platforms, the S. rochei chromosome harbors at least 35 secondary metabolite biosynthetic gene clusters, including those for the 28-membered polyene macrolide pentamycin and the azoxyalkene compound KA57-A.
Subject terms: Genome, Biosynthesis
Introduction
The filamentous Gram-positive soil bacterial genus Streptomyces is well characterized by its prolific potential to produce a vast array of secondary metabolites, including agriculturally and clinically useful antibiotics. Unlike other bacteria, their chromosomes are linear with a size of around 8–9 Mb. Streptomyces linear replicons harbor terminal inverted repeat (TIR) sequences at both ends and the 5′-ends are covalently bound to terminal protein (TP)1. Streptomyces linear chromosomes frequently undergo spontaneous deletions, leading to DNA rearrangements including amplification, arm replacement, and circularization2–4.
To date, over 1,141 Streptomyces strains have been sequenced and deposited in the GenBank database (ftp://ftp.ncbi.nlm.nih.gov/genomes/genbank/bacteria/) (as of 26th of June 2019), including the actinorhodin producer Streptomyces coelicolor A3(2)5, the avermectin producer Streptomyces avermitilis6, and the streptomycin producer Streptomyces griseus7 (Table 1). Streptomyces species have a great potential to produce over 20 secondary metabolites, including polyketides, non-ribosomal peptides, terpenoids, aminoglycosides, siderophores, and others. However, most of these biosynthetic gene clusters are poorly expressed or are not at all under normal culture conditions. Thus, Streptomyces genomes are a valuable source for natural product discovery.
Table 1.
General features of the chromosomes of Streptomyces rochei 7434AN4 and five other Streptomyces species.
| Speciesa | Length (bp) | G + C content (%) | CDS (no.) | Average CDS length (bp) | TIR (bp) | rRNAs (no.) | tRNA (no.) |
|---|---|---|---|---|---|---|---|
| S. rochei 7434AN4 | 8,364,802 | 71.7 | 7,568 | 973 | 53,892 | 7 | 67 |
| S. coelicolor A3(2) | 8,667,507 | 72.1 | 7,825 | 991 | 21,653 | 6 | 63 |
| S. avermitilis MA-4680 | 9,025,608 | 70.7 | 7,582 | 1,027 | 49 | 6 | 68 |
| S. griseus IFO13350 | 8,545,929 | 72.2 | 7,138 | 1,055 | 132,910 | 6 | 66 |
| S. hygroscopicus 5008 | 10,145,833 | 71.9 | 8,849 | 952 | 14 | 6 | 68 |
| S. scabies 87.22 | 10,148,695 | 71.5 | 8,746 | 1,005 | 18,488 | 6 | 75 |
Streptomyces rochei 7434AN4 produces two structurally unrelated polyketides, lankacidin C and lankamycin (Fig. 1)8,9, and carries three characteristic linear plasmids (pSLA2-L, -M, and -S), whose nucleotide sequences were previously determined (Table 2). The 210,614-bp largest linear plasmid, pSLA2-L (Accession number; AB088224), carries 143 open reading frames (ORFs), including the biosynthetic gene clusters for lankacidin, lankamycin, uncharacterized type-II polyketide, and carotenoid10. This plasmid carries many regulatory genes and the biosynthetic gene for the signaling molecules SRBs (Streptomyces rochei butenolides) (Fig. 1) that induce lankacidin and lankamycin production in S. rochei11. The 113,464-bp linear plasmid pSLA2-M (AB597522) comprises of 121 ORFs and carries several self-defense genes including a CRISPR (clustered regularly interspaced short palindromic repeats) cluster and a ku70/ku80-like gene12. Both plasmids harbor a tap-tpg gene pair that encodes a telomere-associated protein and a TP necessary for end patching of linear replicons13. The 17,526-bp smallest plasmid pSLA2-S (AB905437) consists of 17 ORFs and does not contain a tap-tpg gene pair in contrast to pSLA2-L and -M. Basic features of the three linear plasmids are shown in Table 2.
Figure 1.
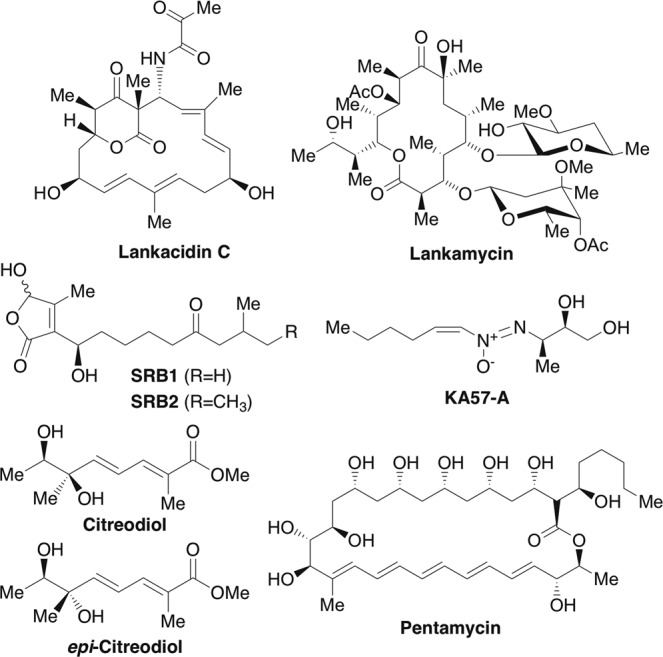
Secondary metabolites produced by Streptomyce rochei 7434AN4 and its mutants. Polyketide antibiotics; lankacidin C and lankamycin. Signaling molecules; SRB1 and SRB2. Azoxyalkene; KA57-A. Polyketides; citreodiol, epi-citreodiol, and pentamycin.
Table 2.
General features of the chromosome and three plasmids of S. rochei 7434AN4.
| Repliconsa | Length (bp) | G + C content (%) | CDS (no.) | Average CDS length (bp) | TIR (bp) |
|---|---|---|---|---|---|
| Chromosome | 8,364,802 | 71.7 | 7,568 | 973 | 53,892 |
| pSLA2-L | 210,614 | 72.8 | 143 | 1,330 | 1,992 |
| pSLA2-M | 113,464 | 69.7 | 121 | 718 | 352 |
| pSLA2-S | 17,526 | 69.7 | 22 | 508 | 817 |
Regarding the genomic structures in S. rochei 7434AN4, the right end sequences of pSLA2-L and -M are almost identical (99.9%) up to 14.6 kb from the end12. Partial sequencing and Southern blot analysis revealed that both ends of the chromosome are identical to each other, and share 98.5% homology with the right end of pSLA2-L and -M up to 3.1 kb. Furthermore, a truncated tpg homolog was detected in one contig14. In addition, curing of pSLA2-L from strain 51252, which harbors only pSLA2-L, caused terminal deletions of the chromosome followed by circularization in mutant 2–399,14. These results suggest that the tap-tpg of pSLA2-L or -M functions for terminal replication to maintain a linear topology of the chromosome. This hypothesis was supported by complementation and curing experiments of the tap-tpg of pSLA2-M14. However, the absence of a tap-tpg pair on the chromosome still remained to be proved by genome sequencing.
Streptomyces species are well known for their three characteristics: possession of linear replicons, complex morphological differentiation, and an ability to produce secondary metabolites. In this study, we have determined the complete nucleotide sequence of the linear chromosome of Streptomyces rochei 7434AN4 and extensively analyzed these three characteristics in comparison with other Streptomyces strains hitherto characterized.
Results and Discussion
Nucleotide sequencing and physical analysis of the linear chromosome of S. rochei 7434AN4
The complete nucleotide sequence of the linear chromosome of S. rochei 7434AN4 was obtained by assembling a combination of reads from the long-read PacBio RS-II sequencing together with short-read Illumina Genome Analyzer IIx (GAIIx) sequencing and Roche 454 pyrosequencing. The 26,119,215 trimmed reads (217-fold coverage of the whole genome) obtained through Illumina GAIIx sequencing were assembled using ABySS protocol to give 340 contigs with >500 bp length. Then, the PacBio RS-II sequencing independently generated 598.1 Mb of sequence data (69-fold coverage). After extensive read assembly and correction among these sequencings with the help of sequence data from Roche 454 pyrosequencing, eight contigs were obtained (Fig. 2). Contigs 1 and 2 harbored a 3.1-kb homologous sequence with the right end of pSLA2-L and -M, locating them at both ends of the chromosome. Opposite boundaries in contigs 1 and 2 harbored downstream of 5S-23S rRNA-encoding genes (rDNAs) (gray and blue arrows, respectively, in Fig. 2-ii). Five contigs, 3, 4, 6, 7, and 8, contained downstream of 5S-23S rDNAs and upstream of 16S rDNA (red arrows in Fig. 2-ii) at both ends. Contig 5 harbored 16S rDNA at both ends, indicating the presence of seven rRNA operons (16S-23S-5S) on the S. rochei chromosome. Unlinked contig gaps were filled by conventional PCR amplification using KOD-plus Neo DNA polymerase and some group-specific 16S and 5S rRNA primer sets (Table S1). Seven amplified PCR fragments (ca. 6 kb) covering each rRNA operon were sequenced and the connectivities of eight contigs were confirmed. The final assembled sequence revealed an 8,364,802-bp linear chromosome with a G + C content of 71.7% and 7,568 predicted coding DNA sequences (CDSs). Since we previously reported the nucleotide sequences of the three linear plasmids, pSLA2-L (210,614 bp)10, -M (113,464 bp)12, and -S (17,526 bp)14, the gross genomic sequence of strain 7434AN4 has now been determined to be 8,706,406 bp.
Figure 2.

Schematic representation of the S. rochei chromosome. Scale bars are drawn in megabases. (i) AseI physical map. The possible core region of the S. rochei chromosome (1.31–6.80 Mb) is marked as yellow. (ii) Distribution of rRNA-encoding gene (rDNA) operons and eight assembled contigs. Gray and blue arrows are 5S-23S rDNA operon, while red is 16S rDNA. (iii) Distribution of tRNAs. (iv) Distribution of secondary metabolite gene clusters. (v) Distribution of CDSs according to direction of transcription (+ strand, upper line; − strand, lower line). (vi) GC-skew for 10-kb window and 500-bp step. The putative oriC (gene) locus is indicated by a blue arrow. Main features were generated by DNA plotter software (https://www.sanger.ac.uk/science/tools/dnaplotter)15.
The linear topology of the chromosome was further supported by pulsed-field gel electrophoresis (PFGE) analysis of AseI and DraI digest (Fig. S2). All of the predicted AseI fragments, namely six fragments larger than 610 kb, three fragments ranging 285–450 kb, and five fragments below 225 kb were detected, except for the largest fragment AseI-A (~2.25 Mb). Southern hybridization analysis of PFGE fragments probed by PCR fragments containing a flanking AseI site agreed with the calculated AseI map (Fig. S2). All of the predicted DraI fragments, four larger than 945 kb, three ranging 285–450 kb, and three below 225 kb, were detected (Fig. S2).
General features of the linear chromosome of S. rochei 7434AN4
The general features of the S. rochei 7434AN4 chromosome are summarized in Tables 1 and 2. In addition, features including distribution of rDNA operons, tRNAs, BGCs, and CDSs according to direction of transcription (+ strand, upper line; − strand, lower line) as well as GC-skew diagram (Fig. 2) were generated by DNA plotter software (https://www.sanger.ac.uk/science/tools/dnaplotter)15. Linear chromosomes and linear plasmids of Streptomyces generally contain TIRs at both ends1. Previous Southern blot analysis with the pSLA2-L end probe indicated that the size of the TIRs of the 7434AN4 chromosome is shorter than 70 kb14. The present study revealed that the 7434AN4 chromosome has 53,892-bp TIRs (Fig. 3a). The inside ends of the TIRs were analyzed by Southern hybridization using a 3.0-kb PCR fragment (nt 52,079–55,053) as a probe (Fig. 3b). When 7434AN4 total DNA was digested with BamHI, two expected signals appeared at 3.5 kb (left-region of chromosome = ch-L) and 4.6 kb (right-region of chromosome = ch-R). When digested with XhoI, two signals were observed at 5.9 kb (ch-L) and 8.6 kb (ch-R). These results coincide with the obtained sequence data. The length of TIR varies among Streptomyces species, generally from tens to hundreds kilobases16. Ubiquitous presence of relatively long TIRs at both ends of Streptomyces linear replicons led to the idea that they might function to maintain a linear topology17. TIRs potentially provide a suitable location for homologous recombination, when one TIR is lost by terminal deletion. If recombination occurs inside of TIRs, it regenerates intact TIRs and if does outside of TIRs, it results in arm replacement18,19, both of which could recover intact termini. However, exceptionally short TIRs were found in S. hygroscopicus 5008 (14 bp)20 and in S. avermitilis (49 bp)6 (Table 1).
Figure 3.

Terminal inverted repeat (TIR) of the S. rochei chromosome. (a) Nucleotide sequence comparison of inside ends of the TIR regions. Identical sequences are indicated by asterisks. Ch-L, chromosome left region; Ch-R, chromosome right region. (b) Southern hybridization analysis of the TIR regions. PCR fragment harboring nt 52,079–55,053 was used as a DNA probe to distinguish the TIR boundary. λ-DNA digested with HindIII was used as a DNA size marker. Lane M, λ/HindIII marker; lane 1, S. rochei 7434AN4 total DNA digested with BamHI; lane 2, S. rochei 7434AN4 total DNA digested with XhoI. (c) Restriction map of the left and right TIR regions of the S. rochei 7434AN4 chromosome. TIR regions of the chromosome are shown by thick black lines. Terminal proteins attached to the 5′-ends are indicated by filled circles. Some important restriction sites are indicated. Bg, BglII; Ba, BamHI; Xh, XhoI; Ec, EcoRI; Kp, KpnI.
Telomere-associated protein and terminal protein coded by tap and tpg, respectively, are necessary for terminal replication of Streptomyces linear replicons13. However, only a truncated tpg homolog (SRO_0241, 136 aa) without a counterpart of tap was found on the S. rochei chromosome. This gene is identical to the truncated tpg homolog in contig 95 reported previously by our group14 and shows a short but high homology only to the central part of tpg; the gene product of SRO_0241 (136 aa) shows 84% (29/35) and 69% (24/35) identity to that of tpgR1 of pSLA2-L (184 aa) and tpgRM of pSLA2-M (184 aa), respectively. This result suggests that the original tpg gene on the chromosome might have been truncated at both 5′- and 3′-sides to generate a fusion gene SRO_0241 with no function. Thus, the absence of a tap-tpg gene pair on the chromosome of strain 7434AN4 has now been confirmed by genome sequencing. In addition, our hypothesis that the lack of tap-tpg is rescued in S. rochei 7434AN4 by introducing pSLA2-L and -M14, has also been proved. In this connection, the smallest plasmid pSLA2-S (17,526 bp) lacks a tap-tpg gene pair. It is reasonable that we could not obtain a mutant carrying only pSLA2-S from S. rochei 7434AN4, although all other mutants carrying possible combinations of three linear plasmids were obtained9.
The S. rochei chromosome contained seven rRNA operons (Fig. 2 and Table S2) and 67 tRNA genes (from 43 families) (Table S3). The replication origin oriC of Streptomyces is generally located between dnaA and dnaN, and contains at least 19 dnaA box-like sequences21. A putative oriC of the S. rochei 7434AN4 chromosome was located at nt. 4,097,668–4,098,732 about 80 kb from the center toward the left end, which also contains 19 dna-like boxes (Figs S2 and 2).
TTA codons are rare in Streptomyces species due to their high G + C content (typically more than 70%); for example, TTA-bearing genes comprised 1.7 and 3.4% of the S. coelicolor and S. avermitilis genomes, respectively22. On the S. rochei 7434AN4 chromosome, 225 CDSs (2.9%) contain TTA codons (Table S4) (SRO_0031 and SRO_7538 are duplicates since they are in the TIR regions of the chromosome). Distribution of TTA-bearing CDSs in five Streptomyces genomes, S. coelicolor, S. avermitilis, S. griseus, S. hygroscopicus, S. scabies, and S. rochei, were analyzed by reciprocal BLAST-P search (Table S5). Among 225 TTA-bearing CDSs on the S. rochei chromosome, 182 (81%) are specific for S. rochei and 30 (13%) are shared with the closely related species S. hygroscopicus, suggesting that TTA-bearing CDSs are species-specific in Streptomyces. Since TTA, one of the six leucine codons, is rare in streptomycetes, bldA, a gene for UUA-specific tRNA, has a crucial role in morphological differentiation and antibiotic production23–25. Among 225 TTA-bearing CDSs on the S. rochei chromosome, 17 CDSs (yellow boxes in Table S4) are involved in secondary metabolite biosynthesis, including polyketide synthases and non-ribosomal peptide synthetases. In the pentamycin biosynthetic gene cluster (BGC) (Fig. S4), five of 12 CDSs (pemA1, pemA2, pemA5, pemC, and pemR) contain a TTA codon. On the contrary, the BGC for filipin (=14-deoxo-pentamycin) in S. avermitilis contains only one TTA-bearing CDS (pteR; a homolog of pemR), suggesting that pentamycin production in S. rochei is strictly controlled under bldA-dependent regulon.
Comparative analysis of the S. rochei chromosome with the genomes of S. coelicolor A3(2), S. avermitilis, S. griseus, and S. hygroscopicus
Most Streptomyces species, including S. coelicolor A3(2), S. avermitilis, and S. griseus, have six rrn operons (16S-23S-5S), however, S. rochei 7434AN4 has seven rrn operons (Table 1). Unusual number of rrn operons were also reported for S. albus J1074 (seven operons)26 and for S. xiamenensis 318 (five operons)27. As shown in Fig. S3, six rrn operons (rrnA, rrnC, rrnD, rrnE, rrnF, and rrnG) in S. rochei are located between the highly conserved ORFs; for example, rrnA operon between putative beta-lactamase (SRO_2104; upstream of 16S) and putative aminotransferase (SRO_2103; downstream of 5S), and rrnG between phosphoenolpyruvate-dependent sugar phosphotransferase (SRO_6100; upstream of 16S) and CDP-alcohol phosphatidyltransferase (SRO_6101; upstream of 5S). The boundary regions around the seventh rrn operon, designated as rrnB in S. rochei (SRO_2782 at upstream of 16S, and SRO_2781 at upstream of 5S) are apparently different from those in S. albus J1074.
We then compared all CDSs on the chromosome of five Streptomyces strains, S. coelicolor A3(2), S. avermitilis, S. griseus, S. hygroscopicus, and S. rochei, using two independent in silico analyses; (1) orthologous clustering analysis by OrthoVenn Analysis Software (http://www.bioinfogenome.net/OrthoVenn/)28 and (2) pair-wise genome alignments by GenomeMatcher, a graphical interface for comparative genomics29. In ortholog clustering analysis, the CDSs on the S. rochei chromosome were classified into 5,370 clusters, among which 3,363 orthologs (44.4%) were shared in 5 strains (Fig. 4). In pair-wise genome alignments, the S. rochei chromosome contains a highly conserved core region (around nt 1.31–6.83 Mb) compared with the four reference strains (Fig. S5). However, a large genomic inversion was detected in S. rochei. When compared with S. avermitilis, a 1.54 Mb inversion was observed at 3.34–4.88 Mb region of the S. rochei chromosome, which corresponds to the 6.08–4.31 Mb region of the S. avermitilis chromosome.
Figure 4.
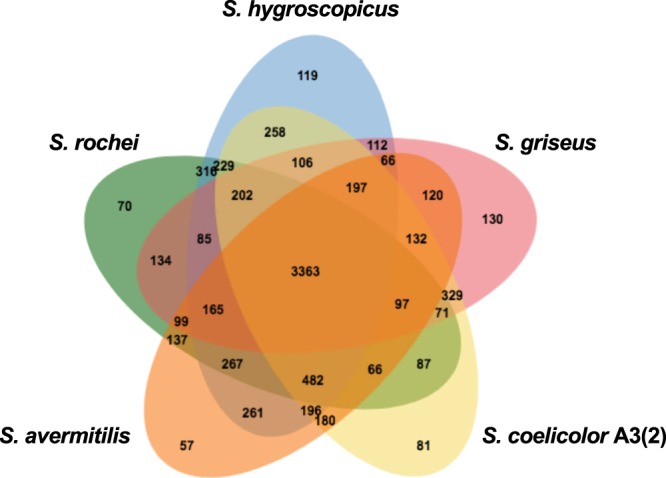
Venn diagram of the number of shared and unique genes between S. rochei and four other Streptomyces strains. OrthoVenn, a web-based application (http://www.bioinfogenome.net/OrthoVenn/) was used in this analysis. Other Streptomyces strains used in this analysis are S. coelicolor A3(2), S. avermitilis MA-4680, S. griseus IFO13350, and S. hygroscopicus 5008.
Biosynthetic gene clusters (BGCs) for secondary metabolites
We have focused on secondary metabolite biosynthetic machineries and regulatory pathways coded on pSLA2-L4,11,30–33. Several mutations including regulatory genes and major biosynthetic pathways coded on pSLA2-L led to activation of “silent” secondary metabolite clusters for pentamycin, citreodiol, epi-cireodiol, and KA57-A (Fig. 1)34–36. However, none of these biosynthetic gene clusters were found on pSLA2-L, suggesting their presence on the chromosome. Based on the 2ndFind database search (http://biosyn.nih.go.jp/2ndFind/) together with antiSMASH platform37, 35 BGCs for secondary metabolites were predicted on the S. rochei chromosome. These BGCs were classified into the following groups (Table 3); 8 for polyketides (PKSs), 8 for non-ribosomal peptides (NRPSs), 3 for hybrid PKS/NRPSs, 3 for lantibiotics, 5 for terpenes, 3 for siderophores, 1 for azoxyalkene, 1 for pseudosugar, 1 for butyrolactone, 1 for melanine, and 1 for ectoine (Table 3). The total length of BGCs (588 kb) occupied 7.03% of the S. rochei chromosome, which is comparable to other Streptomyces strains (594 kb and 6.6% for S. avermitilis).
Table 3.
List of secondary metabolite biosynthetic gene clusters in the S. rochei chromosome.
| Clustera | Product type | Location | Length (bp) | Possible metabolites |
|---|---|---|---|---|
| 1 | PKS-NRPS | SRO_0155-SRO_0157 | 12,171 | |
| 2 | Siderophore | SRO_0365-SRO_0370 | 8,474 | |
| 3 | Terpene | SRO_0471-SRO_0472 | 2,153 | 2-Methylisoborneol |
| 4 | PKS-NRPS | SRO_0501-SRO_0517 | 49,191 | |
| 5 | PKS | SRO_0730-SRO_0739 | 15,137 | |
| 6 | Lantipeptide-NRPS | SRO_0832-SRO_0848 | 34,433 | |
| 7 | PKS-NRPS | SRO_1003-SRO_1008 | 13,813 | |
| 8 | NRPS-Lantipeptide | SRO_1184-SRO_1189 | 11,055 | |
| 9 | Terpene | SRO_1200-SRO_1204 | 6,711 | Hopene |
| 10 | NRPS | SRO_1300-SRO_1346 | 70,044 | |
| 11 | NRPS | SRO_1551-SRO_1557 | 21,989 | |
| 12 | PKS-NRPS | SRO_1613-SRO_1615 | 11,979 | |
| 13 | Terpene | SRO_1735-SRO_1737 | 4,337 | Geosmin |
| 14 | Azoxyalkene and type I PKS | SRO_1819-SRO_1850 | 36,010 | KA57-A |
| 15 | Siderophore | SRO_2036-SRO_2038 | 4,744 | |
| 16 | NRPS | SRO_2051-SRO_2053 | 5,458 | |
| 17 | NRPS | SRO_2500-SRO_2506 | 9,881 | |
| 18 | Terpene | SRO_2585-SRO_2586 | 2,410 | Albaflavenone |
| 19 | Butyrolactone | SRO_3382 | 932 | |
| 20 | NRPS-Lantipeptide | SRO_3845-SRO_3849 | 6,875 | |
| 21 | Type II PKS | SRO_4037-SRO_4044 | 7,077 | Spore pigment |
| 22 | Siderophore | SRO_4684-SRO_4688 | 5,997 | |
| 23 | Melanin | SRO_4761-SRO_4762 | 1,446 | Melanin |
| 24 | Ectoine | SRO_5592-SRO_5595 | 3,165 | 5-Hydroxyectoine |
| 25 | Type I PKS | SRO_6166-SRO_6168 | 12,248 | |
| 26 | Type III PKS | SRO_6250-SRO_6251 | 2,284 | |
| 27 | NRPS | SRO_6287-SRO_6291 | 9,037 | |
| 28 | NRPS | SRO_6304-SRO_6312 | 46,390 | |
| 29 | Type I PKS (Iterative) | SRO_6377-SRO_6381 | 12,789 | |
| 30 | Pseudosugar | SRO_6636-SRO_6646 | 12,173 | |
| 31 | NRPS | SRO_7209-SRO_7215 | 44,813 | |
| 32 | Type I PKS | SRO_7221-SRO_7235 | 81,482 | Pentamycin |
| 33 | Type I PKS (Iterative) | SRO_7329-SRO_7331 | 9,456 | |
| 34 | NRPS | SRO_7379-SRO_7381 | 3,029 | |
| 35 | Terpene | SRO_7452-SRO_7458 | 8,979 | Carotenoid |
aSecondary metabolite gene clusters were predicted by 2nd Find and antiSMASH. The range of gene clusters for unknown secondary metabolites was estimated on the basis of putative functions of gene products.
Some chromosome-borne metabolites were obtained by genome mining on S. rochei36. The lkcA mutant (lkcA; an NRPS-PKS hybrid gene involved in lankacidin biosynthesis) overproduced three UV-active compounds, pentamycin, citreodiol, and epi-citreodiol (Fig. 1)34. Comparison of the pentamycin cluster (pem) with the filipin (=14-deoxypentamycin) cluster (pte) of S. avermitilis38 revealed their high homology (79–92% identities in ORFs) (Fig. S4), except for two additional genes, a P450 monooxygenase gene SRO_7222 (pemI) and a ferredoxin gene SRO_7221 (pemJ), in the former. At this stage, we have not yet identified the biosynthetic gene cluster for citreodiol and epi-citreodiol (ctr cluster). According to the reported feeding experiment39, the ctr cluster might include a C-methyltransferase (C-MT) gene for introduction of two methyl groups at C-2 and C-6. Two iterative type-I PKSs containing a C-MT domain (SRO_6380 and SRO_7330) are potential candidates for biosynthesis of citreodiols, whose gene inactivation is in progress in our laboratory. We isolated an azoxyalkene compound KA57-A (Fig. 1) from a genetically engineered strain KA57, which contains triple mutations on srrB (a tetR-type receptor gene), lkcF-KR1 (a ketoreductase domain 1 of lkcF for lankacidin biosynthesis), and lkmE (a type-II thioesterase gene for lankamycin biosynthesis) coded on pSLA2-L35. KA57-A has a unique azoxy group (N = N+-O−) and its biosynthetic gene (azx) cluster was located at nt 2,061,273–2,097,283 of the chromosome by comparison with the BGC for valanimycin40. It is noteworthy that production of these four metabolites coded on the chromosome was activated by mutation of the genes coded on pSLA2-L, indicating that the linear plasmid affects not only on a topology of the linear chromosome but also on secondary metabolite production in S. rochei.
Streptomyces species produce branched-chain fatty acids for both primary and secondary metabolism41. Branched-chain amino acids, isoleucine, valine, and leucine, are converted to the corresponding 2-oxoacids, which were then decarboxylated to form 2-methylbutyryl-CoA, isobutyryl-CoA, and isovaleryl-CoA, respectively, by the branched-chain 2-oxoacid dehydrogenase complex (Fig. S6). In the biosynthesis of Streptomyces signaling molecules42, branched-chain β-ketoacyl-CoA esters (C8-C13 in length) are condensed with a dihydroxyacetone phosphate unit by specific enzyme such as AfsA (in S. griseus), ScbA (in S. coelicolor), BarX (in S. virginiae), and SrrX (in S. rochei). In S. rochei, the branched-chain fatty acid starter units for SRB1 and SRB2 (Fig. 1) are isobutyrate and (S)-2-methylbutyrate, respectively. Furthermore, the macrolide skeleton of lankamycin (Fig. 1) is also derived from one (S)-2-methylbutyrate starter unit and six malonyl-CoA extender units. The biosynthetic gene cluster bkdFGH for isobutyrate and (S)-2-methylbutyrate43 was also found on the S. rochei chromosome (SRO_3599, 3598, and 3597 for bkdF, G and H, respectively) (Fig. S6).
Other genes
Comparative analysis of protein families with other four Streptomyces genome (S. coelicolor A3(2), S. avermitilis, S. griseus, and S. hygroscopicus) (Table S6) revealed that S. rochei 7434AN4 has relatively larger proportions of two-component histidine kinase gene homologs (113 vs 58 on average) and ABC transporter-related genes (338 vs 220 on average), reflecting a great extent of signal transduction and material transport. Strain 7434AN4 harbors 36 sigma factors and 21 ECF sigma factors. Homologous gene encoding the principal sigma factor σhrdB 44 was identified as SRO_2011. Two sporulation-related sigma factors, σBldN/AdsA for aerial mycelium formation in S. coelicolor A3(2)/S. griseus45,46 and σWhiG for onset of spore formation in S. coelicolor A3(2)47, were also identified as SRO_3261 and SRO_2209, respectively. In addition, other important genes, σR for response to oxidative stress in S. coelicolor A3(2)48 and σshbA for governing σhrdB in S. griseus49 were also identified as SRO_2590 and SRO_2981, respectively.
CRISPR (clustered regularly interspaced short palindromic repeats) is an RNA-dependent immune system widely distributed in bacteria and archaea against infection of foreign genetic elements including phages and plasmids50. The CRISPR-associated genes (cas genes) were identified from SRO_1948 to SRO_1955 (Table S7). Flanking this cluster, 19 DNA repeats (CGGTTCACCTCCGCCTGCGCGGAGCGGAC; 29 bases) were located upstream at nt. 2,205,703–2,206,838 and 6 repeats were downstream at nt. 2,216,850–2,217,183 (Table S8). All the Cas proteins showed considerable similarity with annotated cas gene products in other Streptomyces strains. The linear plasmid pSLA2-M has 49 repeat sequences at the right end of the CRISPR cluster (ORF94-ORF101)12, however, whose consensus sequences, 5′-GTGGCGGTCGCCCTCCGGGGTGACCGAGGATCGCAAC-3′ (37 bases), are different from that on the chromosome.
Analysis of three plasmidless mutants, S. rochei 2-39, YN-P7, and YN-P145
We previously prepared three plasmidless mutants, 2-39, YN-P7, and YN-P145 by protoplast regeneration of S. rochei 51252 that contains only pSLA2-L9,14. In the case of mutant 2-39, chromosomal deletion at both ends followed by circularization was confirmed by cloning of the fusion junction14. Although mutant 2-39 lost around 20% (1.55 Mb) of the chromosome (Fig. 5), seven rRNA operons were still conserved. PCR amplification showed that strains YN-P7 and YN-P145 also keep seven rrn operons, suggesting the importance of all rrn operons in S. rochei. Three mutants exhibited a different phenotype when grown on YM solid medium (Fig. 5); mutants YN-P7 and YN-P145 showed a “white” phenotype, while strain 2-39 a “bald” phenotype (Fig. 5a). To analyze morphological differentiation more precisely, their colonies were observed by scanning electron microscopy (SEM) (Fig. 5b). Chain elongation of aerial mycelium of mutant 2-39 stopped at an early stage, while mutant YN-P7 produced longer but collapsed hyphae. On the other hand, mutant YN-P145 produced partially spiral spore chains although their development is significantly lesser than that in strain 51252. Based on the Illumina sequence data of the three mutants, the bald strain 2-39 was found to suffer a larger chromosomal deletion from the right end (1,090 kb) compared with strains YN-P7 (913 kb) and YN-P145 (934 kb) (Fig. 5c). On the other hand, the deletions at the left chromosomal end of strains 2-39, YN-P7, and YN-P145 were 458 kb, 76 kb, and 603 kb, respectively. Based on the phenotype-genotype correlation in strains 2-39, YN-P7 and YN-P145, we speculate that essential gene(s) responsible for converting vegetative hyphae into aerial hyphae is(are) located at nt 7,275–7,431 kb of the 7434AN4 chromosome. This region harbors 138 ORFs (SRO_6607-SRO_6744), among which some gene(s) may be responsible for aerial hyphae formation in S. rochei.
Figure 5.

Morphological differentiation of three plasmidless mutants of S. rochei and their chromosomal deletion. (a) Spore formation of S. rochei strains. Strains (2-39, YN-P7, YN-P145, and their parent 51252) were grown on YM agar medium at 28 °C for 5 days. (b) Scanning electron microscopy (SEM) of surface grown colonies. (c) Chromosomal deletions in mutants 2-39, YN-P7, and YN-P145.
Conclusion
In this study, we have determined the nucleotide sequence of the 8,364,802-bp linear chromosome of S. rochei 7434AN4, which in turn revealed the gross genome size (8,706,406 bp) of this strain including the three linear plasmids, pSLA2-L, -M, and -S. General features of the linear chromosome were presented; it carries seven rrn operons, 67 tRNA genes, 225 TTA-containg CDSs, and 53.9-kb TIRs at both ends. In particular, the absence of a tpg-tap gene pair on the chromosome has proved our hypothesis that the tpg-tap pairs of pSLA2-L and/or pSLA2-M function to maintain a linear topology of the chromosome in strain 7434AN4.
In silico analysis indicated the presence of 35 secondary metabolites gene clusters on the chromosome, whose functions are not known in most cases. Therefore, we could expect that studies on their functions and regulation, particularly interaction with the regulatory genes coded on pSLA2-L will lead to a discovery of new antibiotics and their improved production.
Materials and Methods
Strains, plasmids, oligonucleotides, and culture media
All the strains, plasmids, and oligonucleotides used in this study were listed in Table S1. YEME liquid medium (0.3% yeast extract, 0.5% peptone, 0.3% malt extract, 1.0% D-glucose, 34% sucrose, 5 mM MgCl2, and 0.5% glycine) was used for preparation of total genomic DNA. YM medium (0.4% yeast extract, 1.0% malt extract, and 0.4% D-glucose, pH 7.3) was used for routine cultivation.
DNA sequencing and assembly
S. rochei 7434AN4 was sequenced using hybrid approach of next-generation sequencing platforms; PacBio RS-II, Illumina GAIIx, and Roche 454 sequencers.
Genomic DNA of strain 7434AN4 was subjected to paired-end sequencing using Illumina GAIIx sequencing system (San Diego, CA, USA) according to the manufacture’s protocol. The 26,119,215 trimmed reads with 217-fold coverage of the whole genome were assembled using ABySS 1.3.751. Illumina read data has been deposited as DRA Accession DRA003131 and DRA003132, Bioproject: PRJDB3565, Biosample: SAMD00027156. Independently, long-read sequencing was performed on PacBio RS-II sequencing system (Pacific Biosciences; Menlo Park, CA, USA). The filtered subreads with 598,149,379 bp in length (69-fold coverage) from PacBio RS-II was then assembled using the Hierarchical Genome Assembly Process (HGAP). The assembly consists of 175 contigs of 8,204,607 bp with an average length of 46,883 bp. Both sequence data was extensively compared and corrected with a help of Roche 454 pyrosequencing to obtain eight contigs (Fig. 2; length of contigs 1-8 were 2,393,094 bp, 1,685,790 bp, 1,286,111 bp, 750,875 bp, 720,184 bp, 567,335 bp, 501,529 bp, and 462,928 bp, respectively. Details for Roche 454 pyrosequencing were described previously14. Sequence gaps among eight contigs were then religiously filled and connected by conventional PCR amplification. Complete nucleotide sequence of the S. rochei 7434AN4 chromosome has been deposited at DDBJ under Accession number AP018517.
Sequence annotation and comparative analysis
Putative coding sequences (CDSs), tRNA-, and rRNA-coding sequences were predicted using Microbial Genome Annotation Pipeline (MiGAP) platform (https://www.migap.org/) and FramePlot 2.3.2 (http://www0.nih.go.jp/~jun/cgi-bin/frameplot.pl)52. Their putative annotation was manually confirmed by a BLASTP program (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins). Main features including distribution of rDNA operons, tRNAs, BGCs, and CDSs according to direction of transcription (+ strand, upper line; − strand, lower line) as well as GC-skew diagram (Fig. 2) were generated by DNA plotter software (https://www.sanger.ac.uk/science/tools/dnaplotter)15. The locus of oriC was predicted manually based on the genome information of other Streptomyces species. The protein families were clustered with OrthoVenn Analysis Software (http://www.bioinfogenome.net/OrthoVenn/), a web platform for comparison and annotation of orthologous gene clusters among multiple species28. The comparative analysis of the chromosomes between S. rochei 7434AN4 and other Streptomyces species was performed using GenomeMatcher software (http://www.ige.tohoku.ac.jp/joho/gmProject/gmhomeJP.html)29 and bl2seq program, which is embedded in the application bundled. Secondary metabolite gene clusters were predicted by either 2ndFind software, a web-based analytical tool (http://biosyn.nih.go.jp/2ndFind/), or antiSMASH 2.0, a web-based analysis platform (http://antismash.secondarymetabolites.org/)37. CRISPRs were predicted using a CRISPRFinder (http://crispr.i2bc.paris-saclay.fr/Server/), an online program.
DNA manipulation and Southern hybridization
Streptomyces strains were grown in liquid YM medium in Sakaguchi flask at 28 °C for 3 days. DNA manipulation of Streptomyces species53 was carried out according to standard procedure. Genome DNA sample of S. rochei 7434AN4 for PFGE was obtained according to the method as described previously9 with a slight modification. Polymerase chain reaction (PCR) was performed on a GeneAtlas G02 Thermal Cycler (Astec Co. Ltd., Fukuoka, Japan) using KOD-plus Neo DNA polymerase (Toyobo, Osaka, Japan) according to the manufacture’s protocol. Southern blot analysis (Figs 3, 5 and S2) was performed as described previously14.
Scanning electron microscopy (SEM)
The surface morphology of S. rochei strain 51252 and three plasmidless mutants was observed by scanning electron microscopy (SEM) after growing on YM agar plate for 5 days. For the preparation of specimens, agar plugs were fixed with 1% osmium tetroxide solution for 12 h, and then dehydrated by lyophilization. The resulting specimens were coated with platinum (2 nm) and observed by a Jeol JSM-5900 Scanning Electron Microscope.
Supplementary information
Acknowledgements
We thank to Dr. D. Kajiya and Mrs. T. Amimoto (N-BARD, Hiroshima University) for measurement of the high resolution mass spectra, and Mrs. K. Koike (N-BARD, Hiroshima University) for SEM analysis. This work was supported by Grants-in-Aid for Scientific Research on Innovative Areas (23108515, 25108718 and 17H05446 to K.A.) from Ministry of Education, Culture, Sports, Science and Technology of Japan (MEXT), Grants-in-Aid for Scientific Research (B) (16H04917 to K.A.) from the Japan Society for the Promotion of Science (JSPS), and the Sasakawa Scientific Research Grant from the Japan Science Society to Y.N. This work was partly supported by a JSPS A3 Foresight Program. A.A.F. and R.M. were supported by the Indonesia Endowment Fund for Education (LPDP). Sequencing analysis using an Illumina GAIIx sequencer was supported by the Grant in Aid for Scientific Research on Innovative Areas (22108010 to J.I.) from MEXT.
Author Contributions
Y.N., Z.C., A.A.F., K.I., H.K. and K.A. designed the experiments, Y.N., Z.C., A.A.F., Y.M. and K.I. performed the experiments, Y.N., A.A.F., A.T., Y.M., R.M., Y.Y., K.I. and K.A. analyzed the data, Y.S., H.Y., M.T., A.L., J.I., M.K. and T.S. performed the next-generation sequencing and extensive assembly, and Y.N., K.I., H.K. and K.A. wrote the manuscript with input from all of the authors. All the authors approved the final version of the manuscript.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary information accompanies this paper at 10.1038/s41598-019-47406-y.
References
- 1.Lin Y-S, Kieser HM, Hopwood DA, Chen CW. The chromosomal DNA of Streptomyces lividans 66 is linear. Mol Microbiol. 1993;10:923–933. doi: 10.1111/j.1365-2958.1993.tb00964.x. [DOI] [PubMed] [Google Scholar]
- 2.Chen CW, et al. The linear chromosomes of Streptomyces: Structure and dynamics. Actinomycetol. 1994;8:103–112. doi: 10.3209/saj.8_103. [DOI] [Google Scholar]
- 3.Volff JN, Altenbuchner J. Genetic instability of the Streptomyces chromosome. Mol Microbiol. 1998;27:239–246. doi: 10.1046/j.1365-2958.1998.00652.x. [DOI] [PubMed] [Google Scholar]
- 4.Kinashi H. Antibiotic production, linear plasmids and linear chromosomes in Streptomyces. Actinomycetol. 2008;22:20–29. doi: 10.3209/saj.SAJ220104. [DOI] [Google Scholar]
- 5.Bentley SD, et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2) Nature. 2002;417:141–147. doi: 10.1038/417141a. [DOI] [PubMed] [Google Scholar]
- 6.Ikeda H, et al. Complete genome sequence and comparative analysis of the industrial microorganism Streptomyces avermitilis. Nat Biotechnol. 2003;21:526–531. doi: 10.1038/nbt820. [DOI] [PubMed] [Google Scholar]
- 7.Ohnishi Y, et al. Genome Sequence of the Streptomycin-Producing Microorganism Streptomyces griseus IFO 13350. J Bacteriol. 2008;190:4050–4060. doi: 10.1128/JB.00204-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hayakawa T, Tanaka T, Sakaguchi K, Otake N, Yonehara H. A linear plasmid-like DNA in Streptomyces sp. producing lankacidin group antibiotics. J Gen Appl Microbiol. 1979;25:255–260. doi: 10.2323/jgam.25.255. [DOI] [Google Scholar]
- 9.Kinashi H, Mori E, Hatani A, Nimi O. Isolation and characterization of linear plasmids from lankacidin-producing Streptomyces species. J Antibiot. 1994;47:1447–1455. doi: 10.7164/antibiotics.47.1447. [DOI] [PubMed] [Google Scholar]
- 10.Mochizuki S, et al. The large linear plasmid pSLA2-L of Streptomyces rochei has an unusually condensed gene organization for secondary metabolism. Mol Microbiol. 2003;48:1501–1510. doi: 10.1046/j.1365-2958.2003.03523.x. [DOI] [PubMed] [Google Scholar]
- 11.Arakawa K, Tsuda N, Taniguchi A, Kinashi H. The butenolide signaling molecules SRB1 and SRB2 induce lankacidin and lankamycin production in Streptomyces rochei. Chembiochem. 2012;13:1447–1457. doi: 10.1002/cbic.201200149. [DOI] [PubMed] [Google Scholar]
- 12.Yang Y, et al. pSLA2-M of Streptomyces rochei is a composite linear plasmid characterized by self-defense genes and homology with pSLA2-L. Biosci Biotechnol Biochem. 2011;75:1147–1153. doi: 10.1271/bbb.110054. [DOI] [PubMed] [Google Scholar]
- 13.Bao K, Cohen SN. Recruitment of terminal protein to the ends of Streptomyces linear plasmids and chromosomes by a novel telomere-binding protein essential for linear DNA replication. Genes Dev. 2003;17:774–785. doi: 10.1101/gad.1060303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nindita Y, et al. The tap-tpg gene pair on the linear plasmid functions to maintain a linear topology of the chromosome in Streptomyces rochei. Mol Microbiol. 2015;95:846–858. doi: 10.1111/mmi.12904. [DOI] [PubMed] [Google Scholar]
- 15.Carver T, Thomson N, Bleasby A, Berriman M, Parkhill J. DNAPlotter: circular and linear interactive genome visualization. Bioinformatics (Oxford, England) 2009;25:119–20. doi: 10.1093/bioinformatics/btn578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen CW, Huang CH, Lee HH, Tsai HH, Kirby R. Once the circle has been broken: dynamics and evolution of Streptomyces chromosomes. Trends Genet. 2002;18:522–529. doi: 10.1016/S0168-9525(02)02752-X. [DOI] [PubMed] [Google Scholar]
- 17.Qin Z, Stanley N, Cohen SN. Survival mechanisms for Streptomyces linear replicons after telomere damage. Mol Microbiol. 2002;45:785–794. doi: 10.1046/j.1365-2958.2002.03051.x. [DOI] [PubMed] [Google Scholar]
- 18.Fischer G, Wenner T, Decaris B, Leblond P. Chromosomal arm replacement generates a high level of intraspecific polymorphism in the terminal inverted repeats of the linear chromosomal DNA of Streptomyces ambofaciens. Proc. Natl. Acad. Sci. USA. 1998;95:14296–14301. doi: 10.1073/pnas.95.24.14296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Uchida T, Miyawaki M, Kinashi H. Chromosomal arm replacement in Streptomyces griseus. J Bacteriol. 2003;185:1120–1124. doi: 10.1128/JB.185.3.1120-1124.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wu H, et al. Genomic and transcriptomic insights into the thermo-regulated biosynthesis of validamycin in Streptomyces hygroscopicus 5008. BMC Genomics. 2012;13:337. doi: 10.1186/1471-2164-13-337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jakimowicz D, et al. Structural elements of the Streptomyces oriC region and their interactions with the DnaA protein. Microbiology. 1998;144:1281–1290. doi: 10.1099/00221287-144-5-1281. [DOI] [PubMed] [Google Scholar]
- 22.Chandra G, Chater KF. Evolutionary flux of potentially bldA-dependent Streptomyces genes containing the rare leucine codon TTA. Anton Leeuw. 2008;94:111–126. doi: 10.1007/s10482-008-9231-5. [DOI] [PubMed] [Google Scholar]
- 23.Leskiw BK, Lawlor EJ, Fernandez-Abalos JM, Chater KF. TTA codons in some genes prevent their expression in a class of developmental, antibiotic-negative, Streptomyces mutants. Proc Natl Acad Sci USA. 1991;88:2461–2465. doi: 10.1073/pnas.88.6.2461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Takano E, et al. A rare leucine codon in adpA is implicated in the morphological defect of bldA mutants of Streptomyces coelicolor. Mol Microbiol. 2003;50:475–486. doi: 10.1046/j.1365-2958.2003.03728.x. [DOI] [PubMed] [Google Scholar]
- 25.White J, Bibb M. bldA dependence of undecylprodigiosin production in Streptomyces coelicolor A3(2) involves a pathway-specific regulatory cascade. J Bacteriol. 1997;179:627–633. doi: 10.1128/jb.179.3.627-633.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zaburannyi N, Rabyk M, Ostash B, Fedorenko V, Luzhetskyy A. Insights into naturally minimised Streptomyces albus J1074 genome. BMC Genomics. 2014;15:97. doi: 10.1186/1471-2164-15-97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Xu M-J, et al. Deciphering the streamlined genome of Streptomyces xiamenensis 318 as the producer of the anti-fibrotic drug candidate xiamenmycin. Sci Rep. 2016;6:18977. doi: 10.1038/srep18977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wang Y, Coleman-Derr D, Chen G, Gu YQ. OrthoVenn: a web server for genome wide comparison and annotation of orthologous clusters across multiple species. Nucl Acids Res. 2015;43:W78–84. doi: 10.1093/nar/gkv487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ohtsubo Y, Ikeda-Ohtsubo W, Nagata Y, Tsuda M. GenomeMatcher: A graphical user interface for DNA sequence comparison. BMC Bioinform. 2008;9:376. doi: 10.1186/1471-2105-9-376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Arakawa K, Sugino F, Kodama K, Ishii T, Kinashi H. Cyclization mechanism for the synthesis of macrocyclic antibiotic lankacidin in Streptomyces rochei. Chem Biol. 2005;12:249–256. doi: 10.1016/j.chembiol.2005.01.009. [DOI] [PubMed] [Google Scholar]
- 31.Arakawa K, Kodama K, Tatsuno S, Ide S, Kinashi H. Analysis of the loading and hydroxylation steps in lankamycin biosynthesis in Streptomyces rochei. Antimicrob Agents Chemother. 2006;50:1946–1952. doi: 10.1128/AAC.00016-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Arakawa K, Mochizuki S, Yamada K, Noma T, Kinashi H. γ-Butyrolactone autoregulator-receptor system involved in lankacidin and lankamycin production and morphological differentiation in Streptomyces rochei. Microbiology. 2007;153:1817–1827. doi: 10.1099/mic.0.2006/002170-0. [DOI] [PubMed] [Google Scholar]
- 33.Yamamoto S, He Y, Arakawa K, Kinashi H. Gamma-butyrolactone-dependent expression of the Streptomyces antibiotic regulatory protein gene srrY plays a central role in the regulatory cascade leading to lankacidin and lankamycin production in Streptomyces rochei. J Bacteriol. 2008;190:1308–1316. doi: 10.1128/JB.01383-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cao Z, Yoshida R, Kinashi H, Arakawa K. Blockage of the early step of lankacidin biosynthesis caused a large production of pentamycin, citreodiol and epi-citreodiol in Streptomyces rochei. J Antibiot. 2015;68:328–333. doi: 10.1038/ja.2014.160. [DOI] [PubMed] [Google Scholar]
- 35.Kunitake H, Hiramatsu T, Kinashi H, Arakawa K. Isolation and biosynthesis of an azoxyalkene compound produced by a multiple gene disruptant of Streptomyces rochei. ChemBioChem. 2015;16:2237–2243. doi: 10.1002/cbic.201500393. [DOI] [PubMed] [Google Scholar]
- 36.Arakawa K. Manipulation of metabolic pathway controlled by signaling molecules, inducers of antibiotic production, for genome mining in Streptomyces spp. Antonie Leeuw. 2018;111:743–751. doi: 10.1007/s10482-018-1052-6. [DOI] [PubMed] [Google Scholar]
- 37.Blin K, et al. antiSMASH 2.0-a versatile platform for genome mining of secondary metabolite producers. Nucl Acids Res. 2013;41:W204–12. doi: 10.1093/nar/gkt449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ikeda H, Shin-ya K, Ōmura S. Genome mining of the Streptomyces avermitilis genome and development of genome-minimized hosts for heterologous expression of biosynthetic gene clusters. J Ind Microbiol Biotechnol. 2014;41:233–250. doi: 10.1007/s10295-013-1327-x. [DOI] [PubMed] [Google Scholar]
- 39.Shizuri Y, Nishiyama S, Imai D, Yamamura S. Isolation and stereostructures of citreoviral, citreodiol, and epicitreodiol. Tetrahedron Lett. 1984;25:4771–4774. doi: 10.1016/S0040-4039(01)81515-X. [DOI] [Google Scholar]
- 40.Garg RP, Ma Y, Hoyt JC, Parry RJ. Molecular characterization and analysis of the biosynthetic gene cluster for the azoxy antibiotic valanimycin. Mol Microbiol. 2002;46:505–517. doi: 10.1046/j.1365-2958.2002.03169.x. [DOI] [PubMed] [Google Scholar]
- 41.Han L, Lobo S, Reynolds KA. Characterization of beta-ketoacyl-acyl carrier protein synthase III from Streptomyces glaucescens and its role in initiation of fatty acid biosynthesis. J Bacteriol. 1998;180:4481–4486. doi: 10.1128/jb.180.17.4481-4486.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Takano E. γ-Butyrolactones: Streptomyces signaling molecules regulating antibiotic production and differentiation. Curr Opin Microbiol. 2006;9:287–294. doi: 10.1016/j.mib.2006.04.003. [DOI] [PubMed] [Google Scholar]
- 43.Denoya CD, et al. A second branched-chain alpha-keto acid dehydrogenase gene cluster (bkdFGH) from Streptomyces avermitilis: its relationship to avermectin biosynthesis and the construction of a bkdF mutant suitable for the production of novel antiparasitic avermectins. J Bacteriol. 1995;177:3504–3511. doi: 10.1128/jb.177.12.3504-3511.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Buttner MJ, Chater KF, Bibb MJ. Cloning, disruption, and transcriptional analysis of three RNA polymerase sigma factor genes of Streptomyces coelicolor A3(2) J Bacteriol. 1990;172:3367–3378. doi: 10.1128/jb.172.6.3367-3378.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bibb MJ, Molle V, Buttner MJ. σBldN, an extracytoplasmic function RNA polymerase sigma factor required for aerial mycelium formation in Streptomyces coelicolor A3(2) J Bacteriol. 2000;182:4606–4616. doi: 10.1128/JB.182.16.4606-4616.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yamazaki H, Ohnishi Y, Horinouchi S. An A-factor-dependent extracytoplasmic function sigma factor (σAdsA) that is essential for morphological development in Streptomyces griseus. J Bacteriol. 2000;82:4596–4605. doi: 10.1128/JB.182.16.4596-4605.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chater KF, et al. The developmental fate of S. coelicolor hyphae depends upon a gene product homologous with the motility sigma factor of B. subtilis. Cell. 1989;59:133–143. doi: 10.1016/0092-8674(89)90876-3. [DOI] [PubMed] [Google Scholar]
- 48.Paget MS, Kang JG, Roe JH, Buttner MJ. σR, an RNA polymerase sigma factor that modulates expression of the thioredoxin system in response to oxidative stress in Streptomyces coelicolor A3(2) EMBO J. 1998;17:5776–5782. doi: 10.1093/emboj/17.19.5776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Otani H, Higo A, Nanamiya H, Horinouchi S, Ohnishi Y. An alternative sigma factor governs the principal sigma factor in Streptomyces griseus. Mol Microbiol. 2013;87:1223–1236. doi: 10.1111/mmi.12160. [DOI] [PubMed] [Google Scholar]
- 50.Haft DH, Selengut J, Mongodin EF, Nelson KE. A guild of 45 CRISPR-associated (Cas) protein families and multiple CRISPR/Cas subtypes exist in prokaryotic genomes. PLoS Comput Biol. 2005;1:e60. doi: 10.1371/journal.pcbi.0010060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Simpson JT, et al. ABySS: a parallel assembler for short read sequence data. Genome Res. 2009;19:1117–1123. doi: 10.1101/gr.089532.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ishikawa J, Hotta K. FramePlot: a new implementation of the frame analysis for predicting protein-coding regions in bacterial DNA with a high G+C content. FEMS Microbiol Lett. 1999;174:251–253. doi: 10.1111/j.1574-6968.1999.tb13576.x. [DOI] [PubMed] [Google Scholar]
- 53.Kieser, T., Bibb, M. J., Buttner, M. J., Chater, K. F. & Hopwood, D. A. Practical Streptomyces Genetics. (John Innes Foundation, Norwich, 2000).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.