

Introduction
Sixty years ago, the ethnomycologist R. G. Wasson discovered an ancient mushroom cult in Oaxaca, Mexico.1, 2 The famous mycologist R. Heim classified the psychoactive mushroom species in the genera Psilocybe and the eminent natural product chemist Albert Hofmann published the isolation, structural elucidation and synthesis of the new alkaloids psilocybin and psilocin in 1958. These indole derivatives were also found in a number of other Psilocybe species encountered in Asia and Europe and it is now known that other psychoactive species exist in the genera Pluteus, Panaeolus, Inocybe and Gymnopilus.1, 2
Hallucinogenic mushrooms of the main genus Psilocybe are known from countries with a wet climate. Besides the cultivation of subtropical species such as Psilocybe cubensis (Earle) Singer, a range of other naturally growing species are commonly found in Europe. They deserve increasing attention because some species can yield up to thousands of fruiting bodies in relatively small spaces.1, 2
The species Psilocybe semilanceata (Fr.) Kummer contains comparatively high amounts of psilocybin (around 1% dry weight) and some baeocystin, which is the monomethyl analogue of psilocybin,3, 4 whereas the phenolic compound psilocin is only found in traces in these mushrooms (Figure 1). This small species usually grows in only scattered patterns on pasture. In recent decades, however, the psychoactive mycoflora of Europe has been transformed deeply. In 1946, E. Wakelfield described a new species from Kew Gardens (London, UK) that became known as Psilocybe cyanescens Wakefield.5, 6 This mushroom typically exhibits strong blueing behaviour after bruising and also spontaneously during ageing. This species grows on wood chips and rich compost derived from plant residues. It has been suggested that these mushrooms grew adventitiously in botanical gardens following importation of plant material from foreign countries. In 1962, this species was also found in the Pacific Northwest, USA1 and Kubicka first discovered other blueing mushrooms in woods near Sazava (Czech Republic) in 1942.1, 5 The species was named Psilocybe bohemica Sebek and further descriptions are available in the Czech mycological literature.7
Figure 1.
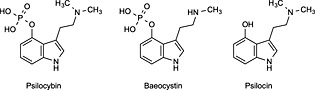
Chemical structures of psilocybin, baeocystin and psilocin. These indole alkaloids are commonly detected in a range of mushrooms of the Psilocybe genus.
Krieglsteiner shed new light a variety of mycological collections obtained from wood debris about 30 years ago5, 6 It is now clear, however, that Psilocybe cyanescens was not in this collection. One of the present authors (J. G.) has studied these mushrooms since 1986 and found genuine Psilocybe bohemica in the Czechoslovakia in 1986 and later in 1989.1 It appears that the distribution of Psilocybe cyanescens has been expanding in Germany, and possibly other European countries, during the last 30 years.1 The reason for this increasing expansion is thought to relate to increasing use of bark and wood chips used for decoration but also for preventive measures when dealing with certain weeds. Correspondingly, such locations have been observed to yield collections of up to thousands of these large and very potent mushrooms.1, 8 Additionally, the analysis of a new species from North America associated with wood chips was also described.9, 10 The unusually large and potent Psilocybe azurescens Stamets & Gartz has also been found in Germany.11 In January 1994, a new species collected from grass land in South Africa was described and later named Psilocybe natalensis by Gartz et al.12 The present case report describes the discovery of a new Psilocybe species.
Experimental
Mushroom species
The collections obtained from various species were compared with the new Psilocybe germanica and samples were deposited in the herbarium of the botanical museum in Berlin/Dahlem (B). Over the years, many hundreds of fresh fruit bodies were encountered from each species. Psilocybe bohemica: November 1986 and 1989 (near Sazava; Czech Republic) and autumn 2013 and 2014 (near Eibenstock, Saxony, Germany). Psilocybe azurescens: October 1991 and 1996 (Astoria, Oregon, USA), October 1998 (Harz Mountains, Germany), October 2000 (Rostock, Germany), autumn 2003 and 2004 (near Leipzig, Germany) and autumn 2014 (near Tharandt, Saxony, Germany). Psilocybe cyanescens: October 1991 and 1996 (Astoria, Eugene, Oregon, USA), autumn almost every year (Leipzig and other locations in Saxony, Germany since 1991). When Psilocybe germanica were discovered in autumn 2014, each of the above mentioned species were encountered at the same time. Chemical analyses of the new species were carried out using established methods previously reported in earlier studies.3, 4, 8, 9
Chemicals and solvents were from Carl Roth (Karlsruhe, Germany) and Sigma‐Aldrich (Seelze, Germany). Psilocybin standard was from Merck (Darmstadt, Germany), psilocin was from Sigma‐Aldrich and baeocystin was available from previous work following isolation Inocybe aeruginascens Babos.13 Silica gel TLC plates were obtained from Macherey‐Nagel (Düren, Germany).
Sample preparation and chemical analyses of the new species were carried out using established TLC and HPLC methods previously reported in earlier studies.4, 14 Briefly, dried, powdered mushrooms (0.1 g) were extracted with methanol (10 mL) for 12 h using a magnetic stirrer at room temperature. Mobile phases used for TLC analysis were n‐butanol:acetic acid:water (2:1:1, v/v) (mobile phase 1) (Rf psilocybin: 0.28; Rf baeocystin: 0.36; Rf psilocin: 0.51) and methanol/aqueous ammonia (25%) (100:1.5, v/v) (mobile phase 1) (Rf psilocybin: 0.10; Rf baeocystin: 0.05: Rf psilocin: 0.39). The methanol extracts were analyzed using each TLC mobile phase systems. For detection, p‐dimethylaminobenzaldehyde (1 g in 20 mL solvent obtained from 5 mL phosphoric acid in 15 mL methanol) was employed. High‐performance liquid chromatography analysis employed UV detection at 267 nm using a LiChrosorp RP‐18 column (250 × 4.6 mm, 5 µm) at a flow rate of 2.0 mL/min. The mobile phase was based on a citrate‐phosphate buffer and ethanol (9:1) (300 mL 0.1 M citric acid and 160 mL 0.1 M NaH2PO4). The Injection volume was 10 μL.
Results and discussion
Latin diagnosis
Whenever a new species is encountered, the provision of a Latin description and diagnosis is required and provided as follows.
Psilocybe germanica Gartz & Wiedemann, sp.nov.
Species sectionis Caerulescentium
Pileus cum largo umbone, calvus,symmetrus, in senectute solum raro curvatus quodam modo in altitudinem, margo non undosus, cutis pilei detrahi non potest, in statu umido fuscus et quodam modo fluidus, hygrophanus, in statu umido non perlucidus, non canaliculatus in statu arido albus, tum umbo plerumque, sua sponte colorem canum vel caeruleum accipit, in pressione vel sua sponte in senectute passim colorem caeruleum accipit, caro in pressione colorem caeruleum accipit, reliquiae veli non exsistunt, a‐4 cm in latitudinem, odor iucunde aromaticus.
Lamellae primo coloris clare fusci, tum in senectute coloris purpurei vel fusci, dense positae, semper alternantes adnexa adnatam sequens.
Stipes albus, semper curvatus, saepe multipliciter, 5–9 cm in longitudinem, 3‐ 7 mm in latitudinem, etiam adulescens velum partiale cum velamine non habens, orbem non habens, tenax et flexibilitis, non valde durus, stamina subtilia mycelis non habens in superficie, interdum densae vel firmae conglomerationes, quae in iniquitate consistunt, videri possunt; in basi stipitis saepe rhizomorphae, quae reliquias ligni stricte continent, in senectute cavus, in pressione celerrime et ubique obscure caerulescens; in senectute etiam sua sponte, interdum in pressione incipiente quodam modo virescens, tum semper celeriter obscure caerulescens; typica magna crassificatio ad acumen vel paulum sub acumine; fungorum adulescentium crassitudo circiter duplo maior quam reliqui stipitis, interdum nodosissimo genere; in senectute crassificatio ut artus adnirationem movet, cum pileus et pars stipitis superior valde infrigitur; crassificatio intestinae partis consistit in strato quasi lignoso et iam pridem disposito, in pressione non caerulescente.
Sporis ellipsoideis, 7–12 / 5.5–7.5 µm, poro germinativo praeditis, cheilocystidiis numerosis, lageniformibus, 25–33 / 6.0–8.2 µm, pleurocystidiis nullis
Crescit in reliquiis lignorum, quae tabe diverse progrediente afficiuntur; etiam in cortice et in diversis arborum fruticumque generibus; in Germania tempus crescendi imprimis de mense Septembri usque ad Decembrem, in quendam modum patiens brevis frigoris nocturni; incrementum etiam in aliis terris exspectari potest. Holotypus in herbario (B), prope Dippoldiswalde, 8th October 2014.
Description of the new species Psilocybe germanica Gartz & Wiedemann
Pileus: 1–4 cm in diameter, always broadly umbonate, rarely flattening to a little turn up in age with a persistent broad umbo; no traces of a veil at any stages, hygrophanous, moist deep brown, lacking a separable pellicle, margin not wavy, not striate, not translucent when moist, fading in drying to whitish; during development soon a spontaneous gray‐bluish colouration on the umbo occurs, at age an additional strong blueing develops, particularly after rains and during freezing; bruising of the white flesh also yields a strong blue colour, odour pleasantly aromatic.
Lamellae: at first brownish then dark purple‐brown, closely set, alternating adnexed to adnate.
Stipes: 5–9 × 0.3–0.7 cm, characteristic enlarged at the pileus, sometimes the thickening often impressed as a joint, in which the upper end of the handle with the cap then drops; dry, white, not very hard, not flexuous, always curved, often multiple times, without traces of a veil, no fine mycelia on it, sometimes solid lumps visible as a rub, easily staining deep blue when touched at all parts; at first touch sometimes green discoloring until a very fast‐moving to deep blue; the wood‐like layer in the thickening shows no blueing at all, stuffed with white mycelia at first, later hollow, rhizomorphs on the base, keeping wood substrate together.
Microscopic features: spores: 9–12 × 5.5–7.5 µm, with a clear germ pore, cheilocystidia lageniform, numerous, 25–33 / 6.0–8.3 µm, pleurocystidia absent.
Habit and distribution: Gregarious to cespitose on deciduous on wood chips from various plants, also on bark in mixture with soil and other wood debris. Observed growing is from September to December. Until now, the mushrooms have only been observed to grow in parks. Holotypus: a gathering (8th October 2014, Dippoldiswalde, Germany) is deposited in the herbarium of the botanical museum Berlin / Dahlem (B).
The new Psilocybe germanica showed a unique combination of features related to stipes and pileus. Another species with such joint‐like thickening has not yet been described until now. As can be seen in Figure 2 there are very clear differences in comparison to Psilocybe azurescens (Figure 2A) and Psilocybe cyanescens (Figure 2B). In contrast to the new species (Figure 2C), both other species show yellow caps after drying. They show a cortina, which left traces on the caps and the stipes in age of the mushrooms. The margin shows a striate structure when wet. The structure of the stipes found in the new species was also different, i.e. the stipe seen in Psilocybe germanica appeared softer. As a consequence, the blueing reaction is more readily observed compared to stipes of Psilocybe azurescens and Psilocybe cyanescens. The wavy appearance is only characteristic for Psilocybe cyanescens.
Figure 2.

A: Psilocybe azurescens. B: Psilocybe cyanescens. C: The new species Psilocybe germanica.
Figure 3 shows a large fruiting of the new species after a short freezing during the night. Exposure to cold temperature resulted in a strong blueing reaction, sometimes across the entire mushroom. Figure 4 shows various forms and developmental stages of the new species. Compared to Psilocybe bohemica (Figure 5), significant differences were observed. In contrast to Psilocybe germanica, this species had a striate and translucent cap when wet. The caps are deep yellow when wet and often whitish in colour when dry, with some blue zones that do not show any particular structure. In most cases, the caps are not umbonate upon ageing. Whereas Psilocybe bohemica display an easily detectable cortina and stipes that are scattered with fine fibrils, the new species Psilocybe germanica exhibits stipes of more regular shapes that are not curved and occasionally enlarged at the base. In contrast to the new species, Psilocybe bohemica can build up only a few spores in age of the fruiting bodies and often the mushrooms are completely sterile.5, 6
Figure 3.

Large fruiting of Psilocybe germanica. Exposure to cold temperature resulted in a strong blueing reaction, sometimes across the entire mushroom.
Figure 4.

Various forms and developmental stages of the new species Psilocybe germanica.
Figure 5.

Psilocybe bohemica. In contrast to Psilocybe germanica, a striate and translucent cap were observed when wet.
The tiny species Psilocybe serbica Moser & Horak showed features for caps and stipes similar to Psilocybe bohemica.15 The microscopic features are comparable to other wood‐loving Psilocybe species2, 5, 6, 9, 10, 11 as is the case with Psilocybe germanica. Chemical analyses of five Psilocybe germanica mushrooms samples revealed the presence of significant amounts of psilocybin and baeocystin (Table 1). This was consistent with qualitative TLC analysis of 30 samples from 2013 and 35 extracted samples from 2014 and psilocin was not detected (data not shown).
Table 1.
Amounts of indole alkaloids detected in five Psilocybe germanica samples and results obtained from previous analyses (%, dry weight)
| Sample | Psilocybin | Baeocystin | Psilocin |
|---|---|---|---|
| 1 | 0.82 | 0.15 | nd * |
| 2 | 0.90 | 0.11 | nd |
| 3 | 0.76 | 0.18 | nd |
| 4 | 0.66 | 0.21 | nd |
| 5 | 1.12 | 0.30 | nd |
| P. cyanescens 16 | 0.34 − 0.98 | 0.01 − 0.05 | 0.01 − 0.93 |
| P. cyanescens 17 | 0.25 − 0.85 | 0.01 − 0.03 | 0.04 − 0.36 |
| P. semilanceata 3, 4 | 0.19 − 1.34 | 0.03 − 0.38 | nd |
| P. semilanceata 18 | 0.55 − 1,96 | 0,11 − 034 | − |
| P. semilanceata 19 | 0.41 − 1.41 | 0.10 − 0.60 | traces |
| P. semilanceata 14 | 0,91 − 1,05 | qualitative | nd − 0.02 |
| P. semilanceata 17 | 0.05 − 1.70 | nd − 0.36 | nd − 0.36 |
| P. bohemica 14 | 0.25 − 1.14 | nd | nd − 0.07 |
| P. bohemica 17 | 0.28 − 0.80 | 0.01 − 0.03 | nd − 0.02 |
| P. bohemica 20 | 0.11 − 1,35 | 0.01 − 0.03 | 0.01 − 0.02 |
| P. azurescens 9 | 1.17 − 1.78 | 0.19 − 0.41 | 0.19 − 0.49 |
nd: not detected.
Psilocybe germanica showed similar alkaloid levels to what is known about Psilocybe semilanceata with only traces of psilocin. Psilocybe azurescens is generally considered more potent and, in addition to psilocybin and baeocystin, is also displaying comparatively large amounts of psilocin.9 Both Psilocybe cyanescens and Psilocybe bohemica typically show the presence of psilocybin and traces of baeocystin. A large variation in psilocybin levels are frequently encountered. In contrast to Psilocybe bohemica,16 Psilocybe cyanescens can also contain large amounts of psilocin.16 In addition, the new indole derivative aeruginascin13, 21 was not detected in any of the mushrooms. As far as it is currently known, this trimethylammonium analog of psilocybin has only been reported to occur in the hallucinogenic species Inocybe aeruginascens Babos.13, 21 Psilocybe semilanceata is the best studied psychoactive mushroom species in the world1, 14, 17, 18, 19, 22, 23 and is known to contain psilocybin and baeocystin (Table 1). Previous investigations in 1994 have shown that the use pure methanol yielded the authentic indole derivatives from dried mushrooms and the use of aqueous solvents resulted in hydrolysis of psilocybin and baeocystin to give psilocin.4
Conclusion
Psilocybe germanica sp.nov. was described as a new taxon from Germany. This species was found to be autumnal and lignicolous, living in soils enriched with deciduous wood‐debris, and featured strong blueing behaviour after bruising and ageing. It contained high amounts of psilocybin and baeocystin. This mushroom appears to be the first described species derived from wood chips that contains significant amounts of baeocystin in the absence of psilocin. The novel species was differentiated from other psychoactive taxons such as Psilocybe cyanescens, Psilocybe azurescens and Psilocybe bohemica in combination from features of the stipes and caps. As it was observed previously with the potent psychoactive species Psilocye cyanescens, it is expected that Psilocybe germanica might be found to achieve a remarkably wide area of distribution in the future given the modern use of mulch in parks and gardens.
Gartz, J. , and Wiedemann, G. (2015) Discovery of a new caerulescent Psilocybe mushroom in Germany: Psilocybe germanica sp.nov.. Drug Test. Analysis, 7: 853–857. doi: 10.1002/dta.1795.
The copyright line for this article was changed on 30 April 2015
References
- 1. Gartz J.. Magic Mushrooms Around the World. A Scientific Journey Across Cultures and Time, Arnshaugk, Neustadt/ Orla, Germany: 2014. [Google Scholar]
- 2. Stamets P.. Psilocybe Mushrooms of the World. An Identification Guide, Ten Speed Press, Berkeley, CA, USA: 1996. [Google Scholar]
- 3. Gartz J.. Further investigations on psychoactive mushrooms of the genera Psilocybe, Gymnopilus, Conocybe . Ann. Mus. Civ. Rovereto Sez. St. Sc. Nat. 1991, 17, 265. [Google Scholar]
- 4. Gartz J.. Extraction and analysis of indole derivatives from fungal biomass. J. Basic Microbiol. 1994, 34, 17. [DOI] [PubMed] [Google Scholar]
- 5. Krieglsteiner G. J.. Studien zum Psilocybe cyanescens – Komplex in Europa. Beiträge zur Kenntnis der Pilze Mitteleuropas. 1984, 1, 61. [Google Scholar]
- 6. Krieglsteiner G. J.. Studien zum Psilocybe cyanescens‐ callosa‐ semilanceata‐ Komplex in Europa. Beiträge zur Kenntnis der Pilze Mitteleuropas. 1986, 2, 57. [Google Scholar]
- 7. Semerdzieva M., Veselsky J.. Léčivé Houby Dříve a Nyní, Academia, Prague: 1986. [Google Scholar]
- 8. Gartz J.. Anmerkungen zu den blauenden Psilocybe‐ Arten von Holzresten in Europa. Südwestdeutsche Pilzrundschau. 2014, 50, 31. [Google Scholar]
- 9. Stamets P., Gartz J.. A new caerulescent Psilocybe from the Pacific Coast of Northwestern America. Integration. 1995, 6, 21. [Google Scholar]
- 10. Stamets P., Gartz J.. A new caerulescent Psilocybe from the Pacific Coast of Northwestern North America. Mycotaxon. 1995, 11, 476. [Google Scholar]
- 11. Gminder A.. Psilocybe azurescens Stamets & Gartz. Erste Freilandfunde in Europa. Südwestdeutsche Pilzrundschau. 2001, 37, 31. [Google Scholar]
- 12. Gartz J., Reid D., Eicker A., Smith M. T.. Psilocybe natalensis sp.nov. ‐ The first indigenous blueing member of the Agaricales of South Africa. Integration. 1995, 6, 29. [Google Scholar]
- 13. Jensen N., Gartz J., Laatsch H.. Aeruginascin, a trimethylammonium analogue of psilocybin from the hallucinogenic mushroom Inocybe aeruginascens . Planta Med. 2006, 72, 665. [DOI] [PubMed] [Google Scholar]
- 14. Semerdzieva M., Wurst M., Koza T., Gartz J.. Psilocybin in Fruchtkörpern von Inocybe aeruginascens . Planta Med. 1986, 52, 83. [Google Scholar]
- 15. Moser M., Horak E.. Psilocybe serbica spec.nov., eine neue Psilocybin und Psilocin bildende Art aus Serbien. Zeitschrift für Pilzkunde. 1968, 37, 137. [Google Scholar]
- 16. Gartz J.. Observations on the Psilocybe cyanescens complex of Europe and North America. Ann. Mus. Civ. Rovereto. 1996, 12, 209. [Google Scholar]
- 17. Stijve T., Kuyper T. W.. Occurrence of psilocybin in various higher fungi from several European countries. Planta Med. 1985, 51, 385. [DOI] [PubMed] [Google Scholar]
- 18. Christiansen A. L., Rasmussen K. E.. Analysis of indole alkaloids in Norwegian Psilocybe semilanceata using high‐performance liquid chromatography and mass spectrometry. J. Chromatogr. 1982, 244, 357. [Google Scholar]
- 19. Brenneisen R., Borner S.. The occurrence of tryptamine derivatives in Psilocybe semilanceata . Z. Naturforsch. 1988, 43C, 511. [Google Scholar]
- 20. Gartz J., Müller G. K.. Analysis and cultivation of fruiting bodies and mycelia of Psilocybe bohemica . Biochem. Physiol. Pflanzen. 1989, 184, 337. [Google Scholar]
- 21. Gartz J.. Psychedelika ‐ Historisch betrachtet, Arnshaugk, Neustadt/Orla, Germany: 2014. [Google Scholar]
- 22. Petersen‐Bjergaard S., Sannes E., Rasmussen K. E., Tønnesen F.. Determination of psilocybin in Psilocybe semilanceata by capillary zone electrophoresis. J. Chromatogr. B Biomed. Sci. Appl. 1997, 694, 375. [DOI] [PubMed] [Google Scholar]
- 23. Andersson C., Kistinsson J., Gry J.. Occurrence and use of hallucinogenic mushrooms containing psilocybin alkaloids. Nordic Council of Ministers, Copenhagen, 2009.