

Abstract
Defensive behaviors, such as withdrawing your hand to avoid potentially harmful approaching objects, rely on rapid sensorimotor transformations between visual and motor coordinates. We examined the reference frame for coding visual information about objects approaching the hand during motor preparation. Subjects performed a simple visuomanual task while a task-irrelevant distractor ball rapidly approached a location either near to or far from their hand. After the distractor ball appearance, single pulses of transcranial magnetic stimulation were delivered over the subject's primary motor cortex, eliciting motor evoked potentials (MEPs) in their responding hand. MEP amplitude was reduced when the ball approached near the responding hand, both when the hand was on the left and the right of the midline. Strikingly, this suppression occurred very early, at 70–80 ms after ball appearance, and was not modified by visual fixation location. Furthermore, it was selective for approaching balls, since static visual distractors did not modulate MEP amplitude. Together with additional behavioral measurements, we provide converging evidence for automatic hand-centered coding of visual space in the human brain.
Introduction
In daily life, sensorimotor integration is required for the execution of a wide range of movements, each of which poses different computational challenges (Rizzolatti et al., 1998; Graziano, 2006). Consider the ongoing processes of movement and response selection while you wash the dishes: although you might still attempt to catch a slippery glass as it is falling, at some point you must withdraw your hand to avoid injury.
The majority of human research on coordinate transformations for hand movements has studied goal-directed reaching and grasping (Crawford et al., 2004; Medendorp et al., 2005; Beurze et al., 2006; Fernandez-Ruiz et al., 2007; Brozzoli et al., 2009), whereas the reference frames in which avoidance movements are coded have been relatively neglected. However, electrophysiological experiments in macaques in which three-dimensional stimuli were moved rapidly toward the monkey's body have revealed body part-centered representations of visual space (Graziano et al., 2004). For example, certain neurons in the ventral and dorsal premotor cortex represent objects in hand-centered coordinates (Graziano et al., 1994, 1997; Fogassi et al., 1996, 1999; Graziano, 1999) and may be involved in the preparation and guidance of movements both toward and away from objects (as suggested by Rizzolatti, 1987; Maravita et al., 2003; Cooke and Graziano, 2004; Avillac et al., 2005). Neuroimaging and neuropsychological studies have suggested the involvement of possibly homologous structures in the hand-centered coding of space in the human brain (di Pellegrino et al., 1997; Bremmer et al., 2001; Farnè et al., 2003, 2005; Makin et al., 2007; Quinlan and Culham, 2007) (for review, see Makin et al., 2008).
In the current study, we used an active motor task to determine the reference frame for coding the location of real, three-dimensional objects approaching the hand. Subjects abducted their right index finger in response to a visual “go” signal and were simultaneously presented with a task-irrelevant distractor ball, rapidly approaching a location either near to or far from their responding hand.
To obtain an index of corticospinal excitability with which to assess the effects of rapidly approaching distractor stimuli, we used single pulse transcranial magnetic stimulation (TMS), applied to the left primary motor cortex, eliciting motor-evoked potentials (MEPs) in the subject's right hand. We hypothesized that the hand-centered coding of approaching visual stimuli should have, as one consequence, some modulatory effect on corticospinal excitability as a function of the distance of the approaching object from the hand (near vs far). We report that corticospinal excitability is indeed modulated in a hand-centered manner, while subjects are preparing to respond to the targets.
In a set of complementary experiments, we examined the relative contributions of visual fixation position and distractor type [approaching ball vs static light-emitting diode (LED)] to this hand-centered coding of space. We further examined the process of “proactive inhibition,” the ability to inhibit in advance one movement to execute another (Koch et al., 2006; Mars et al., 2007; van den Hurk et al., 2007; van Elswijk et al., 2007), in relation to the suppression of possible avoidance responses elicited by the approaching ball.
Materials and Methods
Subjects
A total of 23 healthy right-handed subjects participated in seven experiments (supplemental Table S1, available at www.jneurosci.org as supplemental material). The subjects gave written, informed consent and were screened for contraindications to TMS. The experiments were conducted in accordance with the Declaration of Helsinki and were approved by the local ethical review board.
Data acquisition and apparatus
Physiological measurements.
MEPs were recorded from the right first dorsal interosseus (FDI) muscle using three self-adhesive electrodes (Neuroline; Ambu) in a belly-to-tendon manner. EMG data were acquired using a BioPac system (BioPac Systems). EMG signals were sampled for 2 s at 2 kHz, amplified 1000 times, and bandpass filtered (25–250 Hz). At the end of each trial, trigger pulses were sent to allow automated identification of trials. Data were stored for off-line analysis (see below).
Transcranial magnetic stimulation.
A Magstim Super Rapid stimulator and figure-of-eight, 70-mm-diameter TMS coil were used. The position and orientation of the TMS coil and the intensity of the single pulses was optimized for each subject to elicit reliable MEPs in the FDI, following standard procedures (Hallett, 2007). Resting motor threshold (RMT) was defined as the minimum TMS intensity (in percentage of maximum stimulator output) required to elicit only 5 of 10 MEPs with peak-to-peak amplitudes equal to or above 0.5 mV, while the subject's right hand was relaxed and placed on the right side of the workspace. We set this (relatively) high amplitude criterion for the RMT and stimulated at 110% of this RMT to elicit MEPs on virtually every trial and to allow detection of MEP suppression (Cantello et al., 2000) (supplemental Table S1, available at www.jneurosci.org as supplemental material).
Visual stimuli.
Two types of visual stimuli were used as distractors: balls and LEDs. The balls were 5-cm-diameter red foam spheres, mounted on the end of 55-cm-long aluminum rods (stimulus arms). The LEDs were 5 mm in diameter, green for the central go signal, and red for peripheral distractors/fixations and the central “stop” LED.
A custom apparatus controlled stimulus delivery (see Fig. 1A). Two stimulus arms were mounted on rotating wheels positioned opposite the subject and occluded from the subject's sight. After a cue, the experimenter released one of the stimulus arms from its vertical position, and the ball fell under gravity toward the workspace, entering the subject's field of view (see supplemental Videos 1, 2, available at www.jneurosci.org as supplemental material). The ball traversed ∼37 cm in ∼100 ms after becoming visible to the subject, having a mean velocity when visible of ∼370 cm/s, and stopped with minimal bouncing ∼3 cm above the workspace. The landing position of the ball was either directly above the middle finger of the subject's hand (near trials) or 30 cm laterally to the hand (far trials).
Figure 1.
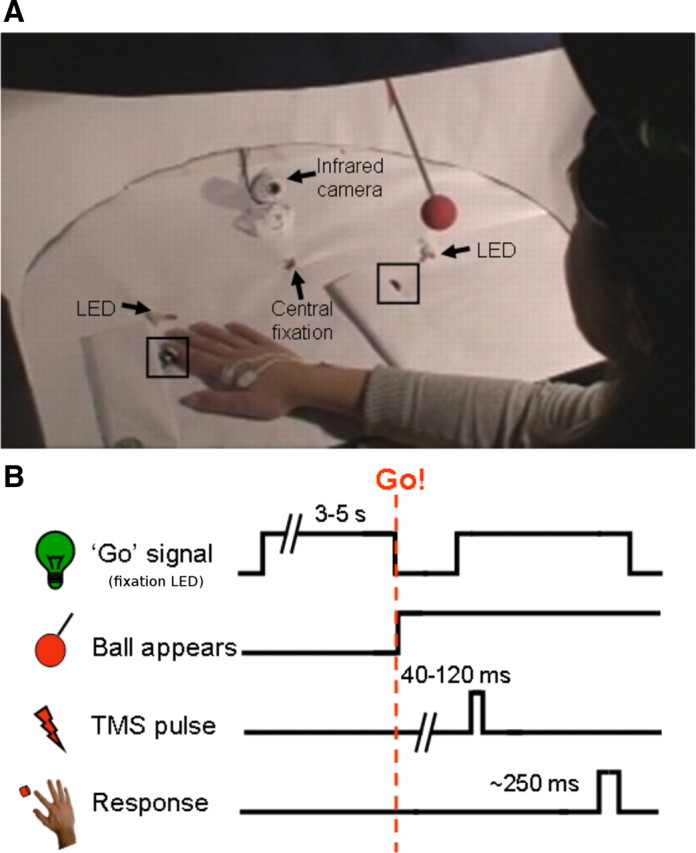
General experimental setup and design. A, Experimental setup. B, Time course. Subjects positioned their right hand 17.5 cm to the left (shown in A) or right of the central fixation LED. They were instructed to respond to a transient (130 ms) offset of the fixation LED by abducting their right index finger to push a button (black square) while ignoring the distractor ball (red sphere). The distractor ball appeared simultaneously with the go signal, approaching a position pseudorandomly either on the left or right (shown in A) of fixation. Between 40 and 120 ms after the appearance of the distractor ball, a single pulse of TMS was delivered to the hand area of the left hemisphere primary motor cortex, eliciting a MEP in the right FDI muscle.
During their trajectories, the stimulus arms interrupted the beams of two lasers, triggering pulses from two detectors mounted on the apparatus. The second laser was positioned to intersect the stimulus arm at the point at which the distractor balls first became visible to the subject, thus providing a precise timing signal for distractor ball appearance. The first laser was positioned close to the start of the movement of the stimulus arms and was used in experiments in which a stop signal or peripheral visual cue was provided in advance of distractor ball appearance. The time taken for the stimulus arms to pass between the two laser beams was approximately constant, with a trial-to-trial timing range of <5 ms.
Additional apparatus.
Two response buttons were firmly attached, with the button facing rightward, 20 cm to the left and 15 cm to the right of the visual midline, so that the middle finger of the subject's hand in both positions was ∼17.5 cm from the midline. For experiments 2, 3, 4, 6, and S1, a small infrared camera was mounted 5 cm behind the central fixation LED, allowing the experimenter to monitor the subject's eye position. A large ventilator was placed under the workspace near the subject, providing constant background noise and airflow, minimizing any acoustic and airflow artifacts induced by stimulus delivery. In pilot experiments, blindfolded subjects were unable to determine the position of balls with respect to their hand.
Design and procedure
Because the seven experiments differed in several respects from each other, the general design and procedures are described here. For additional details, see Table 1, supplemental material (available at www.jneurosci.org), and the individual section for each experiment in Results.
Table 1.
Experimental conditions
| Experiment | N | Blocks | Trials perblock | Trial proportion (%) |
TMS time point(s) | Comments | ||
|---|---|---|---|---|---|---|---|---|
| Hand position(L/R) | Distractor position(L/R/none) | Task(go/stop/stay) | ||||||
| (1) Time course | 10 | 4 | 50 | 50/50 | 50/50/0 | 100/0/0 | 40, 60, 80, 100, 120 | — |
| (2) Eye position | 10 | 2 | 60 | 50/50 | 40/40/20 | 100/0/0 | 70, 80 (75)a | Two fixation positions (L/R); eye monitoring |
| (3) Go/stop balls | 11 (10b) | 4 | 72 | 50/50 | 33/33/33 | 75/25/0 | 70, 80, 90, 100, 110, 120; (80, 110)c | Eye monitoring |
| (4) Go/stop LEDs | 10 (9b) | 4 | 72 | 50/50 | 33/33/33 | 75/25/0 | 70, 80, 90, 100, 110, 120; (80, 110)c | Eye monitoring |
| (5) Go/stay balls/LEDs | 10 | 4 | 48 | 50/50 | 25/25/0d | 50/0/50 | 70 | Behavioral data analyzed |
| (6) Selectivity | 6 | 4 | 60 | 50/50 | 50/50/0 | 100/0/0 | 65, 70, 75, 80, 85 | MEPs from FDI and ADM; eye monitoring |
| (S1) Exog. attention | 10 | 2 | 60 | 50/50 | 50/50/0 | 75/0/25 | 70, 80, 75e | Run with experiment 2; eye monitoring |
N, Sample size; L, left; R, right.
aNo-distractor trials.
bOne dataset from experiment 3 discarded because of insufficient valid trials; one dataset from experiment 4 lost because of hardware failure.
cStop trials.
dPer distractor type (balls, LEDs).
eStay trials.
The two principal variables in every experiment were the position of the right hand (on the left or right of the workspace, in separate, counterbalanced blocks), and the position of the distractor stimulus (left or right of the workspace, pseudorandomized trial-by-trial). Additional variables for each experiment are detailed in Table 1. Subjects performed a short (10–20 trials) practice session before each experiment.
Trials began with the illumination of the central fixation LED, on which subjects fixated for the duration of the trial. To maintain subjects' attention during stimulation and to ensure that hand movement was task-relevant, subjects performed a speeded abduction of their right index finger, pushing a button in response to the target, which was a brief (130 ms) offset of the fixation LED. In trials with a distractor ball, the offset of the fixation was simultaneous with the appearance of the ball, triggered by the interruption of the second laser beam (see supplemental Videos 1, 2, available at www.jneurosci.org as supplemental material). Targets and distractors were presented 3–5 s after trial onset (see Fig. 1B). A single pulse of TMS was applied, at different time points (between 40 and 120 ms) after distractor appearance, over the hand area of the subject's left primary motor cortex. In trials with distractor LEDs, one of two LEDs, positioned underneath the endpoint of the trajectory of each ball was illuminated for 130 ms simultaneously with the go signal. On these trials, and on other trials without distractor balls, the timing of the go signal was determined by using the distribution of target delays with respect to trial onset from experiment 1. These had an approximately uniform rectangular distribution between 750 and 1500 ms.
Analysis
Data analysis was performed with custom software written in Matlab (available at http://www.neurobiography.info).
MEPs.
The peak-to-peak MEP amplitude was determined automatically by finding the minimum and maximum values within a search window, starting ∼20 ms and ending ∼70 ms after TMS. In all cases, individual MEPs were visually inspected and rejected if they were contaminated with a voluntary contraction or large fluctuations in the baseline before the TMS pulse. MEP amplitudes <0.25 mV were discarded as being indistinguishable from baseline. A mean ± SE of 10.5 ± 2.1% of trials across experiments was discarded because of outlying reaction times (RTs) (<150 or >1500 ms), noisy EMG background, or the absence of MEPs.
To rule out the possibility that differences in MEP amplitude were attributable to differences in RT for near and far trials, we regressed out the effect of RT on MEP amplitude and report here only the corrected MEP amplitudes (additional data are provided in supplemental Tables S2 and S3, and supplemental material, available at www.jneurosci.org).
To correct for between-subject differences in baseline MEP amplitude (e.g., because of electrode placement or muscle size), peak-to-peak MEP amplitudes were standardized with respect to the mean MEP amplitude on no-ball trials (collapsing across hand positions), separately for each subject and TMS time point. In experiments in which a no-ball condition was not performed (experiments 1, 5, 6, and S1), the mean MEP amplitude across both near and far conditions was used for standardization purposes (supplemental Tables S2, S3, available at www.jneurosci.org as supplemental material). The effects of these standardization procedures on the statistical significance of the reported results were minimal (supplemental Table S3, available at www.jneurosci.org as supplemental material).
EMG.
EMG data were rectified and the mean EMG baseline activity was compared statistically between experimental conditions, to verify that changes in the baseline EMG activity did not confound our results (supplemental Table S4, available at www.jneurosci.org as supplemental material). We also extracted the onset of voluntary EMG activity, defined as the first point after MEP offset at which the mean EMG over the next 10 ms was ≥3.09 SD above baseline (Hodges and Bui, 1996). These onsets were used to determine EMG “twitches” in experiment 5b.
Statistical analysis.
We used separate two-tailed t tests, rather than omnibus ANOVAs, to test novel hypotheses (e.g., experiments 1, 4, 5, and S1) for two reasons: (1) Since MEPs were standardized for each TMS time point separately, pooling across near and far distractor conditions, there could not be a main effect of TMS time point; (2) in experiments without a separate baseline, data from near and far conditions were not independent. In these cases, the near data were compared against the null hypothesis of 1.0. In other cases, near and far conditions were compared directly. For experiments 2, 3, and 6, we tested one-tailed hypothesis-driven predictions concerning differences between the effects of near and far distractors, using t tests and planned comparisons. To protect against false positives, we used a Bonferroni correction for multiple comparisons. Throughout this manuscript, corrected p values are reported. In addition, we performed numerous bootstrapping and resampling procedures to construct null distributions for each of the critical reported comparisons. In every case, this bootstrapping procedure supported the inferences derived from the t tests, so we report only the standard parametric tests in this manuscript. The bootstrap statistics are presented in supplemental Table S6 (available at www.jneurosci.org as supplemental material). Additional details of the methods are available in supplemental material (available at www.jneurosci.org).
Results
Experiment 1: early hand-centered modulation of corticospinal excitability
We first determined whether and when corticospinal excitability was modulated as a function of the distance of a rapidly approaching three-dimensional visual distractor ball with respect to the subject's hand (experiment 1). Subjects made a speeded right index finger abduction movement in response to a central go signal (the offset of the central fixation LED) while a distractor ball appeared, approaching the workspace on the left or right side (Fig. 1; supplemental Videos 1, 2, available at www.jneurosci.org as supplemental material). On one-half of the trials within each block, and unpredictably, the distractor ball fell near to the subject's hand, and in the other one-half it fell far from their hand. The go signal was simultaneous with, and triggered by, the appearance of the distractor ball. The subjects were instructed to ignore the distractor stimuli and respond only to the target. We found no significant differences between near and far trials with respect to behavioral performance [supplemental Table S2 (available at www.jneurosci.org as supplemental material) shows RT, error, and d′ data across all seven experiments].
For 10 subjects, TMS was applied at five time points (40–120 ms, in 20 ms intervals) after the appearance of the distractor ball (Table 1, experiment 1). This enabled us to measure corticospinal excitability over most of the interval between the appearance of the ball and the very earliest voluntary EMG responses.
Across all TMS time points, the mean MEP amplitude for trials with near distractor balls (standardized mean ± SE amplitude, 0.973 ± 0.011) was significantly lower than for trials with far distractor balls (1.027 ± 0.011; t(9) = 2.41; p = 0.039). Considering each time point separately, the effect of distractor ball distance was significant only at 80 ms after distractor appearance [t(9) = 4.08; p = 0.014, comparing the mean of near distractor ball trials (mean ± SE, 0.91 ± 0.022) against 1.0, and after correcting for five comparisons] (Fig. 2A). For the other four time points, no significant effects were found, suggesting a return to baseline (i.e., no differences between near and far distractor balls) within 20 ms. Within the 80 ms TMS time point, when the hand was on the left of fixation, distractor balls appearing on the left resulted in smaller MEPs than distractor balls appearing on the right, and vice versa for the right hand position (interaction between distractor ball position and hand position; F(1,9) = 16.63; p = 0.003) (Fig. 2B). That is, balls approaching near the hand reduced corticospinal excitability compared with balls falling far away. These results demonstrate a significant modulation of corticospinal excitability at 80 ms after visual stimulation, which is consistent with a hand-centered coding of the approaching objects.
Figure 2.

Rapid modulation of corticospinal excitability by visual stimuli in hand-centered coordinates. A, Mean ± SE peak-to-peak MEP amplitude for near (dark gray) and far (light gray) distractor balls across five TMS time points (40–120 ms). Early after distractor onset (40–80 ms), mean peak-to-peak MEP amplitude in trials with near distractor balls was lower compared with trials with far distractors, but this difference was significant only at the 80 ms time point (*p < 0.05). The near condition was tested against 1.0, and p values were corrected for five comparisons. B, Mean ± SE peak-to-peak MEP amplitude for distractor balls presented to the left and right of fixation for the two hand positions separately, at the 80 ms time point. The difference between peak-to-peak MEP amplitudes for left and right distractor balls reversed when the position of the right hand shifted from left (filled circles) to right (open circles). This pattern was reflected in a significant interaction between hand position and distractor ball position. The asterisks denote statistical significance of the interaction between hand and ball positions, as evaluated with ANOVA (**p < 0.01).
Experiment 2: hand-centered coding of approaching objects is independent of eye position
We next examined whether corticospinal excitability during response preparation is also modulated by the distance of the distractor ball from the location of subjects' visual fixation, or rather is fixed predominantly in hand-centered coordinates, similarly to certain neuronal populations in macaque premotor and parietal cortices (Graziano et al., 1994, 1997; Fogassi et al., 1996, 1999; Graziano, 1999).
With another group of 10 subjects, we repeated the experimental design described above, with some modifications (Table 1, experiment 2). Rather than maintaining fixation centrally throughout the experiment, at the beginning of each trial, one of two fixation LEDs was illuminated, pseudorandomly either on the left or on the right side of the workspace, underneath the end of the left and right distractor ball trajectories, respectively. Thus, in one-half of the trials with a distractor ball, the ball appeared peripherally, on the opposite side to the visual fixation position, whereas in the remaining trials, the ball appeared parafoveally, on the same side as visual fixation. Based on the results of experiment 1 (i.e., a significant effect at 80 ms and a nonsignificant trend at earlier time points), two TMS time points were tested, at 70 and 80 ms after distractor ball appearance.
We predicted a significant reduction of corticospinal excitability when the distractor ball fell near to compared with far from the hand (i.e., in a hand-centered manner) and that this hand-centered representation would be found regardless of the distance of the distractor ball from the fixation position. We used one-tailed planned comparisons for these hypothesis-driven predictions.
A significant effect was found for the 70 ms TMS time point, in which, as predicted, and in accordance with the results of experiment 1, the mean MEP amplitude for near-hand distractor ball trials (mean ± SE amplitude, 0.936 ± 0.045) was significantly lower than for far-hand distractor ball trials (1.013 ± 0.034; F(1,9) = 3.79; p = 0.020) (Fig. 3). There was also a weak (power, 0.259) and nonsignificant trend toward an effect of fixation position, with smaller MEPs for trials with distractor balls presented on the same side as visual fixation, compared with on the opposite side (0.954 ± 0.042 vs 0.995 ± 0.035, respectively; F(1,9) = 2.35; p = 0.080). Note that fixation position was manipulated orthogonally to hand position. There was no effect of distractor ball distance at the 80 ms time point, possibly indicating that the hand-centered effect is maximal at 70 ms after visual presentation or that there is some variability between subjects (see experiment 6). These results suggest that, 70 ms after the presentation of a visual distractor, corticospinal excitability reflects the coding of visual objects primarily with respect to the distance from the hand and does so independently from any potential contribution of visual fixation position.
Figure 3.

Hand-centered modulation of corticospinal excitability by visual distractors is independent of eye position. Mean ± SE peak-to-peak MEP amplitude with TMS delivered 70 ms after distractor ball appearance shows a significant effect of distractor ball distance relative to the hand (dark grays, near; light grays, far). MEP amplitudes were modulated primarily by the distance of the distractor balls from the hand and less, and independently, by the distance from visual fixation. The asterisk denotes statistical significance of planned comparisons using hand-centered predictions (*p < 0.05).
Experiment 3: near distractor balls suppress corticospinal excitability during response preparation
The first two experiments provided evidence for decreased corticospinal excitability after presentation of approaching visual distractor balls near to compared with far from subjects' hands. We hypothesized that this early reduction reflects the proactive inhibition of an undesirable response (Koch et al., 2006; Mars et al., 2007; van den Hurk et al., 2007; van Elswijk et al., 2007), such as an avoidance movement elicited by the approaching ball. We therefore measured the effect of voluntary inhibition of the motor response at the time of distractor presentation (Coxon et al., 2006, 2007) to examine whether this would suppress MEP amplitudes to a similar level as found with near distractor balls. Additionally, we included a no-distractor “baseline” condition to assess whether near balls decrease, or else far balls increase, corticospinal excitability. For this purpose, we repeated the basic design of experiment 1 with a different group of 10 subjects (Table 1, experiment 3). This time, 25% of the trials contained a central red stop signal, presented randomly 200–300 ms before the go signal, instructing subjects to inhibit their response to the upcoming go signal. We also included trials (33%) in which no visual distractor was presented (“no-ball” control trials).
We predicted that a process of proactive inhibition by near distractor balls should result in the following: (1) lower MEP amplitudes for near trials at early TMS time points (70–90 ms), compared with both far and with no-ball trials; (2) no differences for late TMS time points (100–120 ms) between either near and far, or near and no-ball trials; and (3) no significant difference in MEP amplitudes on near distractor trials between go and stop conditions, for the early (70–90 ms) TMS time points.
In the go trials when a distractor ball was presented, we again found a hand-centered reduction of MEP amplitudes, with a significant effect of distractor distance (near vs far), when averaged across the early TMS time points (70–90 ms; near, 0.954 ± 0.025; far, 1.011 ± 0.016; t(9) = 2.87; p = 0.037, corrected for two comparisons) (Fig. 4), but not across the late TMS time points (100–120 ms; t(9) = 1.75; p = 0.226). Looking separately at the 70, 80, and 90 ms TMS time points, only at 70 ms were there significantly lower MEP amplitudes for near than far distractor trials (0.946 ± 0.028 vs 1.036 ± 0.038, respectively; t(9) = 3.07; p = 0.040, corrected for three comparisons). This confirms that the reduction of corticospinal excitability peaks at the 70 ms time point, and then diminishes, replicating the findings of experiment 2.
Figure 4.

Suppression of corticospinal excitability is found for distractor balls approaching the hand, but not for static LED flashes. Mean ± SE peak-to-peak MEP amplitude for near (dark gray) and far (light gray) distractor balls (filled columns) and distractor LEDs (striped columns), pooled over the 70–90 ms TMS time points. When subjects responded to the go signal, peak-to-peak MEP amplitudes were significantly suppressed by distractor balls approaching near to compared with far from the hand, and also for near compared with control trials with no ball. No such suppression was found when distractor LEDs (positioned at the end of the trajectories of the balls) were illuminated instead of the approaching balls. When subjects voluntarily inhibited their motor responses after a stop signal, no significant difference was found between MEP amplitudes on near and far distractor trials, regardless of the distractor type. The asterisks denote statistical significance of t tests between near and no-ball (*p < 0.05), and near and far distractor ball trials (*p < 0.05, one-tailed corrected for two comparisons). n.s., Not significant.
Furthermore, mean MEP amplitude for early TMS time points on trials with near distractor balls was significantly reduced compared with no-distractor control trials (t(9) = 1.89; p = 0.046, compared against 1.0), whereas MEPs for trials with far distractor balls were not significantly different from no-ball trials (Fig. 4). These results show that MEP amplitudes were reduced for the near condition, and not enhanced for the far condition, extending the results of experiments 1 and 2.
Additional support for the early suppression of corticospinal excitability after near distractor balls is provided by comparison of MEP amplitudes between stop trials in which subjects were instructed to inhibit their motor responses before ball appearance, and go trials with either near or far distractor balls. For early TMS time points, mean MEP amplitudes on stop versus go trials with near distractor balls did not differ significantly (t(9) = 0.54; corrected p = 1, with a statistical power of 0.82, based on the effect size of the near vs far go conditions for the early TMS time points) (Fig. 4). This was not the case when comparing MEP amplitudes between go and stop trials for far distractor balls (t(9) = 2.85; p = 0.038, corrected for two comparisons).
Finally, MEP amplitudes on stop trials were not significantly different between near and far distractor ball trials, either for the early or the late TMS time points (Fig. 4). That is, after voluntary inhibition of motor responses, we did not detect any hand-centered modulation of MEP amplitudes. We cannot, however, definitively rule out the possibility that hand-centered modulation may still exist in the stop trials but was not observed because of possible floor effects on MEP amplitudes.
Although these results do not allow conclusions regarding the precise mechanisms involved in these forms of inhibition, they do at least support the hypothesis that an inhibitory process is responsible for the decrease in corticospinal excitability after the appearance of the near ball.
The data presented thus far provide clear evidence for the suppression of corticospinal excitability after the presentation of rapidly approaching three-dimensional visual stimuli near to relative to far from the hand. Together, these results might best be explained in terms of a form of response competition between the ongoing go task and a newly emerging motor plan, elicited by the approaching ball, possibly for an avoidance response. To examine this possibility further, we needed to establish that such effects were not evident when the distractor was not a rapidly approaching object.
Experiment 4: hand-centered coding of visual space is observed with approaching balls, but not static LED flashes
We next determined whether the hand-centered suppression that we identified after distractor ball presentation can be attributed merely to the sudden occurrence of visual events near the hand. We therefore repeated experiment 3 (with the additional stop trials) on a different group of 10 subjects (Table 1, experiment 4). This time, the moving distractor balls were replaced with static distractor LEDs, positioned beneath the endpoints of the trajectories of the balls. One of the distractor LEDs flashed for 130 ms, simultaneously with the central go signal, either near to or far from the responding hand. We predicted that corticospinal excitability would not be modulated by the static visual distractor LEDs. Specifically, we expected to find no reduction of MEP amplitudes after presentation of distractor LEDs near to relative to far from the hand.
Overall, the subjects' percentage correct performance in this experiment was similar to that of experiment 3 (98.2 ± 0.6 vs 98.1 ± 0.3% correct; d′ = 3.71 ± 0.03 vs 3.79 ± 0.06, respectively). No significant differences in performance were found with respect to the distance of the distractor LED from the hand, the trial type (go vs stop), or between the two experiments. Regarding the MEP amplitudes, as predicted, we found no difference between near and far distractor LED trials, both for early and late TMS time points (Fig. 4). Furthermore, and contrary to the results of experiment 3, significantly smaller MEPs were found on stop compared with go trials with near distractor LEDs, specifically for early TMS time points (t(8) = 3.47; p = 0.017, one-tailed, corrected for two comparisons).
These results are similar to certain results from neurophysiological studies of bimodal neurons in premotor cortex, which showed visual selectivity for three-dimensional objects approaching toward and receding from the hand, and lower responses to static or two-dimensional visual stimuli (Graziano et al., 1997).
Experiment 5a: behavioral indices show hand-centered response inhibition for approaching balls, but not for static LEDs
Additional support for the inhibition of motor responses in a hand-centered reference frame derives from behavioral error rates in a “go/stay” task. This experiment was conducted with a different group of 16 subjects (Table 1, experiment 5). In this experiment, on 50% of the trials the go signal did not appear. Subjects were instructed to respond as quickly as possible, only on go trials, and to withhold responses when the go signal was not presented (“stay”). A visual distractor, which was pseudorandomly either moving (a ball) or static (an LED), was presented on every trial, either near to or far from the responding hand. TMS pulses were applied to the motor cortex contralateral to the responding hand. The MEP data are reported in the following section (experiment 5b).
On go trials, performance with near distractor balls was significantly worse than on trials with far distractor balls (92.9 ± 1.3 vs 97.3 ± 1.0% correct, respectively; t(15) = 3.02; p = 0.017, two-tailed, corrected; d′ = 3.16 ± 0.09 vs 3.51 ± 0.07, respectively; t(15) = 3.16; p = 0.013, two-tailed, corrected for two comparisons). As presented in Figure 5A, when a ball approached the responding hand, subjects were more likely not to respond (i.e., there were more missed responses) than when the ball appeared far from the hand. The effects were similar for both left (p = 0.003) and right hand positions separately (p = 0.07). In contrast, performance with near and far distractors on stay trials did not differ significantly (96.0 ± 1.1 vs 97.1 ± 0.9%; d′ = 3.30 ± 0.06 vs 3.35 ± 0.12, respectively).
Figure 5.

Behavior demonstrates selective response inhibition by distractor balls approaching the hand. Mean ± SE percentage of “hits” for distractor balls (A) and LEDs (B) presented to the left or right of fixation, separately for the two hand positions (filled circles, left; open circles, right). When the probability of responses on each trial was 0.5 (i.e., a go signal appeared in only one-half of the trials), subjects were prone to inhibit their response to the targets (i.e., to miss) with a concurrent near distractor ball. No such hand-centered modulation of performance was evident with static LED distractors. The asterisk denotes significance of the interaction between hand position and ball position (*p < 0.05, two-tailed corrected for two comparisons).
This hand-centered modulation of behavioral performance was also selective for approaching balls, compared with static flashing distractors. Overall, performance was nearly identical between trials with distractor balls (95.8 ± 0.5% correct; d′ = 3.33 ± 0.06) and LEDs (95.9 ± 0.6% correct; d′ = 3.33 ± 0.08). Contrary to what was found for balls, however, no differences were found between near and far distractor LEDs on go trials (Fig. 5B).
These results converge with the MEP data obtained from experiment 4, which showed that the decrease in corticospinal excitability during response preparation is selective for a ball approaching the hand and is not merely attributable to sudden visual changes near the hand in the peripheral visual field. Since the ball and LED distractors differed in several respects (e.g., size, luminance), it is impossible at present to establish definitively which aspects are critical for driving the hand-centered suppression that we report here. However, we found no behavioral evidence (i.e., in RT, percentage correct, or d′ measures) to suggest that the two distractor types varied in salience (which might be expected to affect behavioral performance) (see also supplemental Table S2, available at www.jneurosci.org as supplemental material).
Experiment 5b: hand-centered enhancement of corticospinal excitability with response uncertainty
We suggest that our results thus far may best be explained in terms of proactive inhibition during an action selection process: to perform the go task as rapidly and successfully as possible, any avoidance response evoked by the approaching ball needs to be inhibited. According to this account, if subjects were not able sufficiently to prepare a motor response before the appearance of the go signal, the effects of proactive inhibition would not be observed. To test this directly, we used the go/stay task in which, while a distractor appeared on every trial (either a ball or an LED), the go signal appeared on only 50% of the trials (as described above) (Table 1, experiment 5). In this way, we introduced a substantial degree of response uncertainty, forcing subjects to adopt a more conservative response preparation strategy. Subjects were encouraged to perform the task quickly and to refrain from making errors, both misses and false alarms.
Two hypotheses were entertained: first, it is possible that, when no motor response is prepared, there should be no difference between MEP amplitudes on near and far trials, regardless of the visual distractor type (moving ball vs static LED) or response type (go vs stay). Second, in the absence of any preparation to push the response button, the avoidance movement elicited by the approaching ball would remain unchallenged by any competing responses. This might result in greater MEP amplitudes on near relative to far ball go trials. For this experiment, TMS was delivered 70 ms after the appearance of the distractor. Since this experiment was designed to test both a negative (i.e., no difference between near and far ball trials with response uncertainty) and a positive prediction (MEP enhancement with near vs far balls), we used a larger group of subjects (n = 16; power = 0.998, based on the near vs far effect size for the 80 ms TMS time point of experiment 1).
Contrary to the first hypothesis, and consistent with the second, the TMS results continued to support the notion of hand-centered modulation of MEP amplitude for the go ball trials only. This was demonstrated by a significant effect of distance from the hand for the distractor balls (t(15) = 2.58; p = 0.042, two-tailed, corrected for two comparisons), but not for the LEDs. Furthermore, the MEPs were now larger for the near ball trials (1.044 ± 0.019), compared with the far ball trials (0.992 ± 0.022). No such trend was found for the stay trials. That is, with no advanced response preparation, MEP amplitude may now reflect the additional excitatory effects of the distractor ball approaching the hand (i.e., a potentially avoidance-related response), early in the process of response selection.
This interpretation is further supported when we focus, post hoc, on the trials in which subjects produced a voluntary EMG response (here called a muscle “twitch”) (see supplemental material, available at www.jneurosci.org) after the MEP, compared with trials in which no twitch was produced. This analysis was performed regardless of MEP size, whether a response was required, or whether a button press was registered. Within these twitch trials, we found a significant increase of MEP amplitudes for near compared with far trials, and specifically for the distractor balls (t(15) = 3.13; p = 0.014, two-tailed, corrected for two comparisons) (Fig. 6A), but not for the LEDs. Importantly, this result did not depend on differences in EMG baseline activity before the MEP (supplemental Table S4, available at www.jneurosci.org as supplemental material). By sharp contrast, on trials in which no voluntary EMG response occurred, there were no significant effects of distractor distance, either for balls or LEDs. This finding provides direct support for our interpretation of proactive inhibition during response selection: when competition between the avoidance and the go responses was eliminated or, at least, desynchronized, the hand-centered suppression of MEP amplitudes was reversed and became hand-centered enhancement, specifically on those trials in which the hand was most ready to respond.
Figure 6.

With response uncertainty, late muscle activation predicts hand-centered enhancement of MEP amplitude by approaching balls. Mean ± SE peak-to-peak MEP amplitude for distractor ball trials in which a voluntary EMG activity onset (twitch) was recorded after the MEP (A) and for trials in which no late muscle activation was recorded (B). On the EMG twitch trials, near distractor balls elicited greater MEP amplitudes, compared with far distractor balls, both in the left and the right hand positions. Importantly, this was not the case for the left and right no twitch trials. The asterisks denote statistical significance of the interaction between hand position and ball position, as evaluated with ANOVA (**p < 0.01, corrected for two comparisons).
Additional evidence for the enhancement of MEP amplitudes for “near” trials during response uncertainty is provided in experiment S1, reported in supplemental material (available at www.jneurosci.org). This experiment additionally demonstrates that any contribution of exogenous attentional orienting cannot be solely responsible for the hand-centered coding of approaching objects reported here.
Experiment 6: precise time course and muscle specificity of hand-centered coding of approaching objects
An additional experiment was run to answer questions concerning the precise time course of the hand-centered modulation of corticospinal excitability. Specifically, across the replications of our basic finding, slight differences arose in latency between the 80 ms (experiments 1 and S1) and the 70 ms time points (experiments 2, 3, and 5b). We hypothesized that the hand-centered effect starts or peaks at ∼70 ms and has a duration of ∼10–20 ms, perhaps with some intersubject variability in its time course. For this reason, the effect may also be seen at 80 ms after ball appearance.
To test this hypothesis, we repeated experiment 1 with a new group of six subjects, with several minor adjustments (see supplemental material, available at www.jneurosci.org). First, we used five TMS time points between 65 and 85 ms after ball appearance, in 5 ms intervals. Second, we recorded MEPs from both the FDI and the abductor digiti minimi (ADM) muscles. We hypothesized that the excitability of the ADM, which is not directly involved in the task-related index finger abduction response, should not be in competition with any responses elicited by the approaching ball, and therefore should not show hand-centered suppression of MEP amplitudes, contrary to the FDI.
The mean MEP amplitudes across the high-resolution time course are presented in Figure 7. As predicted, we observed the greatest effect of distractor ball position on MEP amplitudes at the 70 ms time point, in which MEP amplitudes were again significantly smaller for near (mean ± SE, 0.862 ± 0.053) compared with far trials (t(5) = 2.64; p = 0.023, comparing the mean of near distractor ball trials against 1, one-tailed). The mean MEP amplitude pooled across the 70, 75, and 80 ms time points was also significantly lower than one, suggesting that the effect endured for as long as 10 ms. The time course of the effect in individual subjects confirmed this suggestion, with each subject showing at least a 7% suppression of MEP amplitudes for near compared with far trials of at least two adjacent time points (i.e., of ∼10 ms), typically at 70–80 ms after ball appearance.
Figure 7.

Hand-centered modulation of corticospinal excitability by approaching balls: high-resolution time course. Mean ± SE peak-to-peak MEP amplitude for near and far distractor balls across five TMS time points, at 5 ms temporal resolution (65–85 ms). The captions are as in Figure 2. The asterisk denotes statistical significance of one-tailed t tests between the near condition and the null hypothesis of 1.0 (*p < 0.05).
In the ADM, no significant differences were found between the near and far conditions at any of the five time points (all values of p ≥ 0.49) nor did we observe any clear trends across the time course (supplemental Fig. S2, available at www.jneurosci.org as supplemental material). This result should be interpreted with some caution, however, given the relatively small number of subjects and the fact that MEPs were not elicited from one subject's ADM muscle. However, given that we found a significant effect in the FDI in the same group of subjects, even after excluding all the data of the subject without MEPs in the ADM (p = 0.039), this result further strengthens our interpretation of proactive inhibition during response selection at 70–80 ms after distractor ball appearance.
Discussion
In the current study, we provide direct evidence for hand-centered coding of approaching visual stimuli during motor preparation. Such a coding was manifested in modulations of corticospinal excitability evoked by presenting three-dimensional visual distractors rapidly approaching subjects' hands. These hand-centered effects were repeatedly observed across independent experiments, occurred when the responding hand was positioned both on the left and on the right of the body midline (Figs. 2, 6, 7), regardless of visual fixation position (Fig. 3), and were dependent on motor response preparation. Furthermore, they did not occur for static distractors (Fig. 4), suggesting that this hand-centered modulation may be specific for potentially aversive approaching distractors, although the latter point awaits confirmation by future studies.
The latency at which visual spatial information began selectively to influence corticospinal excitability was 70 ms after ball appearance, with an effective duration of as short as 10 ms. In this narrow time window, hand-centered processing of visual information had already occurred, even while the visual distractor ball was still falling toward the hand, highlighting the rapidity of the hand-centered coding of visual space identified here.
Proactive inhibition of avoidance movements during response selection
In our study, when a go response was required on every trial and the subjects were able to prepare the appropriate response (i.e., pushing the response button) in advance, a distractor ball approaching near the hand reduced corticospinal excitability at 70–80 ms after ball appearance (experiments 1, 2, 3, and 6) (Figs. 2–4, 7). This reduction may reflect the inhibition of an avoidance-related motor plan, which automatically emerges only when the ball approaches the responding hand (experiment 3) (Fig. 4). Indeed, flexible motor behavior requires the ability to inhibit one movement while concurrently executing another (i.e., selective inhibition) (Coxon et al., 2007). Such inhibition can be effective to suppress undesirable movements not only after they have been initiated but also proactively before any EMG response is released (Boulinguez et al., 2008).
Numerous TMS studies have shown suppression of MEP amplitude during instructed delay periods (foreperiods), which serves to hold back the premature execution of responses (Hasbroucq et al., 1997, 1999a,b; Touge et al., 1998; Davranche et al., 2007). Similarly, during response selection, MEP amplitudes are suppressed for the nonprepared effector and increased for the prepared response (Koch et al., 2006; van Elswijk et al., 2007), but only when sufficient foreperiod information about the response was available to the subjects (Mars et al., 2007; van den Hurk et al., 2007). Likewise, in our study, the reduction of corticospinal excitability by the distractor was evoked only when both advance preparation was possible, and when an approaching, potentially aversive distractor was used. When response uncertainty was introduced, such that the go response could not confidently have been prepared in advance, and a temporal offset was therefore created between the task-related and the avoidance responses, corticospinal excitability was enhanced for active trials in which the ball approached near the responding hand (experiments 4 and S1) (Fig. 6; supplemental Fig. S1, available at www.jneurosci.org as supplemental material). Conversely, for a muscle that did not participate in the preparation for the go response (i.e., the ADM), no hand-centered modulation of corticospinal excitability was observed. In other words, when competition between the avoidance and go responses is eliminated (or temporally desynchronized), no suppression of the avoidance response is required, and an active component of the avoidance movement may be revealed in the MEP amplitude. Our results therefore indicate an interaction between the motor consequences of the rapid visual processing of objects approaching the hand and the voluntary task-related motor plans for that hand.
Mechanisms underlying rapid visual modulation of corticospinal excitability
Threat-induced freezing
The early onset of corticospinal modulations in the present study (70–80 ms after the appearance of the ball) reflects the very rapid transformation of visuomotor information. One possible framework within which to interpret our results could be threat-induced startle or freezing responses. A recent functional magnetic resonance imaging study (Butler et al., 2007) reported decreased activity in bilateral primary motor cortex during periods in which subjects expected to receive painful stimuli (compared with a condition without pain expectation). Using TMS, Cantello et al. (2000) found a significant reduction of corticospinal excitability at 55–75 ms after the onset of unexpected, sudden, and very bright task-irrelevant light flashes. Similar results were reported after the presentation of electrical shocks (Maertens de Noordhout et al., 1992) or loud auditory stimuli (Furubayashi et al., 2000). Pain-inducing stimulation to the hand also causes immediate (Urban et al., 2004) and long-lasting (Farina et al., 2001; Svensson et al., 2003) decreases of corticospinal excitability for distal arm muscles. Furthermore, MEPs were inhibited at later stages of processing (>1 s after stimulus presentation) when subjects passively viewed movies of painful simulation applied to specific muscles on a model (Avenanti et al., 2005). It has been suggested that such inhibition may act as a sort of motor “decerebration,” allowing the spinal motor system freely to develop protective responses after noxious stimulation (Farina et al., 2003).
We believe that the results described in the studies mentioned above do not directly correspond with our findings, and that pain- or startle-related inhibition cannot explain the results we report here, for two reasons. First, it cannot explain the reversal of the direction of hand-centered modulation when subjects could not confidently prepare a response in advance. Second, the time course of startle-related inhibition based on TMS studies (Cantello et al., 2000) just precedes the corticospinal modulations reported here, which are restricted to 70–80 ms after distractor ball appearance. Furthermore, Cantello et al. (2000) did not provide (or search for) evidence for any hand-centered mechanisms, so any direct links with our results remain speculative. We therefore propose that, although fear-induced startle might play some role in the mechanism under investigation, our hand-centered modulation represents a later stage of selection between an appropriate avoidance/defensive response and the task-related response.
Cortical mechanisms
Numerous cortical areas have been shown to modulate corticospinal excitability and may therefore contribute to the results reported here. These include the primary somatosensory area (Avenanti et al., 2007), posterior parietal areas, and the premotor cortex (for review, see O'Shea et al., 2008).
Cooke and Graziano (2003) studied macaque muscle activity during defensive movements evoked by aversive cutaneous stimulation. They distinguished between startle-related EMG activity occurring as early as ∼20–30 ms after stimulus onset and later muscle activity starting ∼70 ms after stimulus onset [Cooke and Graziano (2003), their Fig. 2C]. Only the latter activity was spatially specific to the muscles involved in the defensive response. Very similar motor responses were evoked by electrical macrostimulation of bimodal regions of the premotor cortex (Graziano et al., 2002). Indeed, M1 and the premotor cortex are densely interconnected, both in humans and in monkeys (Shimazu et al., 2004; Dum and Strick, 2005; Koch et al., 2006; O'Shea et al., 2007; Davare et al., 2008). The modulation of MEP amplitude with respect to approaching balls that we report here may therefore reflect ongoing activity in the premotor cortex.
Some premotor neurons are active after visual stimulation at time periods similar to the ones we have described and are thought to encode the significance of visual cues for response selection (Cisek and Kalaska, 2005). In humans, the premotor cortex has been shown to participate in the visuomotor transformations required to configure hand posture with respect to objects (Davare et al., 2006). Furthermore, recent studies have shown a direct influence of premotor over primary motor cortex as early as 75 ms after a go signal (Koch et al., 2006; O'Shea et al., 2007). Although any comparisons between data drawn from monkeys and humans, and using such different methods, should be made with caution, given the timing and the spatial specificity of the above responses with respect to visual events, these mechanisms fit very well with our results and suggest the involvement of human premotor areas with the hand-centered coding of visual space. Following Band and van Boxtel (1999), we tentatively suggest that one source of the hand-centered modulation that we report may be the premotor cortex, the site of modulation may be the primary motor cortex, and the modulation is manifested by changes in MEP amplitudes (Band and van Boxtel, 1999).
Pruszynski et al. (2008) demonstrated rapid arm muscle responses that were sensitive to the direction and distance of visual targets at 95 ms after a go signal. This timing fits very well with our findings, assuming a 20–25 ms conduction time between the primary motor cortex and the intrinsic hand muscles. Pruszynski and colleagues suggested that such rapid motor responses are capable of all the sophistication attributed to voluntary control, within the constraints of their limited processing time. Additional research is required to determine the precise neurophysiological mechanisms that might enable such rapid and sophisticated visuomotor spatial processing in the human brain.
Subcortical mechanisms
In monkeys, spinal interneurons have been shown to participate in the control of movement during preparation periods, displaying both excitatory changes in the same direction as the subsequent movement-related activity, and inhibitory changes that may reflect superimposed mechanisms suppressing muscular output (Prut and Fetz, 1999). It is possible that the hand-centered modulation that we observed reflects such subcortical movement preparation, although it is important to note that, in our results, the critical movement selection processes occurred based on the position of the visual distractor ball relative to the responding hand, so any candidate subcortical source of the reported modulations of corticospinal excitability would have to have access to such information.
Hand-centered visual space and attention
In our experiments, both near and far balls were distractors; however, it is possible that near distractors were more salient than far distractors and that this somehow resulted in reduced levels of corticospinal excitability. Such differences in salience could have been the result of subjects covertly attending toward their responding hand, despite the instructions and the central visual task. We addressed these concerns in several experiments.
First, when the balls were substituted with LEDs positioned near and far from the hand, we found no significant hand-centered modulations of corticospinal excitability (experiment 4) (Fig. 3) or of percentage correct performance (experiment 5) (Fig. 5). Second, we manipulated subject's overt attention, by shifting their fixation between positions on the same or opposite side as the impending distractor ball (experiment 2). When shifting the fixation position with respect to the ball and hand positions, we found a small, but nonsignificant difference between balls approaching the same side, and balls approaching the opposite side of fixation (Fig. 3). At the same time, however, the distance of the ball from the hand (manipulated orthogonally and independently from the distance from fixation) significantly modulated corticospinal excitability. Finally, we used a cueing paradigm to attract subjects' exogenous covert visuospatial attention toward versus away from the distractor ball, just before its appearance (experiment S1; supplemental Fig. S1A; supplemental material, available at www.jneurosci.org). Again, this manipulation did not significantly modulate corticospinal excitability, while the distance of the distractor from the hand remained most effective (albeit marginally significantly), regardless of whether subjects' attention was cued toward or away from the hand (supplemental Fig. S1B, available at www.jneurosci.org as supplemental material).
We therefore conclude that the role of overt and covert visuospatial attention in the results that we report here is marginal, or, at most, complementary, to the role of hand position. A rapid and automatic hand-centered response to potentially aversive approaching visual stimuli is a better description of our results as a whole.
Conclusions
We demonstrate here that the human motor system is rapidly informed about potential collisions between nearby objects and specific body parts that are about to move. This specialized system for transforming nearby sensory inputs into rapid and appropriate motor outputs is ideally suited to serve as a sensorimotor interface for driving defensive movements away from potentially harmful approaching objects.
Footnotes
This work was supported by an Avenir grant and a Travel Award from Boehringer Ingleheim Fonds (T.R.M.). N.P.H. was supported by the Royal Commission for the Exhibition of 1851 (Imperial College, London, UK), The Lady Davis Fellowship Trust, and the Interdisciplinary Center for Neural Computation (Hebrew University, Jerusalem, Israel). C.B. was supported by the European Mobility Fellowship and the Avenir grant. Y.R. was supported by Marie Curie Research and Training Network Discos, European Science Foundation Eurochore BASIC, and Hospices Civiles de Lyon. A.F. was supported by Inserm Avenir Grant R05265CS and Agence Nationale de la Recherche Grant R08118CS. We thank Francesco Pavani for helpful comments, Paul C. J. Taylor for comments on previous versions of this manuscript, and our subjects.
References
- Avenanti A, Bueti D, Galati G, Aglioti SM. Transcranial magnetic stimulation highlights the sensorimotor side of empathy for pain. Nat Neurosci. 2005;8:955–960. doi: 10.1038/nn1481. [DOI] [PubMed] [Google Scholar]
- Avenanti A, Bolognini N, Maravita A, Aglioti SM. Somatic and motor components of action simulation. Curr Biol. 2007;17:2129–2135. doi: 10.1016/j.cub.2007.11.045. [DOI] [PubMed] [Google Scholar]
- Avillac M, Denève S, Olivier E, Pouget A, Duhamel JR. Reference frames for representing visual and tactile locations in parietal cortex. Nat Neurosci. 2005;8:941–949. doi: 10.1038/nn1480. [DOI] [PubMed] [Google Scholar]
- Band GPH, van Boxtel GJM. Inhibitory motor control in stop paradigms: review and reinterpretation of neural mechanisms. Acta Psychol. 1999;101:179–211. doi: 10.1016/s0001-6918(99)00005-0. [DOI] [PubMed] [Google Scholar]
- Beurze SM, Van Pelt S, Medendorp WP. Behavioral reference frames for planning human reaching movements. J Neurophysiol. 2006;96:352–362. doi: 10.1152/jn.01362.2005. [DOI] [PubMed] [Google Scholar]
- Boulinguez P, Jaffard M, Granjon L, Benraiss A. Warning signals induce automatic EMG activations and proactive volitional inhibition: evidence from analysis of error distribution in simple RT. J Neurophysiol. 2008;99:1572–1578. doi: 10.1152/jn.01198.2007. [DOI] [PubMed] [Google Scholar]
- Bremmer F, Schlack A, Shah NJ, Zafiris O, Kubischik M, Hoffmann K, Zilles K, Fink GR. Polymodal motion processing in posterior parietal and premotor cortex: a human fMRI study strongly implies equivalencies between humans and monkeys. Neuron. 2001;29:287–296. doi: 10.1016/s0896-6273(01)00198-2. [DOI] [PubMed] [Google Scholar]
- Brozzoli C, Pavani F, Urquizar C, Cardinali L, Farnè A. Grasping actions remap peripersonal space. Neuroreport. 2009;20:913–917. doi: 10.1097/WNR.0b013e32832c0b9b. [DOI] [PubMed] [Google Scholar]
- Butler T, Pan H, Tuescher O, Engelien A, Goldstein M, Epstein J, Weisholtz D, Root JC, Protopopescu X, Cunningham-Bussel AC, Chang L, Xie XH, Chen Q, Phelps EA, Ledoux JE, Stern E, Silbersweig DA. Human fear-related motor neurocircuitry. Neuroscience. 2007;150:1–7. doi: 10.1016/j.neuroscience.2007.09.048. [DOI] [PubMed] [Google Scholar]
- Cantello R, Civardi C, Cavalli A, Varrasi C, Vicentini R. Effects of a photic input on the human cortico-motoneuron connection. Clin Neurophysiol. 2000;111:1981–1989. doi: 10.1016/s1388-2457(00)00431-4. [DOI] [PubMed] [Google Scholar]
- Cisek P, Kalaska JF. Neural correlates of reaching decisions in dorsal premotor cortex: specification of multiple direction choices and final selection of action. Neuron. 2005;45:801–814. doi: 10.1016/j.neuron.2005.01.027. [DOI] [PubMed] [Google Scholar]
- Cooke DF, Graziano MS. Defensive movements evoked by air puff in monkeys. J Neurophysiol. 2003;90:3317–3329. doi: 10.1152/jn.00513.2003. [DOI] [PubMed] [Google Scholar]
- Cooke DF, Graziano MS. Sensorimotor integration in the precentral gyrus: polysensory neurons and defensive movements. J Neurophysiol. 2004;91:1648–1660. doi: 10.1152/jn.00955.2003. [DOI] [PubMed] [Google Scholar]
- Coxon JP, Stinear CM, Byblow WD. Intracortical inhibition during volitional inhibition of prepared action. J Neurophysiol. 2006;95:3371–3383. doi: 10.1152/jn.01334.2005. [DOI] [PubMed] [Google Scholar]
- Coxon JP, Stinear CM, Byblow WD. Selective inhibition of movement. J Neurophysiol. 2007;97:2480–2489. doi: 10.1152/jn.01284.2006. [DOI] [PubMed] [Google Scholar]
- Crawford JD, Medendorp WP, Marotta JJ. Spatial transformations for eye-hand coordination. J Neurophysiol. 2004;92:10–19. doi: 10.1152/jn.00117.2004. [DOI] [PubMed] [Google Scholar]
- Davare M, Andres M, Cosnard G, Thonnard JL, Olivier E. Dissociating the role of ventral and dorsal premotor cortex in precision grasping. J Neurosci. 2006;26:2260–2268. doi: 10.1523/JNEUROSCI.3386-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davare M, Lemon R, Olivier E. Selective modulation of interactions between ventral premotor cortex and primary motor cortex during precision grasping in humans. J Physiol. 2008;586:2735–2742. doi: 10.1113/jphysiol.2008.152603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davranche K, Tandonnet C, Burle B, Meynier C, Vidal F, Hasbroucq T. The dual nature of time preparation: neural activation and suppression revealed by transcranial magnetic stimulation of the motor cortex. Eur J Neurosci. 2007;25:3766–3774. doi: 10.1111/j.1460-9568.2007.05588.x. [DOI] [PubMed] [Google Scholar]
- di Pellegrino G, Làdavas E, Farnè A. Seeing where your hands are. Nature. 1997;388:730. doi: 10.1038/41921. [DOI] [PubMed] [Google Scholar]
- Dum RP, Strick PL. Frontal lobe inputs to the digit representations of the motor areas on the lateral surface of the hemisphere. J Neurosci. 2005;25:1375–1386. doi: 10.1523/JNEUROSCI.3902-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farina S, Valeriani M, Rosso T, Aglioti S, Tamburin S, Fiaschi A, Tinazzi M. Transient inhibition of the human motor cortex by capsaicin-induced pain: a study with transcranial magnetic stimulation. Neurosci Lett. 2001;314:97–101. doi: 10.1016/s0304-3940(01)02297-2. [DOI] [PubMed] [Google Scholar]
- Farina S, Tinazzi M, Le Pera D, Valeriani M. Pain-related modulation of the human motor cortex. Neurol Res. 2003;25:130–142. doi: 10.1179/016164103101201283. [DOI] [PubMed] [Google Scholar]
- Farnè A, Demattè ML, Làdavas E. Beyond the window: multisensory representation of peripersonal space across a transparent barrier. Int J Psychophysiol. 2003;50:51–61. doi: 10.1016/s0167-8760(03)00124-7. [DOI] [PubMed] [Google Scholar]
- Farnè A, Demattè ML, Làdavas E. Neuropsychological evidence of modular organization of the near peripersonal space. Neurology. 2005;65:1754–1758. doi: 10.1212/01.wnl.0000187121.30480.09. [DOI] [PubMed] [Google Scholar]
- Fernandez-Ruiz J, Goltz HC, DeSouza JF, Vilis T, Crawford JD. Human parietal “reach region” primarily encodes intrinsic visual direction, not extrinsic movement direction, in a visual motor dissociation task. Cereb Cortex. 2007;17:2283–2292. doi: 10.1093/cercor/bhl137. [DOI] [PubMed] [Google Scholar]
- Fogassi L, Gallese V, Fadiga L, Luppino G, Matelli M, Rizzolatti G. Coding of peripersonal space in inferior premotor cortex (area F4) J Neurophysiol. 1996;76:141–157. doi: 10.1152/jn.1996.76.1.141. [DOI] [PubMed] [Google Scholar]
- Fogassi L, Raos V, Franchi G, Gallese V, Luppino G, Matelli M. Visual responses in the dorsal premotor area F2 of the macaque monkey. Exp Brain Res. 1999;128:194–199. doi: 10.1007/s002210050835. [DOI] [PubMed] [Google Scholar]
- Furubayashi T, Ugawa Y, Terao Y, Hanajima R, Sakai K, Machii K, Mochizuki H, Shiio Y, Uesugi H, Enomoto H, Kanazawa I. The human hand motor area is transiently suppressed by an unexpected auditory stimulus. Clin Neurophysiol. 2000;111:178–183. doi: 10.1016/s1388-2457(99)00200-x. [DOI] [PubMed] [Google Scholar]
- Graziano MS. Where is my arm? The relative role of vision and proprioception in the neuronal representation of limb position. Proc Natl Acad Sci U S A. 1999;96:10418–10421. doi: 10.1073/pnas.96.18.10418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graziano MS. Progress in understanding spatial coordinate systems in the primate brain. Neuron. 2006;6:7–9. doi: 10.1016/j.neuron.2006.06.011. [DOI] [PubMed] [Google Scholar]
- Graziano MS, Yap GS, Gross CG. Coding of visual space by premotor neurons. Science. 1994;266:1054–1057. doi: 10.1126/science.7973661. [DOI] [PubMed] [Google Scholar]
- Graziano MS, Hu XT, Gross CG. Visuospatial properties of ventral premotor cortex. J Neurophysiol. 1997;77:2268–2292. doi: 10.1152/jn.1997.77.5.2268. [DOI] [PubMed] [Google Scholar]
- Graziano MS, Taylor CS, Moore T. Complex movements evoked by microstimulation of precentral cortex. Neuron. 2002;34:841–851. doi: 10.1016/s0896-6273(02)00698-0. [DOI] [PubMed] [Google Scholar]
- Graziano MS, Gross CG, Taylor CSR, Moore T. A system of multimodal areas in the primate brain. In: Spence C, Driver J, editors. Crossmodal space and crossmodal attention. Oxford: Oxford UP; 2004. pp. 51–67. [Google Scholar]
- Hallett M. Transcranial magnetic stimulation: a primer. Neuron. 2007;55:187–199. doi: 10.1016/j.neuron.2007.06.026. [DOI] [PubMed] [Google Scholar]
- Hasbroucq T, Kaneko H, Akamatsu M, Possamaï CA. Preparatory inhibition of cortico-spinal excitability: a transcranial magnetic stimulation study in man. Brain Res Cogn Brain Res. 1997;5:185–192. doi: 10.1016/s0926-6410(96)00069-9. [DOI] [PubMed] [Google Scholar]
- Hasbroucq T, Kaneko H, Akamatsu M, Possamaï CA. The time-course of preparatory spinal and cortico-spinal inhibition: an H-reflex and transcranial magnetic stimulation study in man. Exp Brain Res. 1999a;124:33–41. doi: 10.1007/s002210050597. [DOI] [PubMed] [Google Scholar]
- Hasbroucq T, Osman A, Possamaï CA, Burle B, Carron S, Dépy D, Latour S, Mouret I. Cortico-spinal inhibition reflects time but not event preparation: neural mechanisms of preparation dissociated by transcranial magnetic stimulation. Acta Psychol. 1999b;101:243–266. doi: 10.1016/s0001-6918(99)00007-4. [DOI] [PubMed] [Google Scholar]
- Hodges PW, Bui BH. A comparison of computer-based methods for the determination of onset of muscle contraction using electromyography. Electroencephalogr Clin Neurophysiol. 1996;101:511–519. doi: 10.1016/s0013-4694(96)95190-5. [DOI] [PubMed] [Google Scholar]
- Koch G, Franca M, Del Olmo MF, Cheeran B, Milton R, Alvarez Sauco M, Rothwell JC. Time course of functional connectivity between dorsal premotor and contralateral motor cortex during movement selection. J Neurosci. 2006;26:7452–7459. doi: 10.1523/JNEUROSCI.1158-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maertens de Noordhout A, Rothwell JC, Day BL, Dressler D, Nakashima K, Thompson PD, Marsden CD. Effect of digital nerve stimuli on responses to electrical or magnetic stimulation of the human brain. J Physiol. 1992;447:535–548. doi: 10.1113/jphysiol.1992.sp019016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makin TR, Holmes NP, Zohary E. Is that near my hand? Multisensory representation of peripersonal space in human intraparietal sulcus. J Neurosci. 2007;27:731–740. doi: 10.1523/JNEUROSCI.3653-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makin TR, Holmes NP, Ehrsson HH. On the other hand: dummy hands and peripersonal space. Behav Brain Res. 2008;191:1–10. doi: 10.1016/j.bbr.2008.02.041. [DOI] [PubMed] [Google Scholar]
- Maravita A, Spence C, Driver J. Multisensory integration and the body schema: close to hand and within reach. Curr Biol. 2003;13:R531–R539. doi: 10.1016/s0960-9822(03)00449-4. [DOI] [PubMed] [Google Scholar]
- Mars RB, Bestmann S, Rothwell JC, Haggard P. Effects of motor preparation and spatial attention on corticospinal excitability in a delayed-response paradigm. Exp Brain Res. 2007;182:125–129. doi: 10.1007/s00221-007-1055-4. [DOI] [PubMed] [Google Scholar]
- Medendorp WP, Goltz HC, Crawford JD, Vilis T. Integration of target and effector information in human posterior parietal cortex for the planning of action. J Neurophysiol. 2005;93:954–962. doi: 10.1152/jn.00725.2004. [DOI] [PubMed] [Google Scholar]
- O'Shea J, Sebastian C, Boorman ED, Johansen-Berg H, Rushworth MF. Functional specificity of human premotor-motor cortical interactions during action selection. Eur J Neurosci. 2007;26:2085–2095. doi: 10.1111/j.1460-9568.2007.05795.x. [DOI] [PubMed] [Google Scholar]
- O'Shea J, Taylor PC, Rushworth MF. Imaging causal interactions during sensorimotor processing. Cortex. 2008;44:598–608. doi: 10.1016/j.cortex.2007.08.012. [DOI] [PubMed] [Google Scholar]
- Pruszynski JA, Kurtzer I, Scott SH. Rapid motor responses are appropriately tuned to the metrics of a visuo-spatial task. J Neurophysiol. 2008;100:224–238. doi: 10.1152/jn.90262.2008. [DOI] [PubMed] [Google Scholar]
- Prut Y, Fetz EE. Primate spinal interneurons show pre-movement instructed delay activity. Nature. 1999;401:590–594. doi: 10.1038/44145. [DOI] [PubMed] [Google Scholar]
- Quinlan DJ, Culham JC. fMRI reveals a preference for near viewing in the human parieto-occipital cortex. Neuroimage. 2007;36:167–187. doi: 10.1016/j.neuroimage.2007.02.029. [DOI] [PubMed] [Google Scholar]
- Rizzolatti G. Functional organization of inferior area 6. Ciba Found Symp. 1987;132:171–186. doi: 10.1002/9780470513545.ch11. [DOI] [PubMed] [Google Scholar]
- Rizzolatti G, Luppino G, Matelli M. The organization of the cortical motor system: new concepts. Electroencephalogr Clin Neurophysiol. 1998;106:283–296. doi: 10.1016/s0013-4694(98)00022-4. [DOI] [PubMed] [Google Scholar]
- Shimazu H, Maier MA, Cerri G, Kirkwood PA, Lemon RN. Macaque ventral premotor cortex exerts powerful facilitation of motor cortex outputs to upper limb motoneurons. J Neurosci. 2004;24:1200–1211. doi: 10.1523/JNEUROSCI.4731-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svensson P, Miles TS, McKay D, Ridding MC. Suppression of motor evoked potentials in a hand muscle following prolonged painful stimulation. Eur J Pain. 2003;7:55–62. doi: 10.1016/s1090-3801(02)00050-2. [DOI] [PubMed] [Google Scholar]
- Touge T, Taylor JL, Rothwell JC. Reduced excitability of the cortico-spinal system during the warning period of a reaction time task. Electroencephalogr Clin Neurophysiol. 1998;109:489–495. doi: 10.1016/s0924-980x(98)00050-2. [DOI] [PubMed] [Google Scholar]
- Urban PP, Solinski M, Best C, Rolke R, Hopf HC, Dieterich M. Different short-term modulation of cortical motor output to distal and proximal upper-limb muscles during painful sensory nerve stimulation. Muscle Nerve. 2004;29:663–669. doi: 10.1002/mus.20011. [DOI] [PubMed] [Google Scholar]
- van den Hurk P, Mars RB, van Elswijk G, Hegeman J, Pasman JW, Bloem BR, Toni I. Online maintenance of sensory and motor representations: effects on corticospinal excitability. J Neurophysiol. 2007;97:1642–1648. doi: 10.1152/jn.01005.2006. [DOI] [PubMed] [Google Scholar]
- van Elswijk G, Kleine BU, Overeem S, Stegeman DF. Expectancy induces dynamic modulation of corticospinal excitability. J Cogn Neurosci. 2007;19:121–131. doi: 10.1162/jocn.2007.19.1.121. [DOI] [PubMed] [Google Scholar]