

Abstract
Psychostimulants, including amphetamine, act as antihyperkinetic agents in humans with hyperkinetic disorder such as attention-deficit hyperactivity disorder and are known to be effective in enhancing attention-related processes; however, the underlying mechanisms have not been adequately addressed. Mice lacking the Adcyap1 gene encoding the neuropeptide pituitary adenylate cyclase-activating polypeptide (Adcyap1−/−) display psychomotor abnormalities, including increased novelty-seeking behavior and hyperactivity. In this study, Adcyap1−/− mice showed sensory-motor gating deficits, measured as deficits in prepulse inhibition (PPI), and showed normal PPI in response to amphetamine. Amphetamine also significantly decreased hyperlocomotion in Adcyap1−/− mice, and this paradoxical antihyperkinetic effect depended on serotonin 1A (5-HT1A) receptor signaling. c-Fos-positive neurons were increased in the prefrontal cortex in amphetamine-treated Adcyap1−/− mice, suggesting increased inhibitory control by prefrontal neurons. Additionally, amphetamine produced an antihyperkinetic effect in wild-type mice that received the 5-HT1A agonist 8-hydroxy-2-(di-n-propylamino)tetralin. These results indicate that Adcyap1−/− mice act as a model of hyperlocomotion and PPI deficits and suggest that 5-HT1A-mediated pathways are important determinants of the psychostimulant-elicited, rate-dependent effects that are in a negative function of the baseline rate of activity.
Keywords: neuropeptide, knock-out mice, psychostimulant, hyperactivity, prepulse inhibition, serotonin 5-HT1A receptor
Introduction
Pituitary adenylate cyclase-activating polypeptide (PACAP) is a neuropeptide originally isolated from ovine hypothalamus based on its ability to stimulate adenylate cyclase in rat anterior pituitary cell cultures and a member of the vasoactive intestinal peptide (VIP)/secretin/glucagon family. It exerts multiple activities as a neurotransmitter or neuromodulator via three G-protein-linked receptors, one PACAP-specific (PAC1) receptor and two receptors that are shared with VIP (VPAC1 and VPAC2) (Arimura, 1998; Vaudry et al., 2000; Hashimoto et al., 2006). Our recently developed mice lacking the Adcyap1 gene encoding the neuropeptide PACAP (Adcyap1−/−) have marked phenotypes, including behavioral abnormalities (Hashimoto et al., 2001; Shintani et al., 2002; Kawaguchi et al., 2003; Tanaka et al., 2004). Adcyap1−/− mice are born in the expected Mendelian ratios but show a high early mortality rate before weaning. The surviving Adcyap1−/− females exhibit reduced fertility, which is partly attributable to reduced mating frequency, and inadequate maternal behavior. Furthermore, Adcyap1−/− mice display remarkable behavioral changes, including hyperlocomotion and jumping behavior in an open field, and increased novelty-seeking behavior. These salient phenotypes may be attributable to, at least in part, perturbed monoamine neurotransmission, because serotonin (5-HT) metabolism is slightly decreased in the cerebral cortex and striatum of Adcyap1−/− mice, and hyperactive behavior is ameliorated by the antipsychotic drug haloperidol (Hashimoto et al., 2001). However, the mechanisms involved and the pathophysiological significance still remain unclear.
It is commonly accepted that changes in dopaminergic tone highly correlate with alterations in locomotor activity. Psychostimulants such as amphetamine and methylphenidate are indirect agonists that facilitate the action of catecholamines including dopamine (DA), and their effects on motor activity have been hypothesized as being rate dependent. Low baseline rates of activity are increased by stimulants, whereas higher rates are increased to a lesser extent, or even decreased, as a result of drug treatment, such that stimulant-induced change is a negative linear function of the baseline rate of activity (Solanto, 1998, 2002). However, the underlying mechanism remains essentially unknown.
Prepulse inhibition (PPI) is the phenomenon in which a weak prepulse stimulus attenuates the response to a subsequent startling stimulus, providing an operational measure of sensorimotor gating and a cross-species form of information processing, deficient in patients with schizophrenia and some other neuropsychiatric disorders, including comorbid attention-deficit hyperactivity disorder (ADHD) and Tourette's syndrome (Castellanos et al., 1996; Geyer et al., 2001). Although psychostimulants reduce PPI in normal subjects, they are known to be effective in enhancing attention-related processes (Solanto, 2002) as well as attending PPI (Hawk et al., 2003) in ADHD children. However, stimulant effects on PPI have not been adequately assessed clinically, and with respect to stimulant response of PPI, there is no animal model showing a positive response. To date, understanding of the therapeutic action mechanisms of stimulants is still in its infancy.
In the present study, we demonstrated that Adcyap1−/− mice showed psychopathological aspects of hyperactivity and PPI deficits, as well as beneficial responses to amphetamine. Our results give new insights into the mechanisms underlying the therapeutic effects of psychostimulants.
Materials and Methods
Animals.
All animal experiments were performed in accordance with protocols approved by the Animal Care and Use Committee of Graduate School of Pharmaceutical Sciences, Osaka University. Generation of Adcyap1−/− mice by a gene-targeting technique has been reported previously (Hashimoto et al., 2001). The null mutation was backcrossed onto an Institute of Cancer Research mouse background. Wild-type and Adcyap1−/− mice used were obtained from the intercross of heterozygous animals.
PPI analysis.
Acoustic startle responses were measured in a startle chamber (SR-LAB; San Diego Instruments, San Diego, CA) using standard methods described previously (Sakaue et al., 2003). The testing session started with a 5 min acclimatization to the startle chamber in the presence of 65 dB background white noise. Testing consisted of 40 120-dB pulses alone and 10 pulses preceded (100 ms) by a prepulse of 66, 68, 71, or 77 dB. Pulses were randomly presented with an average of 15 s between pulses. Twelve no-stimulus trials were included to assess spontaneous activity during testing that was routinely observed to be 10–20 (arbitrary unit). For drug treatments, animals were placed in the startle chamber just after intraperitoneal injection of amphetamine or 30 min after intraperitoneal injection of haloperidol. PPI was calculated as a percentage score: PPI (%) = (1 − [(startle response for pulse with prepulse)/(startle response for pulse alone)]) × 100.
Locomotor activity.
Locomotor activity was quantified in plastic activity monitoring boxes (30 × 30 × 30 cm) for 90 min using an infrared photocell beam detection system Acti-Track (Panlab, Barcelona, Spain) after intraperitoneal injection of drug or saline. The number of jumps was scored for 90 min using video recordings by experienced observers blinded to the mouse genotypes.
Measurement of rectal temperature.
Rectal temperature was recorded with a Physitemp Bat 12 digital thermometer (Physitemp Instruments, Clifton, NJ) before and after intraperitoneal drug injection.
Immunohistochemistry.
After intraperitoneal injection of amphetamine or saline, mice were placed back into their boxes. Two hours after injection, mice were deeply anesthetized with pentobarbital, perfused transcardially with saline, followed by a solution of 4% paraformaldehyde in PBS. Frontal sections (30 μm) containing medial prefrontal cortices (prelimbic cortex and infralimbic cortex) at +1.42 mm from the bregma and dorsomedial striatum at +0.50 mm from the bregma (Franklin and Paxinos, 1997) were cut and processed for immunohistochemistry with anti-c-Fos rabbit polyclonal primary antibody (sc-52; Santa Cruz Biotechnology, Santa Cruz, CA) and biotin-labeled anti-rabbit IgG secondary antibody (Nichirei, Tokyo, Japan). c-Fos-positive nuclei were counted manually by experienced observers blinded to the mouse genotypes.
Statistical analysis.
Statistically significant differences were assessed by ANOVA, followed by post hoc Mann–Whitney U test or Tukey's multiple comparison test, where applicable.
Results
PPI deficits in Adcyap1−/− mice
To investigate a possible role of PACAP in sensorimotor gating, PPI was measured in Adcyap1−/− mice and their wild-type littermate controls. There was no significant difference in startle amplitudes elicited at 100 or 120 dB between the two groups. Pulse intensities of 65 and 77 dB, selected as background noise and the highest prepulse intensity, respectively, elicited negligible startle when not paired with the startle stimulus in both groups (Fig. 1A, bottom). Adcyap1−/− mice showed diminished PPI at 71 and 77 dB prepulse intensities compared with wild-type mice (Fig. 1A, top). To examine the developmental changes of PPI deficits in Adcyap1−/− mice, PPI was tested at postnatal weeks 4, 6, and 8. There was no significant difference in startle amplitudes between the two groups at all studied ages (Fig. 1B, bottom). At postnatal week 4, PPI levels were similar between the two groups (Fig. 1B, top). Although wild-type mice showed an age-dependent increase in PPI, there was no significant increase in PPI in Adcyap1−/− mice from 4 to 8 weeks of age. At postnatal weeks 6 and 8, PPI levels were lower in Adcyap1−/− mice when compared with wild-type mice by 44 and 35%, respectively. PPI deficits in Adcyap1−/− mice were also seen at 14 weeks of age (data not shown).
Figure 1.

Startle amplitudes without prepulses and PPI of the startle reflex in Adcyap1−/− mice. A–D, PPI levels (top) and startle responses (bottom) at different pulse intensities in 8-week-old mice (A), at 77 dB prepulse followed by 120 dB startle pulses in mice at postnatal weeks 4, 6, and 8 (B), or in 8-week-old mice after pretreatment with haloperidol (C) or amphetamine (D). The number of wild-type (open bars) and Adcyap1−/− (closed bars) mice are indicated above the bars. Data are expressed as means ± SEM.
Effects of psychostimulants on PPI deficits in Adcyap1−/− mice
Dopaminergic systems have been postulated to be involved in the control of PPI (Swerdlow and Geyer, 1998); therefore, the effects of the D2 dopamine receptor blocking antipsychotic haloperidol on PPI deficits were assessed in Adcyap1−/− mice. Haloperidol (0.02 or 0.2 mg/kg) failed to improve PPI deficits in Adcyap1−/− mice (Fig. 1C, top) and showed no significant effect on startle amplitudes in both groups (Fig. 1C, bottom). We assessed possible effects of psychostimulants on the PPI deficits in Adcyap1−/− mice and found that amphetamine, at a clinically relevant dose range (2 mg/kg) (Gainetdinov and Caron, 2000), reversed PPI deficits to the control level in wild-type mice (Fig. 1D, top). Consistent with a previous report (Ralph et al., 2001b), a higher dose of amphetamine (10 mg/kg) tended to reduce PPI in wild-type mice, although there were no significant changes in PPI levels between the two groups. Startle amplitudes were reduced in Adcyap1−/− mice that received 10 mg/kg amphetamine (Fig. 1D, bottom).
Effects of haloperidol and psychostimulants on abnormal jumping behavior in Adcyap1−/− mice
We examined the effects of haloperidol and psychostimulants on explosive jumping behavior in Adcyap1−/− mice. Haloperidol effectively reduced the number of jumps in Adcyap1−/− mice (Fig. 2A). Interestingly, amphetamine and methylphenidate also reduced the number of jumps in Adcyap1−/− mice (Fig. 2B,C).
Figure 2.

A–C, Number of jumps in Adcyap1−/− mice after pretreatment with haloperidol (A), amphetamine (B), and methylphenidate (C). The number of wild-type (open bars) and Adcyap1−/− (closed bars) mice are indicated above the bars. Data are expressed as means ± SEM.
Amphetamine-evoked paradoxical antihyperkinetic effect
Adcyap1−/− mice maintained high initial levels of locomotor activity with reduced thigmotaxis or wall-hugging behavior, an index of anxiety, during the open field test (Hashimoto et al., 2001). As expected, wild-type mice responded to amphetamine (2 mg/kg) with increased locomotor activity (Fig. 3A); however, in sharp contrast, amphetamine (2 mg/kg) paradoxically attenuated hyperlocomotion in Adcyap1−/− mice (Fig. 3B). Likewise, 10 mg/kg amphetamine increased locomotor activity in wild-type mice and still produced antihyperkinetic effects in Adcyap1−/− mice (data not shown). Adcyap1−/− mice entered the center region more often than wild-type mice. Amphetamine (2 mg/kg) inhibited such aberrant behavior and, instead, increased thigmotaxis as seen in wild-type mice (Fig. 3C).
Figure 3.

Locomotor activity in the open field test. A, B, Locomotor activity in wild-type (A) and Adcyap1−/− (B) mice that received 2 mg/kg amphetamine (closed circles) or saline (open circles). n = 16 per group. ∗p < 0.05 and ∗∗p < 0.01 versus saline. C, Representative locomotor patterns of saline-treated (top panels) or 2 mg/kg amphetamine-treated (bottom panels) wild-type (left panels) and Adcyap1−/− (right panels) mice during 25–30 min of a 90 min recording in an open field test. Data are expressed as means ± SEM.
Possible involvement of 5-HT1A receptor signaling in the psychobehavioral changes
We explored possible neurochemical alterations relevant to psychobehavioral changes in Adcyap1−/− mice and found reduced hypothermic response to 5-HT1A agonists. The 5-HT1A agonists 8-hydroxy-2-(di-n-propylamino)tetralin (8-OH-DPAT) or buspirone significantly decreased rectal temperature in wild-type mice, whereas the response was markedly attenuated in Adcyap1−/− mice (Fig. 4). Therefore, we examined the possible involvement of 5-HT1A signaling in the amphetamine-evoked antihyperkinetic effect in Adcyap1−/− mice. The selective 5-HT1A receptor antagonist N-[2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl]-N-(2-pyridinyl)cyclohexanecarboxamide (WAY-100635) (0.3 mg/kg) blocked the action of amphetamine (Fig. 5B). We also found that amphetamine produced paradoxical calming effects in wild-type mice that received 8-OH-DPAT (0.05 mg/kg) (Fig. 5A). Neither WAY-100635 nor 8-OH-DPAT alone influenced locomotor activity in wild-type and Adcyap1−/− mice (Fig. 5C,D).
Figure 4.

5-HT1A agonist-induced hypothermia. Wild-type (open circles) and Adcyap1−/− (closed circles) mice were injected intraperitoneally with 0.1 mg/kg 8-OH-DPAT (A), 0.5 mg/kg 8-OH-DPAT (B), or 10 mg/kg buspirone (C). Rectal temperature was measured at the indicated times. n = 7–8 per group. ∗p < 0.05 and ∗∗p < 0.01 versus wild-type mice. Data are expressed as means ± SEM.
Figure 5.

Locomotor activity in the open field test. A–D, Locomotor activity in wild-type (A, C) and Adcyap1−/− (B, D) mice that received 0.05 mg/kg 8-OH-DPAT (triangles), 0.3 mg/kg WAY-100635 (squares), or saline (circles) either alone (open symbols) or in combination with 2 mg/kg amphetamine (closed symbols). The results of the experiment with amphetamine or saline alone are the same as those in Figure 3. n = 15–16 per group. ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001 versus saline alone; †p < 0.05, ††p < 0.01, and †††p < 0.001 versus amphetamine. Data are expressed as means ± SEM.
c-Fos-positive neurons were increased in prefrontal cortex in amphetamine-treated Adcyap1−/− mice
Presynaptic 5-HT1A autoreceptors have been shown to mediate the hypothermic response, and receptor density correlates with the hypothermic response to 5-HT1A agonists (Aguirre et al., 1998). Therefore, we performed reverse transcription-PCR analysis to quantify 5-HT1A and other 5-HT receptor mRNA levels, as well as microarray analysis. However, to date, we have not confirmed changes in expression of mRNA for 5-HT receptors and other genes that are probably responsible for altered psychomotor functions in Adcyap1−/− mice (data not shown).
There is evidence implicating the prefrontal cortex in the pathophysiology of motor dysregulation as well as PPI deficits (Swerdlow and Geyer, 1998; Goldman-Rakic et al., 2000). Therefore, we examined c-Fos expression as a marker for postsynaptic activity to define the pattern of neurons excited by amphetamine (Fig. 6). The number of c-Fos-positive neurons increased in the medial prefrontal cortex in amphetamine-treated Adcyap1−/− mice compared with wild-type mice, suggesting hyperactivation of Adcyap1−/− prefrontal cortical neurons by amphetamine. Amphetamine-induced increase in the number of c-Fos-positive neurons in the dorsomedial striatum (Fig. 6) and other regions, including the cingulate cortex and nucleus accumbens (data not shown), was not significantly different between the two groups.
Figure 6.
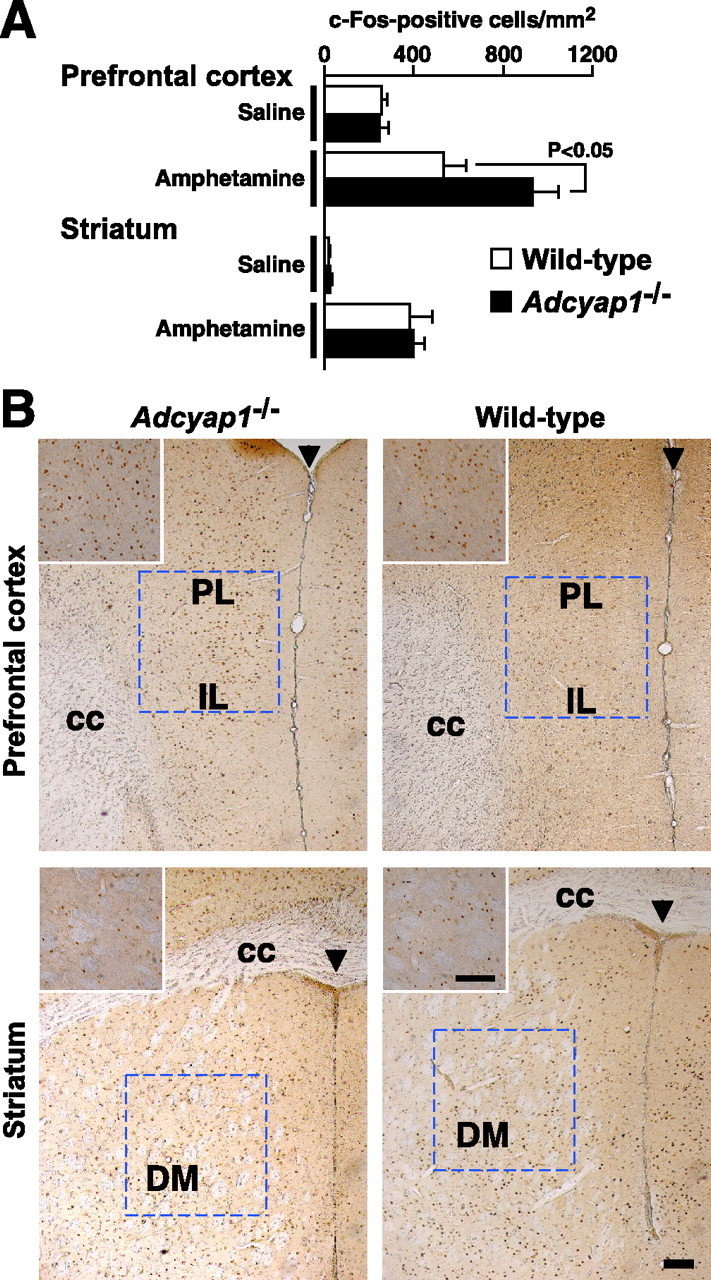
c-Fos-positive neurons in medial prefrontal cortex and dorsomedial striatum in amphetamine-treated Adcyap1−/− mice. A, Number of c-Fos-positive neurons in the outlined regions in B in wild-type (open bars) and Adcyap1−/− (closed bars) mice that received 10 mg/ml amphetamine. n = 6–7 per group. B, Photomicrographs showing representative c-Fos labeling in medial prefrontal cortex (top panels) and dorsomedial striatum (bottom panels) in amphetamine-treated Adcyap1−/− (left panels) and wild-type (right panels) mice. Insets, High magnifications of c-Fos staining. PL, Prelimbic cortex; IL, infralimbic cortex; DM, dorsomedial striatum; cc, corpus callosum; arrowheads, midline. Scale bars, 100 μm. Data are expressed as means ± SEM.
Discussion
Psychostimulants are controlled substances, because they have long-term sensitizing effects and a potential for abuse, and they may also be neurotoxic. However, psychostimulant treatment has long been recognized to attenuate hyperactivity, paradoxically, and often to improve cognitive performance (Castellanos and Tannock, 2002; Solanto, 2002; Garland and Kirkpatrick, 2004). Animal studies have demonstrated that psychostimulants have biphasic effects on motor activity and cognitive processes (Solanto, 1998), but underlying mechanisms remain mostly uncharacterized. We demonstrated that Adcyap1−/− mice showed psychopathological changes, including PPI deficits, and investigated the effects of psychostimulants on hyperactivity, PPI deficits, and excessive jumping activity, as well as the role of 5-HT1A signaling in the paradoxical actions of amphetamine on hyperactivity. These findings seem a long way from the clinical disorders of hyperkinesia and cognitive impairment, but we hope to provide models with some typical features of psychostimulant responses in these disorders.
There is considerable evidence for the involvement of dopaminergic systems in the control of PPI (Geyer et al., 2001). We demonstrated that, although hyperlocomotion (Hashimoto et al., 2001) and jumping behavior in Adcyap1−/− mice were effectively attenuated by haloperidol (a D2 antagonist), PPI deficits were not reversed by haloperidol. Other than DA, glutamatergic systems are important for modulating PPI (Geyer et al., 2001). PPI is reduced in rodents and humans by noncompetitive NMDA antagonists, such as phencyclidine and dizocilpine [(+)-5-methyl-10,11-dihydro-5H-dibenzo [a,d] cyclohepten-5,10-imine maleate]. Several studies have shown that PACAP can potentiate NMDA receptor functions (Stella and Magistretti, 1996; Liu and Madsen, 1997; Pellegri et al., 1998). Recently, we showed that Adcyap1−/− mice do not exhibit inflammatory or neuropathic pain, and PACAP is required for functional coupling of neuronal nitric oxide synthase to NMDA receptors in the spinal cord for chronic pain to occur (Mabuchi et al., 2004). This raises the possibility that similar mechanisms might be involved in psychomotor changes in Adcyap1−/− mice. PPI deficits in Adcyap1−/− mice may therefore be ascribable in part to NMDA hypofunction.
One of the most striking findings in our study was that amphetamine completely reversed PPI deficits in Adcyap1−/− mice and, to our knowledge, this is the first animal model showing PPI deficits and paradoxical responses to psychostimulants. DA transporter knock-out mice were shown to exhibit PPI deficits, which were improved by the D2 receptor antagonist raclopride (Ralph et al., 2001a) and the 5-HT2A receptor antagonist M100907 (Barr et al., 2004). Therefore, it will be interesting to investigate the effects of psychostimulants on PPI deficits in DA transporter knock-out mice.
Adcyap1−/− mice did not show a significant increase in PPI from 4 to 8 weeks of age, suggesting that the process of sensorimotor gating remains functionally immature. Developmental issues are considered to be particularly relevant to understanding PPI deficits in humans, because PPI appears to develop in children 5–8 years of age (Ornitz et al., 1990). In addition, Adcyap1−/− mice showed a slight decrease in 5-HT metabolite 5-hydroxyindoleacetic acid in their brain (Hashimoto et al., 2001), and this was manifested at 4 weeks of age (the earliest age tested) (our unpublished data). Several lines of evidence suggest that PACAP acts as a neurotrophic factor and plays diverse roles in mammalian neurogenesis (Arimura, 1998; Vaudry et al., 2000). Therefore, it is conceivable that developmental defects in 5-HT systems, or the relative balance of tone between 5-HT and other neurotransmitter systems, such as the glutamatergic system, may contribute to PPI deficits and hyperactivity in Adcyap1−/− mice. However, this does not exclude the possibility that PACAP is actively involved in psychological functions. Three different PACAP knock-out lines (Gray et al., 2001; Hamelink et al., 2002; Colwell et al., 2004) developed separately from our colony have been reported to also show dysfunction of lipid and carbohydrate metabolism, cold hypersensitivity, impaired catecholamine regulation in the sympathoadrenal axis, and deficits in the circadian light response, suggesting putative developmental and/or neuroplastic abnormalities in these mutant mice.
It has been postulated that prefrontal cortex dysregulation may lead to disinhibition in targets of the prefrontal cortex projection, with a possible relevance to dysregulation of motor functions and PPI (Swerdlow and Geyer, 1998; Goldman-Rakic et al., 2000). In vivo microdialysis showed that basal and amphetamine-induced release of extracellular DA and 5-HT in the prefrontal cortex did not differ significantly between Adcyap1−/− and wild-type mice (DA, F(20,180) = 0.198; not significant; 5-HT, F(20,180) = 1.069; not significant). However, the number of c-Fos-positive neurons increased in the medial prefrontal cortex of Adcyap1−/− mice compared with wild-type mice after amphetamine administration. This result raises the possibility that hyperactivation of prefrontal cortical neurons by amphetamine might result in an increased inhibitory control by prefrontal neurons in Adcyap1−/− mice. It has been demonstrated that c-fos mRNA expression in the frontal cortex is increased by environmental novelty, but this effect is not further increased by amphetamine (Badiani et al., 1998). Likewise, when exposed to novelty (alone), the increase in c-Fos-positive neurons in the prefrontal cortex tended to be greater in Adcyap1−/− mice compared with wild-type mice, but the effect of novelty was similar to that of amphetamine in the respective groups of mice (data not shown). Additional studies to define and characterize these cell populations responsible for the effects of amphetamine and novelty will help to investigate the neuronal mechanisms for psychostimulant treatment.
Studies in DA transporter knock-out mice suggest the tantalizing possibility that hyperkinetic behavior might be controlled through precise targeting of 5-HT receptors, or even through enhanced availability of 5-HT precursors (Gainetdinov et al., 1999). The latter possibility is still controversial, because therapeutic efficacy of selective serotonergic drugs is not commonly recognized in treating hyperkinetic disorder (Gainetdinov and Caron, 2000; Popper, 2000; Davids et al., 2003). The reason for this may be simply a result of the large multiplicity of 5-HT receptor subtypes and existence of multiple 5-HT autoreceptors, having sometimes opposing or no effects on locomotion (Geyer, 1996; Lucki, 1998). Regarding the former possibility (precise targeting of 5-HT receptors), the present study showed that WAY-100635 blocked the amphetamine-elicited antihyperkinetic effect in Adcyap1−/− mice, and that amphetamine produced a paradoxical calming effect in 8-OH-DPAT-treated wild-type mice. Previous reports show that 8-OH-DPAT attenuates psychostimulant-induced behavioral sensitization and increment in locomotor activity, where both presynaptic and postsynaptic 5-HT1A receptor-dependent mechanisms are suggested to be involved (Przegalinski and Filip, 1997; Przegalinski et al., 2000; Carey et al., 2005). However, the paradoxical calming effect of psychostimulant plus 5-HT1A receptor agonist combination has hitherto not been addressed and may provide provocative clues into the mechanisms of the therapeutic effects of psychostimulants. Together with previous reports, the present data suggest that the targeting of 5-HT1A receptors, adjunctive to psychostimulants, are promising for pharmaceutical intervention in hyperkinetic disorder.
Histochemical studies have shown that PACAP immunoreactivity is present in several brain regions involved in the DA and 5-HT systems, including the cerebral cortex, nucleus accumbens, amygdala, hypothalamus, substantia nigra, ventral tegmental area, and dorsal raphe nucleus (Masuo et al., 1993; Piggins et al., 1996). PAC1 receptors are expressed broadly in both the target areas and nuclei of origin of these monoaminergic systems (Hashimoto et al., 1996). VPAC1 and VPAC2 receptors are also expressed in these systems (Usdin et al., 1994). These observations suggest a functional relationship between PACAP and monoaminergic neurons. Serotonergic cell bodies are located mainly in the raphe nuclei, whereas 5-HT-containing terminals are widely distributed in the brain. Among 5-HT receptor subtypes, 5-HT1A receptors are expressed presynaptically as the primary somatodendritic autoreceptor on serotonergic raphe neurons and postsynaptically in a variety of other neurons (Barnes and Sharp, 1999). Although there is no report of colocalization analysis between PACAP/PACAP receptors and 5-HT-containing elements, it is plausible that PACAP modulates the serotonergic system both at the origin and innervation sites. The reduced hypothermic response to 5-HT1A agonists in Adcyap1−/− mice supports the functional coupling between the two systems.
The supplemental figure (available at www.jneurosci.org as supplemental material) shows a schematic representation of possible relationships between locomotor activity and degree of stimulation of 5-HT1A systems in amphetamine-treated Adcyap1−/− and wild-type mice. Neither WAY-100635 nor 8-OH-DPAT alone influenced locomotor activity in both groups, indicating that changes in 5-HT1A systems per se do not influence locomotor activity. The observations that WAY-100635 blocked the amphetamine-elicited antihyperkinetic effect in Adcyap1−/− mice, and that amphetamine induced hypokinesia in 8-OH-DPAT-treated wild-type mice, indicate that the 5-HT1A relative activity has a great influence on the effects of amphetamine. Psychostimulants have been hypothesized to exert rate-dependent effects that show a negative linear correlation with the baseline rate of activity (Solanto, 1998, 2002). The present results suggest that the stimulation level of 5-HT1A systems, or the relative balance with other 5-HT receptor subtypes or neurotransmitter systems, might be involved in the rate-dependent effects elicited by psychostimulants.
Many psychiatric disorders are multifactorial, reflecting longitudinal and complex interactions of causative agents, including genetic and environmental factors. Pathogenesis remains poorly understood; therefore, animal models provide useful tools to investigate the mechanisms underlying human diseases and for the design of new treatments (Lipska and Weinberger, 2000). Although, Adcyap1−/− mice do not provide an animal model of some specific psychiatric disorder per se, the present study may provide insights into the pathophysiology and etiology of hyperkinetic disorder and other disorders including disrupted PPI.
Footnotes
*K.T. and N.S. contributed equally to this work.
This work was supported in part by Grants-in-Aid for Scientific Research and for Young Scientists from Japan Society for the Promotion of Science and by grants from Sankyo Foundation of Life Science and Taisho Pharmaceutical Company.
References
- Aguirre N, Ballaz S, Lasheras B, Del-Rio J (1998). MDMA (“Ecstasy”) enhances 5-HT1A receptor density and 8-OH-DPAT-induced hypothermia: blockade by drugs preventing 5-hydroxytryptamine depletion. Eur J Pharmacol 346:181–188. [DOI] [PubMed] [Google Scholar]
- Arimura A (1998). Perspectives on pituitary adenylate cyclase activating polypeptide (PACAP) in the neuroendocrine, endocrine, and nervous systems. J Physiol (Japan) 48:301–331. [DOI] [PubMed] [Google Scholar]
- Badiani A, Oates MM, Day HE, Watson SJ, Akil H, Robinson TE (1998). Amphetamine-induced behavior, dopamine release, and c-fos mRNA expression: modulation by environmental novelty. J Neurosci 18:10579–10593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes NM, Sharp T (1999). A review of central 5-HT receptors and their function. Neuropharmacology 38:1083–1152. [DOI] [PubMed] [Google Scholar]
- Barr AM, Lehmann-Masten V, Paulus M, Gainetdinov RR, Caron MG, Geyer MA (2004). The selective serotonin-2A receptor antagonist M100907 reverses behavioral deficits in dopamine transporter knockout mice. Neuropsychopharmacology 29:221–228. [DOI] [PubMed] [Google Scholar]
- Carey RJ, DePalma G, Damianopoulos E, Shanahan A, Muller CP, Huston JP (2005). Evidence that the 5-HT1A autoreceptor is an important pharmacological target for the modulation of cocaine behavioral stimulant effects. Brain Res 1034:162–171. [DOI] [PubMed] [Google Scholar]
- Castellanos FX, Tannock R (2002). Neuroscience of attention-deficit/hyperactivity disorder: the search for endophenotypes. Nat Rev Neurosci 3:617–628. [DOI] [PubMed] [Google Scholar]
- Castellanos FX, Fine EJ, Kaysen D, Marsh WL, Rapoport JL, Hallett M (1996). Sensorimotor gating in boys with Tourette's syndrome and ADHD: preliminary results. Biol Psychiatry 39:33–41. [DOI] [PubMed] [Google Scholar]
- Colwell CS, Michel S, Itri J, Rodriguez W, Tam J, Lelievre V, Hu Z, Waschek JA (2004). Selective deficits in the circadian light response in mice lacking PACAP. Am J Physiol Regul Integr Comp Physiol 287:R1194–R1201. [DOI] [PubMed] [Google Scholar]
- Davids E, Zhang K, Tarazi FI, Baldessarini RJ (2003). Animal models of attention-deficit hyperactivity disorder. Brain Res Brain Res Rev 42:1–21. [DOI] [PubMed] [Google Scholar]
- Franklin KBJ, Paxinos G (1997). In: The mouse brain in stereotaxic coordinates San Diego: Academic.
- Gainetdinov RR, Caron MG (2000). An animal model of attention deficit hyperactivity disorder. Mol Med Today 6:43–44. [DOI] [PubMed] [Google Scholar]
- Gainetdinov RR, Wetsel WC, Jones SR, Levin ED, Jaber M, Caron MG (1999). Role of serotonin in the paradoxical calming effect of psychostimulants on hyperactivity. Science 283:397–401. [DOI] [PubMed] [Google Scholar]
- Garland M, Kirkpatrick P (2004). Atomoxetine hydrochloride. Nat Rev Drug Discov 3:385–386. [DOI] [PubMed] [Google Scholar]
- Geyer MA (1996). Serotonergic functions in arousal and motor activity. Behav Brain Res 73:31–35. [DOI] [PubMed] [Google Scholar]
- Geyer MA, Krebs-Thomson K, Braff DL, Swerdlow NR (2001). Pharmacological studies of prepulse inhibition models of sensorimotor gating deficits in schizophrenia: a decade in review. Psychopharmacology 156:117–154. [DOI] [PubMed] [Google Scholar]
- Goldman-Rakic PS, Muly EC III, Williams GV (2000). D1 receptors in prefrontal cells and circuits. Brain Res Brain Res Rev 31:295–301. [DOI] [PubMed] [Google Scholar]
- Gray SL, Cummings KJ, Jirik FR, Sherwood NM (2001). Targeted disruption of the pituitary adenylate cyclase-activating polypeptide gene results in early postnatal death associated with dysfunction of lipid and carbohydrate metabolism. Mol Endocrinol 15:1739–1747. [DOI] [PubMed] [Google Scholar]
- Hamelink C, Tjurmina O, Damadzic R, Young WS, Weihe E, Lee HW, Eiden LE (2002). Pituitary adenylate cyclase-activating polypeptide is a sympathoadrenal neurotransmitter involved in catecholamine regulation and glucohomeostasis. Proc Natl Acad Sci USA 99:461–466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimoto H, Nogi H, Mori K, Ohishi H, Shigemoto R, Yamamoto K, Matsuda T, Mizuno N, Nagata S, Baba A (1996). Distribution of the mRNA for a pituitary adenylate cyclase-activating polypeptide receptor in the rat brain: an in situ hybridization study. J Comp Neurol 371:567–577. [DOI] [PubMed] [Google Scholar]
- Hashimoto H, Shintani N, Tanaka K, Mori W, Hirose M, Matsuda T, Sakaue M, Miyazaki J, Niwa H, Tashiro F, Yamamoto K, Koga K, Tomimoto S, Kunugi A, Suetake S, Baba A (2001). Altered psychomotor behaviors in mice lacking pituitary adenylate cyclase-activating polypeptide (PACAP). Proc Natl Acad Sci USA 98:13355–13360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimoto H, Shintani N, Baba A (2006). New insights into the central PACAPergic system from the phenotypes in PACAP- and PACAP receptor-knockout mice. Ann NY Acad Sci in press. [DOI] [PubMed]
- Hawk LW Jr, Yartz AR, Pelham WE Jr, Lock TM (2003). The effects of methylphenidate on prepulse inhibition during attended and ignored prestimuli among boys with attention-deficit hyperactivity disorder. Psychopharmacology 165:118–127. [DOI] [PubMed] [Google Scholar]
- Kawaguchi C, Tanaka K, Isojima Y, Shintani N, Hashimoto H, Baba A, Nagai K (2003). Changes in light-induced phase shift of circadian rhythm in mice lacking PACAP. Biochem Biophys Res Commun 310:169–175. [DOI] [PubMed] [Google Scholar]
- Lipska BK, Weinberger DR (2000). To model a psychiatric disorder in animals: schizophrenia as a reality test. Neuropsychopharmacology 23:223–239. [DOI] [PubMed] [Google Scholar]
- Liu GJ, Madsen BW (1997). PACAP38 modulates activity of NMDA receptors in cultured chick cortical neurons. J Neurophysiol 78:2231–2234. [DOI] [PubMed] [Google Scholar]
- Lucki I (1998). The spectrum of behaviors influenced by serotonin. Biol Psychiatry 44:151–162. [DOI] [PubMed] [Google Scholar]
- Mabuchi T, Shintani N, Matsumura S, Okuda-Ashitaka E, Hashimoto H, Muratani T, Minami T, Baba A, Ito S (2004). Pituitary adenylate cyclase-activating polypeptide is required for the development of spinal sensitization and induction of neuropathic pain. J Neurosci 24:7283–7291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masuo Y, Suzuki N, Matsumoto H, Tokito F, Matsumoto Y, Tsuda M, Fujino M (1993). Regional distribution of pituitary adenylate cyclase activating polypeptide (PACAP) in the rat central nervous system as determined by sandwich-enzyme immunoassay. Brain Res 602:57–63. [DOI] [PubMed] [Google Scholar]
- Ornitz EM, Guthrie D, Lane SJ, Sugiyama T (1990). Maturation of startle facilitation by sustained prestimulation. Psychophysiology 27:298–308. [DOI] [PubMed] [Google Scholar]
- Pellegri G, Magistretti PJ, Martin JL (1998). VIP and PACAP potentiate the action of glutamate on BDNF expression in mouse cortical neurones. Eur J Neurosci 10:272–280. [DOI] [PubMed] [Google Scholar]
- Piggins HD, Stamp JA, Burns J, Rusak B, Semba K (1996). Distribution of pituitary adenylate cyclase activating polypeptide (PACAP) immunoreactivity in the hypothalamus and extended amygdala of the rat. J Comp Neurol 376:278–294. [DOI] [PubMed] [Google Scholar]
- Popper CW (2000). Pharmacologic alternatives to psychostimulants for the treatment of attention-deficit/hyperactivity disorder. Child Adolesc Psychiatr Clin N Am 9:605–646. [PubMed] [Google Scholar]
- Przegalinski E, Filip M (1997). Stimulation of serotonin (5-HT)1A receptors attenuates the locomotor, but not the discriminative, effects of amphetamine and cocaine in rats. Behav Pharmacol 8:699–706. [DOI] [PubMed] [Google Scholar]
- Przegalinski E, Siwanowicz J, Baran L, Filip M (2000). Activation of serotonin (5-HT)1A receptors inhibits amphetamine sensitization in mice. Life Sci 66:1011–1019. [DOI] [PubMed] [Google Scholar]
- Ralph RJ, Paulus MP, Fumagalli F, Caron MG, Geyer MA (2001a). Prepulse inhibition deficits and perseverative motor patterns in dopamine transporter knock-out mice: differential effects of D1 and D2 receptor antagonists. J Neurosci 21:305–313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ralph RJ, Paulus MP, Geyer MA (2001b). Strain-specific effects of amphetamine on prepulse inhibition and patterns of locomotor behavior in mice. J Pharmacol Exp Ther 298:148–155. [PubMed] [Google Scholar]
- Sakaue M, Ago Y, Baba A, Matsuda T (2003). The 5-HT1A receptor agonist MKC-242 reverses isolation rearing-induced deficits of prepulse inhibition in mice. Psychopharmacology 170:73–79. [DOI] [PubMed] [Google Scholar]
- Shintani N, Mori W, Hashimoto H, Imai M, Tanaka K, Tomimoto S, Hirose M, Kawaguchi C, Baba A (2002). Defects in reproductive functions in PACAP-deficient female mice. Regul Pept 109:45–48. [DOI] [PubMed] [Google Scholar]
- Solanto MV (1998). Neuropsychopharmacological mechanisms of stimulant drug action in attention-deficit hyperactivity disorder: a review and integration. Behav Brain Res 94:127–152. [DOI] [PubMed] [Google Scholar]
- Solanto MV (2002). Dopamine dysfunction in AD/HD: integrating clinical and basic neuroscience research. Behav Brain Res 130:65–71. [DOI] [PubMed] [Google Scholar]
- Stella N, Magistretti PJ (1996). Vasoactive intestinal peptide (VIP) and pituitary adenylate cyclase-activating polypeptide (PACAP) potentiate the glutamate-evoked release of arachidonic acid from mouse cortical neurons. Evidence for a cAMP-independent mechanism. J Biol Chem 271:23705–23710. [DOI] [PubMed] [Google Scholar]
- Swerdlow NR, Geyer MA (1998). Using an animal model of deficient sensorimotor gating to study the pathophysiology and new treatments of schizophrenia. Schizophr Bull 24:285–301. [DOI] [PubMed] [Google Scholar]
- Tanaka K, Hashimoto H, Shintani N, Yamamoto A, Baba A (2004). Reduced hypothermic and hypnotic responses to ethanol in PACAP-deficient mice. Regul Pept 123:95–98. [DOI] [PubMed] [Google Scholar]
- Usdin TB, Bonner TI, Mezey E (1994). Two receptors for vasoactive intestinal polypeptide with similar specificity and complementary distributions. Endocrinology 135:2662–2680. [DOI] [PubMed] [Google Scholar]
- Vaudry D, Gonzalez BJ, Basille M, Yon L, Fournier A, Vaudry H (2000). Pituitary adenylate cyclase-activating polypeptide and its receptors: from structure to functions. Pharmacol Rev 52:269–324. [PubMed] [Google Scholar]