

Abstract
Error signals are vital to motor learning. However, we know little about pathways that transmit error signals for learning in voluntary movements. Here we show that microstimulation of the midbrain tegmentum can induce learning in saccadic eye movements in monkeys. Weak electrical stimuli delivered ∼200 ms after saccades in one horizontal direction produced gradual and marked changes in saccade gain. The spatial and temporal characteristics of the produced changes were similar to those of adaptation induced by real visual error. When stimulation was applied after saccades in two different directions, endpoints of these saccades gradually shifted in the same direction in two dimensions. We conclude that microstimulation created powerful learning signals that dictate the direction of adaptive shift in movement endpoints. Our findings suggest that the error signals for saccade adaptation are conveyed in a pathway that courses through the midbrain tegmentum.
Keywords: motor learning, electrical stimulation, midbrain, saccade, adaptation, macaque, monkey
Introduction
Motor learning ensures the accuracy of the movements we execute daily. Vital to learning is the information about the error that results from executed movements. Visual error drives learning in many types of movements, including saccades, voluntary rapid eye movements that shift the direction of gaze from one location in space to another (Hopp and Fuchs, 2004). Learning in saccades, called saccade adaptation, is induced by visual error, an offset between target and eye positions, that is present immediately after saccades (Wallman and Fuchs, 1998; Shafer et al., 2000; Noto and Robinson, 2001; Fujita et al., 2002). The cerebellum plays a central role in saccade adaptation (Barash et al., 1999; Inaba et al., 2003; Scudder and McGee, 2003). In particular, recent studies indicate that the cerebellar oculomotor vermis is at least one of the sites for plasticity (Robinson et al., 2002; Robinson and Noto, 2005). It is unknown, however, what pathway brings the error signal to the cerebellum or how the error is encoded at or upstream of the learning site.
We thought that microstimulation could be a useful tool to probe a pathway that transmits error signals for saccade adaptation (Mauk et al., 1986; Carey et al., 2005). The visual and posterior parietal cortices and the frontal eye field, among other cortical areas, are potential sources of error signals and project to the superior colliculus, a key midbrain structure that issues saccade signals to the premotor reticular circuitry (Scudder et al., 2002). The colliculus also has indirect access to the cerebellum via both the nucleus reticularis tegmenti pontis (Crandall and Keller, 1985; Yamada and Noda, 1987; Noda et al., 1990) and the inferior olive (Frankfurter et al., 1976; Brodal and Brodal, 1981, 1982). In addition, the frontal eye field, supplementary motor area, and motor cortex project to the parvocellular part of the red nucleus in the midbrain, the origin of descending fibers to the inferior olive (Strominger et al., 1979; Stanton et al., 1988; Tokuno et al., 1995; Burman et al., 2000).
As a first step to identify the pathway that transmits the error signal for motor learning in saccades, we investigated the effect of local microstimulation in the medial part of the mesencephalic tegmentum, a region that contains fibers from two candidate structures that instruct the cerebellum, superior colliculus, and parvocellular red nucleus. If a bundle of fibers conveys the error signals, its activation at an appropriate timing after movement is expected to induce adaptation. We report here that timed stimulation of the medial part of the mesencephalic tegmentum induces gradual and marked changes in saccade amplitude and direction. The temporal and spatial characteristics of these changes are similar to those of behavioral adaptation induced by real visual error. Taking advantage of the directional specificity of saccade adaptation, we further show that microstimulation creates signals that are equivalent to the neural correlate of visual error in two-dimensional space.
Materials and Methods
Animals.
Two rhesus monkeys (Macaca mulatta; male; 4.8 and 3.9 kg) were prepared for eye movement recording by the magnetic search coil method (Fuchs and Robinson, 1966). Anesthesia was introduced with ketamine hydrochloride (15–20 mg/kg, i.m.) and maintained by inhalation of isoflurane. A coil of Teflon-coated stainless steel wire was implanted on one eye to measure eye movements. Stainless steel tubes were attached to the skull with bone screws and dental acrylic for painless immobilization of the head. A recording chamber (Crist Instrument, Hagerstown, MD) was implanted over the posterior part of the skull under general anesthesia to gain access to the midbrain. After surgery, antibiotics (500 mg of cefazolin sodium) was given intramuscularly for 3 d to prevent infection. After recovery from the surgery, the monkeys were trained to follow a small jumping target with the eye. During training and experiments, the animal's head was painlessly immobilized by stainless steel rods inserted into the tubes in the acrylic implant. All surgical and experimental protocols were approved by the Animal Care and Use Committee at the University of Tsukuba and by the Animal Care Committee at the Tsukuba Primate Research Center.
Behavioral task.
The monkey sat in a primate chair in a darkened booth with its head restrained during experiment. The animal was required to make saccades toward a target spot (∼0.3°) presented on a monitor screen, which was 35.5 cm from its eyes. Whenever the monkey maintained its eye position within 1.5° of the target continuously for 1.0–1.5 s, the target jumped to another position and the animal was rewarded with a small amount of apple juice. The animal followed the target spot reliably for several thousand trials in a single experiment, which lasted 2–2.5 h. One monkey (monkey I) had been used in other studies and repeatedly subjected to an adaptation paradigm with intrasaccadic target steps. The other monkey (monkey M) was naive to any adaptation paradigm.
Once we set the stimulating electrode at an appropriate position for microstimulation (see below), we first sampled saccades to target steps of three amplitudes (5, 10, and 15°) in each of eight directions (both horizontal and vertical directions and the diagonals) in monkey I as control data for transfer test. We recorded in both monkeys a few hundred saccades to 10° horizontal or vertical target steps (prestimulation session) and then applied microstimulation to the paramedian midbrain after each saccade in either one or both directions (stimulation session). Microstimulation was continued for ∼70 min unless the endpoint shift amounted to ∼5°. In the poststimulation session in monkey I, we again sampled saccades to target steps of the three amplitudes in each of the eight directions (transfer test). In the exploratory tracks at the start of this study, we found that a robust change in saccade vector could be induced without extinguishing the target after saccades. Therefore, in the present study, we used a permanently visible target in the stimulation session.
Recording and microstimulation.
A glass-coated tungsten microelectrode was inserted through a guide tube (23 gauge spinal needle) in the tracks tilted 15° laterally to the left in a frontal plane. In the first several tracks in each monkey, we explored the midbrain by recording single-unit and multiunit activity to locate the oculomotor nuclei, a structure near the midline that we could identify easily by its characteristic burst-tonic multiunit firing (“singing”) associated with vertical saccades. Vertical eye movement-related activity was often followed by leftward or rightward burst-tonic activity, indicating, respectively, the entrance into the right or left medial rectus subdivision. After we were oriented about the location of the nuclei on both sides, we advanced the electrode into a region 0.5–2.5 mm lateral to the oculomotor nucleus and applied microstimulation. Trains of rectangular cathodal pulses (pulse width, 0.2 ms; rate, 400/s; train length, 50 ms) were delivered from a constant current stimulator. Stimulation onset was triggered a set interval after target movement. Small eye movements (<1°) were evoked with short latencies from most stimulation sites probably attributable to current spread to the nearby medial longitudinal fasciculus (MLF) or oculomotor nucleus. Stimulus intensity was usually set slightly higher than the threshold for evoking eye movements and limited below 80 μA in all experiments (range, 25–80 μA; typically 40–50 μA). Because the most effective time window for visual error to induce saccade adaptation with the intrasaccadic step (ISS) paradigm is known to be the first 100–150 ms after saccade end (Shafer et al., 2000), we set the timing of microstimulation slightly later than this, taking into account the time required for error signal transmission from the retina to the midbrain. The time from the target step to the stimulus train were set so that in most of the trials electrical stimuli were delivered ∼200–250 ms after saccade end. The mean delay from saccade end ranged from 147 to 232 ms (n = 47 sessions) in monkey I and 192–300 ms (n = 11sessions) in monkey M. The delay from saccade end varied across saccades because of the variability in saccade reaction time and duration.
Data analysis.
Horizontal and vertical components of eye position and target position were digitized on-line at 1 kHz, and neural activity and stimulus artifact were sampled at 50 kHz and stored in a hard disk with an interface (Micro1401; Cambridge Electronic Design, Cambridge, UK). Data were analyzed off-line on a computer using homemade programs that ran on an analysis software (Spike2; Cambridge Electronic Design). Saccade onset and end were defined by an eye velocity threshold criterion of 20°/s. Visually guided saccades with latencies <60 ms were regarded as anticipatory and not analyzed. Parameters of saccades and target steps, i.e., positions, timings, and peak eye velocities, were exported to statistics programs (JMP; SAS Institute, Cary, NC) to calculate other saccade characteristics. The target eccentricity was defined as the difference between the target and the eye positions at saccade onset. The gain of a saccade to a target step in a specific direction (horizontal or vertical) was defined as the ratio of the saccade size to the target eccentricity in that direction. The saccade endpoint relative to the initial target location was calculated as the difference between eye position at saccade end and initial target position. We simply call it “saccade endpoint” in this paper. The amount of gain change (see Figs. 1c, 5a), shift direction, and shift amplitude (see Figs. 6b, 7c) were estimated by the difference in average between the last 50 saccades in the prestimulation session and the last 50 saccades in stimulation session. The gain change elicited for the first 400 and 950 saccades was obtained by subtracting the average for the last 50 prestimulation saccades from the average for the 376th–425th and 926th–975th saccades in stimulation session, respectively. Immediate changes in the gain at the beginning of stimulation trials were quantified as a difference in average between the last 50 prestimulation saccades and the next 50 saccades. The endpoint shift was defined as a vector that connects the average endpoint for the last 50 saccades in the prestimulation session and that for the last 50 saccades in stimulation session. Numerical data are presented as a mean ± SD in the text.
Figure 1.

Effect of microstimulation coupled with horizontal saccades in one direction. a, A photomicrograph of a frontal section of the midbrain showing a stimulation site (asterisk) in one experiment. Arrows indicate two electrolytic lesions made for reconstruction. The area surrounded by broken line is expanded in Figure 8 to show the extent of effective sites across all experiments. III, Oculomotor nucleus; NRTP, nucleus reticularis tegmenti pontis; Pn, pontine nuclei. b, Sample records collected before (Pre), during (Stim), and after (Post) stimulation session. Black traces in each panel show, from top to bottom, stimulus mark and horizontal and vertical eye positions. A small vertical arrow indicates a tiny eye movement evoked directly by stimulation. Gray traces superimposed on eye positions are target positions. c, Course of gain change. Gain of leftward and rightward saccades is plotted as a function of saccade number. Vertical broken lines indicate start of stimulation session. Stimulation was coupled only with leftward saccades. d, Comparison of gain change in stimulation-coupled and noncoupled saccades. Gain decreases and increases are shown in the left and right panels, respectively. Filled circles connected by solid lines and open circles with broken lines indicate data in monkey I and monkey M, respectively. Horizontal arrows indicate the experiment shown in b and c.
Figure 5.
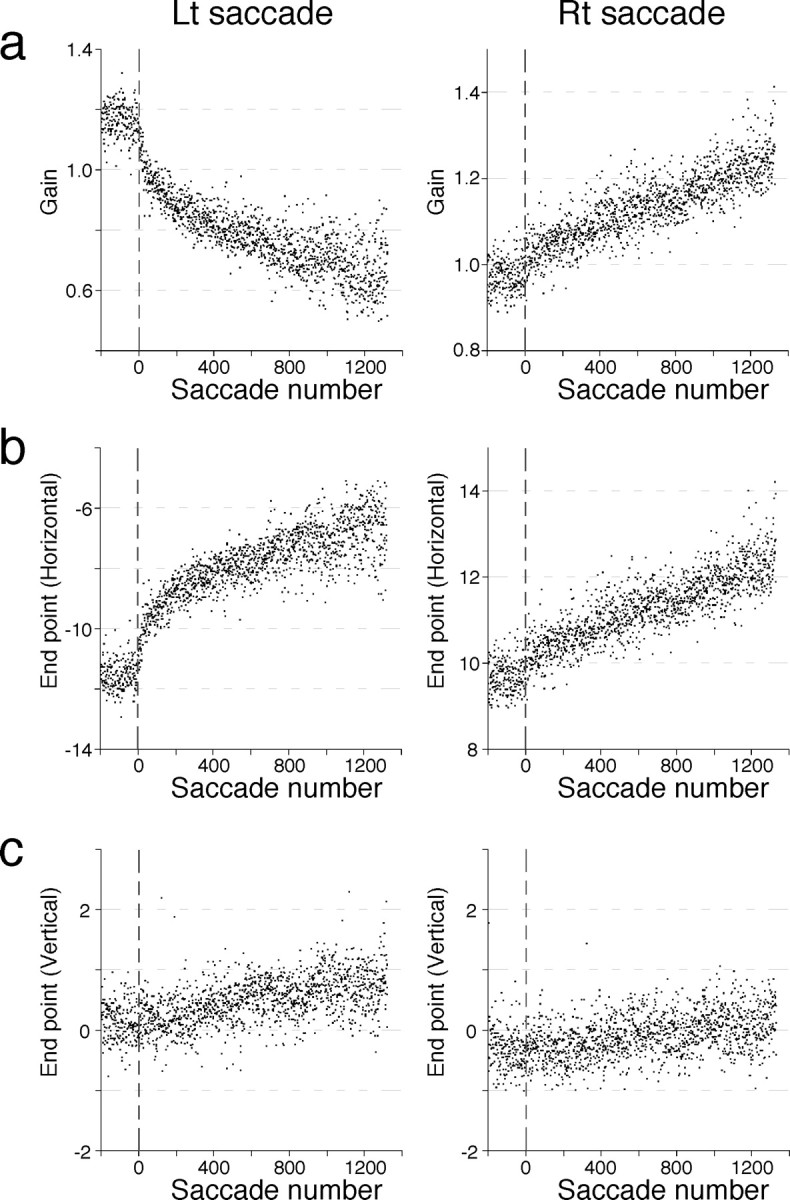
Effect of microstimulation applied shortly after both leftward (Lt) and rightward (Rt) saccades. a, Horizontal gain plotted against saccade number for leftward (left panel) and rightward (right panel) saccades, which show a marked decrease and increase in gain, respectively. b, The horizontal component of movement endpoints (relative to initial target location) of leftward and rightward saccades, both of which exhibit gradual rightward shifts in endpoints. On the ordinate, −10° and 10° indicate target positions to which the animal made leftward and rightward saccades, respectively. c, The vertical component of movement endpoints of leftward and rightward saccades, both of which show small, gradual upward shifts in endpoints.
Figure 6.
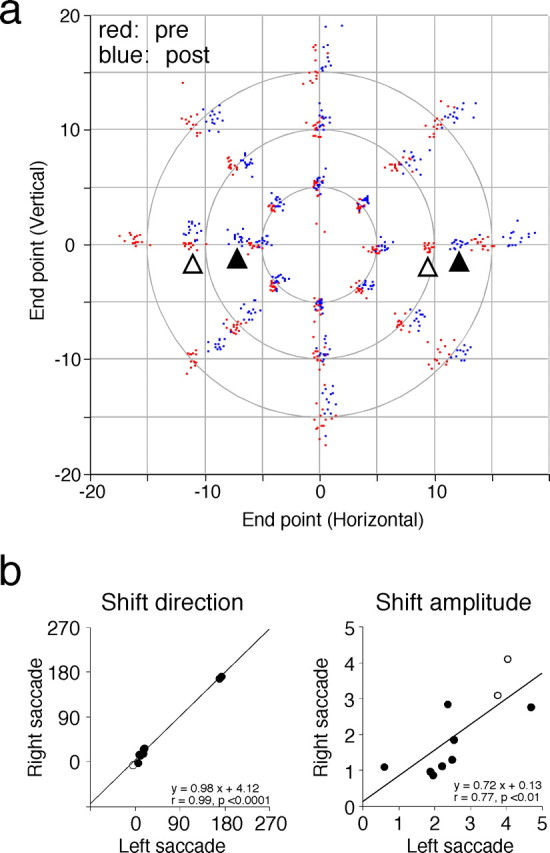
Similar adaptive shifts in endpoints elicited when both leftward and rightward saccades are coupled with microstimulation. a, Transfer of the induced adaptive change to saccades to target steps of various vectors. Data are from the same experiment shown in Figure 5 and shown in the same format as in Figure 2a. Two pairs of open and filled triangles indicate clusters of endpoints in prestimulation (red) and poststimulation (blue) session for saccades to 10° leftward and rightward target steps, respectively. b, Similarity in adaptive shifts in endpoint between leftward and rightward saccades. Summary of 10 experiments. Filled and open circles indicate experiments in monkey I and monkey M, respectively. Left, Shift direction (degrees) for rightward saccades is plotted against that for leftward saccades. For both ordinate and abscissa, 0, 90, 180, and 270 indicate right, up, left, and down, respectively. Right, Shift amplitude (degrees) for rightward saccades is plotted against that for leftward saccades.
Figure 7.
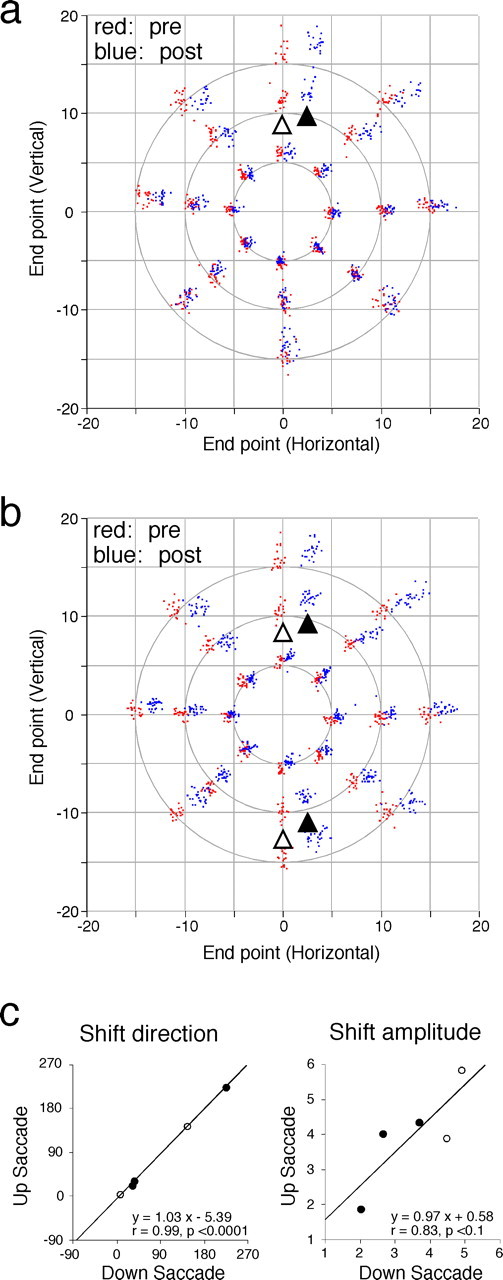
Adaptive shifts in movement endpoint of vertical saccades that are coupled with microstimulation. a, Transfer of the induced adaptive change to saccades to target steps of various vectors when microstimulation followed saccades to 10° upward target steps with a short delay. Shown in the same format as in Figure 2a. Open and filled triangles indicate clusters of endpoints in prestimulation (red) and poststimulation (blue) session for saccades to 10° upward target steps, respectively. b, Results of a similar transfer test when microstimulation followed both saccades to 10° upward and downward target steps with short delays. Two pairs of open and filled triangles indicate endpoint shifts for upward and downward saccades, respectively. c, Similarity in adaptive shifts in endpoint between upward and downward saccades. Summary of five experiments. Filled and open circles indicates experiments in monkey I and monkey M, respectively. Left, Shift direction (degrees) for upward saccades is plotted against that for downward saccades. For both ordinate and abscissa, 0, 90, 180, and 270 indicate right, up, left, and down, respectively. Right, Shift amplitude (degrees) for upward saccades is plotted against that for downward saccades.
Histology.
In the last experiment in monkey I, electrolytic lesions were made along the track 4.0 mm above and 2.0 mm below the microstimulation site by passing direct cathodal current (20–25 μA for 30–40 s). The dorsal lesion was made at the dorsal border of the right oculomotor nuclei as judged by recorded unit activity (see Fig. 1a, top arrow; also shown in Fig. 8). Later on the same day, the animal was killed with an overdose of pentobarbital and perfused intracardially first with 2 L of saline and then with 3 L of 4% paraformaldehyde. The brain was removed, postfixed for several days, and cut into serial frontal sections (100 μm thick). The sections were mounted on slide glasses, dehydrated, stained with cresyl violet, and coverslipped for histological examination. Locations of stimulation sites were estimated with reference to the marking lesions on the basis of the rostrocaudal and mediolateral coordinates of the track and the depth reading.
Figure 8.

Location of effective stimulation sites and summary of adaptive shifts produced from these sites, drawn on a photomicrograph of a frontal section from monkey I. Filled circles indicate the reconstructed stimulation sites. Arrows indicate shift vectors, i.e., the direction and amplitude of elicited adaptive shifts in saccade endpoint. III, Oculomotor nucleus.
Results
We investigated in two monkeys (monkeys I and M) whether microstimulation of the midbrain tegmentum could mimic error signals and produce changes in saccade vector that are similar to adaptation induced by real visual error, e.g., the error created by stepping the target during saccades (Hopp and Fuchs, 2004). Most of the stimulation sites were located ventrolateral to the oculomotor nuclei, between the MLF and the decussation of the brachium conjunctivum (BCX) (see Figs. 1a, 8).
Gain changes induced by microstimulation of the midbrain tegmentum
In the first series of experiments, we required the monkey to make saccades to horizontal target steps and examined the effect of microstimulation applied in conjunction with either rightward or leftward saccades. Figure 1a–c shows the result of one such experiment. The stimulation site was located ∼2.0 mm ventrolateral to the oculomotor nucleus on the right side (Fig. 1a). Figure 1b illustrates sample raw eye-movement records obtained from prestimulation, stimulation, and poststimulation trials. Before microstimulation, the amplitude of saccades to 10° leftward target steps was ∼10° (Fig. 1b, left). During stimulation trials, an electrical pulse train (40 μA for 20 times, 400 Hz, 50 ms) was applied shortly after every leftward saccade. Target movement triggered stimulation. Therefore, the delay between saccade end and stimulation onset was variable because of variability in the animal's reaction time and saccade duration. Here we adjusted the stimulation delay after target movement so that the average lag from saccade end was 189 ± 49 ms (957 saccades). Note that microstimulation evoked only a tiny eye movement (arrow). This movement was probably the result of minor current spread to the nearby MLF or oculomotor nucleus. Over the course of hundreds of trials, saccades to leftward target steps gradually decreased their amplitude and fell short of the target. It is noteworthy that microstimulation did not prevent the monkey from making leftward corrective saccades to bring the eye on target. The reduction in amplitude was still evident after microstimulation was discontinued (Fig. 1b, right). To compare the effect of microstimulation on leftward and rightward saccades, we plotted the gain of individual saccades as a function of the number of saccades in each direction. As Figure 1c shows, the gain of leftward saccades, which were associated with microstimulation, progressively decreased with an approximately exponential course, eventually changing from 1.024 ± 0.040 (last 50 prestimulation saccades) to 0.586 ± 0.099 (last 50 saccades). In contrast, the gain of rightward saccades, which were not followed by microstimulation, was 1.155 ± 0.058 (last 50 prestimulation) and 1.177 ± 0.047 (last 50 saccades), remaining relatively constant for most of the stimulation trials after a small transient increase near the start of stimulation (1.243 ± 0.069, first 50 saccades).
Figure 1d summarizes the results of 14 experiments in which saccades in one horizontal direction were coupled with microstimulation (30–60 μA) and exhibited clear changes in horizontal gain (> 0.1). Both decreases and increases in gain were produced in each monkey depending on the stimulation site and the direction of saccades (see below). The magnitude of gain decrease for stimulation-coupled saccades (0.107–0.438, average of 0.297) was significantly larger than that for noncoupled saccades (−0.158 to 0.012, average of −0.061; p < 0.05; n = 8; Wilcoxon's signed rank test). Similarly, the gain increase for stimulation-coupled saccades (0.210–0.363, average of 0.270) was significantly larger than that for noncoupled saccades (−0.029 to 0.113, average of 0.037; p < 0.05; n = 6). Thus, microstimulation resulted in progressive gain changes for saccades that were coupled with it. The gain changes for noncoupled saccades were much smaller, mostly <0.1. In all of the 14 experiments, the horizontal gain of stimulation-coupled saccades changed gradually.
The prestimulation gain of stimulation-coupled and noncoupled saccades were substantially different (by >0.1) from unity in four and eight experiments, respectively (e.g., leftward saccades in Fig. 5 and rightward saccades in Fig. 1c). This was attributable to incomplete recovery from the gain change induced in the preceding experiment. In addition, an immediate gain change at the start of stimulation trials (as in Fig. 1c) was observed for saccades not coupled with microstimulation in 12 of 14 experiments (p < 0.05), ranging from −0.147 to 0.120 (negative indicates decrease). Their absolute value had a mean of 0.073 ± 0.037 (n = 12). We currently do not have a plausible explanation for this initial, jump-like change in gain.
Behavioral adaptation is relatively specific to the vector of adapted saccades (Straube et al., 1997; Noto et al., 1999). To examine the spatial specificity of microstimulation-induced changes, we recorded saccades to target movements of various vectors before and after stimulation session in 8 of the 14 experiments. Figure 2a shows the distribution of endpoints of prestimulation (red) and poststimulation (blue) saccades (same experiment as in Fig. 1a–c). Endpoints of saccades to 10° leftward target steps (open triangle) shifted by ∼3° in the rightward direction (filled triangle). Saccades to 15° leftward target steps showed a larger shift and a comparable gain change, whereas saccades to 5° target steps showed a smaller gain change. This pattern of more gain change transfer to larger saccades than to smaller saccades is characteristic of adaptation induced by real visual error (Noto et al., 1999). Figure 2a also shows that the amount of endpoint shift decreases as the saccade direction moves away from the “adapted” direction. Similar patterns of transfer were observed in seven other experiments, as shown in the summary plot of percentage transfer for saccade amplitude (Fig. 2b) and saccade direction (Fig. 2c).
Figure 2.

Spatial characteristics of microstimulation-induced adaptive changes. a, Transfer of the induced change to saccades to target steps of various vectors. Data are from the same experiment shown in Figure 1, b and c. Saccade endpoints relative to initial target location before (red) and after (blue) stimulation session are shown in two dimensions. Open and filled triangles indicate clusters of endpoints in prestimulation and poststimulation session for saccades to 10° leftward target steps. b, Summary for eight experiments (monkey I). Relative amplitude of endpoint shift is shown for saccades to 5, 10, and 15° target movements in the adapted direction. c, Similar plot of relative shift amplitude for saccades to 10° target movements in five directions. Abscissa indicates directional difference from the adapted direction. Positive indicates clockwise deviation.
Saccade adaptation induced by visual error is known to have a gradual, approximately exponential course, its magnitude and rate being highly variable (Straube et al., 1997; Scudder et al., 1998; Robinson et al., 2003). We quantified the size of gain changes elicited in stimulation-coupled saccades in the two monkeys. The total gain changes observed in individual experiments had a mean of −0.295 ± 0.111 (range of −0.106 to −0.438; n = 8) and 0.267 ± 0.062 (range of 0.210–0.367; n = 6) for gain decreases and increases, respectively. The total number of stimulation-coupled saccades ranged from 345 to 1419 (885 ± 289; n = 14). The gain was clearly far from reaching the steady state in the majority of experiments. Therefore, it was difficult to estimate reliable rate constants using exponential fit. Instead, we calculated the gain changes that occurred for the first 400 and 950 stimulation-coupled saccades. The gain change elicited for the 400 saccades was −0.217 ± 0.133 for gain decrease (n = 7) and 0.142 ± 0.102 for gain increase (n = 6). The gain change achieved at the 950th saccade was −0.269 ± 0.163 (n = 3) and 0.168 ± 0.040 (n = 3) for gain decreases and increases, respectively. Although precise comparison of the rate constant is not possible, the magnitude of these microstimulation-induced gain changes seems similar to that reported for behavioral adaptation that was elicited by stepping the target during saccades (ISS) (Straube et al., 1997; Scudder et al., 1998; Robinson et al., 2003). We also compared the gain changes observed in the present study in monkey I to those elicited by 35% ISS in the same monkey in our previous study (Kojima et al., 2004). The stimulation-induced gain decrease for the first 400 saccades (absolute value) had a mean of 0.288 ± 0.019 (n = 4, monkey I). This was significantly larger than the ISS-induced gain decrease for 400 saccades, 0.140 ± 0.038 (n = 9, monkey I; p < 0.01). The stimulation-elicited gain increase for 400 saccades, 0.149 ± 0.132 (n = 4, monkey I), was not significantly different from the ISS-induced gain increase for 400 saccades, 0.116 ± 0.034 (n = 5, monkey I; p = 0.60). Given the huge variability in the rate of saccade adaptation across experiments (Straube et al., 1997; Robinson et al., 2003), it seems difficult to conclude that microstimulation elicits faster gain changes than the visual error does.
We next asked how stimulation timing affected the induction of the gradual, progressive changes in saccade gain. Figure 3 shows the result of one experiment in which we tested two different timings. When microstimulation (30 μA) was applied 1055 ± 75 ms after rightward saccades, their horizontal gain did not change (∼1000 ms lag). The gain of the last 50 (339th–388th) saccades, 1.120 ± 0.068, was not significantly different from that of the first 50 saccades, 1.126 ± 0.053. When the same stimulus train was applied 222 ± 55 ms after rightward saccades, their gain started to increase and continued to do so over the next several hundred saccades (∼200 ms lag). The gain of 339th–388th saccades in the 200 ms lag period, 1.261 ± 0.057, was significantly larger than that of the first 50 saccades in this period, 1.128 ± 0.055. We obtained similar results in another experiment, changing the timings from 1006 ± 175 to 241 ± 49 ms. The results suggest that timing of microstimulation is crucial to induce adaptive gain changes, 1.0 s after saccades being too late to exert consistent effects on saccade vector.
Figure 3.

Effect of stimulation timing on the induction of adaptation-like gain changes. The gain of stimulation-coupled horizontal saccades is plotted against saccade number. The gain did not change when microstimulation was given with an ∼1000 ms lag. When the lag was reduced to ∼200 ms, the gain began to show progressive increase that lasted throughout the stimulation session.
At nearly all stimulation sites that were effective for inducing progressive changes in saccade gain, small eye movements were evoked at a relatively low current (30–60 μA) with latencies of 7–13 ms. These movements were probably caused by the current spread to the MLF or oculomotor nucleus. The size of eye movements evoked directly by microstimulation (typically <1°) did not change over the course of stimulation session. The direction or size of these evoked movements were not related to whether or how the gain of stimulation-coupled saccades changed over many trials. Furthermore, in one experiment, we set the stimulus current subthreshold for evoking eye movements (Fig. 4, left). When leftward saccades were coupled with this stimulation, their gain decreased gradually to ∼0.6 (Fig. 4, right). This result confirms that directly evoked eye movements are not necessary in producing adaptation-like gain changes.
Figure 4.

Adaptation-like gain changes induced by microstimulation that did not directly evoke eye movements. Left, Traces indicate, top to bottom, eye velocity, stimulation mark, and horizontal and vertical eye position. Gray traces that accompany eye position traces indicate target position. Right, Saccade gain plotted against saccade number. Leftward saccades, which were coupled with this subthreshold stimulation (30 μA), exhibited a marked gain decrease over ∼650 saccades. Rt, Right; Lt, left.
Microstimulation generates signals representing the direction of error in two dimensions
The results described so far indicate that microstimulation in the medial midbrain tegmentum generated an instructive signal that dictates the adaptive change in saccades. We hypothesized that the signal created by microstimulation mimics neural correlate of visual error that specifies the direction of progressive shifts in endpoints of stimulation-coupled saccades. In the second series of experiments, we tested this hypothesis by applying microstimulation after both leftward and rightward saccades during a single stimulation session. Figure 5 shows the result of one experiment. Weak electrical currents (25 μA) were delivered in the midbrain tegmentum on the right side, the estimated stimulation site being ∼1.0 mm dorsolateral to that in Figure 1a. As shown in Figure 5a, the gain of leftward saccades gradually decreased to ∼0.7, whereas that of rightward saccades increased to ∼1.25. Microstimulation thus exerted opposite effects on the gains of leftward and rightward saccades. This is the result of rightward shifts in the endpoints of both leftward and rightward saccades (Fig. 5b). There was also a smaller but significant upward shifts in endpoints for saccades in both directions (Fig. 5c). The start points of saccades did not shift significantly (data not shown). Thus, microstimulation produced gradual shifts in endpoints in the same direction (right-up) for both rightward and leftward saccades. This is consistent with our hypothesis that microstimulation generates signals equivalent to the neural correlate of visual error.
Figure 6a shows in two dimensions how saccades of various vectors changed their endpoints after saccades to 10° rightward and leftward target steps were repeatedly coupled with microstimulation (same experiment as in Fig. 5). The endpoints of the stimulation-coupled saccades shifted rightward and slightly upward after ∼1300 saccades (pairs of open and filled triangles). These “adaptive” shifts transferred to larger and smaller horizontal saccades in a similar manner as adaptation induced by a behavioral paradigm. Again, the magnitude of endpoint shifts was progressively reduced as the directional difference increased.
Figure 6b summarizes 10 experiments in which both rightward and leftward saccades were coupled with microstimulation. We regarded the elicited shift in saccade endpoint as a two-dimensional vector and calculated its direction and amplitude. The shift vector direction of rightward saccades is plotted against that of leftward saccades (Fig. 6b, left). The data points are situated close to the line, y = x, indicating that endpoints of rightward and leftward saccades shifted in the same direction. In addition, there is a significant positive correlation between the amplitude of shift vectors of rightward saccades and that of leftward saccades (Fig. 6b, right).
In a third set of experiments, we further examined the effect of coupling vertical saccades with microstimulation, requiring the monkey to follow a target jumping along the vertical meridian by 10°. In the experiment shown in Figure 7a, upward saccades were coupled with microstimulation applied to a site close to that in Figure 1a. Clear changes in endpoints were seen for upward but not for downward saccades. The endpoint of upward, stimulation-coupled saccades shifted in the right-up direction (open and filled triangles). Again, the adaptive shift transferred to saccades of different vectors in the same manner as behavioral adaptation; progressively less transfer as the saccade vector departed from that of adapted saccades. In two other experiments, we coupled upward saccades with microstimulation and obtained similar results.
In contrast, when we next coupled both upward and downward saccades with microstimulation, the endpoints of both changed. Figure 7b shows the results of one experiment. Repeated coupling in each direction resulted in both upward and downward saccades shifting their endpoints in similar directions. Figure 7c summarizes results of five experiments. As was the case with horizontal saccades, the shift vector directions were similar for saccades in the two vertical directions (Fig. 7c, left). The shift amplitude of upward saccades tended to vary with that of downward saccades (Fig. 7c, right).
The data for horizontal and vertical saccades indicate that the shift vector direction is determined by the stimulation site independent of the direction of stimulation-coupled saccades. Furthermore, the induced shifts in endpoints of rightward, leftward, and upward saccades had similar directions when they were coupled with microstimulation at the identical site in single experiments. These results strongly suggest that microstimulation created signals equivalent to two-dimensional visual error signals.
Location of effective stimulation sites
We intended to stimulate the medial part of the tegmentum at the rostrocaudal level of the oculomotor nuclei. In monkey I, microstimulation sites were estimated with reference to the marking lesions and drawn on a photomicrograph of a frontal section. Figure 8, which is an expansion of the area bordered by the broken line in Figure 1a, shows the reconstructed sites of stimulation (filled circles). An arrow originating from a filled circle represents the size and direction of the gradual shift in endpoint produced by stimulation of that site. Most of the sites (33 of 41) that were tested for inducing adaptation were located in this transverse plane. Histological examination of serial sections revealed that the plane was ∼1.5 mm rostral to the caudal edge of the oculomotor nuclei, close to the caudal edge of the magnocellular red nucleus.
Most stimulation sites were located in the area that was bordered medially by the MLF, ventrally by the traversing fibers of the BCX and laterally by dorsolaterally coursing fibers from the BCX and by the magnocellular red nucleus. Sizable shifts in saccade endpoints (>1°) were induced from 20 sites in monkey I. The shift amplitude ranged from 1.1 to 4.7° (mean of 2.7 ± 1.2). Many stimulation sites were found to induce large, progressive shifts in endpoints of stimulation-coupled saccades. The majority of the endpoint shift vectors had an ipsilateral component along with varying sizes of vertical component. Some stimulation sites had shift vectors with a contralateral component, e.g., the five sites shown on the left side of the midbrain. Dorsal and lateral sites may tend to elicit contralaterally directed endpoint shifts. Microstimulation in sites outside the boundary of Figure 8, in the periaqueductal gray, ∼4 mm lateral of the oculomotor nuclei and immediately ventral to the BCX, induced only small, often nonsignificant, shifts in endpoints (0.1–0.7°). Two other sites in planes 1.0 mm caudal and 0.5 mm rostral to the plane in Figure 8, respectively, were found to induce shifts (>1°) in the left and left-up directions. Both were located on the left side and 1–2 mm lateral to the midline. Note that these reconstructed locations may have some estimation error, particularly in the depth along the electrode track.
In the other animal (monkey M), sizable shifts in endpoint (>1°) were induced from five sites. Four of these were in the right midbrain and located 1–2 mm ventrolateral to the oculomotor nuclei. The remaining one site was estimated to be in a comparable region of the left midbrain. The amplitude of the shift vectors ranged between 1.1 and 5.4° (mean of 3.4 ± 1.6). The direction of the shift vectors was either nearly ipsilateral or ipsilateral-upward. Furthermore, the gradual shift in saccade endpoint was induced in this monkey from a region that extended 3 mm rostrocaudally. This suggests that the stimulated neural element responsible for eliciting adaptation-like changes may be passing fibers rather than cell bodies. In fact, we encountered few units near most stimulation sites during recording that preceded microstimulation in each experiment.
To summarize, the induced gradual shift in saccade endpoint at many tested sites had an ipsiversive component with a varying degree of vertical component. Sites located more laterally and dorsally tended to have shift vectors directed contralaterally. The direction of the shift vectors ranged from down-horizontal to up-horizontal and appeared to be dependent on the stimulation site. These results suggest that visual error may be encoded in the activity of descending fibers in the medial part of the mesencephalic tegmentum. Different error signals seem to be represented in different parts of the fiber tract.
Discussion
Microstimulation induces motor learning by generating error signals
The major finding of this work is that repetitive coupling of saccades with microstimulation of the midbrain tegmentum results in marked changes in movement vector. Three characteristics of these changes resemble those of adaptation induced by real visual error such as the error created by stepping the target during saccades (Hopp and Fuchs, 2004). First, the gain of microstimulation-coupled saccades exhibited a gradual course over more than 1000 saccades, and the magnitude of the gain change was similar to that reported previously for behavioral adaptation (Straube et al., 1997; Scudder et al., 1998; Robinson et al., 2003). Second, examining the transfer to saccades of different vectors indicated that the produced change was about as specific to the vector of stimulation-coupled saccades as behaviorally adapted saccades are (Noto et al., 1999). Third, microstimulation was effective when applied with an ∼150–300 ms delay but caused no change in saccade endpoint when given with an ∼1000 ms delay. The presence of an effective period shortly after saccades is a temporal property of the visual error that induces adaptation (Shafer et al., 2000; Fujita et al., 2002). These similarities suggest that microstimulation created instructive signals for motor learning in saccades.
To our knowledge, only two studies have reported that electrical stimulation of some relevant structure generates signals that instruct motor learning; the inferior olive was activated for eye blink conditioning (Mauk et al., 1986) and the middle temporal visual cortical area for smooth pursuit adaptation (Carey et al., 2005). In the present study, we took advantage of directional specificity of saccade adaptation to further investigate the nature of “instruction” and asked whether the activity created by microstimulation is equivalent to the neural correlate of visual error, the key sensory stimulus for adaptation. When saccades to horizontal target steps in one direction were coupled with microstimulation, the induced adaptive changes could be viewed as alterations in gain. It might be argued that the produced signals instructed the saccadic system to increase or decrease the gain. However, when both rightward and leftward saccades were followed by the same electrical stimuli, we observed a gain increase for saccades in one direction and a concomitant gain decrease for saccades in the opposite direction. This finding clearly indicates that the instruction is not “modify the gain per se.” Rather, the generated signals seemed to dictate the direction of the shift in saccade endpoints that was about to be induced. This was confirmed by the finding that endpoints of upward and downward saccades, when both were coupled with microstimulation, shifted gradually in similar directions. In addition, endpoints of horizontal and vertical saccades exhibited adaptive shifts in similar directions when they were coupled with microstimulation at the identical site in single experiments.
Interactions of electrical stimulation with concurrent visual error
As described in Materials and Methods, we made the target visible after saccades in the present study. Endpoints of saccades coupled with microstimulation therefore progressively deviated from the target, creating a visual error after movements. Such errors could have resulted in adaptation that opposed and potentially nullified the stimulation-induced gradual changes. However, the saccade endpoint continued to shift in a fixed direction often >3° despite the presence of the consistent and ever-growing visual error. This result indicates that stimulation created powerful instructive signals for adaptation. We speculate that the effect of the real visual error was masked by that of microstimulation-induced error signals or the transmission of real error signals was impaired by electrical stimulation.
Interaction of microstimulation with visual error also provides a clue about the site of stimulation in the saccade adaptation circuitry. In normal situations, the error signal for adaptation and the signal for a corrective saccade are both derived from the same visual error on the retina. The two signals share a common path at least in early visual stages but are expected to later be carried by two distinct pathways. One is a well known path for saccade generation (Scudder et al., 2002) and the other is a path dedicated solely to the transmission of learning signals. Our data seem to suggest that we applied microstimulation somewhere in the error signal pathway downstream of the branching point. Although the postsaccadic visual error did not seem to produce clear behavioral adaptation, it did induce corrective saccades that brought the eye onto the target (Figs. 1b, 4). Thus, the real visual error, which apparently failed to counteract stimulation-induced learning, could still elicit the normal corrective movements. Put another way, microstimulation did not elicit “wrong” corrective saccades that would have brought the eye farther away from the target. Such wrong movements would have been expected if we had stimulated upstream of the branching point, creating the signals from which both corrective motor command and learning signals were formed.
Possible structures that transmit error signals for saccade adaptation
The current study suggests that error signals for saccade adaptation are conveyed by neural elements in the medial part of the mesencephalic tegmentum. Because we were able to explore the midbrain tegmentum only to a limited extent mediolaterally and rostrocaudally, it is difficult to specify a structure that was responsible for inducing adaptation-like changes in saccades. Nevertheless, at least three descending systems can be suggested as candidates on the basis of anatomical data. The first two originate from the superior colliculus: the efferent fibers that target the nucleus reticularis tegmenti pontis, a major precerebellar nucleus that projects to the cerebellar oculomotor vermis (Yamada and Noda, 1987) and the collicular output fibers that project to the medial accessory nucleus of the inferior olive (MAO) (Frankfurter et al., 1976), which in turn projects to the oculomotor vermis (Brodal and Brodal, 1981, 1982). The descending fibers from the colliculus, which probably include tectoreticular and tectoolivary fibers, course ventromedially outside the MLF before descending in the contralateral predorsal bundle (Frankfurter et al., 1976; Moschovakis et al., 1988). A third descending tract that could be responsible for the observed changes in saccades originates from the parvocellular part of the red nucleus and terminates in the MAO (Strominger et al., 1979; Burman et al., 2000). The rubroolivary fibers descend in the region lateral to the MLF at the level of the oculomotor nucleus. Many of the effective stimulation sites found in our study appear to correspond to the locations of these descending tracts. There is a possibility, however, that artificial activation of neural elements other than these tracts may have resulted in modification of saccade vectors by directly or indirectly producing learning signals. Also, our results do not necessarily indicate that the pathway stimulated here is the one that normally conveys the learning signal. Additional studies are needed to identify the structure responsible for the observed effects.
To date, two studies have examined inferior olive signals during saccade adaptation by recording complex spikes from Purkinje cells in the cerebellar vermis. One of the studies shows that climbing fiber response encodes the direction of visual error and thus supports the olivary relay of error signals (Soetedjo and Fuchs, 2006). The other study shows that the pattern of climbing fiber activity is related to the error signal as well as to the stabilization of adaptation (Catz et al., 2005). Thus, the results of both studies make the tectoolivary and rubroolivary tracts likely candidates for the error signal pathways. These tracts have also been assumed to transmit the learning signal in a recent model of saccade adaptation (Fujita, 2005).
We conclude that microstimulation of the medial part of the midbrain tegmentum creates powerful instructive signals that guide learning in saccades. Our results provide a basis for future studies to identify the pathway that brings the error signals to the adaptation site.
Footnotes
This work was supported by the 21st Century COE program (Promotion of Kansei Science) and Grant-in-Aid 16300129 (K.Y., Y.I.) from the Japanese government. Y. Kojima is a Research Fellow of the Japan Society for the Promotion of Science (16.12119). We are grateful to the veterinary staff at the Tsukuba Primate Center for their expert help in surgery and daily care of the animals, Kozo Kobayashi for building a primate chair, Akiko Ohgami for histological work, and Albert Fuchs at the University of Washington National Primate Research Center for providing target display programs.
References
- Barash S, Melikyan A, Sivakov A, Zhang M, Glickstein M, Thier P. Saccadic dysmetria and adaptation after lesions of the cerebellar cortex. J Neurosci. 1999;19:10931–10939. doi: 10.1523/JNEUROSCI.19-24-10931.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodal P, Brodal A. The olivocerebellar projection in the monkey. Experimental studies with the method of retrograde tracing of horseradish peroxidase. J Comp Neurol. 1981;201:375–393. doi: 10.1002/cne.902010306. [DOI] [PubMed] [Google Scholar]
- Brodal P, Brodal A. Further observations on the olivocerebellar projection in the monkey. Exp Brain Res. 1982;45:71–83. doi: 10.1007/BF00235764. [DOI] [PubMed] [Google Scholar]
- Burman K, Darian-Smith C, Darian-Smith I. Macaque red nucleus: origins of spinal and olivary projections and terminations of cortical inputs. J Comp Neurol. 2000;423:179–196. doi: 10.1002/1096-9861(20000724)423:2<179::aid-cne1>3.0.co;2-#. [DOI] [PubMed] [Google Scholar]
- Carey MR, Medina JF, Lisberger SG. Instructive signals for motor learning from visual cortical area MT. Nat Neurosci. 2005;8:813–819. doi: 10.1038/nn1470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Catz N, Dicke PW, Thier P. Cerebellar complex spike firing is suitable to induce as well as to stabilize motor learning. Curr Biol. 2005;15:2179–2189. doi: 10.1016/j.cub.2005.11.037. [DOI] [PubMed] [Google Scholar]
- Crandall WF, Keller EL. Visual and oculomotor signals in nucleus reticularis tegmenti pontis in alert monkey. J Neurophysiol. 1985;54:1326–1345. doi: 10.1152/jn.1985.54.5.1326. [DOI] [PubMed] [Google Scholar]
- Frankfurter A, Weber JT, Royce GJ, Strominger NL, Harting K. An autoradiographic analysis of the tecto-olivary projection in primates. Brain Res. 1976;118:245–257. doi: 10.1016/0006-8993(76)90710-1. [DOI] [PubMed] [Google Scholar]
- Fuchs AF, Robinson DA. A method for measuring horizontal and vertical eye movement chronically in the monkey. J Appl Physiol. 1966;21:1068–1070. doi: 10.1152/jappl.1966.21.3.1068. [DOI] [PubMed] [Google Scholar]
- Fujita M. Feed-forward associative learning for volitional movement control. Neurosci Res. 2005;52:153–165. doi: 10.1016/j.neures.2005.02.006. [DOI] [PubMed] [Google Scholar]
- Fujita M, Amagai A, Minakawa F, Aoki M. Selective and delay adaptation of human saccades. Brain Res Cogn Brain Res. 2002;13:41–52. doi: 10.1016/s0926-6410(01)00088-x. [DOI] [PubMed] [Google Scholar]
- Hopp JJ, Fuchs AF. The characteristics and neuronal substrate of saccadic eye movement plasticity. Prog Neurobiol. 2004;72:27–53. doi: 10.1016/j.pneurobio.2003.12.002. [DOI] [PubMed] [Google Scholar]
- Inaba N, Iwamoto Y, Yoshida K. Changes in cerebellar fastigial burst activity related to saccadic gain adaptation in the monkey. Neurosci Res. 2003;46:359–368. doi: 10.1016/s0168-0102(03)00098-1. [DOI] [PubMed] [Google Scholar]
- Kojima Y, Iwamoto Y, Yoshida K. Memory of learning facilitates saccadic adaptation in the monkey. J Neurosci. 2004;24:7531–7539. doi: 10.1523/JNEUROSCI.1741-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mauk MD, Steinmetz JE, Thompson RF. Classical conditioning using stimulation of the inferior olive as the unconditioned stimulus. Proc Natl Acad Sci USA. 1986;83:5349–5353. doi: 10.1073/pnas.83.14.5349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moschovakis AK, Karabelas AB, Highstein SM. Structure-function relationship in the primate superior colliculus. I. Morphological classification of efferent neurons. J Neurophysiol. 1988;60:232–262. doi: 10.1152/jn.1988.60.1.232. [DOI] [PubMed] [Google Scholar]
- Noda H, Sugita S, Ikeda Y. Afferent and efferent connections of the oculomotor region of the fastigial nucleus in the macaque monkey. J Comp Neurol. 1990;302:330–348. doi: 10.1002/cne.903020211. [DOI] [PubMed] [Google Scholar]
- Noto CT, Robinson FR. Visual error is the stimulus for saccade gain adaptation. Brain Res Cogn Brain Res. 2001;12:301–305. doi: 10.1016/s0926-6410(01)00062-3. [DOI] [PubMed] [Google Scholar]
- Noto CT, Watanabe S, Fuchs AF. Characteristics of simian adaptation fields produced by behavioral changes in saccade size and direction. J Neurophysiol. 1999;81:2798–2813. doi: 10.1152/jn.1999.81.6.2798. [DOI] [PubMed] [Google Scholar]
- Robinson FR, Noto CT. Role of the cerebellar oculomotor vermis in saccade adaptation. Soc Neurosci Abstr. 2005;31:986–984. [Google Scholar]
- Robinson FR, Fuchs AF, Noto CT. Cerebellar influences on saccade plasticity. Ann NY Acad Sci. 2002;956:155–163. doi: 10.1111/j.1749-6632.2002.tb02816.x. [DOI] [PubMed] [Google Scholar]
- Robinson FR, Noto CT, Bevans SE. Effect of visual error size on saccade adaptation in monkey. J Neurophysiol. 2003;90:1235–1244. doi: 10.1152/jn.00656.2002. [DOI] [PubMed] [Google Scholar]
- Scudder CA, Batourina EY, Tunder GS. Comparison of two methods of producing adaptation of saccade size and implications for the site of plasticity. J Neurophysiol. 1998;79:704–715. doi: 10.1152/jn.1998.79.2.704. [DOI] [PubMed] [Google Scholar]
- Scudder CA, Kaneko CS, Fuchs AF. The brainstem burst generator for saccadic eye movements: a modern synthesis. Exp Brain Res. 2002;142:439–462. doi: 10.1007/s00221-001-0912-9. [DOI] [PubMed] [Google Scholar]
- Shafer JL, Noto CT, Fuchs AF. Temporal characteristics of error signals driving saccadic gain adaptation in the macaque monkey. J Neurophysiol. 2000;84:88–95. doi: 10.1152/jn.2000.84.1.88. [DOI] [PubMed] [Google Scholar]
- Soetedjo R, Fuchs AF. Complex spike activity of Purkinje cells in the oculomotor vermis during behavioral adaptation of monkey saccades. J Neurosci. 2006;26:7741–7755. doi: 10.1523/JNEUROSCI.4658-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanton GB, Goldberg ME, Bruce CJ. Frontal eye field efferents in the macaque monkey. II. Topography of terminal fields in midbrain and pons. J Comp Neurol. 1988;271:493–506. doi: 10.1002/cne.902710403. [DOI] [PubMed] [Google Scholar]
- Straube A, Fuchs AF, Usher S, Robinson FR. Characteristics of saccadic gain adaptation in rhesus macaques. J Neurophysiol. 1997;77:874–895. doi: 10.1152/jn.1997.77.2.874. [DOI] [PubMed] [Google Scholar]
- Strominger NL, Truscott TC, Miller RA, Royce GJ. An autoradiographic study of the rubroolivary tract in the rhesus monkey. J Comp Neurol. 1979;183:33–45. doi: 10.1002/cne.901830104. [DOI] [PubMed] [Google Scholar]
- Tokuno H, Takada M, Nambu A, Inase M. Somatotopical projections from the supplementary motor area to the red nucleus in the macaque monkey. Exp Brain Res. 1995;106:351–355. doi: 10.1007/BF00241130. [DOI] [PubMed] [Google Scholar]
- Wallman J, Fuchs AF. Saccadic gain modification: visual error drives motor adaptation. J Neurophysiol. 1998;80:2405–2416. doi: 10.1152/jn.1998.80.5.2405. [DOI] [PubMed] [Google Scholar]
- Yamada J, Noda H. Afferent and efferent connections of the oculomotor cerebellar vermis in the macaque monkey. J Comp Neurol. 1987;265:224–241. doi: 10.1002/cne.902650207. [DOI] [PubMed] [Google Scholar]