

Abstract
The neuromuscular junction (NMJ) is a unique, specialized chemical synapse that plays a crucial role in transmitting and amplifying information from spinal motor neurons to skeletal muscles. NMJ complexity ensures closely intertwined interactions between numerous synaptic vesicles, signaling molecules, ion channels, motor neurons, glia, and muscle fibers, making it difficult to dissect the underlying mechanisms and factors affecting neurodegeneration and muscle loss. Muscle fiber or motor neuron cell death followed by rapid axonal degeneration due to injury or disease has a debilitating effect on movement and behavior, which adversely affects the quality of life. It thus becomes imperative to study the synapse and intercellular signaling processes that regulate plasticity at the NMJ and elucidate mechanisms and pathways at the cellular level. Studies using in vitro 2D cell cultures have allowed us to gain a fundamental understanding of how the NMJ functions. However, they do not provide information on the intricate signaling networks that exist between NMJs and the biological environment. The advent of 3D cell cultures and microfluidic lab-on-a-chip technologies has opened whole new avenues to explore the NMJ. In this perspective, we look at the challenges involved in building a functional NMJ and the progress made in generating models for studying the NMJ, highlighting the current and future applications of these models.
Neuromuscular Junction
The neuromuscular junction (NMJ) is a large cholinergic synapse responsible for muscle functioning. Any structural changes or degeneration of this vital junction could result in motor neuron cell death and muscle atrophy. The NMJ comprises three integral components working in tandem—a presynaptic motor neuron terminal, the intrasynaptic basal lamina1 in the synaptic cleft, and the postsynaptic muscle membrane.2 In response to stimuli, activated multimeric neuronal voltage-gated channels open, leading to a calcium influx into the cell. The calcium induces the acetylcholine (ACh)-filled synaptic vesicles to fuse with the neuronal cell membrane, releasing the neurotransmitter into the synaptic cleft. ACh binds to the highly dense nicotine acetylcholine receptors (nAChR) on the post synaptic muscle leading to the depolarization of the membrane through activation of the sodium channels. This depolarization further activates voltage-gated L-type calcium channels and ryanodine receptors in the muscle, triggering calcium release from the sarcolemma. The released calcium binds to troponin C, inducing the motion of the motor protein elements. Figure 1 shows a schematic of synaptic transmission at the NMJ.
Figure 1.

The NMJ: influx of calcium (Ca2+) into the motor neuron leads to release of the neurotransmitter, ACh into the synaptic cleft. ACh binds to the receptors on skeletal muscle membranes opening up ion channels, leading to muscle depolarization and contraction.
Even though the preformed AChR clusters on the post synaptic membrane may have a role in determining where innervations of muscle fibers occur,3 studies have shown that the extending axons of motor neurons form synapses at a different location.4 There is species-dependent pre- and post-synaptic differentiation with differences observed in NMJ formation between rodents and zebrafish.5 Many parallel signaling pathways are involved in generating a functional motor unit. Post synaptic maturation and AChR clustering is also regulated by factors such as neuregulin 1 and WNT ligands and receptors such as ERBB.6
The NMJ is further complicated by the different types of motor neurons and muscle fibers involved (Figure 2). The α, β, and γ lower motor neurons are of varying sizes, and they innervate different muscle fibers (slow- and fast-twitch muscle) to form slow, fast fatigable, and fast fatigue-resistant motor units.7 The relatively smaller number of motor neurons also necessitates one motor neuron innervating several muscle fibers. For instance, the 400 motor neurons innervating the gastrocnemius muscle supplies approximately 300 000 muscle fibers.8
Figure 2.

The complexity involved in a functional NMJ: the complexity of the NMJ is highlighted here, indicating how species, muscle type, synapse location, type of neurotransmitters, glial cells, and signaling pathways all play a role in defining function and development.
The NMJ was initially explored as a synapse to understand synaptic plasticity, development, and function due to its size and easy access. However, the rising prevalence of neurodegenerative and muscular disorders also necessitated a model to study the pathophysiology underlying diseases, such as amyotrophic lateral sclerosis (ALS) and myasthenia gravis, where the denervation at the NMJ results in the weakening of limbs and increasing unsteadiness in gait.9 Understanding the pathogenesis of these disorders at the NMJ is hence critical for early diagnosis and effective treatment. This pressing imperative to investigate new treatment strategies underlines the need for better models that can provide some much needed insights into the mechanisms involved in NMJ disruption.
Advances in healthcare have led to shifts in age demographics with the percentage of the older population increasing each year. Age-associated degeneration of the NMJ has thus increased the urgency for the development of robust NMJ models to understand the mechanism involved in motor neuron death and muscle loss apart from evaluating novel therapeutic interventions for the same. Fabrication of NMJ has also been aimed toward tissue engineering the stretch reflex arc and may have applications in the field of robotics and prosthetics.10
Over the last few decades, the NMJ has been extensively studied in animal models, primarily to understand synaptogenesis.5,9 However, while these studies have resulted in better clarity, models such as the SOD1 mice for ALS have not translated well into effective treatment for human ALS, calling into question their efficacy.11 Although animal models have been at the forefront of research into the NMJ,9,12 it also presents several impediments in the form of ethical issues involving animal welfare, limited time points to study developmental regulation, and the influence of other factors such as migrating cells, disease, organ failure, and immune response13 in the test animal. The failure to adequately translate animal research to clinical benefits14 gives impetus to explore other high throughput in vitro techniques. It is further challenging to examine the NMJs of humans during the early stages of the disease or over long periods of time due to absence of reliable biomarkers and invasive biopsies. Researchers have tried to overcome this hurdle by trying to recreate the NMJ using in vitro models for a more high-throughput analysis that includes understanding disease pathways, creating therapeutic targets, drug discovery, and toxicology. The advent of stem cells15 and human induced pluripotent stem cells (iPSCs) provide attractive opportunities to study the human NMJ using in vitro models.16
The role of signaling factors and signaling pathways, cell types, receptors and mechanisms involved, changes that occur with age and disease, maturation at both the pre- and post-synaptic terminals, and the varied responses to drugs are still subject to debate.6 This is especially true with respect to the developmental regulation at the NMJ and the structural changes that accompany aging or disease. Early in vitro studies involved 2D motor neuron–skeletal muscle dissociated cocultures and organotypic cultures to simplify the system under study and to augment and complement the accompanying animal research into the NMJ. The development of specialized lab-on-a-chip (LOC) and microfluidics technology has created high-throughput in vitro models enabling investigators to narrow down, isolate, and study the NMJ at the cellular and molecular levels. More recently, the advent of three dimensional (3D) bioprinting has led to the fabrication of the NMJ in tissue-like environments, closely mimicking the NMJ development and functionality found in vivo.
An important facet of NMJ in vitro models and one of the reasons for the paucity of work in this area is the complexity involved in culturing two different cell types harvested from different environments in vivo. Both cell types often require different culture conditions (culture medium, growth, and differentiation factors) when cultured separately, and this complexity renders it difficult to create these NMJ in vitro models. The differing mechanical properties of muscle and neuron, for example, would require the right culture substrate which in turn should allow for neuronal process outgrowth and muscle differentiation.17
The following sections examine past and current techniques used to study the NMJ, and trace the progress made toward building high-throughput 3D in vitro NMJ models for research into synaptogenesis, disease, aging, and drug discovery. We discuss the novel results that research in this area has yielded, highlight the advantages that microfluidic tools add to in vitro models, and propose ways to further bolster such systems, bringing it closer to in vivo conditions.
2D in Vitro NMJ Models
The functional significance of the NMJ became the focus of interest at the turn of the 20th century when Harrison observed the contraction of a myotome still left attached to a piece of medullary cord in a frog embryo tissue culture. Early research involved removing sections of the embryonic spinal cord from test animals with the muscle still attached to the developing ganglia. Studies of the NMJ using in vitro methods commenced with an examination of the nerve–muscle connections in the spinal cord–somite regions of a chick and mouse embryo.18 These studies gave important insights into the development and function at post- and pre-synaptic structures including synaptic vesicles. Organotypic cocultures are still used to explore the NMJ. Nerve–muscle coculture models have pointed to the muscle as being a therapeutic target for degenerative diseases such as spinal muscular atrophy (SMA). In the case of patients suffering from ALS, experiments conducted using the patient serum revealed no neurotoxicity in organotypic cocultures.19
Later research into 2D NMJ models involved culturing dissociated motor neurons and muscle cells on surfaces coated with extracellular matrix (ECM) proteins and covered with cell culture medium. Figure 3 shows a simple schematic of a 2D coculture of motor neurons and muscle cells using this strategy.
Figure 3.
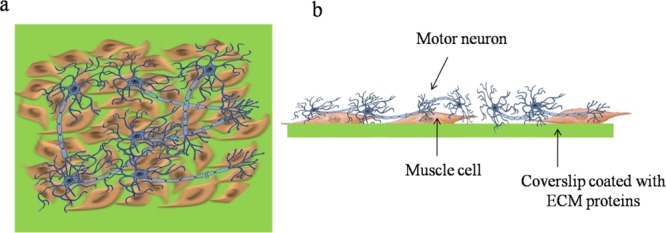
2D cocultures of dissociated motor neuron and muscle cells. (a) 2D culture: top view of muscle myocytes and motor neurons plated on a coverslip. The motor neurons are usually cultured on top of the muscle cells, which are seeded on a coverslip coated with ECM proteins. (b) 2D culture: side view of muscle cells and motor neurons seeded on a coverslip.
Disease pathology has also been studied using in vitro 2D NMJ models. Cultures of dystrophic embryonic myotubes and neurons have been employed to understand dystrophy. Human skeletal muscle from children with Duchenne muscular dystrophy was found to form stable, mature, functional motor end plates with fetal mouse spinal cord explants but displayed lesions after 2–4 months in culture in contrast to normal muscle20
In vitro NMJ models provide the opportunity to use stem cells, genetically modified cells, and human cells, finding possible applications in drug discovery, personalized medicine, and for exploring disease pathology. Motor neurons derived from multipotent spinal cord precursor cells have formed synaptic junctions with skeletal myocytes.21 Murine HB9::GFP transgenic embryonic stem cells, differentiated into motor neurons, formed NMJs in a coculture with myotubes.22 Motor neurons differentiated from human spinal cord stem cells were first shown to form synapses with embryonic rat skeletal myotubes under serum-free conditions15 in a preliminary study. A later study resulted in the development of a unique in vitro human NMJ from human spinal cord stem cells and human skeletal muscle stem cells in a defined environment.23 Human mesenchymal cells that were genetically engineered to express enhanced green fluorescent protein and the motor neuron associated transcription factors Olig and HB9 could be induced to become motor neuron-like cells that formed functional motor units with differentiated C2C12 myotubes.24
Though a plethora of reports are available on 2D models for NMJ, they do not perfectly mimic the native tissue architecture. Moreover, most of the studies were performed on static cultures that do not reproduce the dynamic flow conditions encountered in biological systems. Long-term maintenance of the 2D cultures also remains a challenge. Improved models of the NMJ need to be developed to overcome these issues. Micro-electro-mechanical systems (MEMS)-based systems offer a dynamic localized environment to probe the events occurring at the NMJ.
MEMS & Microfluidic Tools: Compartmentalized In Vitro NMJ Models
Tissue engineering applications that have been explored till date for the study of the NMJ include regenerative strategies for tackling disease conditions such as dystrophy and building prosthetic devices. Recent research has focused on extending in vitro NMJ models toward building the stretch reflex arc10 where the E18 rat skeletal muscle innervated by DRG neurons and grown on surface-modified MEMS cantilever devices in a defined system demonstrated functional interaction between muscle and sensory neurons.25 Contractile function was quantified using a laser and photo-detector unit to sense cantilever deflection from motor neuron innervated muscle twitching.
The advent of devices incorporating microfluidics technology has allowed for a compartmentalized study on the effects of different stimuli. With precise control over the cell micro-environment including diffusion and fluid flow, microfluidics presents a novel platform to study the NMJ. Motor neurons and muscle in two different chambers connected by axons in the microchannels, enabling spatiotemporal control over the cells,26 help in studying localized actions at the soma, along the axon, and at the NMJ. Microtunnels or microgrooves were initially used to study hippocampal neurons and were plated on microelectrode arrays to study parameters such as the propagation speed and direction of action potentials.27 Campenot first demonstrated independent control of the proximal and distal fluid environment of the cell soma and neurites to understand the effect of nerve growth factor, leading to the establishment of Campenot chambers.28 Microfluidic devices allowed for the growth of neurons in wells or chambers, with axonal growth along narrow tunnels 10 μm wide and 3 μm high to a second, isolated chamber, allowing for a separation of cell body and axons.29 The dimensions of the microchannels limit the size of the neurites entering it and restrict cell bodies from crossing over to other chambers while allowing for communication between the two chambers through their projections.
The fabrication of these microfluidic devices is relatively easy,30 generally made using poly(dimethylsiloxane) (PDMS) and are now available commercially.31 Such devices have proved to be effective for the visualization and manipulation of synapses32 and to study retrograde33 and anterograde34 axonal transport. Central nervous system injury was investigated using microfluidic platforms by exploring the role of myelin-associated inhibitors in axonal regeneration and quantifying the time taken for axonal growth in response to these soluble factors.35 Myogenic differentiation in response to chemical,36 electrical,37 and mechanical stimuli38 has been studied using microdevices. The ability to enforce geometric constraints on cells39 and provide chemical guidance cues40 allows for control on growth, differentiation, movement, and alignment.
One of the first studies toward a functional NMJ, incorporating microcompartments and microconduits on microelectrode arrays, involved rat cervical ganglion neurons and ventricular myocytes.41 Synaptic pathways between the two cell types were confirmed by stimulating and recording evoked electrical responses using the microelectrodes. One of the studies, looking specifically at the synaptic innervation of skeletal muscle using microdevices, was reported by Park et al. In this work, a microfluidic platform with two microchambers (6.35 mm × 12.75 mm × 4.76 mm) connected by 100 parallel microchannels (10 μm wide and 500 μm long) was developed for a functional NMJ between mouse embryonic stem cell-derived motor neurons and C2C12 myotubes.42 The coculture was maintained in neuronal culture medium with supplementation of growth factors for axonal growth. Seven days after the axons crossed through the micro-tunnels to the second chamber, NMJs were identified by staining for bungarotoxin, which binds to nAChR and helps visualize axons anchored on myotubes. In order to mimic the complex in vivo cellular interactions, embryonic rat spinal motor neurons were cocultured with spinal glial cells from postnatal day 2 (P2) rat pups in the poly-l-laminin-coated proximal chamber of a microfluidic device.43 As axons started extending into the distal chamber, the P2 rat hind limb skeletal muscle myocytes were plated in the collagen-coated chamber and a serum-free medium was used for both cell types. The plating of specific cell types in the proximal and distal chambers in both these studies were timed such that the neurites from motor neurons can reach mature differentiated myotubes that can form NMJs.
The applications of microfluidics in studying the NMJ were highlighted by Zahavi et al.44 Using a compartmentalized microfluidic device with microchannels that enable bidirectional communication between motor neurons and muscle, the group was able to demonstrate that trophic factors such as glial derived neurotrophic factor (GDNF) act differently at the axon and on the soma, having spatially distinct effects. Motor neurons from spinal cord explants of the transgenic HB9::GFP mice and the FM-4-64 (N-(3-triethylammoniumpropyl)-4-(6-(4-(diethylamino)phenyl)hexatrienyl)pyridinium dibromide) dyes were used to enable visualization of vesicular release at the sites of NMJ, whereas myoblasts infected with lentiviral vectors, GDNF–mCherry (GDNF tagged with red fluorescent protein mCherry) were used to visualize retrograde GDNF transport from muscle to neuron for the first time. This work underlines the significance of in vitro microfluidic models in investigating the molecular events at the NMJ.
3D in Vitro NMJ Models
3D cell culture systems have become more relevant over the last decade as they closely mimic in vivo conditions as opposed to routine 2D cultures Figure 4.
Figure 4.
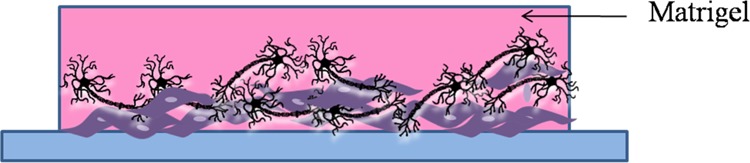
3D culture: side view of muscle cells and motor neurons cocultured in matrigel. The muscle cells are usually allowed to differentiate into muscle fibers in the matrigel before the motor neurons are added on top of them and differentiated to form neuron-muscle contacts.
Typically, 3D cocultures to study the NMJ involve culturing muscle cells in hydrogels like Matrigel or ECM matrices and then differentiating motor neurons over them. 3D cell cultures tend to be more functionally relevant with respect to contractile muscle. Extensive work has been done to tissue engineer skeletal muscle constructs using 3D cultures.45 Collagen I is a major component of native ECM and has adequate mechanical properties needed for contraction and force generation. Collagen gel constructs have been used to study early skeletal muscle development and muscle aging.46 Collagen, however, has less stability and degrades faster, proving to be ineffective for long-term cultures.47 Matrigel, a commercially available decellularized ECM, allowed for better differentiation of muscle progenitor cells compared to collagen.48 Other attractive alternatives to collagen are fibrinogen, thrombin, or fibrin. Silk fibroin has even better mechanical strength and stability when compared to collagen and may be another promising material for 3D cultures to mimic the NMJ.49 Tissue-engineered muscle has been generated from myotubes using fibrin gels, where cell-mediated contraction evoked response to electrical stimuli.50
A more recent study has fabricated three-dimensional human skeletal muscle tissue using myoblasts incorporated into a hydrogel mixture that contains thrombin and fibrinogen and seeded onto a custom-made device used to provide the uniaxial stress stimulation.51 The myoblasts are separately differentiated as human embryonic stem cells into post-mitotic motor neurons, which are maintained in Matrigel and cultured in a medium containing specific differentiation factors such as insulin. Once the differentiated motor neurons formed clusters, they were transferred to the hydrogel muscle suspensions and the culture medium was accordingly changed and supplemented with growth factors. This study added to the mounting evidence that a 3D environment aids in the maturation and alignment of mature fibers, in the increased presence and clustering of ACh receptors, the upregulation of muscle markers such as myosin heavy chain, and spontaneous electrical activity accompanied by twitching. Another similar study explored a long term (35 days) human skeletal muscle coculture along with iPSC-derived motor neurons in 3D collagen constructs, where the motor neurons maintained their cholinergic phenotype for around 21 days.52
3D neuronal cultures should enable the growth, differentiation, and proliferation of neuronal cells, allow for and guide neurite outgrowth, and induce the formation of functional synapses. The culture of both skeletal muscle in 3D environments and motor neuron cells in proximity to skeletal muscles cells have demonstrated improved maturation, contraction outputs, and upregulation of muscle and neuronal markers.53 Muscle-derived cells differentiate, fuse, and align in 3D collagen constructs as per the physiological architecture found in vivo and axonal growth cones from motor neurons were able to move through the collagen toward the myotubes and form synaptic structures.53 In a similar study, primary muscle-derived cells and mixed ventral horn cells were cultured on 3D fibrin constructs to create a tissue-engineered NMJ. Muscle function of the NMJ construct was assessed using electrical stimulation, and the upregulation of muscle and neuronal markers such as desmin, MAP-2, antisynaptic vesicle protein-2 (SV-2), and myosin heavy chain were assessed using confocal microscopy and reverse transcription polymerase chain reaction (RT-PCR).54 In all these studies, the muscle-derived cells are allowed to grow, differentiate, and become confluent before the spinal motor neurons are plated with them to make the NMJ. Frega et al. designed a unique strategy to form 3D neuronal cultures from hippocampal neurons grown on layers of laminin and poly-l-lysine-coated 40 μm glass beads. Placing these 3D layers over a neuronal monolayer on top of microelectrode arrays created network connectivity and enabled measurement of both spontaneous and evoked field potentials. Collagen gel cultures have been used to study the effect of inflammatory cytokines such as interleukin 1β and tumor necrosis factor on neurite outgrowth.55 Kaplan’s group has developed a “functional brain-like cortical tissue” using 3D neuronal cultures in silk fibroin-based materials in a compartmentalized manner.56 The silk fibroin provided both the required electrical connectivity and mechanical stability to mimic the spongy brain tissue.
Work on developing 3D models using motor neuron and muscle cells has been increasing in recent years. As demonstrated by studies with 2D cultures, dimensional restraints disallow proper contractile functioning of muscle monolayers in response to stimulation from innervating motor neurons.57 The difficulty in culturing motor neurons and muscle in compatible 3D constructs/gels with the appropriate mechanical, electrical, and chemical cues for cell differentiation, proliferation, contraction, and neurite outgrowth has made it harder to construct 3D NMJs.
In a seminal report, Morimoto et al., have presented some of the first results in 3D in vitro NMJ models.58 Using PDMS stamps, they patterned C2C12 mouse muscle myoblast-loaded Matrigel onto PDMS substrates. The patterned muscle cells were cultured on the PDMS substrates to form free-standing muscle fibers anchored to fibronectin-coated glass plates fixed at both ends of the substrate. Mouse neural stem cells were then plated on these free-standing fibers after reducing the volume of culture medium to enable attachment of the “neurospheres” to the fibers. After ensuring attachment, the neural stem cells were then differentiated into neurons, and they formed functional NMJs that can be stimulated to trigger muscle contractions after 14 days in culture. Although 3D cultures provide a substrate or scaffold that mimic physiological environments, the organization and alignment of muscle fibers and motor neurons are equally important for the guidance of axons and communication between pre- and post-synaptic structures. Micropatterned NMJs in a dish on mechanically patterned ECM strip have been formed, demonstrating increased differentiation and fusion of the myotubes compared to 3D cultures and larger ACh receptors.59 An increased expression of NMJ receptors such as MuSK and Lrp4 and an increased contraction of the myotube fibers when stimulated by ACh confirm the presence of a fully functional NMJ.
More research is required in this area to generate viable, long-term 3D in vitro models for the NMJ. As detailed earlier, both skeletal muscle and neuronal cells do well in collagen, matrigel, and silk fibroin gels. Collagen type I gels would provide the desired porosity for axonal growth60 and the necessary elasticity for contractility. Fibrin–collagen61 or hyaluronic acid–collagen62 constructs could be used for added mechanical stability.
Microfluidics and 3D In Vitro NMJ Models
The ability to combine both 3D cultures and microfluidic devices to study the NMJ is an attractive prospect. Microfluidics would enable spatial and temporal control over the microenvironment while allowing for a precise, specific, and localized interrogation of the system. The 3D environment would provide for better cell survival, growth, differentiation, and functionality making long-term studies using in vitro cultures possible. Putting together these models would provide the best of both worlds while overcoming some of the disadvantages inherent in each technique. Motor neurons and muscle cells can be compartmentalized while providing spatial guidance cues and signaling gradients using perfusion or 3D hydrogels63 for better integration and formation of synapses.64 Shimizu et al., have shown a promising start in this area by engineering functional 3D contractile C2C12 skeletal muscle myofibers in microchannels,65 while Yang et al. have confirmed the role of in vivo paracrine signals in the differentiation of human neuronal stem cells cultured in a 3D ECM hydrogel-filled microfluidic device.66 Yang’s work is noteworthy because it involves the interaction between two cell types in two different chambers and the cell–cell interaction here induces stem cell differentiation via paracrine signaling.
Other work in this area has included the differentiation of neuroepithelial stem cells into functional dopaminergic neurons in a 3D microfluidic device67 and the development of a photo-cross linked polymer gelatin methacrylate, which could be incorporated into a microfluidic device to culture neural stem cells.68 Work has been done on an in vitro platform by Uzel et al.,69 where 3D hydrogel cultures of mouse embryonic stem cell-derived motor neurons and skeletal muscle were compartmentalized using PDMS chambers separated by flexible pillars that serve to direct neurite outgrowth as well as deflect upon muscle contraction. Figure 5 schematically depicts this idea behind combining microfluidics and 3D cultures to study the NMJ.
Figure 5.
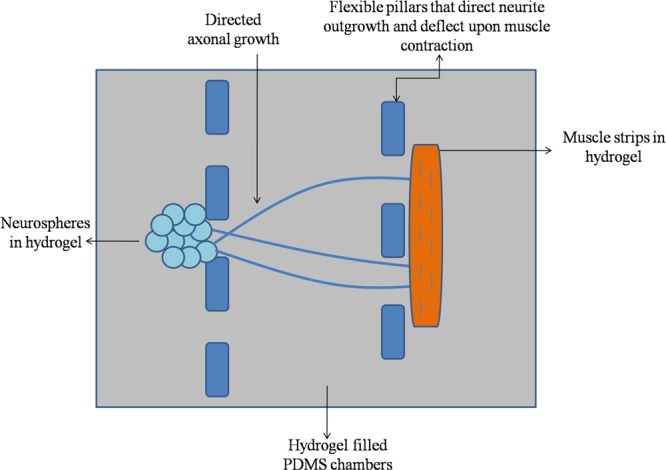
Combining 3D hydrogel cultures with microfluidic PDMS chambers and anchoring pillars to separate both cell types as well as isolate and study the NMJ.
The arrival of iPSCs has opened new avenues of investigation in different fields including studies on NMJ.16 Recent reports have indicated that iPSCs from patients with neurodegenerative diseases such as ALS70 and SMA71 can be differentiated into motor neurons. Incorporating such cells into standardized 3D microfluidic devices would help identify therapeutic targets and have applications in drug discovery and personalized medicine.
3D Bioprinting
The advent of 3D bioprinting has made it easier to explore tissue engineering in vitro using disassociated cell cultures, cocultures, and 3D hydrogels/ECM proteins, allowing for the use of flexible designs that more accurately capture in vivo conditions. Recent studies have looked at using 3D bioprinting to make in vitro models of the NMJ. C2C12 myoblasts differentiated into multinucleated myotubes and mouse embryonic stem cells differentiated into motor neurons were first cultured separately in ECM proteins to create a muscle ring tissue and embryoid bodies (EB), respectively.72 The engineered muscle tissue, ECM proteins, and EB were all then combined with different hydrogels, which allows for the formation of multi-ringed tissue that are integrated with each other. Cell–gel combinations included C2C12 cells in thrombin, fibrinogen, and matrigel solutions cultured in differentiation factors and tissue molds to form a ring-like structure. Additional cells in gels, including EB containing motor neurons, were then added to the hydrogel ring molds to form the tissue rings. Analysis performed using immunohistochemistry, electron microscopy, and neurite growth measurements showed a greater quantity of differentiated motor neurons—pointing to the effect of the 3D hydrogel environment. The presence of supporting glial cells among the EB is also thought to contribute to the better survival, growth, differentiation, and proliferation of motor neurons as well as the formation and maintenance of the NMJ. The motor neurons, C2C12 muscle myotubes and clusters of postsynaptic functional AChR units were identified by fluorescence imaging. Spontaneous twitching was also observed within individual layers and between the layers of these motor neuron constructs. These tissue construct rings were then placed on bioprinted 3D hydrogel “skeletons” that provide the structural integrity, stability, and connectivity of the muscle–tendon–ligament–bone structures found in the body. EBs showed extended neurite outgrowth into the “tissue” and chemical stimulation leads to contraction of the aligned muscle fiber tissue.72
More recently, a study along similar lines examined developing 3D bioprinted silk fibroin cantilevers as support systems for culturing and aligning human myotubes, and the addition of motor neuron-like cells from human induced stem cells into this system allows for the integration of these cells with muscle and provides a novel in vitro platform comprising NMJ units.73 Such novel 3D bioprinted functional NMJ tissue structures can be used to quantify muscle activity, the effect of chemical and electrical stimulants, and the changes in growth and development of the NMJ units themselves over time and in response to external stimuli.
Current Outlook and Future Perspectives
A combination of novel compartmentalized microfluidic chambers and varied cell culture models are currently being used to understand neuromuscular functioning and signaling pathways, disease pathologies, and to test new therapeutic drugs. Although it is understood that glia play a central role in supporting neuronal cells, the complexity of culturing three different cell types in vitro has limited the introduction of these supporting cells into the NMJ models that comprise motor neurons and skeletal muscle cells. Schwann cells, in particular, play an important role in the development and functioning of the motor neuron. A time-dependent addition of motor neurons (day 5) and Schwann cells (day 9) to skeletal muscle cells in a compartmentalized microfluidic platform showed a significant increase in the viability, number, and length of individual myotubes.74
The development of stable, functional NMJ models has recently led to different groups trying to understand and study disease pathology at the molecular level. Increasingly optogenetics is being explored to identify NMJs and create motor neurons activated by light as opposed to electrical stimulation. Using human iPSC (hiPSC) also allows for human and patient donor-specific NMJ models. The combination of compartmentalized microfluidic chambers, the optogenetic channel-rhodopsin-2 protein-expressing photosensitive motor neurons, and the non-optogenetic skeletal muscle from a single donor was used to study NMJ functionality and connectivity over time as well as the effect of neurotoxin-induced damage. This novel model provided the added advantage of being able to study disease pathology wherein the NMJ acted as a biosensor, being able to detect and change in response to myasthenia gravis autoantibodies in patient serum.75 In this study, motor neurons were stimulated optically and force/contraction measurements were performed using specially developed software. Motor units, motor neurons, skeletal muscle cell viability and functionality were characterized using immunohistochemistry and flow cytometry. The microfluidic platform used for compartmentalization is an updated version of earlier models consisting of compartments separated by channels for axonal growth and pillars for muscle bundle attachment.
ALS is another fatal neurodegenerative disease that involves motor neuron degeneration and muscle atrophy. NMJ dysfunction and damage precedes motor neuron cell death in ALS and groups are now attempting to build patient-specific microfluidic-based chips of the NMJ. Using optogenetic hiPSC-induced motor neurons from patients with sporadic ALS, Kamm et al. were able to assess and quantify neurite outgrowth, muscle contraction, and cell viability and compare it to non-ALS motor units. Pillars used to tether and separate skeletal muscle were used to quantify muscle contraction when motor neurons were activated by light. The efficacy of drugs and inhibitors used to treat ALS was also studied in this model.76
There are now many well-characterized in vitro LOC models of the NMJ that could in the future be a part of the body of “body-on-a-chip” systems. Current models include 3D cultures incorporating compartmentalized motor neurons and skeletal muscle forming functional NMJs with supporting cells such as Schwann cells aiding in the formation of robust motor units. The advent of hiPSCs making these models patient-specific allows us to finally explore the neurodegenerative diseases of the peripheral nervous system at the molecular level and to aid in drug development and study drug side effects. Such “NMJ chips” could be developed into theranostic devices that can be used to monitor and assess NMJ degeneration over time and develop effective treatment strategies.
Although in vitro models and cell cultures cannot precisely mimic in vivo conditions for applications such as drug discovery, the advent of microfluidic designs and lab/organ/body-on-a-chip devices allows for creating environments that come close to physiological conditions. Such in vitro microfluidic models of the NMJ or other tissue would enable the cells to get nutrients and oxygen via fluid circulation as well as be subjected to varied mechanical and chemical stimulation needed for cell differentiation, growth, viability, and proliferation. Such a system would also allow for drug perfusion and delivery to target tissue sites and ways to monitor and assess tissue response. An added advantage of such models is that the use of hiPSCs offers an opportunity to test and study drug effects on human tissue as opposed to animal models. A “bioreactor” model of the NMJ, using a coculture of electrically stimulated skeletal myocytes and motor neurons differentiated from transgenic mouse cell line-derived embryonic stem cells was used to study the efficacy of drugs that affect key signaling pathways during NMJ activation to develop a platform for pharmaceutical testing.77 Dose–response curves from these experiments reveal a concentration-dependent response in this model to drugs that block the synthesis of ACh and act as inhibitors of vesicular ACh transporters and neurotransmission. The model was sensitive to low concentrations of the applied drugs, and the inhibitory concentrations obtained were comparable to other in vitro models. There are however no comparisons done with the half maximal inhibitory concentrations and lethal doses in in vivo models. The inability to replicate precise in vivo physiological conditions and the differences that exist between in vivo and in vitro environments, cell morphology, supporting cells, blood vessels, and other factors are potential pitfalls and limitations to using these models for pharmacokinetics and drug discovery.
Ultimately the “NMJ—on—a—chip” could introduce a new paradigm in understanding the NMJ and in drug testing, leading to a reduction in animal testing. More high quality data is however necessary with in vitro results being comparable to in vivo results. In vitro NMJ models, that closely mimic in vivo conditions, could ultimately replace animal studies and provide an inexpensive, high-throughput tool for treating neurodegenerative diseases, engineer prosthetic devices, and understand aging at the NMJ. The integration of 3D culture with microfluidic compartments for coculture of different cell types may lead to an ideal NMJ mimic in the near future. The increasing awareness and rising global incidence of neurodegenerative disorders have provided the correct quantum of impetus to research on developing realistic NMJ models in vitro, and it is but a matter of time when these systems will be routinely employed for drug discovery and basic science studies.
Acknowledgments
The authors wish to acknowledge SASTRA Deemed University for infrastructural support and funding through Prof. TRR R&D fund. Financial support through DST-FIST (SR/FST/LSI-453/2010) and Early Career Research award (ECR/2016/000006) from the Department of Science & Technology is also gratefully acknowledged.
The authors declare no competing financial interest.
References
- Patton B. L. Basal lamina and the organization of neuromuscular synapses. J. Neurocytol. 2003, 32, 883–903. 10.1023/b:neur.0000020630.74955.19. [DOI] [PubMed] [Google Scholar]
- Punga A. R.; Ruegg M. A. Signaling and aging at the neuromuscular synapse: lessons learnt from neuromuscular diseases. Curr. Opin. Pharmacol. 2012, 12, 340–346. 10.1016/j.coph.2012.02.002. [DOI] [PubMed] [Google Scholar]
- Ziskind-Conhaim L.; Bennett J. I. The effects of electrical inactivity and denervation on the distribution of acetylcholine receptors in developing rat muscle. Dev. Biol. 1982, 90, 185–197. 10.1016/0012-1606(82)90224-x. [DOI] [PubMed] [Google Scholar]
- Anderson M. J.; Cohen M. W. Nerve-induced and spontaneous redistribution of acetylcholine receptors on cultured muscle cells. J. Physiol. 1977, 268, 757–773. 10.1113/jphysiol.1977.sp011880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu H.; Xiong W. C.; Mei L. To build a synapse: signaling pathways in neuromuscular junction assembly. Development 2010, 137, 1017–1033. 10.1242/dev.038711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darabid H.; Perez-Gonzalez A. P.; Robitaille R. Neuromuscular synaptogenesis: coordinating partners with multiple functions. Nat. Rev. Neurosci. 2014, 15, 703–718. 10.1038/nrn3821. [DOI] [PubMed] [Google Scholar]
- Kanning K. C.; Kaplan A.; Henderson C. E. Motor Neuron Diversity in Development and Disease. Annu. Rev. Neurosci. 2010, 33, 409–440. 10.1146/annurev.neuro.051508.135722. [DOI] [PubMed] [Google Scholar]
- Johnson L. R.; Byrne J. H.. Essential Medical Physiology; Academic Press, 2003. [Google Scholar]
- Gonzalez-Freire M.; de Cabo R.; Studenski S. A.; Ferrucci L. The Neuromuscular Junction: Aging at the Crossroad between Nerves and Muscle. Front. Aging Neurosci. 2014, 6, 208. 10.3389/fnagi.2014.00208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo X.; Ayala J. E.; Gonzalez M.; Stancescu M.; Lambert S.; Hickman J. J. Tissue engineering the monosynaptic circuit of the stretch reflex arc with co-culture of embryonic motoneurons and proprioceptive sensory neurons. Biomaterials 2012, 33, 5723–5731. 10.1016/j.biomaterials.2012.04.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benatar M. Lost in translation: treatment trials in the SOD1 mouse and in human ALS. Neurobiol. Dis. 2007, 26, 1–13. 10.1016/j.nbd.2006.12.015. [DOI] [PubMed] [Google Scholar]
- Rizzuto E.; Pisu S.; Musaro A.; Del Prete Z. Measuring Neuromuscular Junction Functionality in the SOD1 Animal Model of Amyotrophic Lateral Sclerosis. Ann. Biomed. Eng. 2015, 43, 2196. 10.1007/s10439-015-1259-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- te Pas F. W.; Everts E.; Haagsman H. P.. Muscle Development of Livestock Animals: Physiology, Genetics and Meat Quality; CABI Pub., 2004. [Google Scholar]
- Hackam D. G. Translating animal research into clinical benefit. Bmj 2007, 334, 163–164. 10.1136/bmj.39104.362951.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo X.; Das M.; Rumsey J.; Gonzalez M.; Stancescu M.; Hickman J. Neuromuscular junction formation between human stem-cell-derived motoneurons and rat skeletal muscle in a defined system. Tissue Eng., Part C 2010, 16, 1347–1355. 10.1089/ten.tec.2010.0040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomson S. R.; Wishart T. M.; Patani R.; Chandran S.; Gillingwater T. H. Using induced pluripotent stem cells (iPSC) to model human neuromuscular connectivity: promise or reality?. J. Anat. 2012, 220, 122–130. 10.1111/j.1469-7580.2011.01459.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engler A. J.; Griffin M. A.; Sen S.; Bönnemann C. G.; Sweeney H. L.; Discher D. E. Myotubes differentiate optimally on substrates with tissue-like stiffness. J. Cell Biol. 2004, 166, 877–887. 10.1083/jcb.200405004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filogamo G.; Gabella G. The development of neuro-muscular correlations, in vertebrates. Arch. Biol. 1967, 78, 9–60. [PubMed] [Google Scholar]
- Liveson J.; Frey H.; Bornstein M. B. The effect of serum from ALS patients on organotypic nerve and muscle tissue cultures. Acta Neuropathol. 1975, 32, 127–131. 10.1007/bf00689566. [DOI] [PubMed] [Google Scholar]
- Peterson E. R.; Masurovsky E. B.; Spiro A. J.; Crain S. M. Duchenne dystrophic muscle develops lesions in long-term coculture with mouse spinal cord. Muscle Nerve 1986, 9, 787–808. 10.1002/mus.880090903. [DOI] [PubMed] [Google Scholar]
- MacDonald S. C.; Fleetwood I. G.; Hochman S.; Dodd J. G.; Cheng G. K. W.; Jordan L. M.; Brownstone R. M. Functional motor neurons differentiating from mouse multipotent spinal cord precursor cells in culture and after transplantation into transected sciatic nerve. J. Neurosurg. 2003, 98, 1094–1103. 10.3171/jns.2003.98.5.1094. [DOI] [PubMed] [Google Scholar]
- Kubo T.; Randolph M. A.; Groger A.; Winograd J. M. Embryonic stem cell-derived motor neurons form neuromuscular junctions in vitro and enhance motor functional recovery in vivo. Plast. Reconstr. Surg. 2009, 123, 139s–148s. 10.1097/prs.0b013e3181923d07. [DOI] [PubMed] [Google Scholar]
- Guo X.; Gonzalez M.; Stancescu M.; Vandenburgh H. H.; Hickman J. J. Neuromuscular junction formation between human stem cell-derived motoneurons and human skeletal muscle in a defined system. Biomaterials 2011, 32, 9602–9611. 10.1016/j.biomaterials.2011.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park H.-W.; Cho J.-S.; Park C.-K.; Jung S. J.; Park C.-H.; Lee S.-J.; Oh S. B.; Park Y.-S.; Chang M.-S. Directed induction of functional motor neuron-like cells from genetically engineered human mesenchymal stem cells. PLoS One 2012, 7, e35244 10.1371/journal.pone.0035244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rumsey J. W.; Das M.; Bhalkikar A.; Stancescu M.; Hickman J. J. Tissue engineering the mechanosensory circuit of the stretch reflex arc: sensory neuron innervation of intrafusal muscle fibers. Biomaterials 2010, 31, 8218–8227. 10.1016/j.biomaterials.2010.07.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ionescu A.; Zahavi E. E.; Gradus T.; Ben-Yaakov K.; Perlson E. Compartmental microfluidic system for studying muscle-neuron communication and neuromuscular junction maintenance. Eur. J. Cell Biol. 2016, 95, 69–88. 10.1016/j.ejcb.2015.11.004. [DOI] [PubMed] [Google Scholar]
- Dworak B. J.; Wheeler B. C. Novel MEA platform with PDMS microtunnels enables the detection of action potential propagation from isolated axons in culture. Lab Chip 2009, 9, 404–410. 10.1039/b806689b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campenot R. B. Local control of neurite development by nerve growth factor. Proc. Natl. Acad. Sci. U.S.A. 1977, 74, 4516–4519. 10.1073/pnas.74.10.4516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan L.; Alagapan S.; Franca E.; Brewer G. J.; Wheeler B. C. Propagation of action potential activity in a predefined microtunnel neural network. J. Neural Eng. 2011, 8, 046031. 10.1088/1741-2560/8/4/046031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald J. C.; Duffy D. C.; Anderson J. R.; Chiu D. T.; Wu H.; Schueller O. J. A.; Whitesides G. M. Fabrication of microfluidic systems in poly(dimethylsiloxane). Electrophoresis 2000, 21, 27–40. . [DOI] [PubMed] [Google Scholar]
- a Taylor A. M.; Blurton-Jones M.; Rhee S. W.; Cribbs D. H.; Cotman C. W.; Jeon N. L. A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat. Methods 2005, 2, 599–605. 10.1038/nmeth777. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Taylor A.; Jeon N. L. Microfluidic and compartmentalized platforms for neurobiological research. Crit. Rev. Bioeng. 2011, 39, 185–200. 10.1615/critrevbiomedeng.v39.i3.20. [DOI] [PubMed] [Google Scholar]
- Taylor A. M.; Dieterich D. C.; Ito H. T.; Kim S. A.; Schuman E. M. Microfluidic local perfusion chambers for the visualization and manipulation of synapses. Neuron 2010, 66, 57–68. 10.1016/j.neuron.2010.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poon W. W.; Blurton-Jones M.; Tu C. H.; Feinberg L. M.; Chabrier M. A.; Harris J. W.; Jeon N. L.; Cotman C. W. β-Amyloid impairs axonal BDNF retrograde trafficking. Neurobiol. Aging 2011, 32, 821–833. 10.1016/j.neurobiolaging.2009.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wisner T. W.; Sugimoto K.; Howard P. W.; Kawaguchi Y.; Johnson D. C. Anterograde transport of herpes simplex virus capsids in neurons by both separate and married mechanisms. J. Virol. 2011, 85, 5919–5928. 10.1128/jvi.00116-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H. J.; Park J. W.; Byun J. H.; Vahidi B.; Rhee S. W.; Jeon N. L. Integrated microfluidics platforms for investigating injury and regeneration of CNS axons. Ann. Biomed. Eng. 2012, 40, 1268–1276. 10.1007/s10439-012-0515-6. [DOI] [PubMed] [Google Scholar]
- Tourovskaia A.; Li N.; Folch A. Localized acetylcholine receptor clustering dynamics in response to microfluidic focal stimulation with agrin. Biophys. J. 2008, 95, 3009–3016. 10.1529/biophysj.107.128173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tandon N.; Marsano A.; Maidhof R.; Numata K.; Montouri-Sorrentino C.; Cannizzaro C.; Voldman J.; Vunjak-Novakovic G. Surface-Patterned Electrode Bioreactor for Electrical Stimulation. Lab Chip 2010, 10, 692–700. 10.1039/b917743d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wan C.-r.; Chung S.; Kamm R. D. Differentiation of embryonic stem cells into cardiomyocytes in a compliant microfluidic system. Ann. Biomed. Eng. 2011, 39, 1840–1847. 10.1007/s10439-011-0275-8. [DOI] [PubMed] [Google Scholar]
- Zhao Y.; Zeng H.; Nam J.; Agarwal S. Fabrication of skeletal muscle constructs by topographic activation of cell alignment. Biotechnol. Bioeng. 2009, 102, 624–631. 10.1002/bit.22080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy J.; Kennedy T. E.; Costantino S. Engineered cell culture substrates for axon guidance studies: moving beyond proof of concept. Lab Chip 2013, 13, 498–508. 10.1039/c2lc41002h. [DOI] [PubMed] [Google Scholar]
- Takeuchi A.; Nakafutami S.; Tani H.; Mori M.; Takayama Y.; Moriguchi H.; Kotani K.; Miwa K.; Lee J.-k.; Noshiro M.; Jimbo Y. Device for co-culture of sympathetic neurons and cardiomyocytes using microfabrication. Lab Chip 2011, 11, 2268–2275. 10.1039/c0lc00327a. [DOI] [PubMed] [Google Scholar]
- Park H. S.; Liu S.; McDonald J.; Thakor N.; Yang I. H.. Neuromuscular junction in a microfluidic device. Conference of the IEEE Engineering in Medicine and Biology Society, 2013; Vol. 2013, pp 2833–2835. [DOI] [PubMed]
- Southam K. A.; King A. E.; Blizzard C. A.; McCormack G. H.; Dickson T. C. Microfluidic primary culture model of the lower motor neuron-neuromuscular junction circuit. J. Neurosci. Methods 2013, 218, 164–169. 10.1016/j.jneumeth.2013.06.002. [DOI] [PubMed] [Google Scholar]
- Zahavi E. E.; Ionescu A.; Gluska S.; Gradus T.; Ben-Yaakov K.; Perlson E. A compartmentalized microfluidic neuromuscular co-culture system reveals spatial aspects of GDNF functions. J. Cell Sci. 2015, 128, 1241–1252. 10.1242/jcs.167544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klumpp D.; Horch R. E.; Kneser U.; Beier J. P. Engineering skeletal muscle tissue - new perspectives in vitro and in vivo. J. Cell. Mol. Med. 2010, 14, 2622–2629. 10.1111/j.1582-4934.2010.01183.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharples A. P.; Player D. J.; Martin N. R. W.; Mudera V.; Stewart C. E.; Lewis M. P. Modellingin vivoskeletal muscle ageingin vitrousing three-dimensional bioengineered constructs. Aging Cell 2012, 11, 986–995. 10.1111/j.1474-9726.2012.00869.x. [DOI] [PubMed] [Google Scholar]
- Leikina E.; Mertts M. V.; Kuznetsova N.; Leikin S. Type I collagen is thermally unstable at body temperature. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 1314–1318. 10.1073/pnas.032307099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grefte S.; Vullinghs S.; Kuijpers-Jagtman A. M.; Torensma R.; Von den Hoff J. W. Matrigel, but not collagen I, maintains the differentiation capacity of muscle derived cellsin vitro. Biomed. Mater. 2012, 7, 055004. 10.1088/1748-6041/7/5/055004. [DOI] [PubMed] [Google Scholar]
- Zhao C.; Yao J.; Masuda H.; Kishore R.; Asakura T. Structural characterization and artificial fiber formation ofBombyx mori silk fibroin in hexafluoro-iso-propanol solvent system. Biopolymers 2003, 69, 253–259. 10.1002/bip.10350. [DOI] [PubMed] [Google Scholar]
- Huang Y. C.; Dennis R. G.; Larkin L.; Baar K. Rapid formation of functional muscle in vitro using fibrin gels. J Appl Physiol 2005, 98, 706–713. 10.1152/japplphysiol.00273.2004. [DOI] [PubMed] [Google Scholar]
- Afshar Bakooshli M.; Lippmann E.; Mulcahy B.; Iyer N.; Nguyen C.; Tung K.; Stewart B.; van den Dorpel H.; Fuehrmann T.; Shoichet M.; Bigot A.; Pegoraro E.; Ahn H.; Ginsberg H.; Zhen M.; Ashton R.; Gilbert P.. A three-dimensional culture model of innervated human skeletal muscle enables studies of the adult neuromuscular junction and disease modeling. 2018, bioRxiv:275545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marani L.; Pardo-Figuerez M.; Capel A. J.; Nilsson Z.; Stolzing A.; Lewis M. P. Development of a human in vitro model of the neuromuscular junction, using induced pluripotent stem cell-derived motor neurons and 3d tissue engineering constructs. Orthop. Proc. 2018, 100-B, 79. 10.1302/1358-992X.2018.16.079. [DOI] [Google Scholar]
- Smith A. S. T.; Passey S. L.; Martin N. R. W.; Player D. J.; Mudera V.; Greensmith L.; Lewis M. P. Creating Interactions between Tissue-Engineered Skeletal Muscle and the Peripheral Nervous System. Cells Tissues Organs 2016, 202, 143–158. 10.1159/000443634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin N. R. W.; Passey S. L.; Player D. J.; Mudera V.; Baar K.; Greensmith L.; Lewis M. P. Neuromuscular Junction Formation in Tissue-Engineered Skeletal Muscle Augments Contractile Function and Improves Cytoskeletal Organization. Tissue Eng., Part A 2015, 21, 2595–2604. 10.1089/ten.tea.2015.0146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H.; Caspar T. W.; Shah S. B.; Hsieh A. H. Effects of proinflammatory cytokines on axonal outgrowth from adult rat lumbar dorsal root ganglia using a novel three-dimensional culture system. Spine J. 2015, 15, 1823–1831. 10.1016/j.spinee.2015.03.017. [DOI] [PubMed] [Google Scholar]
- Tang-Schomer M. D.; White J. D.; Tien L. W.; Schmitt L. I.; Valentin T. M.; Graziano D. J.; Hopkins A. M.; Omenetto F. G.; Haydon P. G.; Kaplan D. L. Bioengineered functional brain-like cortical tissue. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 13811–13816. 10.1073/pnas.1324214111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosseini V.; Ahadian S.; Ostrovidov S.; Camci-Unal G.; Chen S.; Kaji H.; Ramalingam M.; Khademhosseini A. Engineered contractile skeletal muscle tissue on a microgrooved methacrylated gelatin substrate. Tissue Eng., Part A 2012, 18, 2453–2465. 10.1089/ten.tea.2012.0181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morimoto Y.; Kato-Negishi M.; Onoe H.; Takeuchi S. Three-dimensional neuron-muscle constructs with neuromuscular junctions. Biomaterials 2013, 34, 9413–9419. 10.1016/j.biomaterials.2013.08.062. [DOI] [PubMed] [Google Scholar]
- Happe C. L.; Tenerelli K. P.; Gromova A. K.; Kolb F.; Engler A. J. Mechanically patterned neuromuscular junctions-in-a-dish have improved functional maturation. Mol. Biol. Cell 2017, 28, 1950–1958. 10.1091/mbc.e17-01-0046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- a Madaghiele M.; Sannino A.; Yannas I. V.; Spector M. Collagen-based matrices with axially oriented pores. J. Biomed. Mater. Res., Part A 2008, 85A, 757–767. 10.1002/jbm.a.31517. [DOI] [PubMed] [Google Scholar]; b Fukushima K.; Enomoto M.; Tomizawa S.; Takahashi M.; Wakabayashi Y.; Itoh S.; Kuboki Y.; Shinomiya K. The axonal regeneration across a honeycomb collagen sponge applied to the transected spinal cord. J. Evol. Med. Dent. Sci. 2008, 55, 71–79. [PubMed] [Google Scholar]
- Cvetkovic C.; Raman R.; Chan V.; Williams B. J.; Tolish M.; Bajaj P.; Sakar M. S.; Asada H. H.; Saif M. T. A.; Bashir R. Three-dimensionally printed biological machines powered by skeletal muscle. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 10125–10130. 10.1073/pnas.1401577111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang T.-W.; Spector M. Development of hyaluronic acid-based scaffolds for brain tissue engineering. Acta Biomater. 2009, 5, 2371–2384. 10.1016/j.actbio.2009.03.033. [DOI] [PubMed] [Google Scholar]
- Sundararaghavan H. G.; Masand S. N.; Shreiber D. I. Microfluidic generation of haptotactic gradients through 3D collagen gels for enhanced neurite growth. J. Neurotrauma 2011, 28, 2377–2387. 10.1089/neu.2010.1606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Duinen V.; Trietsch S. J.; Joore J.; Vulto P.; Hankemeier T. Microfluidic 3D cell culture: from tools to tissue models. Curr. Opin. Biotechnol. 2015, 35, 118–126. 10.1016/j.copbio.2015.05.002. [DOI] [PubMed] [Google Scholar]
- Shimizu K.; Araki H.; Sakata K.; Tonomura W.; Hashida M.; Konishi S. Microfluidic devices for construction of contractile skeletal muscle microtissues. J. Biosci. Bioeng. 2015, 119, 212–216. 10.1016/j.jbiosc.2014.07.003. [DOI] [PubMed] [Google Scholar]
- Yang K.; Park H.-J.; Han S.; Lee J.; Ko E.; Kim J.; Lee J. S.; Yu J. H.; Song K. Y.; Cheong E.; Cho S.-R.; Chung S.; Cho S.-W. Recapitulation of in vivo-like paracrine signals of human mesenchymal stem cells for functional neuronal differentiation of human neural stem cells in a 3D microfluidic system. Biomaterials 2015, 63, 177–188. 10.1016/j.biomaterials.2015.06.011. [DOI] [PubMed] [Google Scholar]
- Moreno E. L.; Hachi S.; Hemmer K.; Trietsch S. J.; Baumuratov A. S.; Hankemeier T.; Vulto P.; Schwamborn J. C.; Fleming R. M. T. Differentiation of neuroepithelial stem cells into functional dopaminergic neurons in 3D microfluidic cell culture. Lab Chip 2015, 15, 2419–2428. 10.1039/c5lc00180c. [DOI] [PubMed] [Google Scholar]
- Lee Y.; Lee J. M.; Bae P.-K.; Chung I. Y.; Chung B. H.; Chung B. G. Photo-crosslinkable hydrogel-based 3D microfluidic culture device. Electrophoresis 2015, 36, 994–1001. 10.1002/elps.201400465. [DOI] [PubMed] [Google Scholar]
- Uzel S. G. M.; Platt R. J.; Subramanian V.; Pearl T. M.; Rowlands C. J.; Chan V.; Boyer L. A.; So P. T. C.; Kamm R. D. Microfluidic device for the formation of optically excitable, three-dimensional, compartmentalized motor units. Sci. Adv. 2016, 2, e1501429 10.1126/sciadv.1501429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dimos J. T.; Rodolfa K. T.; Niakan K. K.; Weisenthal L. M.; Mitsumoto H.; Chung W.; Croft G. F.; Saphier G.; Leibel R.; Goland R.; Wichterle H.; Henderson C. E.; Eggan K. Induced pluripotent stem cells generated from patients with ALS can be differentiated into motor neurons. Science 2008, 321, 1218–1221. 10.1126/science.1158799. [DOI] [PubMed] [Google Scholar]
- Ebert A. D.; Yu J.; Rose F. F.; Mattis V. B.; Lorson C. L.; Thomson J. A.; Svendsen C. N. Induced pluripotent stem cells from a spinal muscular atrophy patient. Nature 2009, 457, 277–280. 10.1038/nature07677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cvetkovic C.; Rich M. H.; Raman R.; Kong H.; Bashir R. A 3D-printed platform for modular neuromuscular motor units. Microsyst. Nanoeng. 2017, 3, 17015. 10.1038/micronano.2017.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anthony D. T.; Eliad C.; Cairns D. M.; Maria R.; Juanita M.; Mathews J.; Kaplan D. L. Bioinspired Three-Dimensional Human Neuromuscular Junction Development in Suspended Hydrogel Arrays. Tissue Eng., Part C 2018, 24, 346–359. 10.1089/ten.tec.2018.0062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh T.; Vazquez M. Time-Dependent Addition of Neuronal and Schwann Cells Increase Myotube Viability and Length in an In Vitro Tri-culture Model of the Neuromuscular Junction. Regen. Eng. Transl. Med. 2019, 1–12. 10.1007/s40883-019-00095-5.30976657 [DOI] [Google Scholar]
- Vila O. F.; Uzel S. G. M.; Ma S. P.; Williams D.; Pak J.; Kamm R. D.; Vunjak-Novakovic G. Quantification of human neuromuscular function through optogenetics. Theranostics 2019, 9, 1232–1246. 10.7150/thno.25735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osaki T.; Uzel S. G. M.; Kamm R. D. Microphysiological 3D model of amyotrophic lateral sclerosis (ALS) from human iPS-derived muscle cells and optogenetic motor neurons. Sci. Adv. 2018, 4, eaat5847 10.1126/sciadv.aat5847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charoensook S. N.; Williams D. J.; Chakraborty S.; Leong K. W.; Vunjak-Novakovic G. Bioreactor model of neuromuscular junction with electrical stimulation for pharmacological potency testing. Integr. Biol. 2017, 9, 956–967. 10.1039/c7ib00144d. [DOI] [PMC free article] [PubMed] [Google Scholar]