

Abstract
Dengue virus (DENV) is the most prevalent mosquito-borne virus in the world and a major cause of morbidity in the tropics and subtropics. Upregulation of HLA class I molecules has long been considered a feature of DENV infection, yet this has not been evaluated in the setting of natural infection. Natural killer (NK) cells, an innate immune cell subset critical for mounting an early response to viral infection, are inhibited by self HLA class I, suggesting that upregulation of HLA class I during DENV infection could dampen the NK cell response. Here we addressed whether upregulation of HLA class I molecules occurs during in vivo DENV infection and, if so, whether this suppresses the NK cell response. We found that HLA class I expression was indeed upregulated during acute DENV infection across multiple cell lineages in vivo. To better understand the role of HLA class I upregulation, we infected primary human monocytes, a major target of DENV infection, in vitro. Upregulation of total HLA class I is dependent on active viral replication and is mediated in part by cytokines and other soluble factors induced by infection, while upregulation of HLA-E occurs in the presence of replication-incompetent virus. Importantly, blocking DENV-infected monocytes with a pan-HLA class I Fab nearly doubles the frequency of degranulating NK cells, while blocking HLA-E does not significantly improve the NK cell response. These findings demonstrate that upregulation of HLA class I during DENV infection suppresses the NK cell response, potentially contributing to disease pathogenesis.
Keywords: dengue virus, anti-viral response, human leukocyte antigen class I, HLA-E, natural killer cells, monocytes
Introduction
Dengue virus (DENV) is a positive-strand RNA virus of which there are four serotypes (DENV-1 to DENV-4). The virus is transmitted between humans by its vector, Aedes mosquitoes. Each year, an estimated 390 million people are infected with DENV (Bhatt et al., 2013). While most DENV infections are not life-threatening, severe infections can result in hemorrhage, plasma leakage, shock, organ failure, and death (Kyle and Harris, 2008). The incidence of dengue is rapidly rising (World Health Organization, 2012), increasing the need for a better understanding of how the human immune system responds to DENV infection. There is significant interest in elucidating the role of natural killer (NK) cells during DENV infection. NK cells are innate lymphoid cells that play a key role during the early stages of viral infection. Previous studies have shown that NK cells are activated in vivo during DENV infection (Azeredo, 2006; Petitdemange et al., 2016) and that activated NK cells may be an indicator of a positive prognosis (Azeredo, 2006). NK cell activation in response to virally infected cells is dependent on the balance of activating and inhibitory signals from numerous germline-encoded receptors. One such activating receptor, FcRγIIIa (CD16a), mediates antibody-dependent cell cytotoxicity (ADCC), a key bridge between the adaptive and innate immune systems in which antibodies bound to infected cells target them for NK cell killing (Laoprasopwattana et al., 2007; Sun et al., 2017, 2019). NK cells can also kill DENV-infected cells in the absence of ADCC (Costa et al., 2017). Several NK cell receptors, namely DNAM-1, NKG2D, and NKp44 have been implicated in this direct recognition of DENV-infected cells (Beltrán and López-Vergès, 2014; Petitdemange et al., 2014; Costa et al., 2017; Mathew, 2018). However, DENV may also evade the NK cell response, most notably through upregulation of HLA class I (Lobigs et al., 1996; Momburg et al., 2001; Hershkovitz et al., 2008; Glasner et al., 2017; Drews et al., 2018).
HLA class I molecules can bind inhibitory NK cell receptors, mitigating NK cell effector functions against healthy cells. The classical HLA-A, -B, and -C molecules do this by binding to various inhibitory killer-cell immunoglobulin-like receptors (KIRs). The non-classical HLA-E, which presents peptides derived from leader sequences of other HLA molecules, does this by binding to the inhibitory heterodimer CD94/NKG2A (Braud et al., 1998). Viruses can evade NK cell recognition by taking advantage of these inhibitory interactions. In vitro studies have shown flaviviruses, including DENV, upregulate total HLA class I as well as HLA-E, leading to inhibition of NK cell activation (Lobigs et al., 1996; Momburg et al., 2001; Hershkovitz et al., 2008; Glasner et al., 2017; Drews et al., 2018). Immune cells, particularly monocytes, are the main targets of DENV infection in vivo (Durbin et al., 2008). However, previous studies investigating DENV-mediated HLA class I upregulation and its effect on NK cell activation have used mouse and human cell lines derived from non-immune cells or differentiated primary immune cells (Lobigs et al., 1996; Libraty et al., 2001; Momburg et al., 2001; Cheng et al., 2004; Hershkovitz et al., 2008; Nightingale et al., 2008; Shwetank et al., 2013; Glasner et al., 2017; Drews et al., 2018). This has left a critical gap in our understanding of how undifferentiated primary human immune cell expression of HLA class I is affected by DENV infection, and whether any such changes impact NK cell responses to DENV.
We aimed to determine whether upregulation of class I HLAs, including HLA-E, occurs during in vivo DENV infection and, if so, whether this serves to suppress the NK cell response. To address this question, we analyzed peripheral blood mononuclear cell (PBMC) samples from a Panamanian cohort of adult dengue patients and healthy controls for expression of total HLA class I and HLA-E. We then used in vitro DENV-infected primary monocytes to determine mediators of HLA class I upregulation. Finally, we co-cultured primary NK cells with autologous, DENV-infected monocytes in the presence of HLA class I blocking Fabs to determine the impact of HLA class I expression on the NK cell response.
Materials and Methods
DENV Patients and Ethical Statement
Adult DENV patients with <5 days of symptoms consistent with acute DENV infection (fever over 38°C, severe headache, retro-orbital pain, intense myalgia, arthralgia, exanthema, conjunctivitis, diarrhea, chills, nausea, vomiting, abdominal pain, petechiae, and/or bleeding) were recruited at public health institutions (hospitals belonging to the Ministry of Health, the Social Security System in Panama City, Republic of Panama, and suburban areas). Healthy Panamanian control donors volunteered at Gorgas Memorial Institute of Health Studies. All dengue cases were confirmed by qRT-PCR, NS1 antigen, DENV-specific IgM, and IgG serological testing. The study protocol was approved by the IRB of Hospital del Niño (CBIHN-M-0634), then confirmed by the committees of ICGES, CSS, Santo Tomas Hospital, and Stanford University. Anonymous healthy adult PBMC samples for in vitro studies were collected from leukoreduction system chambers purchased from the Stanford Blood Center.
PBMC Sample Processing, Storage, and Thawing
PBMCs were isolated using gradient centrifugation separation by Ficoll-Paque, suspended in freezing media (90% FBS, 10% DMSO), stored at −80°C for 24–72 h, then transferred to liquid nitrogen. PBMCs were thawed, added to complete media (RPMI-1640, 10% FBS, 1% L-glutamine, 1% penicillin/streptomycin), centrifuged, and counted.
Mass Cytometry Staining, Data Acquisition, and Analysis
Antibodies for mass cytometry were conjugated using Maxpar® X8 Antibody Labeling Kits (Fluidigm). PBMCs were stained with 25 μM cisplatin (Enzo Life Sciences), live cell palladium barcoded for 30 min at 4°C (Mei et al., 2015), pooled, and stained with surface antibodies for 30 min at 4°C. Cells were fixed with 2% paraformaldehyde in PBS and permeabilized (eBioscience Permeabilization Buffer) prior to intracellular staining for 45 min at 4°C. Finally, the cells were incubated at 4°C in iridium-191/193 intercalator (DVS Sciences) for up to a week, washed once with CyPBS (10X Rockland PBS diluted to 1X in MilliQ water), washed three times with MilliQ water, and diluted with EQ Four Element Calibration Beads before being run on a Helios mass cytometer (Fluidigm). Raw FCS files were normalized using the Normalizer multivariate curve resolution. Normalized files were then de-barcoded using the ParkerICI Premessa de-barcoder. FlowJo® 10.2 was used to gate on live cells. viSNE analysis was performed in Cytobank.
Human sHLA-E ELISA Testing
Human sera from 6 DENV confirmed patients and 31 healthy donors were diluted 1:10 and assayed with the Human MHCE/HLA-E ELISA Kit (Biomatik) per manufacturer's instructions. Optical densities were used to calculate concentration (ng/mL) with a 4 parametric logistic regression analysis using GraphPad Prism 7.
Monocyte and NK Cell Preparation
Monocytes were isolated from PBMCs by negative selection using a human Pan Monocyte Isolation Kit (Miltenyi). Autologous NK cells were isolated by negative selection using a human NK Cell Isolation Kit (Miltenyi) and cultured in complete RPMI-1640 media with 300 IU/mL of IL-2 (R&D Systems) for 22 h.
DENV Infection of Primary Monocytes and Analysis of HLA Expression
Aedes albopictus C6/36 cells were infected with DENV-2 laboratory strain 429557 (NR-12216). Supernatants were harvested on day 7 or 8 post-infection, filtered, and ultracentrifuged on a D-sorbitol cushion at 59,439 RCF at 4°C for 3 h. Virus was titrated using a Vero cell focus-forming assay (Bayless et al., 2016). Concentrated virus was stored at −80°C. Virus was UV-inactivated at 500 μJ × 100 on ice in flat-bottom 96-well plates using a Stratagene UV Stratalinker. Virus inactivation was verified by focus-forming assays. All experiments were repeated with multiple virus batches. Monocytes were mock-infected, exposed to UV-inactivated DENV-2, or infected with active DENV-2 at a multiplicity of infection (MOI) of 2 for 2 h in infection media (RPMI-1640 media with 2% FBS, 1% penicillin/streptomycin, 1% L-glutamine, and 20 mM HEPES). After 2 h, cells were washed, resuspended in 24 h infection media (infection media without HEPES), and incubated at 37°C, 5% CO2 for 22 or 46 h. Cells were stained with FITC-conjugated anti-CD3 (UCHT1, BioLegend), FITC-conjugated anti-CD7 (CD7-6B7, BioLegend), PE-Cy7-conjugated anti-HLA-E (3D12, BioLegend), PE-conjugated anti-pan HLA class I (W6/32, BioLegend), flavivirus group antigen (4G2, Novus Biologicals) conjugated to Alexa FluorTM 647 using Alexa Fluor 647 Antibody Labeling Kit (Life Technologies), and LIVE/DEADTM Fixable Yellow Dead Cell Stain Kit (Life Technologies) before analysis on a MACSQuant Analyzer and FlowJo® 10.2.
Supernatant Swap Assay
Conditioned supernatants from aforementioned DENV-infected primary monocyte cultures were UV-inactivated as previously described. They were then used to culture primary monocytes from the same donors from which the supernatants were collected. After a 24 h incubation, monocytes were stained with APC-conjugated anti-CD3, APC-conjugated anti-CD7, PE-conjugated anti-pan HLA class I, and Zombie Aqua Fixable Viability dye (BioLegend), then analyzed using a CytekTM Aurora analyzer and FlowJo® 10.2.
Quantification of Cytokine Production by Luminex
The concentrations of cytokines in conditioned supernatants from the aforementioned DENV-infected primary monocyte cultures were assessed in duplicate using a multiplex cytokine assay by Luminex per the manufacturer's instructions.
NK Cell Degranulation Assay
Monocytes were infected with DENV-2 at an MOI of 2 and incubated for 24 h. After incubation, monocytes were left unblocked, blocked with an anti-pan human HLA class I Fab (generated from DX17, BD Bioscience), with an anti-human HLA-E Fab (generated from 3D12, BioLegend), or with isotype-matched control mouse IgG1 Fab (generated from MG1-45, BioLegend) all at 7.3 μg/mL for 30 min before adding autologous, IL-2-activated NK cells at a 1:5 effector to target (E:T) ratio. Fabs were produced using mouse IgG1 Fab F(ab)2 Kits (Thermo Scientific) and verified by gel electrophoresis and Coomassie Blue staining. During the 4 h co-culture, cells were incubated with brefeldin A (eBioscience), monensin (eBioscience), and APC-H7-conjugated anti-CD107a (H4A3, BD Bioscience) per manufacturer's instructions. Cells were stained with PerCP-Cy5.5-conjugated anti-CD3, FITC-conjugated anti-CD7, Alexa Fluor 700-conjugated anti-CD16 (3G8, BioLegend), PE-Cy7-conjugated anti-CD56 (HCD56, BioLegend), and Zombie Aqua Fixable Viability dye, then analyzed using a CytekTM Aurora analyzer and FlowJo® 10.2.
Statistical Analysis
A Friedman test, followed by Dunn's multiple comparisons test was used to determine significant differences between paired data with three conditions. A paired Wilcoxon signed-rank test was used to determine significant differences between DENV– and DENV+ cells. A Friedman test with FDR correction followed by a one-tailed Wilcoxon matched-pairs signed-rank test with a holm correction was used to analyze the Luminex data. Fab blocking data was analyzed using a Friedman test followed by paired Wilcoxon signed-rank tests. All statistical analysis was done using GraphPad Prism 8, R version 3.4.2, R version 3.6.0, and the compare_means function in the open source ggpubr R package.
Results
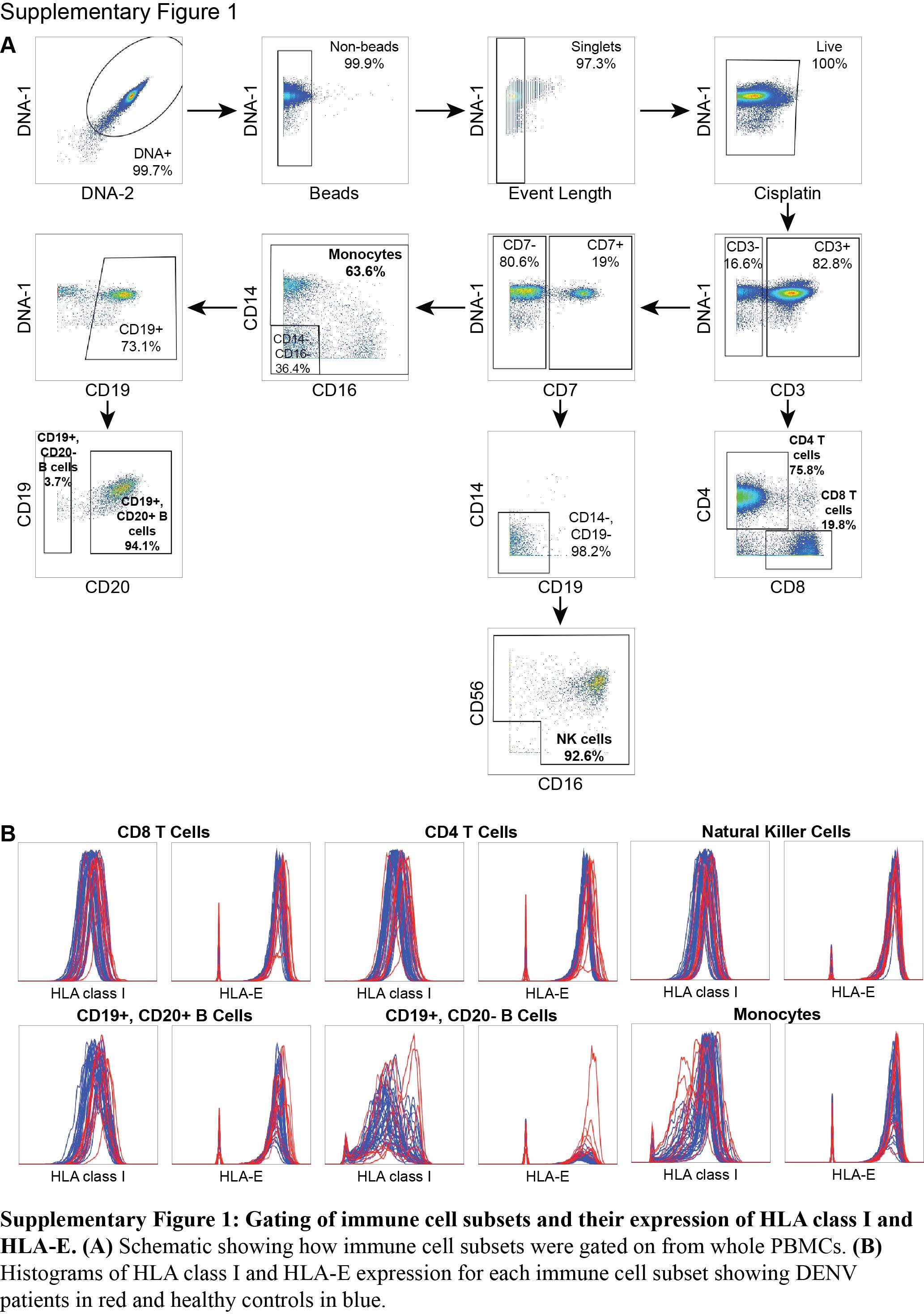
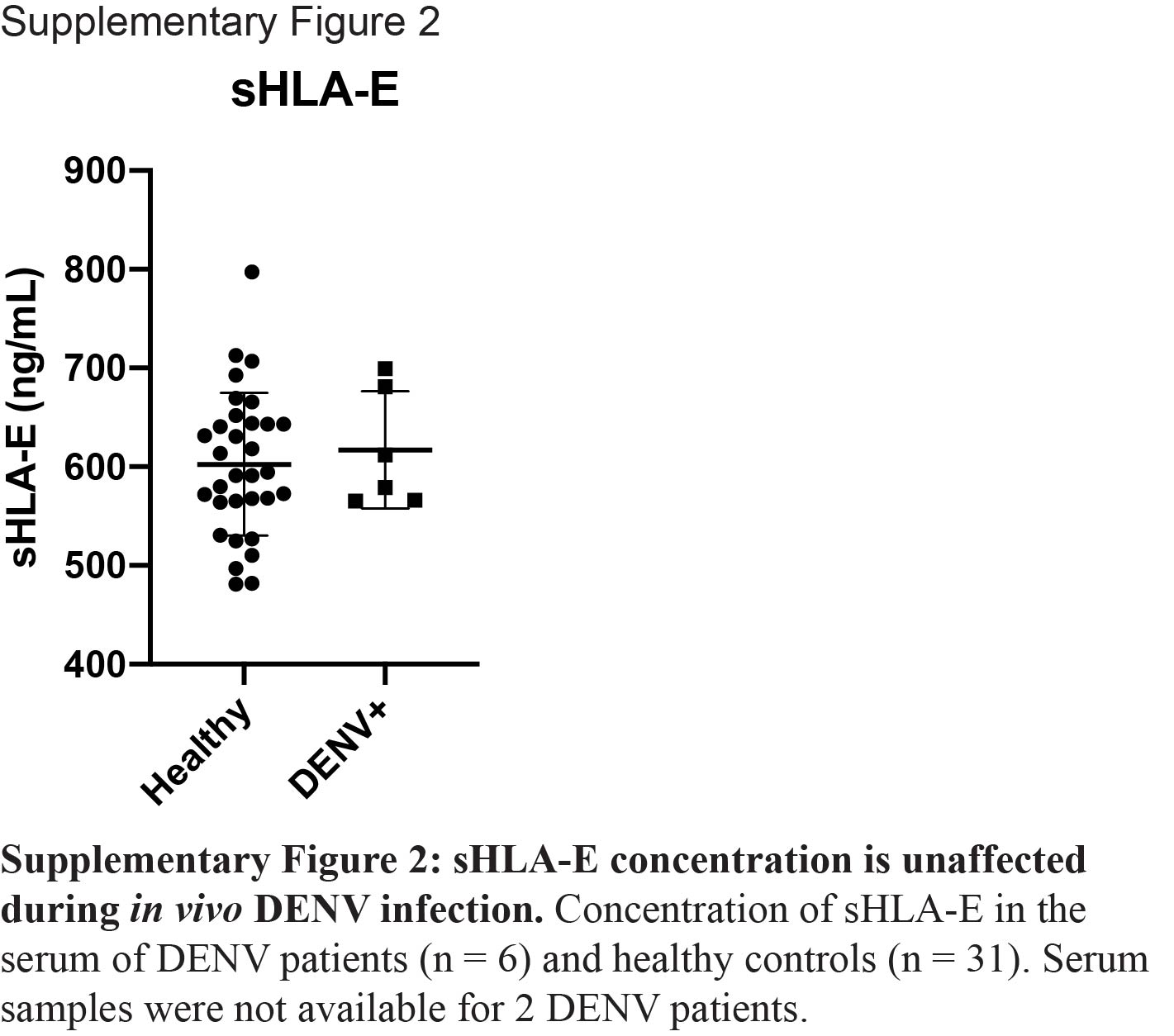
We evaluated HLA class I expression on PBMCs from a Panamanian cohort of 8 qRT-PCR confirmed, DENV-2-infected adults within 5 days of symptom onset and 31 healthy Panamanian adult controls (Supplementary Table 1). The expression patterns of HLA class I across cell subsets were visualized with viSNE. This algorithm separated immune cell subsets into clusters based on expression of key lineage markers; manual gating confirmed cluster identity (Figure 1A and Supplementary Figure 1A). Analysis based on protein expression revealed marked upregulation of total HLA class I and HLA-E in DENV-infected adults compared to healthy controls across multiple immune cell subsets (Figure 1B and Supplementary Figure 1B). As HLA-E can also be shed as soluble HLA-E (sHLA-E), which has been implicated as a potential viral mechanism of NK cell evasion (Shwetank et al., 2013, 2014), we performed an sHLA-E ELISA. There was no significant difference in the concentration of sHLA-E between the DENV-infected adults and healthy controls (Supplementary Figure 2). Together, these findings indicate that upregulation of HLA class I occurs on the cell surface of multiple immune cell subsets during acute in vivo DENV infection.
Figure 1.

HLA class I upregulation occurs during in vivo DENV infection. (A) Visualization of immune cell subsets in PBMCs from acute Panamanian DENV patients and healthy Panamanian controls using viSNE. The plots represent pooled data from n = 8 DENV patients and n = 31 controls. To assure equal donor representation, 4,375 events were used from each DENV patient and 1,129 events were used from each healthy control, resulting in 34,999 pooled events to generate both the DENV+ and healthy control viSNEs. Color key demonstrates major cell populations as determined by gating overlaid upon the viSNE visualization, demonstrating clusters of major cell subsets. (B) viSNE visualization of total HLA class I and HLA-E expression in whole PBMCs from DENV patients and healthy controls, generated as in (A). (C) Representative histogram from a DENV patient illustrating HLA class Ihigh and HLA class Ilow expressing monocytes gated on for generalized linear mixed model (GLMM) analysis. (D) GLMM analysis of markers associated with HLA class Ihigh and HLA class Ilow expressing monocytes. (E) Representative histogram from a DENV patient showing increased HLA class I expression by the monocyte subset gated on using the 10 markers identified in (D) (CD163, CD48, HLA-DR, CD95, LFA-3, HLA-E, ICAM-1, CCR2, Nectin-1, and CD14) compared to total monocytes from the same donor. (F) Summary data from all eight DENV patients. Wilcoxon signed-rank test **P < 0.01.
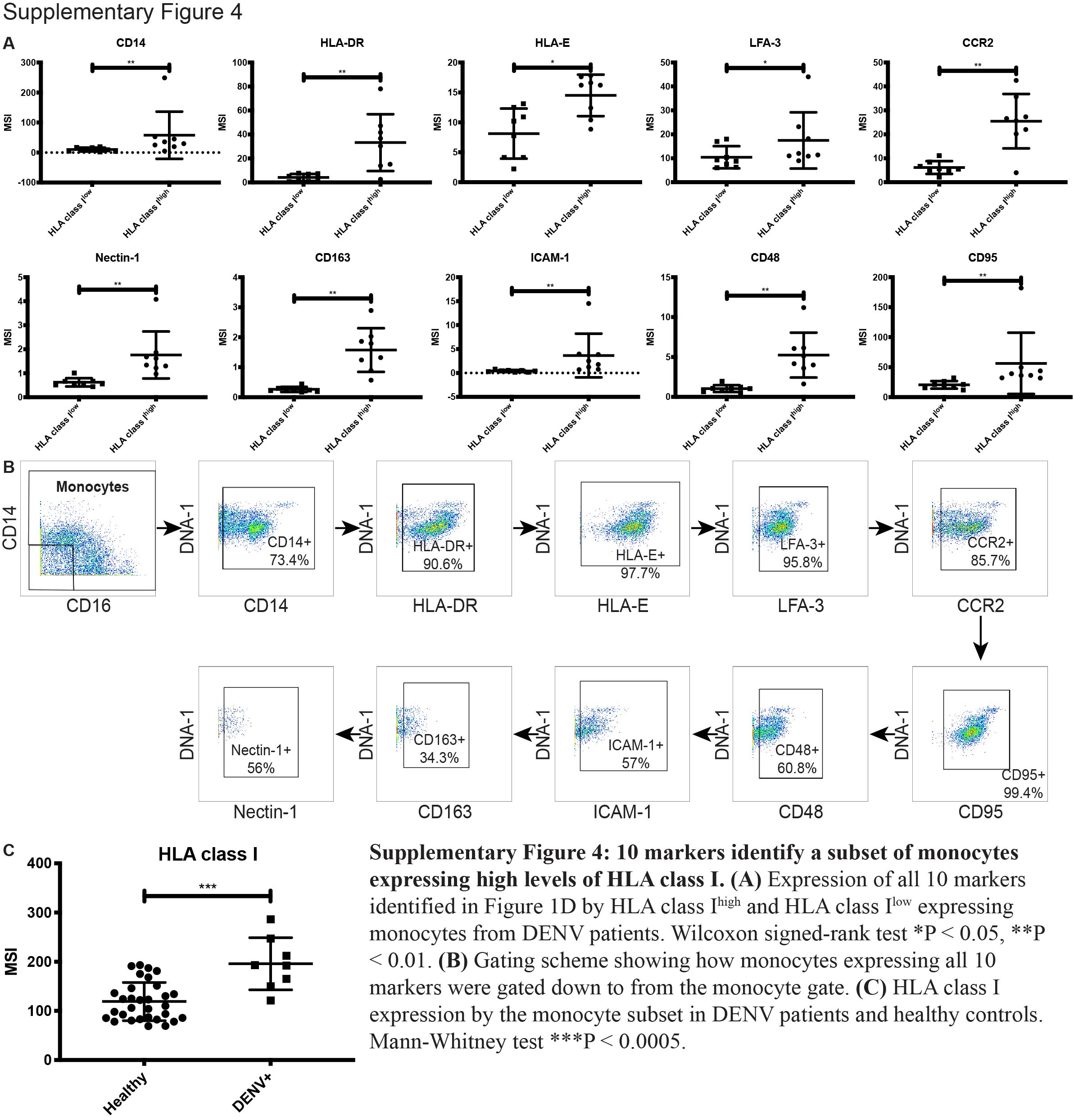
Interestingly, viSNE visualization revealed that upregulation of total HLA class I and HLA-E was not uniform across all monocytes. Instead, there were clear HLA class Ihigh and HLA-Ehigh expressing monocytes. We gated on these cells (Figure 1C and Supplementary Figure 3A) and used an unbiased generalized linear mixed model (GLMM) (Seiler et al., 2019) to identify associated markers. The GLMM identified 10 markers (CD14, HLA-DR, HLA-E, LFA-3, CCR2, CD95, CD48, ICAM-1, CD163, and Nectin-1) whose expression was associated with HLA class Ihigh monocytes (Figure 1D) and 5 markers (HLA class I, CD11b, ULBP-1,2,5,6, CD163, and MICA/B) whose expression was associated with HLA-Ehigh monocytes (Supplementary Figure 3B). We then verified these markers by comparing the expression level of each marker in HLAhigh expressing monocytes to the expression level in HLAlow expressing monocytes from DENV-infected adults (Supplementary Figures 3C, 4A). Finally, we gated down to monocytes expressing all 10 or all 5 markers in DENV-infected adults (Supplementary Figures 3D, 4B), and found that the 10 marker subset and the 5 marker subset expressed HLA class I and HLA-E, respectively, at significantly higher levels than total monocytes from the same donor (Figures 1E,F and Supplementary Figures 3E,F) verifying that combinatorial gating using the aforementioned markers accurately identifies HLA class Ihigh and HLA-Ehigh monocytes. Using the same gating scheme, we gated on the 10 and 5 marker monocyte subsets in healthy controls and found that HLA class I expression by the 10 marker subset and HLA-E expression by the 5 marker subset were both 1.6-fold higher in DENV-infected adults compared to healthy controls (Supplementary Figures 3G, 4C). These results indicate that upregulation of HLA class I and HLA-E by monocytes during in vivo DENV infection only occurs on specific monocyte subsets.
To better understand the effects of HLA class I upregulation, we modeled DENV infection in vitro using primary monocytes isolated from healthy donors. Further, to understand how viral replication vs. the presence of viral proteins alters HLA class I expression, we compared the effects of “active,” replication-competent DENV, with that of UV-inactivated virus incapable of viral replication. At 24 h post-infection (hpi), HLA class I expression did not significantly differ between active DENV, UV-inactivated DENV, or mock-infected conditions (Supplementary Figures 5A,B). By 48 hpi, HLA class I expression was 2.1-fold higher in monocytes infected with active DENV compared to mock (Figures 2A,B). Interestingly, in the active virus cultures, the uninfected bystander monocytes had a modest 1.1- and 1.2-fold higher HLA class I expression at 24 (Supplementary Figures 5C,D) and 48 (Figures 2C,D) hpi, respectively, than infected monocytes in the same culture. This suggests HLA class I upregulation is primarily restricted to infected cells with a modest impact on uninfected cells, likely due to changes in the cytokine milieu. Alternatively, bystander cells in our assay may have been infected at a level below our limit of detection.
Figure 2.

Primary monocytes upregulate HLA class I and HLA-E during in vitro DENV infection. Primary monocytes isolated from whole PBMCs from healthy blood bank donors were mock-infected (blue), exposed to UV-inactivated DENV (orange), or infected with active DENV (red) for 48 h. Representative histograms of HLA class I (A) and HLA-E (E) expression in total monocytes cultured in the respective conditions. Fluorescence minus one (FMO) shown in gray. HLA class I MFI in total monocytes (B) as well as bystander (DENV–) and infected (DENV+) monocytes (D). Representative histograms of HLA class I (C) and HLA-E (G) expression in bystander monocytes (DENV–, green) and infected monocytes (DENV+, purple). Percentage of HLA-Ehigh total monocytes (F) as well as bystander and infected monocytes (H). Two independent experiments measuring HLA class I were performed with 6 donors. The average for each donor is represented in the graphs. n = 12 for HLA-E experiments. Friedman test followed by Dunn's multiple comparisons test was used to analyze total monocytes. Wilcoxon signed-rank test was used to analyze DENV– vs. DENV+ monocytes. *P < 0.05, **P < 0.01, ***P < 0.001.
Expression of HLA-E was also altered during DENV infection. At 24 and 48 hpi, exposure to UV-inactivated DENV or infection with active DENV resulted in a majority of monocytes becoming HLA-Ehigh (Supplementary Figure 5E and Figure 2E). Similarly, at 48 hpi, the percentage of HLA-Ehigh monocytes in the UVed and active DENV cultures was 2.8- and 3-fold higher, respectively, compared to mock-infected monocytes (Figure 2F). The increase of HLA-Ehigh monocytes was 3-fold for both virus conditions compared to mock at 24 hpi (Supplementary Figure 5F). At both time points, bystander monocytes in the active DENV cultures had a bimodal distribution of HLA-Elow and HLA-Ehigh cells, while infected cells were all HLA-Ehigh (Figure 2G and Supplementary Figure 5G). Infected monocytes displayed a 1.3-fold increase in the percentage of HLA-Ehigh monocytes at 24 hpi compared to bystander monocytes (Supplementary Figure 5H), but at 48 hpi there was no longer a significant difference (Figure 2H). These results suggest that the response to viral proteins, rather than viral replication, is the main driver of HLA-E upregulation.
Considering that bystander monocytes expressed higher levels of HLA class I than DENV-infected monocytes, we wanted to test whether secreted factors, such as cytokines, viral proteins, and other molecules produced during active DENV infection were sufficient to upregulate HLA class I. To this end, we UV-treated conditioned supernatants from previous 48 h cultures of primary monocytes that were mock-infected, exposed to UV-inactivated DENV, or infected with active DENV. We then isolated uninfected monocytes from the same donors from which the supernatants were collected and cultured them in the conditioned supernatants for 24 h. Supernatants collected from active, DENV-infected cultures led to a significant, if modest, 1.1-fold increase in HLA class I expression compared to monocytes cultured in supernatants collected from the mock-infected and UV-inactivated DENV cultures (Figures 3A,B). This increase in HLA class I expression shows that soluble factors secreted during active DENV infection could be contributing to HLA class I upregulation. In order to investigate the potential role of cytokines in mediating this increase in HLA class I expression, we used Luminex to determine the concentration of cytokines present in the conditioned supernatants. We found that the concentrations of IFN-α, IFN-β, and TNF-α were 759.2-, 26-, and 271.7-fold higher, respectively, in the active condition compared to mock and 59.9-, 12.3-, and 2.8-fold higher, respectively, in the active condition compared to the UVed condition (Figure 3C). These findings suggest that upregulation of HLA class I during active DENV infection is largely driven by viral replication, and is likely mediated to some extent by cytokines. It is important to note that viral RNA, proteins, and particles present in the supernatant of active infection cultures could also contribute to HLA class I upregulation.
Figure 3.

Soluble factors secreted during active DENV infection upregulate HLA class I expression. Conditioned supernatants from experiments shown in Figure 2 were UV-treated and used to culture primary monocytes from healthy blood bank donors (n = 9). After a 24 h incubation, expression of total HLA class I was analyzed by flow cytometry. Histograms from a single representative donor (A) as well as summary data from all 9 donors (B) are shown. Friedman test followed by Dunn's multiple comparisons test, *P < 0.05, **P < 0.01. (C) Cytokine concentrations in conditioned supernatants from experiments shown in Figure 2 were analyzed by Luminex. Values shown are the average of two reads for each sample (n = 6). Friedman test with FDR correction followed by a one-tailed Wilcoxon matched-pairs signed-rank test with holm correction, *P < 0.05.
Given that NK cell activation is dampened by the expression of self-HLA class I on potential target cells, we investigated the impact of DENV-mediated HLA class I upregulation on NK cell degranulation in response to DENV-infected cells. We co-cultured DENV-infected primary monocytes with autologous primary NK cells in the presence of an isotype-matched control Fab, an anti-pan HLA class I blocking Fab, or an anti-HLA-E blocking Fab for 4 h and measured the percentage of CD107a+ NK cells as a marker of degranulation and killing activity (Figures 4A,B). Fabs were used instead of whole IgG to avoid killing via ADCC following binding of the anti-HLA antibodies to the target cells. Blocking HLA class I on mock-infected monocytes led to a 9.7% frequency of CD107a+ NK cells, a 4.5% increase from 5.2% in the unblocked, mock-infected monocyte condition. Because NK cells can become activated when they are unable to bind self-HLA class I, this result demonstrates that the Fabs were effectively blocking their targeted proteins. Blocking HLA class I on DENV-infected monocytes resulted in nearly double the frequency of CD107a+ NK cells compared to DENV-infected monocytes blocked with the isotype-matched control Fab, increasing from 9.9 to 18.8%. Additionally, HLA class I-blocking of DENV-infected monocytes resulted in a 6.7% increase in CD107a+ NK cells compared to blocking HLA-E. Blocking HLA class I on DENV-infected monocytes also nearly doubled the frequency of CD107a+ NK cells compared to blocking HLA class I on mock-infected cells, increasing from 9.7 to 18.8% (P = 0.0312). For all of the blocking conditions, except HLA-E, the DENV co-cultures had a statistically significant increase in CD107a+ NK cells compared to the mock-infected co-cultures. Thus, these data demonstrate that HLA class I upregulation can dampen the NK cell response to DENV-infected cells.
Figure 4.

Blocking HLA class I improves NK cell responses to DENV-infected cells. Primary NK cells and monocytes were isolated from whole PBMCs from healthy blood bank donors (n = 6). NK cells were activated for 22 h with IL-2. Monocytes were mock-infected (blue) or infected with active DENV (red) at an MOI of 2 for 24 h. Prior to co-culture with autologous NK cells, monocytes were blocked for 30 min with an isotype-matched control Fab, an anti-HLA-E blocking Fab, or an anti-pan HLA class I blocking Fab. Monocytes and NK cells were co-cultured for 4 h before NK cell expression of CD107a was evaluated by flow cytometry. Flow cytometry plots from a single representative donor (A) as well as summary data from all 6 donors (B) are shown. Friedman test followed by paired Wilcoxon signed-rank tests, *P < 0.05.
Discussion
Roughly one-third of the world's population is at risk of acquiring DENV, making it critically important that we elucidate immune factors that contribute both to disease protection and pathogenesis. Mechanisms by which DENV evades the innate immune response by inhibiting the production and signaling of type I IFNs, as well as other aspects of the cellular antiviral response, have been well-reported (Morrison et al., 2012; Green et al., 2014). Similarly, pathways that might promote DENV escape from NK cell recognition have been proposed, including a potential role for the upregulation of HLA class I molecules by DENV-infected cells to inhibit the NK cell response by binding inhibitory KIRs or CD94/NKG2A (Beltrán and López-Vergès, 2014; Petitdemange et al., 2014; Mathew, 2018). However, these data have primarily arisen from in vitro infection of mouse or human cell lines, rather than more physiologic systems such as natural infection or infection of undifferentiated primary human immune cells. Here, we show natural DENV infection leads to increased expression of HLA class I and HLA-E in adult patients. We also found that soluble factors produced during active infection contributed to HLA class I upregulation. Finally, blocking HLA class I on DENV-infected monocytes enhanced the ability of NK cells to degranulate in response to DENV-infected cells. Together, these findings show HLA class I upregulation during active DENV infection suppresses NK cell degranulation.
HLA class I upregulation during flavivirus infection has been previously described and attributed to various mechanisms such as NFκB activation (Kesson and King, 2001), increased transport of peptides into the endoplasmic reticulum for HLA loading (Momburg et al., 2001), and the presence of IFN-β (Glasner et al., 2017). We show replication-competent virus was required for significant HLA class I upregulation, but that HLA class I expression was highest in uninfected bystander monocytes. Further, supernatants collected from active DENV cultures were able to upregulate HLA class I on uninfected monocytes and contained higher concentrations of IFN-α, IFN-β, and TNF-α compared to supernatants from mock-infected or UV-inactivated DENV cultures indicating HLA class I upregulation is mediated at least in part by soluble factors. These findings pose a potential mechanism for HLA class I upregulation in which soluble factors secreted by DENV-infected cells induce increased HLA class I expression on all cells in an effort to promote cytotoxic T lymphocyte responses. However, the DENV-infected cells express lower levels than the bystander cells because DENV may encode proteins that interfere with processes driving HLA class I upregulation to escape the T cell response (Ye et al., 2013; Green et al., 2014; Guzman and Harris, 2015; Glasner et al., 2017).
Interestingly, upregulation of HLA-E likely involves different mechanisms than upregulation of other HLA class I molecules. We observed a significant increase in HLA-E expression in response to active DENV as well as UV-inactivated DENV. This suggests that direct sensing of viral products by innate immune receptors and the resulting cytokines, rather than pathways induced during viral replication, may be the primary contributors to HLA-E upregulation. However, DENV-infected monocytes expressed the highest levels of HLA-E, implying DENV itself is also mediating HLA-E upregulation. Intriguingly, cytomegalovirus proteins can modulate the surface expression of HLA-E by encoding HLA leader sequence mimics with reduced binding affinity to the CD94/NKG2 receptors (Heatley et al., 2013). Similarly, human immunodeficiency virus-1 has been found to upregulate HLA-E expression, resulting in the presentation of a capsid peptide which prevents HLA-E engagement with CD94/NKG2A (Nattermann et al., 2005; Davis et al., 2016). No such mechanisms for modulating HLA-E expression and its affinity for the CD94/NKG2 receptors have been reported for DENV. The NetMHCpan 4.0 server predicts several DENV-2 peptides with strong and weak binding to HLA-E*01:01, making it possible that DENV peptides modulate NKG2A/C binding. Together, these results suggest HLA-E upregulation is mediated by both virus-dependent and virus-independent mechanisms, and could influence NK cell recognition through NKG2A/C.
Here we extend prior studies demonstrating that HLA class I upregulation during flavivirus infection in cell lines can inhibit NK cell activation (Lobigs et al., 1996; Momburg et al., 2001; Hershkovitz et al., 2008; Glasner et al., 2017; Drews et al., 2018). For the first time, we used patient samples and more physiologically relevant in vitro infection and co-culture systems with undifferentiated primary human immune cells. By directly blocking NK cell binding to HLA class I using Fabs, we found HLA class I expression on DENV-infected cells significantly dampens NK cell degranulation. This provides the first direct evidence that upregulation of HLA class I is responsible for inhibition of NK cell responses to flavivirus-infected cells. We did observe some non-specific increase in NK cell degranulation in the presence of the isotype-matched control antibody, but this effect was dwarfed by the increase in NK cell degranulation in response to DENV-infected cells in the presence of HLA class I blocking Fabs. These findings indicate that HLA class I expression dampens the magnitude of the NK cell response.
Previous studies using cytokine-activated endothelial cells showed that blocking surface HLA-E and sHLA-E increased NK cell killing, illustrating the importance of HLA-E expression as a potential NK escape mechanism in some vascular diseases (Coupel et al., 2007). However, similar to Drews et al. we did not observe a significant modulating effect of HLA-E expression on NK cell activity against DENV-infected cells (Drews et al., 2018). HLA-E binds to both inhibitory NK cell receptor CD94/NKG2A and activating receptor CD94/NKG2C (Braud et al., 1998; Valés-Gómez et al., 1999; Kaiser et al., 2005). Notably, HLA-E binds to CD94/NKG2A with higher affinity (Valés-Gómez et al., 1999; Kaiser et al., 2005). The fact that NK cell binding to HLA-E can result in both activating and inhibitory signaling with a dominant advantage toward inhibitory signaling could explain our results and those of Drews et al. HLA-E's greater affinity toward CD94/NKG2A also suggests that increased HLA-E expression on DENV-infected cells might be part of the viral escape strategy. Specifically blocking NKG2A or NKG2C in NK cell-infected cell co-cultures could identify the role of inhibitory signaling through NKG2A vs. activating signaling through NKG2C in DENV recognition.
In contrast to Drews et al. and Shwetank et al. who observed an increase in sHLA-E in the supernatants of DENV-infected HMEC-1 cells and Japanese encephalitis virus-infected human brain microvascular endothelial cells, respectively, we saw no significant increase in sHLA-E in the serum of DENV patients compared to healthy controls (Shwetank et al., 2013; Drews et al., 2018). This suggests that sHLA-E shedding is not increased at the systemic level during in vivo DENV infection and is consequently unlikely to contribute strongly to suppression of the NK cell response.
This study has limitations, the most significant of which is the small sample size of DENV-infected adults in our cohort. Despite the modest numbers, the conclusions we drew from these in vivo data were clear and supported by our in vitro experiments using primary human immune cells. The second limitation is that we were unable to clearly determine the role of HLA-E on the NK cell response to DENV infection given its binding to both inhibitory CD94/NKG2A and activating CD94/NKG2C receptors.
To our knowledge, ours is the first study showing upregulation of HLA class I molecules in acute dengue patient samples, suggesting different drivers of HLA-E upregulation vs. upregulation of other HLA class I proteins during DENV infection, and showing enhanced primary NK cell degranulation upon blocking HLA class I on primary DENV-infected monocytes. Our in vivo HLA class I expression data need to be confirmed with additional DENV cohorts at different time points of the disease, spanning all serotypes and degrees of disease severity, and including pediatric patients. This will be vital to determining what temporal, viral, and age-related factors affect HLA class I upregulation, as well as whether there is a correlation between disease severity and HLA class I expression. Future experiments are also required to determine what factors and pathways mediate HLA class I upregulation in monocytes and how HLA class I expression is modulated in other immune cell subsets. Overall, this study furthers our understanding of the impacts of DENV infection on innate immune cells and their intercellular interactions.
Data Availability
Requests to access the dataset should be directed to Catherine Blish at cblish@stanford.edu.
Ethics Statement
The studies involving human participants were reviewed and approved by the IRB of Hospital del Niño (CBIHN-M-0634), then confirmed by the committees of ICGES, CSS, Santo Tomas Hospital, and Stanford University. Written informed consent to participate in this study was provided by the participants or participants' legal guardian/next of kin.
Author Contributions
JM, DB, CB, and SL-V contributed to the conceptualization, formal analysis, investigation, data curation, and the preparation of the writing of the original draft. JM, DB, AP, LS, AA, and RV contributed to the methodology. DB, EH, LL, CB, and SL-V contributed to the resources. EH and LL contributed to the reviewing and editing of the manuscript. CB and SL-V contributed to the supervision and project administration. DB, CB, and SL-V contributed to the funding acquisition.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank all health institutions, patients, and their families, Luis Bonilla from the Blood Bank of Santo Tomas Hospital for providing blood components, and the Stanford Human Immune Monitoring Center. We thank Dr. Taia Wang for critical reading of the manuscript and Dr. Anne-Maud Ferreira for statistical analysis of the Luminex data.
Footnotes
Funding. This work was supported by NIH R21AI135287 and R21AI130523 to CB, grants 9044.51 from the Ministry of Economy and Finance of Panama and 71-2012-4-CAP11-003 from SENACYT to SL-V, National Science Foundation Graduate Research Fellowship DGE-1656518 to JM and NIH training grant T31-AI-07290 (PI Olivia Martinez). DB (Ph.D. student) and SL-V are members of the Sistema Nacional de Investigación (SNI) of SENACYT, Panama. CB is the Tashia and John Morgridge Faculty Scholar in Pediatric Translational Medicine from the Stanford Maternal Child Health Research Institute and an Investigator of the Chan Zuckerberg Biohub.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2019.00268/full#supplementary-material
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
- Azeredo E. L. (2006). NK cells, displaying early activation, cytotoxicity and adhesion molecules, are associated with mild dengue disease. 143, 345–356. 10.1111/j.1365-2249.2006.02996.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayless N. L., Greenberg R. S., Swigut T., Wysocka J., Blish C. A. (2016). Zika virus infection induces cranial neural crest cells to produce cytokines at levels detrimental for neurogenesis. Cell Host Microbe 20, 423–428. 10.1016/j.chom.2016.09.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beltrán D., López-Vergès S. (2014). NK cells during dengue disease and their recognition of Dengue virus-infected cells. Front. Immunol. 5:192. 10.3389/fimmu.2014.00192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhatt S., Gething P. W., Brady O. J., Messina J. P., Farlow A. W., Moyes C. L., et al. (2013). The global distribution and burden of dengue. Nature 496, 504–507. 10.1038/nature12060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braud V. M., Allan D. S., O'Callaghan C. A., Söderström K., D'Andrea A., Ogg G. S., et al. (1998). HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 391, 795–799. 10.1038/35869 [DOI] [PubMed] [Google Scholar]
- Cheng Y., King N. J. C., Kesson A. M. (2004). Major histocompatibility complex class I (MHC-I) induction by West Nile virus: involvement of 2 signaling pathways in MHC-I up-regulation. J. Infect. Dis. 189, 658–668. 10.1086/381501 [DOI] [PubMed] [Google Scholar]
- Costa V. V., Ye W., Chen Q., Teixeira M. M., Preiser P., Ooi E. E., et al. (2017). Dengue virus-infected dendritic cells, but not monocytes, activate natural killer cells through a contact-dependent mechanism involving adhesion molecules. MBio 8:e00741–17. 10.1128/mBio.00741-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coupel S., Moreau A., Hamidou M., Horejsi V., Soulillou J.-P., Charreau B. (2007). Expression and release of soluble HLA-E is an immunoregulatory feature of endothelial cell activation. Blood 109, 2806–2814. 10.1182/blood-2006-06-030213 [DOI] [PubMed] [Google Scholar]
- Davis Z. B., Cogswell A., Scott H., Mertsching A., Boucau J., Wambua D., et al. (2016). A conserved HIV-1-derived peptide presented by HLA-E renders infected T-cells highly susceptible to attack by NKG2A/CD94-bearing natural killer cells. PLoS Pathog. 12:e1005421. 10.1371/journal.ppat.1005421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drews E., Adam A., Htoo P., Townsley E., Mathew A. (2018). Upregulation of HLA-E by dengue and not Zika viruses. Clin. Transl. Immunol. 7:e1039 10.1002/cti2.1039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durbin A. P., Vargas M. J., Wanionek K., Hammond S. N., Gordon A., Rocha C., et al. (2008). Phenotyping of peripheral blood mononuclear cells during acute dengue illness demonstrates infection and increased activation of monocytes in severe cases compared to classic dengue fever. Virology 376, 429–435. 10.1016/j.virol.2008.03.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glasner A., Oiknine-Djian E., Weisblum Y., Diab M., Panet A., Wolf D. G., et al. (2017). Zika virus escapes NK cell detection by upregulating major histocompatibility complex class I molecules. J. Virol. 91:e00785-17. 10.1128/JVI.00785-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green A. M., Beatty P. R., Hadjilaou A., Harris E. (2014). Innate immunity to Dengue virus infection and subversion of antiviral responses. J. Mol. Biol. 426, 1148–1160. 10.1016/j.jmb.2013.11.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guzman M. G., Harris E. (2015). Dengue. Lancet 385, 453–465. 10.1016/S0140-6736(14)60572-9 [DOI] [PubMed] [Google Scholar]
- Heatley S. L., Pietra G., Lin J., Widjaja J. M. L., Harpur C. M., Lester S., et al. (2013). Polymorphism in human cytomegalovirus UL40 impacts on recognition of human leukocyte antigen-E (HLA-E) by natural killer cells. J. Biol. Chem. 288, 8679–8690. 10.1074/jbc.M112.409672 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hershkovitz O., Zilka A., Bar-Ilan A., Abutbul S., Davidson A., Mazzon M., et al. (2008). Dengue virus replicon expressing the nonstructural proteins suffices to enhance membrane expression of HLA class I and inhibit lysis by human NK cells. J. Virol. 82, 7666–7676. 10.1128/JVI.02274-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaiser B. K., Barahmand-Pour F., Paulsene W., Medley S., Geraghty D. E., Strong R. K. (2005). Interactions between NKG2x immunoreceptors and HLA-E ligands display overlapping affinities and thermodynamics. J. Immunol. 174, 2878–2884. 10.4049/jimmunol.174.5.2878 [DOI] [PubMed] [Google Scholar]
- Kesson A. M., King N. J. C. (2001). Transcriptional regulation of major histocompatibility complex class I by flavivirus West Nile is dependent on NF-kappa B activation. J. Infect. Dis. 184, 947–954. 10.1086/323603 [DOI] [PubMed] [Google Scholar]
- Kyle J. L., Harris E. (2008). Global spread and persistence of dengue. Annu. Rev. Microbiol. 62, 71–92. 10.1146/annurev.micro.62.081307.163005 [DOI] [PubMed] [Google Scholar]
- Laoprasopwattana K., Libraty D. H., Endy T. P., Nisalak A., Chunsuttiwat S., Ennis F. A., et al. (2007). Antibody-dependent cellular cytotoxity mediated by plasma obtained before secondary Dengue virus infections: potential involvement in early control of viral replication. J. Infect. Dis. 195, 1108–1116. 10.1086/512860 [DOI] [PubMed] [Google Scholar]
- Libraty D. H., Pichyangkul S., Ajariyakhajorn C., Endy T. P., Ennis F. A. (2001). Human dendritic cells are activated by Dengue virus infection: enhancement by gamma interferon and implications for disease pathogenesis. J. Virol. 75, 3501–3508. 10.1128/JVI.75.8.3501-3508.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lobigs M., Blanden R. V., Müllbacher A. (1996). Flavivirus-induced up-regulation of MHC class I antigens; implications for the induction of CD8+ T-cell-mediated autoimmunity. Immunol. Rev. 152, 5–19. 10.1111/j.1600-065X.1996.tb00908.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathew A. (2018). Defining the role of NK cells during Dengue virus infection. Immunology 154, 557–562. 10.1111/imm.12928 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mei H. E., Leipold M. D., Schulz A. R., Chester C., Maecker H. T. (2015). Barcoding of live human peripheral blood mononuclear cells for multiplexed mass cytometry. J. Immunol. 194, 2022–2031. 10.4049/jimmunol.1402661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Momburg F., Müllbacher A., Lobigs M. (2001). Modulation of Transporter associated with Antigen Processing (TAP) -mediated peptide import into the endoplasmic reticulum by flavivirus infection. J. Virol. 75, 5663–5671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison J., Aguirre S., Fernandez-Sesma A. (2012). Innate immunity evasion by Dengue virus. Viruses 4, 397–413. 10.3390/v4030397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nattermann J., Nischalke H. D., Hofmeister V., Kupfer B., Ahlenstiel G., Feldmann G., et al. (2005). HIV-1 infection leads to increased HLA-E expression resulting in impaired function of natural killer cells. Antivir. Ther. 10, 95–107. [DOI] [PubMed] [Google Scholar]
- Nightingale Z. D., Patkar C., Rothman A. L. (2008). Viral replication and paracrine effects result in distinct, functional responses of dendritic cells following infection with dengue 2 virus. J. Leukoc. Biol. 84, 1028–1038. 10.1189/jlb.0208105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petitdemange C., Wauquier N., Devilliers H., Yssel H., Mombo I., Caron M., et al. (2016). Longitudinal analysis of natural killer cells in Dengue virus-infected patients in comparison to chikungunya and chikungunya/Dengue virus-infected patients. PLoS Negl. Trop. Dis. 10:e0004499. 10.1371/journal.pntd.0004499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petitdemange C., Wauquier N., Rey J., Hervier B., Leroy E., Vieillard V. (2014). Control of acute Dengue virus infection by natural killer cells. Front. Immunol. 5:209. 10.3389/fimmu.2014.00209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seiler C., Kronstad L. M., Simpson L. J., Le Gars M., Vendrame E., Blish C. A., et al. (2019). Uncertainty Quantification in Multivariate Mixed Models for Mass Cytometry Data. arXiv [stat.AP]. Available online at: http://arxiv.org/abs/1903.07976.
- Shwetank, Date O. S., Carbone E., Manjunath R. (2014). Inhibition of ERK and proliferation in NK cell lines by soluble HLA-E released from Japanese encephalitis virus infected cells. Immunol. Lett. 162, 94–100. 10.1016/j.imlet.2014.07.010 [DOI] [PubMed] [Google Scholar]
- Shwetank, Date O. S., Kim K. S., Manjunath R. (2013). Infection of human endothelial cells by Japanese encephalitis virus: increased expression and release of soluble HLA-E. PLoS ONE 8:e79197. 10.1371/journal.pone.0079197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun P., Morrison B. J., Beckett C. G., Liang Z., Nagabhushana N., Li A., et al. (2017). NK cell degranulation as a marker for measuring antibody-dependent cytotoxicity in neutralizing and non-neutralizing human sera from dengue patients. J. Immunol. Methods 441, 24–30. 10.1016/j.jim.2016.11.005 [DOI] [PubMed] [Google Scholar]
- Sun P., Williams M., Nagabhushana N., Jani V., Defang G., Morrison B. J. (2019). NK cells activated through antibody-dependent cell cytotoxicity and armed with degranulation/IFN-γ production suppress antibody-dependent enhancement of dengue viral infection. Sci. Rep. 9:1109. 10.1038/s41598-018-36972-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valés-Gómez M., Reyburn H. T., Erskine R. A., López-Botet M., Strominger J. L. (1999). Kinetics and peptide dependency of the binding of the inhibitory NK receptor CD94/NKG2-A and the activating receptor CD94/NKG2-C to HLA-E. EMBO J. 18, 4250–4260. 10.1093/emboj/18.15.4250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- World Health Organization (2012). Global Strategy for Dengue Prevention and Control 2012-2020. Available online at: https://apps.who.int/iris/bitstream/handle/10665/75303/9789241504034_eng.pdf
- Ye J., Zhu B., Fu Z. F., Chen H., Cao S. (2013). Immune evasion strategies of flaviviruses. Vaccine 31, 461–471. 10.1016/j.vaccine.2012.11.015 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Requests to access the dataset should be directed to Catherine Blish at cblish@stanford.edu.