

Abstract
Elevated levels of corticosteroid hormones, presumably occupying both mineralocorticoid receptors (MRs) and glucocorticoid receptors (GRs), have been reported to impair synaptic plasticity in the hippocampus as well as the acquisition of hippocampus-dependent memories. In contrast, recent evidence suggests that activation of MRs enhance cognitive functions. To clarify the roles of different steroid receptors in hippocampal plasticity, young adult rats were injected with the GR antagonist RU38486 (mifepristone) or the MR antagonist Spironolactone before the exposure to an acute swim stress. Hippocampal responses to perforant path stimulation were then recorded in anesthetized rats. Stress combined with RU38486 produced a striking facilitation of LTP. Spironolactone enabled only short-term potentiation that reversed to long-term depression (LTD) in the stressed animals. Finally, the blockade of both MRs and GRs led to impairment of long-term potentiation. These findings indicate that MRs and GRs assume opposite roles in regulation of synaptic plasticity after acute exposure to stressors.
Keywords: mineralocorticoid receptor, glucocorticoid receptor, DG hippocampus, acute swim stress, neural plasticity, LTP
Introduction
The hippocampus serves a pivotal role in memory formation (Squire, 1982). It has been suggested that corticosteroid modulation of hippocampal activity and plasticity may underlie some aspects of acute and possibly chronic effects of stress. Based on biochemical and functional characteristics, two types of corticosteroid receptors have been described in the brain (Reul and de Kloet, 1985; Lupien and McEwen, 1997). Type I or the high-affinity mineralocorticoid receptor (MR) is most densely localized in hippocampal and septal neurons. Type II or glucocorticoid receptor (GR) is ubiquitously distributed in the brain, including neurons in the hippocampus, hypothalamus, glial cells, and pituitary cells. These receptors complement each other and put both MRs and GRs in a position to modulate the limbic-hypothalamic-pituitary-adrenal axis (LHPA) responsivity under stressful conditions. The MRs are activated at low corticosterone (CORT) concentrations and may offer tonic inhibition to the LHPA axis (Reul and de Kloet, 1985; Funder, 1986; Dallman et al., 1987). When high corticosterone concentrations are present, MRs saturate, and the GRs take over. The dual action of these receptors in the hippocampus appears to be central for both basal modulation and stress regulation of the LHPA.
Basal corticosteroid levels are associated with the effective induction of long-term potentiation (LTP) in the hippocampus (Diamond et al., 1992). In contrast, elevated levels of corticosteroid hormones (presumably occupying both GRs and MRs) or exposure to stress have been reported to impair LTP and to facilitate long-term depression (LTD) in the hippocampus (Diamond et al., 1992; Pavlides et al., 1995, 1996; Pavlides and McEwen, 1999; Alfarez et al., 2002; Krugers et al., 2005). Little is known about the mechanism through which these stress-induced changes exert their effects, although NMDA receptors seem to be involved (Kim et al., 1996). Stress-mediated changes in glutamate transmission and calcium influx affect the propensity for synaptic plasticity, although not necessarily in the direction of impaired LTP. Furthermore, it has been suggested that the MRs and GRs function in a binary manner at the cellular level (de Kloet et al., 1998). MR activity is considered to maintain the excitability and stability of networks. Conversely, GR activation seems to involve the suppression or normalization of network activity (de Kloet et al., 2005). Previous studies did find differential effects of MRs and GRs on LTP in the DG (Pavlides et al., 1995), but as yet, no conclusive picture of the effects of the different steroid antagonists on in vivo LTP in the dentate gyrus (DG) was obtained.
The following experiments were designed to characterize corticosteroid modulation of hippocampal synaptic plasticity in baseline condition and after stress, depending on the relative activation of MRs and/or GRs.
Materials and Methods
Animals.
All procedures were approved by the Weizmann Institute Institutional Animal Care and Use Committee in accordance with standard legal guidelines.
Male Wistar rats weighing between 240 and 280 g (8–10 weeks of age) were housed four per cage in 75.0 × 55.0 × 15.0 cm Plexiglas cages in temperature-controlled (23 ± 1°C) animal quarters on a 12 h light/dark cycle (lights on from 7:00 A.M. to 7:00 P.M.) with ad libitum access to standard Purina Rat Chow pellets and water.
Drugs.
The MR antagonist Spironolactone (Sigma, St. Louis, MO) and the GR antagonist RU38486 (mifepristone) were dissolved in 1,2-propandiole (20 mg/10 ml) and were injected subcutaneously (20 mg/kg) one-half hour before the exposure to acute swim stress (ASS). The time span was chosen to allow proper passage of the drugs through the blood–brain barrier and their binding to the receptors. The doses of antagonists were based on previous studies (Smriga et al., 1998).
Adrenalectomy.
Rats were anesthetized with ketamine and diazepam (1:1; 0.25 ml/120 g) and adrenalectomized (Adx) 1 week before behavioral and electrophysiological procedures. A ∼2 cm midline incision was made on the dorsal hump. Using the kidney as a landmark, both adrenal glands were removed. Adx rats were provided with normal saline as drinking water for 3 d after the operation.
ASS.
Individual rats were placed for 15 min in a circular water tank (diameter, 0.5 m; height, 0.5 m). Water depth was 40.0 cm, and temperature was maintained at 23 ± 1°C. After exposure to ASS, rats were allowed to rest for one-half hour before anesthesia and electrophysiological recording (Avital et al., 2001; Avital and Richter-Levin, 2005).
Measurements of hippocampal activity and plasticity.
Rats were anesthetized with urethane (21% solution; 1.2 g/kg, i.p.) and placed in a stereotaxic apparatus. A bipolar 125 μm concentric stimulating electrode was placed in the perforant path (PP) (coordinates: 8 mm posterior to bregma, 4 mm lateral to the midline; depth was adjusted to yield maximal response of the DG). A glass pipette (diameter, 2–3 μm) containing a 3 m NaCl solution, was inserted into the DG of the dorsal hippocampus using a hydraulic micro-drive [coordinates: 4 mm posterior to bregma, 2.5 mm lateral to the midline; the depth of the electrode was adjusted to yield the largest field EPSP (fEPSP)]. Evoked responses were amplified and filtered at 1 Hz to 1 kHz. Preparation for recording usually lasted ∼30 min, and there were no significant differences in the preparation time between groups. Recording was allowed to stabilize for 10 min. Baseline field potential responses in the DG to PP stimulation were recorded using stimulus intensity that was 50% of the intensity that evoked maximal asymptotic spike amplitude (monopolar pulses, 100 μs duration). During recording, rectal temperature was maintained at 37 ± 0.5°C. Off-line measurements were made of the slope of the fEPSP, using averages of five successive responses to a given stimulation intensity applied at 0.1 Hz. Although the main parameter that was measured systematically was the population EPSP, parallel changes in population spikes were also noted, and because they changed in a similar manner to those observed with the fEPSPs, there was no systematic analysis of changes in population spikes.
To assess short-term plasticity, paired-pulse responses were obtained. A twin pulse stimulus was delivered at three interstimulus intervals (ISIs) (15, 30, and 60 ms), and averages of five successive responses to a given stimulus intensity, at each ISI, were quantified as the ratio of the second over the first response. LTP was induced by applying high-frequency stimulation (HFS) (five trains of eight 0.4 ms 400 Hz pulses, spaced 10 s apart). Ten measurements, 10 s apart, were taken and averaged every 5 min during 30 min pre-HFS. LTP was computed as the change in the evoked responses measured during 60 min post-HFS. Data were collected and analyzed off-line using Power Lab software.
Statistical analysis.
The results were analyzed by a two-way ANOVA for repeated measures, with the group as a between-subjects factor and the stimulus intensity/ISI/time post-HFS as a within-subject, repeated measure factor. To relate different measures, Pearson’s coefficients were calculated. Results are presented as mean ± SEM.
Results
fEPSPs were recorded from the DG in anesthetized adult rats, in response to stimulation of the PP. Baseline neural responses as well as their dynamic range were similar in the groups that were treated with MR/GR blockers, as well as in the ASS and the adrenalectomized groups (Fig. 1A,B).
Figure 1.

The dynamic range of reactivity to afferent stimulation is similar under control, stress, or drug conditions without (F(4,15) < 1) (A) and with (F(5,42) < 1) (B) stress. This indicates that there was no significant difference among the groups in the excitatory synaptic input to the hippocampus recorded in the DG.
To gain insight into possible alterations in inhibitory/excitatory interaction within the DG, we examined the responses to paired-pulse stimulation applied to the PP. The response to the second stimulus in this paradigm consists normally of an early suppression relative to the first response, caused by a fast feedback inhibitory connection, followed by a late facilitation of the response, relative to the first one. Under control conditions (Fig. 2A), MR or GR blockade did not affect paired-pulse profile along the different ISIs, compared with their control counterparts. Adrenalectomized rats and rats that were treated with the combination of GR and MR antagonists expressed a significant decrease in paired-pulse suppression at 15 ms ISI and a mild facilitation at 30 ms, compared with all other groups. A marked decrease in paired-pulse facilitation was observed at an ISI of 60 ms only in rats that were treated with spironolactone, compared with Adx and GR and MR groups.
Figure 2.

DG responses to paired-pulse stimulation applied to the PP: under control conditions, MR or GR blockade did not affect paired-pulse responses (A). B, Strikingly, after stress, there was a decrease in paired-pulse facilitation at 60 ms ISI compared with controls. GR but not MR blockade restored paired-pulse facilitation in the stressed rats (***p < 0.0001). Adrenalectomy restored paired-pulse facilitation as well, presumably because of the lack of GR activation under this condition (traces of MR block and Adx groups are shown under control and stress conditions).
Acute swim stress increased paired-pulse suppression at 30 and 60 ms ISI (Fig. 2B). GR blockade restored paired-pulse facilitation in the stressed rats, whether it was blocked alone (GR block group) or together with MR [MR plus GR (MR+GR) block and also in the Adx group, in which it could not be activated]. In contrast, spironolactone by itself had no effect on the stress-induced suppression of paired-pulse facilitation.
In response to the tetanic stimulation, control rats expressed a sustained, 63% increase in the slope of fEPSPs. GR antagonist-injected rats expressed a similar level of LTP (84.16 ± 7.64%; n = 4). In contrast, rats that were treated with spironolactone showed no LTP but only a short-term potentiation that lasted for ∼30 min (Fig. 3A). The exposure to acute swim stress resulted in a significant suppression of LTP (Fig. 3B). The application of the spironolactone before ASS led to decreased post-HFS potentiation to the extent that it reversed within 25 min into long-term depression (22.23 ± 3.42%; n = 8).In contrast, rats that were injected with the GR antagonist and subjected to ASS showed a remarkable long-lasting potentiation (228 ± 6.41%; n = 8) (Fig. 3B). Finally, rats that were treated with the combination of GR and MR antagonists or adrenalectomized rats expressed no LTP (Fig. 3C,D). Figure 3E summarizes the level of fEPSP slope potentiation 1 h post-HFS stimulation across the different conditions and treatments. It illustrates that both stress (i.e., increased level of CORT) and MR+GR blockade or adrenalectomy (i.e., functional absence of CORT) have prevented the induction of LTP. Furthermore, blocking MR alone was sufficient to block LTP. In contrast, blocking GR (particularly after the exposure to stress) resulted in a robust enhancement of LTP maintenance.
Figure 3.
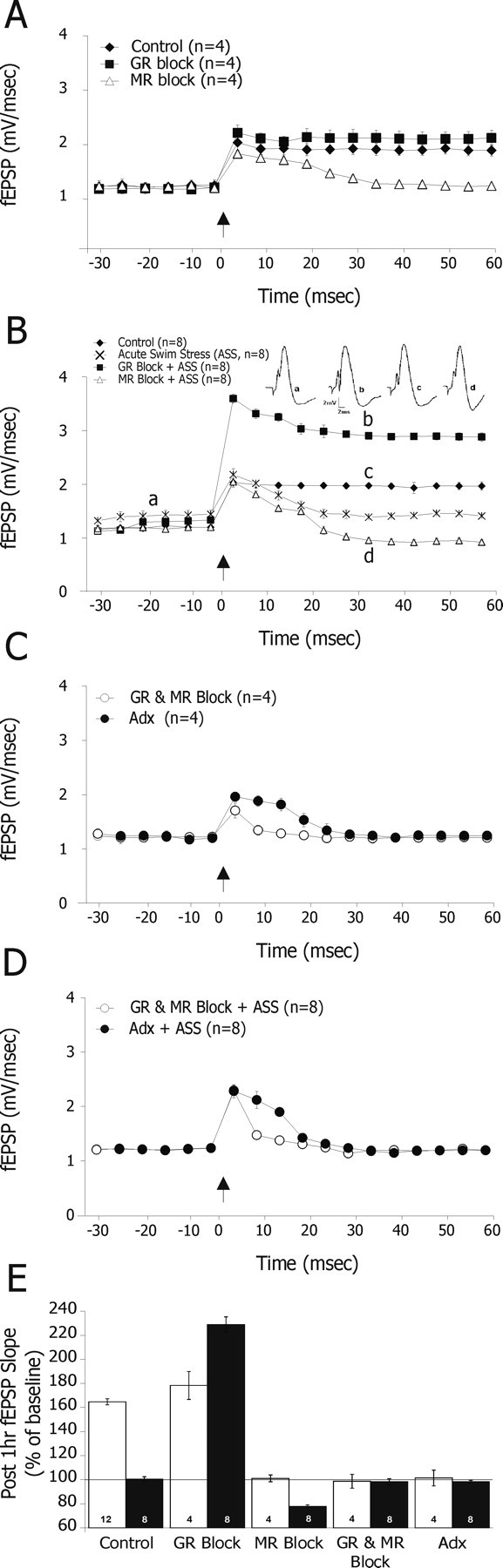
Effects of GR and MR antagonists on LTP. After one-half hour of stable baseline recording, an HFS was applied to the PP, and its consequence was recorded in the DG. A, Control rats expressed a sustained and typical, 63% increase in fEPSPs slope. Rats treated with GR antagonist (RU38486; 20 mg/kg) 75 min before HFS expressed a similar level of LTP (84.16 ± 7.64%; n = 4). In contrast, rats that were treated with the MR antagonist (Spironolactone; 20 mg/kg) showed no LTP but only a short-term potentiation that lasted for ∼30 min. B, Under stress, the difference between these two latter groups was accentuated; rats that were injected with the GR antagonist and subjected to behavioral stress (trace b) showed remarkable and long-lasting potentiation (228 ± 6.41%; n = 8) compared with their counterparts controls (trace c). Interestingly, the application of the MR antagonist before stress led to decreased post-HFS potentiation (trace d) to the extent that it reversed within 25 min to depotentiation/long-term depression (22.23 ± 3.42%; n = 8). C, D, Finally, rats that were either pretreated with the combination of GR and MR antagonists or adrenalectomized rats that showed no LTP. E, The fEPSP slope recorded 1 h post-HFS stimulation compared with baseline level was calculated in all groups. Both MR blockade and an exposure to stress blocked LTP. In contrast, GR blockade facilitated LTP, although mainly under stress.
We then calculated correlation coefficients to examine whether paired-pulse responses along the different ISIs relate to the level of potentiation observed 1 h post-HFS. A negative correlation between the level of potentiation measured 1 h post-HFS and paired-pulse responses at 30 ms ISI was found, under both control and stress conditions (rp = −0.606, p < 0.005; rp = −0.366, p < 0.011, respectively).
A positive correlation was found between paired-pulse responses at 60 ms ISI and the level of potentiation measured only under stress 1 h post-HFS (rp = 0.452; p < 0.001). Thus, the ability of pretreatment with the GR antagonist to reverse the decreased paired-pulse facilitation of stressed rats at 60 ms ISI is indicative of the remarkable potentiation level observed in these rats. Likewise, the decreased paired-pulse facilitation at 60 ms ISI after the exposure to stress or pretreatment with MR antagonist is compatible with the lack of LTP or the appearance of LTD in these rats.
Discussion
The present results demonstrate that activation of MRs is required for neural plasticity in the hippocampus. Blocking MRs (MR block, MR+GR block and Adx groups) suppressed the ability to induce LTP. Furthermore, the blockade of MRs under stress led to a decrease in post-HFS potentiation that actually reversed to long-term depression after ∼30 min. GRs, in contrast, were found to exert an inhibitory effect on the induction of LTP. After the exposure to stress, the blockade of GRs led to a remarkable long-lasting increase (approximately twofold) of LTP, suggesting that the activation of these receptors normally suppresses neural plasticity.
Although urethane is known to stimulate the activation of the HPA axis and to increase the level of circulating corticosterone (Hamstra et al., 1984; Smythe et al., 1987), urethane-induced increase in circulating corticosterone did not affect the ability to induce LTP in control animals (Shirasaka and Wasterlain, 1995; Gilbert and Mack, 1999), indicating that relatively high levels of circulating corticosterone are required to block the induction of LTP in DG.
In this study, blockers were applied subcutaneously. Therefore, possible involvement of effects on peripheral receptors cannot be ruled out. However, to the extent that similar findings were obtained in the in vitro preparations in which the blockers were obtained directly in the hippocampus (Pavlides et al., 1996), it is likely that the effects observed in the current study result from direct blockade of hippocampal MRs or GRs.
There is abundant evidence describing the modulatory effect of stress on learning and memory performance as well as on synaptic plasticity (Kim and Diamond, 2002), indicating that this effect follows an inverted U-shape function (Cahill and McGaugh, 1998; McGaugh, 2000; McGaugh and Roozendaal, 2002). Stress was found to impair LTP in CA1 area, both in vivo (Pavlides et al., 2002; Maroun and Richter-Levin, 2003; Kavushansky et al., 2006) and in vitro (Foy et al., 1987; Shors et al., 1989; Kim et al., 1996). Thompson (1992) as well as Diamond et al. (2005) hypothesized that stress and LTP share similar mechanisms and that stress-induced plasticity simply saturate the feasibility of further potentiation to be induced by HFS. However, results regarding the effects of stress on DG LTP do not easily fall into this proposed framework. Different stressors were reported to either impair (Foy et al., 1987; Diamond and Rose, 1994), enhance (Gerges et al., 2001; Kavushansky et at., 2006), or have no effect (Garcia, 2001) on the ability to induce LTP in this region.
Kortz and Frey (2003) found a bidirectional effect of behavioral stress on the maintenance of DG LTP; handling 15 min after the induction of early LTP resulted in an impairment of LTP, whereas a 2 min swim also 15 min after induction resulted in prolongation of LTP up to 24 h. Although both manipulations increased circulating CORT, handling-dependent LTP impairment was reversed by blockade of GRs but was left unaffected by blockade of MRs. Berger et al. (2006) have recently shown that the inactivation of MR gene brings about a clear acquisition deficit in the water maze task as well as in the eight-arm radial maze. These results are congruent with our current findings that MR blockade suppresses DG LTP, whereas GR blockade, particularly under stress, uncovers a larger potentiation.
This pattern of potentiation/depotentiation 1 h post-HFS correlates with paired-pulse responses at 60 ms ISI. Moreover, we found that GR blockade reversed the stress-related decrease in paired-pulse facilitation (at 60 ms ISI) after stress, an effect that was not observed under MR blockade. This apparent correlation between the effects of stress and of MR blockade on paired-pulse facilitation and on plasticity suggests that the effects on LTP are mediated by a change in postsynaptic reactivity to afferent stimulation and not by a change in probability of transmitter release.
Adrenalectomy was suggested to induce a rapid and selective neural loss in the DG (Sloviter et al., 1993). Such cell loss could potentially contribute to results observed in the adrenalectomized group. However, the dynamic range of reactivity to afferent stimulation was similar in adrenalectomized and control animals. Furthermore, paired-pulse responses of adrenalectomized animals did not differ from controls, suggesting that adrenalectomy-related apoptosis did not contribute significantly to the observed results.
Together, the results indicate that MRs and GRs assume opposite roles in regulating synaptic plasticity, particularly under stressful conditions. Previous studies have already suggested such a differential role for MRs and GRs in the hippocampus (Pavlides et al., 1995). Here, we demonstrate that the functional significance of such opposing roles is emphasized under stressful conditions. The activation of MRs is likely to be a prerequisite for plasticity to occur. However, MRs are high-affinity receptors, thus under normal conditions and certainly after an exposure to a stressor, these receptors are sufficiently occupied to enable plasticity. After stress, the level of circulating CORT increases and the low-affinity GRs may become activated and inhibit the formation of LTP. Nonetheless, increased activation of MRs, presumably together with additional neuromodulators such as noradrenaline, may yet shift the balance to enable plasticity in the DG. Thus, in the DG, the final outcome of an exposure to stress may depend on the exact characteristics of the stress experience and on the relationship between the level of activation of MRs, GRs, and other neuromodulatory systems.
Footnotes
This work was supported in part by European Union Project 512012 to G.R.-L. A.A. was a recipient of a postdoctoral fellowship from the National Institute for Psychobiology in Israel, in memory of Leah M. Smith.
References
- Alfarez DN, Wiegert O, Joels M, Krugers HJ. Corticosterone and stress reduce synaptic potentiation in mouse hippocampal slices with mild stimulation. Neuroscience. 2002;115:1119–1126. doi: 10.1016/s0306-4522(02)00483-9. [DOI] [PubMed] [Google Scholar]
- Avital A, Richter-Levin G. Exposure to juvenile stress exacerbates the behavioural consequences of exposure to stress in the adult rat. Int J Neuropsychopharmacol. 2005;8:163–173. doi: 10.1017/S1461145704004808. [DOI] [PubMed] [Google Scholar]
- Avital A, Goshen I, Kamsler A, Segal M, Iverfeldt K, Richter-Levin G, Yirmiya R. Impaired interleukin-1 signaling is associated with deficits in hippocampal memory processes and neural plasticity. Hippocampus. 2001;13:826–834. doi: 10.1002/hipo.10135. [DOI] [PubMed] [Google Scholar]
- Berger S, Wolfer DP, Selbach O, Alter H, Erdmann G, Reichardt HM, Chepkova AN H, Haas HL, Lipp HP, Schutz G. Loss of the limbic mineralocorticoid receptor impairs behavioral plasticity. Proc Natl Acad Sci USA. 2006;103:195–200. doi: 10.1073/pnas.0503878102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cahill L, McGaugh JL. Mechanisms of emotional arousal and lasting declarative memory. Trends Neurosci. 1998;21:294–299. doi: 10.1016/s0166-2236(97)01214-9. [DOI] [PubMed] [Google Scholar]
- Dallman MF, Akana SF, Jacobson L, Levin N, Cascio CS, Shinsako J. Characterization of corticosterone feedback regulation of ACTH secretion. Ann NY Acad Sci. 1987;512:402–414. doi: 10.1111/j.1749-6632.1987.tb24976.x. [DOI] [PubMed] [Google Scholar]
- de Kloet ER, Vreugdenhil E, Oitzl MS, Joels M. Brain corticosteroid receptor balance in health and disease. Endocr Rev. 1998;19:269–301. doi: 10.1210/edrv.19.3.0331. [DOI] [PubMed] [Google Scholar]
- de Kloet ER, Joels M, Holsboer F. Stress and the brain: from adaptation to disease. Nat Rev Neurosci. 2005;6:463–475. doi: 10.1038/nrn1683. [DOI] [PubMed] [Google Scholar]
- Diamond DM, Rose GM. Stress impairs LTP, hippocampal-dependent memory. Ann NY Acad Sci. 1994;746:411–414. doi: 10.1111/j.1749-6632.1994.tb39271.x. [DOI] [PubMed] [Google Scholar]
- Diamond DM, Bennett MC, Fleshner M, Rose GM. Inverted-U relationship between the level of peripheral corticosterone and the magnitude of hippocampal primed burst potentiation. Hippocampus. 1992;2:421–430. doi: 10.1002/hipo.450020409. [DOI] [PubMed] [Google Scholar]
- Diamond DM, Park CR, Campbell AM, Woodson JC. Competitive interactions between endogenous LTD and LTP in the hippocampus underlie the storage of emotional memories and stress-induced amnesia. Hippocampus. 2005;15:1006–1025. doi: 10.1002/hipo.20107. [DOI] [PubMed] [Google Scholar]
- Foy MR, Stanton ME, Levine S, Thompson RF. Behavioral stress impairs long-term potentiation in rodent hippocampus. Behav Neural Biol. 1987;48:138–149. doi: 10.1016/s0163-1047(87)90664-9. [DOI] [PubMed] [Google Scholar]
- Funder JW. Aldosterone receptors. Adv Exp Med Biol. 1986;196:145–152. doi: 10.1007/978-1-4684-5101-6_10. [DOI] [PubMed] [Google Scholar]
- Garcia R. Stress, hippocampal plasticity, and spatial learning. Synapse. 2001;40:180–183. doi: 10.1002/syn.1040. [DOI] [PubMed] [Google Scholar]
- Gerges NZ, Stringer JL, Alkadhi KA. Combination of hypothyroidism and stress abolishes early LTP in the CA1 but not dentate gyrus of hippocampus of adult rats. Brain Res. 2001;922:250–260. doi: 10.1016/s0006-8993(01)03181-x. [DOI] [PubMed] [Google Scholar]
- Gilbert ME, Mack CM. Field potential recordings in dentate gyrus of anesthetized rats: stability of baseline. Hippocampus. 1999;9:277–287. doi: 10.1002/(SICI)1098-1063(1999)9:3<277::AID-HIPO7>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- Hamstra WN, Doray D, Dunn JD. The effect of urethane on pituitary-adrenal function of female rats. Acta Endocrinol (Copenh) 1984;106:362–367. doi: 10.1530/acta.0.1060362. [DOI] [PubMed] [Google Scholar]
- Kavushansky A, Vouimba RM, Cohen H, Richter-Levin G. Activity and plasticity in the CA1, the dentate gyrus, and the amygdala following controllable vs. uncontrollable water stress. Hippocampus. 2006;16:35–42. doi: 10.1002/hipo.20130. [DOI] [PubMed] [Google Scholar]
- Kim JJ, Diamond DM. The stressed hippocampus, synaptic plasticity and lost memories. Nat Rev Neurosci. 2002;3:453–462. doi: 10.1038/nrn849. [DOI] [PubMed] [Google Scholar]
- Kim JJ, Foy MR, Thompson RF. Behavioral stress modifies hippocampal plasticity through N-methyl-d-aspartate receptor activation. Proc Natl Acad Sci USA. 1996;93:4750–4753. doi: 10.1073/pnas.93.10.4750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kortz V, Frey JU. Stress-related modulation of hippocampal long-term potentiation in rats: involvement of adrenal steroid receptors. J Neurosci. 2003;23:7281–7287. doi: 10.1523/JNEUROSCI.23-19-07281.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krugers HJ, Alfarez DN, Karst H, Parashkouhi K, van Gemert N, Joels M. Corticosterone shifts different forms of synaptic potentiation in opposite directions. Hippocampus. 2005;15:697–703. doi: 10.1002/hipo.20092. [DOI] [PubMed] [Google Scholar]
- Lupien SJ, McEwen BS. The acute effects of corticosteroids on cognition: integration of animal and human model studies. Brain Res Brain Res Rev. 1997;24:1–27. doi: 10.1016/s0165-0173(97)00004-0. [DOI] [PubMed] [Google Scholar]
- Maroun M, Richter-Levin G. Exposure to acute stress blocks the induction of long-term potentiation of the amygdala-prefrontal cortex pathway in vivo. J Neurosci. 2003;23:4406–4409. doi: 10.1523/JNEUROSCI.23-11-04406.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGaugh JL. Memory–a century of consolidation. Science. 2000;287:248–251. doi: 10.1126/science.287.5451.248. [DOI] [PubMed] [Google Scholar]
- McGaugh JL, Roozendaal B. Role of adrenal stress hormones in forming lasting memories in the brain. Curr Opin Neurobiol. 2002;12:205–210. doi: 10.1016/s0959-4388(02)00306-9. [DOI] [PubMed] [Google Scholar]
- Pavlides C, McEwen BS. Effects of mineralocorticoid and glucocorticoid receptors on long-term potentiation in the CA3 hippocampal field. Brain Res. 1999;851:204–214. doi: 10.1016/s0006-8993(99)02188-5. [DOI] [PubMed] [Google Scholar]
- Pavlides C, Watanabe Y, Magarinos AM, McEwen BS. Opposing roles of type I and type II adrenal steroid receptors in hippocampal long-term potentiation. Neuroscience. 1995;68:387–394. doi: 10.1016/0306-4522(95)00151-8. [DOI] [PubMed] [Google Scholar]
- Pavlides C, Ogawa S, Kimura A, McEwen BS. Role of adrenal steroid mineralocorticoid and glucocorticoid receptors in long-term potentiation in the CA1 field of hippocampal slices. Brain Res. 1996;738:229–235. doi: 10.1016/s0006-8993(96)00776-7. [DOI] [PubMed] [Google Scholar]
- Pavlides C, Nivon LG, McEwen BS. Effects of chronic stress on hippocampal long-term potentiation. Hippocampus. 2002;12:245–257. doi: 10.1002/hipo.1116. [DOI] [PubMed] [Google Scholar]
- Reul JM, de Kloet ER. Two receptor systems for corticosterone in rat brain: microdistribution and differential occupation. Endocrinology. 1985;117:2505–2511. doi: 10.1210/endo-117-6-2505. [DOI] [PubMed] [Google Scholar]
- Shors TJ, Seib TB, Levine S, Thompson RF. Inescapable versus escapable shock modulates long-term potentiation in the rat hippocampus. Science. 1989;244:224–226. doi: 10.1126/science.2704997. [DOI] [PubMed] [Google Scholar]
- Sloviter RS, Sollas AL, Dean E, Neubort S. Adrenalectomy-induced granule cell degeneration in the rat hippocampal dentate gyrus: characterization of an in vivo model of controlled neuronal death. J Comp Neurol. 1993;330:324–336. doi: 10.1002/cne.903300304. [DOI] [PubMed] [Google Scholar]
- Smriga M, Nishiyama N, Saito H. Mineralocorticoid receptor-mediated enhancement of neuronal excitability and synaptic plasticity in the dentate gyrus in vivo is dependent on the beta-adrenergic activity. J Neurosci Res. 1998;51:593–601. doi: 10.1002/(SICI)1097-4547(19980301)51:5<593::AID-JNR6>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- Smythe GA, Gleeson RM, Stead BH. Stimulation of the hypothalamic-pituitary-adrenal axis and inhibition of growth hormone release via increased central noradrenaline neuronal activity by urethane anaesthesia in the rat: blockade by clonidine. Aust J Biol Sci. 1987;40:91–96. [PubMed] [Google Scholar]
- Squire LR. The neuropsychology of human memory. Annu Rev Neurosci. 1982;5:241–273. doi: 10.1146/annurev.ne.05.030182.001325. [DOI] [PubMed] [Google Scholar]
- Thompson RF. Memory. Curr Opin Neurobiol. 1992;2:203–208. doi: 10.1016/0959-4388(92)90013-b. [DOI] [PubMed] [Google Scholar]