

Abstract
The melanocortin pathway has been implicated in both metabolism and sexual function. When the melanocortin 4 receptor (MC4R) is knocked out globally, male mice display obesity, low sexual desire, and copulatory difficulties; however, it is unclear whether these phenotypes are interdependent. To elucidate the neuronal circuitry involved in sexual dysfunction in MC4R knockouts, we re-expressed the MC4R in these mice exclusively on Sim1 neurons (tbMC4RSim1 mice) or on a subset of Sim1 neurons, namely oxytocin neurons (tbMC4Roxt mice). The groups were matched at young ages to control for the effects of obesity. Interestingly, young MC4R null mice had no deficits in sexual motivation or erectile function. However, MC4R null mice were found to have an increased latency to reach ejaculation compared to control mice, which was restored in both tbMC4RSim1 and tbMC4Roxt mice. These results indicate that melanocortin signaling via the MC4R on oxytocin neurons is important for normal ejaculation independent of the male’s metabolic health.
Keywords: paraventricular nucleus, erectile dysfunction, melanocortin, obesity
Introduction:
Sexual dysfunction can profoundly affect a man’s quality of life. At least one fifth of men experience some form of sexual dysfunction during their lifetime [1,2]. The correlation between age and worsening sexual function is well-documented [3–7], often related to increased obesity and consequent diabetes and cardiovascular complications [8]. Although treatments such as phosphodiesterase inhibitors exist for erectile dysfunction, responsiveness to these drugs vary, and medications for other forms of sexual dysfunction are limited [3]. Medications targeting potential impairments in brain circuits underlying sexual function have recently garnered interest. For example, selective serotonin reuptake inhibitors, which act by increasing extracellular serotonin, have been used as a treatment for premature ejaculation [2]. The dopamine agonist apomorphine has been explored as a treatment for sexual motivation and erectile dysfunction [9]. Drugs that target the melanocortin system have also been tested as an erectile dysfunction treatment [10,11]. Increasing our understanding of the neurocircuitry underlying aspects of sexual function may lead to more effective treatment options.
Strong preclinical and clinical evidence supports the involvement of the melanocortin system in sexual function [12–15]. Melanocortins, including α-melanocyte stimulating hormone (MSH), βMSH, γMSH, and adrenocorticotropic hormone (ACTH), are products of the proopiomelanocortin (POMC) polypeptide. These products bind to five different melanocortin receptors (MC1R, MC2R, MC3R, MC4R, MC5R) with varying binding affinities [16–18]. Since the MC3R and MC4R are the primary subtypes expressed by the brain [19], these two may mediate the central effects of melanocortins. MC4 receptors, which are G-protein coupled receptors that can couple with Gi/o, Gs, and Gq, can trigger multiple downstream pathways [20]. Evidence suggests that the MC4R, which has an established role in in mediating satiety [21,22], may also mediate sexual behavior and function. Administration of exogenous αMSH, an agonist of the MC3R and MC4R [18], has been found to lead to erection in rodents [23]. At three to six months of age, MC4R null mice show decreased motivation to engage in sexual activity and take longer to reach ejaculation [14]. Male mice with POMC-producing neurons that are insensitive to leptin and insulin show reduced αMSH production and reduced expression of MC4Rs. These mice exhibit decreased sexual motivation as indicated by reduced mounting behavior during the initial sexual encounter; this phenotype is evident at four to six months of age, concurrent with increases in adiposity and insulin insensitivity [24].
POMC neurons project to nuclei that express Sim1, such as the paraventricular nucleus (PVH) [25], the supraoptic nucleus (SON), and regions of the amygdala including the medial amygdala (MeA) and basolateral amygdala (BLA) [26–28]. We previously showed that genetically re-expressing MC4Rs only in Sim1 neurons of MC4RKO mice restores both ejaculation and intromission efficiency at six months of age [29]. However, MC4Rs in Sim1 neurons also play a role in regulating feeding behavior [28,30,31]; these mice weighed twice the amount of control mice at the time of testing [29]. Therefore, it remains to be determined whether MC4Rs on Sim1 neurons directly regulate sexual function, or whether obesity is a necessary component of the dysfunction seen.
Oxytocin neurons comprise a large portion of the Sim1 neuron population [32], and are found in the PVH and SON [33], downstream of POMC neurons. Oxytocin neurons have been heavily implicated in mediating sexual behavior in both animals [34–36] and humans [37,38]. These neurons are postulated to mediate the effects of MC4 receptor agonists on partner preference and female sexual function [39,40]. We hypothesized that oxytocin neurons may mediate MC4R-driven sexual behavior in male mice. We therefore examined the ability of MC4Rs on Sim1 and oxytocin neurons to maintain sexual function using two-month old mice without age-related obesity. Our findings underline the feasibility of identifying specific neuronal targets for the treatment of male sexual dysfunction.
Results:
Before testing sexual behavior, the metabolic profile of MC4RKO mice was investigated, as metabolic diseases are known to be associated with sexual dysfunction. MC4 receptors play an important role in regulating satiety [21,41–44]. Previous studies have shown that MC4RKO mice gain a significant amount of weight over the course of their lives [28,30,31,44]. To confirm these findings, a body composition profile was obtained from the tested mice at two months of age. Nuclear magnetic resonance (NMR) revealed that MC4R manipulation affected weight gain. MC4RKO mice were significantly heavier than WT mice (p=0.0009; Fig. 1A). This increase in weight reflected an increase in fat mass (p<0.0001; Fig. 1B) but not in lean mass compared to WT (p=0.0895; Fig. 1C).
Fig. 1.
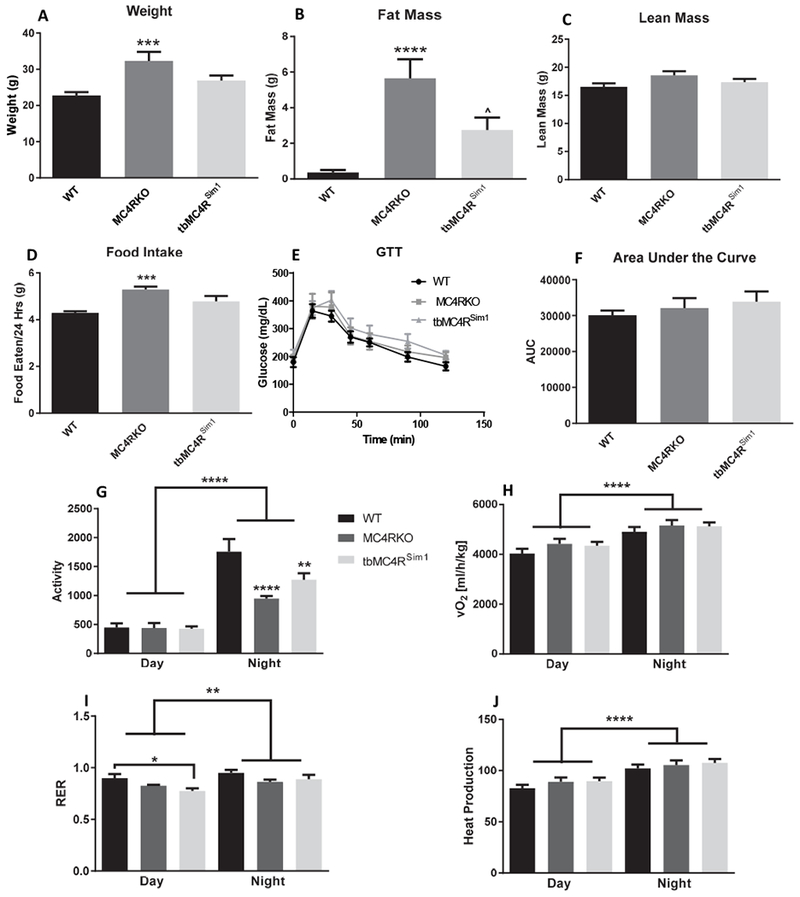
Nuclear Magnetic Resonance (NMR) at 2 months of age. (A) MC4RKO mice had increased weight gain compared to WT and tbMC4RSim1 mice using a one-way ANOVA (N=8-11). (B) Fat mass was increased in MC4RKO mice compared to WT. (C) There was no difference in lean mass at two months of age between groups. (D) MC4RKO mice had increased food intake compared to WT mice. (E) Glucose Tolerance Testing (GTT) revealed no changes between groups. (F) The GTT data was also represented as the area under the curve (AUC). (G) Activity was reduced in MC4RKO mice at night compared with WT mice. (H) VO2 was not significantly different. (I) The respiratory exchange ratio (RER) showed that MC4RKO mice was reduced at night compared to controls. (J) Metabolic cages normalized to lean body mass revealed no difference in heat production. *=compared to WT, #=compared to tbMC4RSim1
Sim1 neurons have been shown to play a role in energy expenditure as well [28,30,31], so we investigated whether the expression of MC4R only on Sim1 neurons rescued these parameters. tbMC4RSim1 body weights did not differ from WT mice (p=0.2105), although they trended towards weighing less than MC4RKO mice (p=0.0797; Fig. 1A). tbMC4RSim1 and WT mice did not differ in fat mass (p=0.0942; Fig. 1B) or lean mass (p>0.9999; Fig. 1C), but they had less fat mass compared to MC4RKO mice (p=0.0301; Fig. 1B). Thus, consistent with previous literature [30], Sim1 neurons appear to be involved with the body composition changes seen in MC4RKO mice.
Food intake over 24 hours was increased in MC4RKO mice (p=0.0006) compared to WT mice (Fig. 1D). Food intake was not different between tbMC4RSim1 and WT mice (p=0.1146) or MC4RKO mice (p=0.1061). The intermediate phenotype displayed by tbMC4RSim1 mice suggest that Sim1 neurons partially contribute to the hyperphagia of MC4R null mice. A glucose tolerance test found no significant difference across groups (F(2, 27)=0.6355, p=0.5374), suggesting that at two months of age, neither MC4RKO nor tbMC4RSim1 mice have a diabetic phenotype (Fig. 1E-F). These results were somewhat surprising, as previous studies have reported hyperglycemia in MC4RKO mice as young as 10 weeks old [44–46]. However, previous reports examined mice on a C57Bl6 background, a strain known to be susceptible to the development of type 2 diabetes [47], whereas the mice in this study were on a mixed C57Bl6/129S background.
Sim1 neurons [31] and MC4R [41] have been shown to affect locomotion. We therefore tested whether knocking out MC4R had an effect on mouse activity. Calorimetric cages demonstrated a significant interaction between genotype and time of day (F(2,45)=6.46; p=0.0034), which is consistent with the nocturnal nature of the mice. MC4RKO mice (p=0.0028) were less active at night compared to WT controls (p=0.002). tbMC4RSim1 were not significantly less active than WT (p=0.0708) or MC4RKO mice (p=0.3476; Fig. 1G). These findings suggest MC4R signaling in Sim1 neurons partially contributes to the locomotion phenotype of MC4RKO mice.
In accord with some [48], but not all previous studies [31], we found no compensatory increase in O2 consumption (F(2,46)=1.625, p=0.2080; Fig. 1H) or thermogenesis (F(2,46)=0.06763, p=0.9347; Fig. 1J) in MC4RKO mice in response to their obese phenotype. Genotype had a significant main effect on RER (F(2, 44)=5.285, p=0.0088) because of the difference between tbMC4RSim1 mice and WT (p=0.0225). This finding indicates greater utilization of fat for energy by tbMC4RSim1 mice during the day compared to control mice; no significance was seen during the night (p=0.4706; Fig. 1I). While Sim1 neuron ablation in adulthood can reduce energy expenditure [49], our studies indicate that this effect does not solely depend on disrupted MC4R signaling in Sim1 neurons.
A high-fat diet control group (WT-HFD) was utilized to control for weight gain independent of MC4R manipulation. These mice were age and weight matched to MC4RKO mice. An NMR study confirmed that the body weight of these mice equaled that of MC4RKO mice (p>0.9999; Fig. 2A).
Fig. 2.
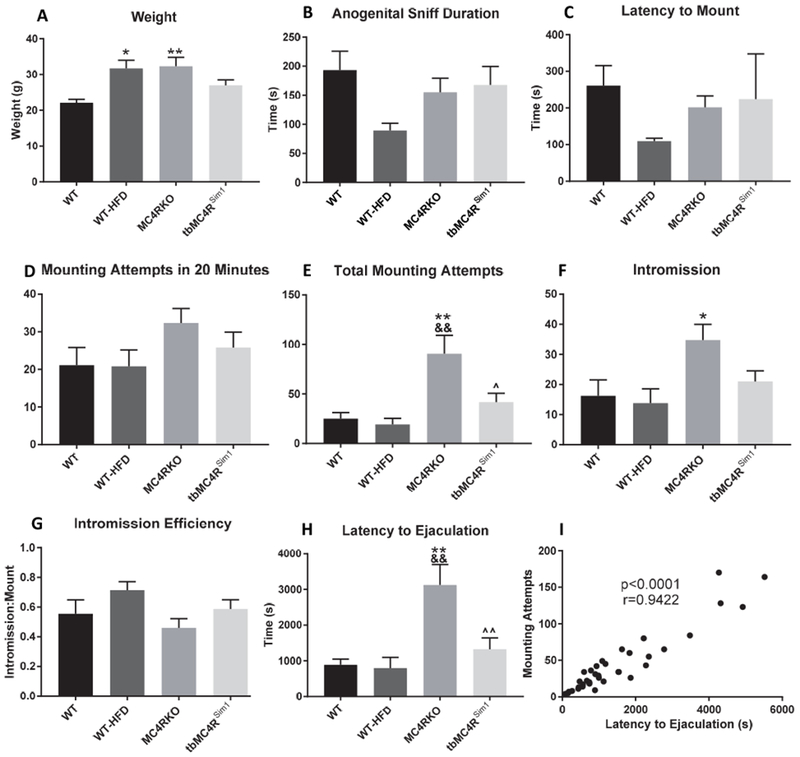
Sexual behavior in WT (n=9), WT-HFD (n=5), MC4RKO (n=9), and tbMC4RSim1 (n=10) mice. (A) WT-HFD mice were weight-matched to MC4RKO mice and both groups weighed significantly more than WT controls. (B) Time spent in the anogenital sniffing phase was not different across groups. (C) Latency to mount phase was not different across groups. (D) Mounting attempts within the first twenty minutes of sexual behavior was not different across groups. (E) Mounting attempts between MC4RKO and all other groups was significantly different. (F) Intromission number was not significantly different between groups. (G) The ratio of successful intromission to mounting attempts, an indicator of intromission efficiency, was not significant across groups. (H) Latency to ejaculation was significantly increased in MC4RKO mice compared to all other groups. (I) There was a significant correlation between latency to ejaculate and mounting attempts. *=compared to WT, &=compared to WT-HFD, ^=compared to MC4RKO
To test the role of the MC4R in sexual behavior, male mice were paired with females and their behavior was recorded. Measures of sexual motivation included anogenital sniffing duration, latency to mount, and number of mounting attempts in the first 20 minutes after pairing the mice [24,50–52]. There was no difference between groups in the duration of the anogenital sniffing phase (F(3,29)=1.62, p=0.2062; Fig. 2B), the latency to mounting behavior (F(3,29)=1.724, p=0.1841; Fig. 2C), or in the number of mounting attempts within the first 20 minutes of sexual activity (F(3,30)=1.439, p=0.2511; Fig. 2D). These results imply increased body weight alone at this age did not impede the level of interest that these mice had in initiating copulation. We therefore conclude that sexual motivation was not different between WT and MC4RKO mice.
To fully analyze sexual function, we examined sexual behavior of the paired mice over a six-hour period. Compared to both WT (p=0.0027) and WT-HFD controls (p=0.0061; Fig. 2E), MC4RKO mice had increased total mounting attempts. MC4RKO mice also reached intromission more times than WT (p=0.0482) but not WT-HFD littermates (p=0.0655; Fig. 2F). The intromission:mount ratio, which is an indication of intromission efficiency and erectile function [53–55] was not different between groups (F=1.558(3,29), p=0.2207; Fig. 2G). Interestingly, there was an increased latency to ejaculation in the MC4RKO mice compared to WT (p=0.0012) and WT-HFD mice (p=0.0049; Fig.2H). Nevertheless, all MC4RKO mice achieved ejaculation within the six hours filmed. We reasoned that if the WT mice were achieving ejaculation more successfully, they would not need as many mounting attempts as MC4RKO males. To visualize the relationship between latency to ejaculate and mounting attempts, we tested the correlation between the two measures and found it to be strongly positive (r=0.9422, p<0.0001, R2=0.8877; Fig. 2I). This finding indicates that the mice that took longer to ejaculate were the same mice that had higher mounting attempts. Overall, MC4RKO mice required a significantly longer period of time to reach ejaculation compared to WT mice and weight-matched controls, despite intact sexual motivation.
tbMC4RSim1 mice were generated to determine whether MC4 receptors on Sim1 neurons were critical to the effects seen. Sim1-cre controls and tbMC4RSim1 mice were tested for sexual behavior at the same time as the other groups. Sim1-cre control males did not differ from WT littermates in mounting frequency (p=0.3468), or any other measure, so the WT group is used as the control group in all figures for ease of interpretation. Although the tbMC4RSim1 mice did not exhibit any altered behavior compared to controls, mounting attempts (p=0.0303) and latency to ejaculation (p=0.009) were all significantly lower than MC4RKO mice (Fig. 2E-I). These results show that expressing MC4Rs only on Sim1 neurons prevents the changes in sexual behavior and ejaculation seen in MC4R null mice.
To narrow down the neurocircuitry involved further, we examined whether expression of MC4Rs on oxytocin neurons, which are a subset of Sim1 neurons in the PVH, could rescue the deficits in sexual performance seen in MC4R null mice [32]. tbMC4Roxt mice were generated and colocalization of MC4R and oxytocin neurons in Oxt-cre controls (Fig. 3A) and tbMC4Roxt mice was confirmed (Fig. 3B). Colocalization was also noted in the SON (Fig. 3C). tbMC4Roxt mice were paired with females at two months of age and sexual behavior was tested at the same time as other groups. Because Oxt-cre mice were not different from WT littermates in mounting frequency (p=0.929), or any other measure, WT mice are shown as the control group in the figures. tbMC4Roxt mice had weights that were comparable to MC4RKO mice (p>0.9999) and significantly greater than WT mice (p=0.0048; Fig. 4A). In terms of sexual motivation, tbMC4Roxt mice were no different from other groups. However, mounting attempts (p=0.0306) and latency to ejaculation (p=0.0273) were significantly lower than MC4RKO mice (Fig. 4B-E). These results show that MC4R expressed only on oxytocin neurons also prevent the alterations in ejaculation seen in MC4R null mice.
Fig. 3.
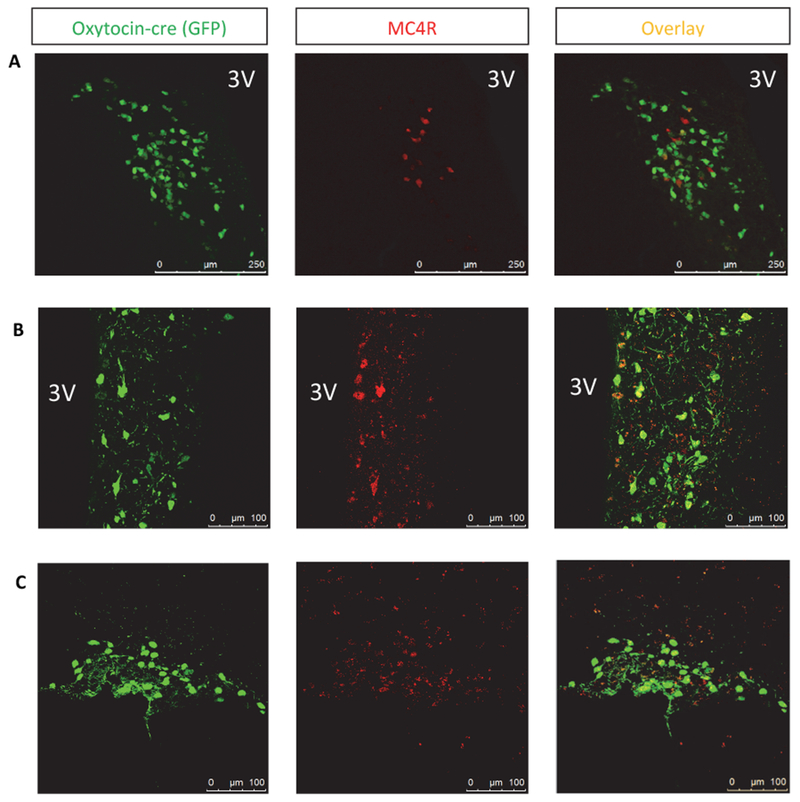
Co-localization was seen between GFP expression in oxytocin neurons (green) and MC4R (red) in both (A) Oxt-cre mice (20× magnification) and (B) tbMC4Roxt mice (40× magnification). Images show the hypothalamus immediately adjacent to the third ventricle (3V). Co-localization was also seen between oxytocin neurons and MC4R in the SON in (C) tbMC4Roxt mice (40× magnification).
Fig. 4.
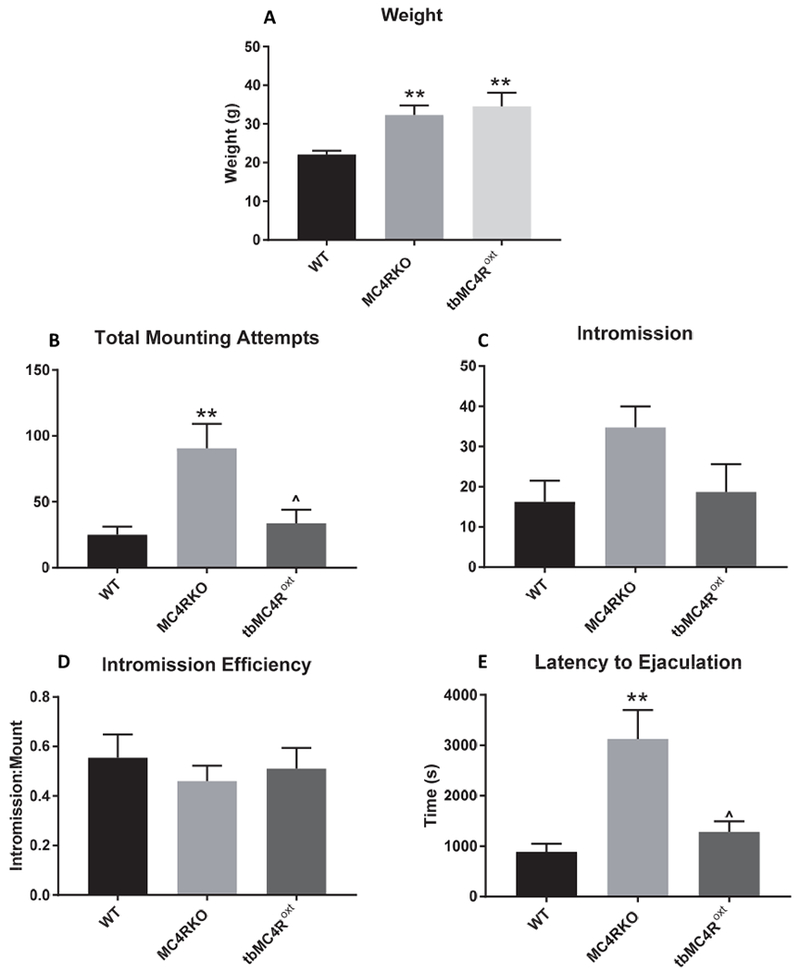
Sexual behavior in WT (n=9), MC4RKO (n=9), and tbMC4Roxt (n=6) mice. WT and MC4RKO data repeated for clarity. (A) tbMC4Roxt mice had a similar weight to MC4RKO mice and both groups weighed significantly more than WT controls. (B) Mounting attempts between MC4RKO and all other groups was significantly different by one-way ANOVA, with Bonferroni’s post-tests. (C) Intromission number was not significantly different between MC4RKO and all other littermates. (D) The ratio of successful intromission to mounting attempts, an indicator of intromission efficiency, was not significant across groups. (E) Latency to Ejaculation was significantly increased in MC4RKO mice compared to all other groups. *=compared to WT, ^=compared to MC4RKO
Serum was collected from mice to assess hormone concentrations. Results showed that knocking out MC4R had no effect on serum testosterone (F=0.3579, p=0.7838), estradiol (F=0.4287, p=0.7341), luteinizing hormone (LH; F=1.451, p=0.2499), or follicle stimulating hormone (FSH; F=3.418, p=0.3315) levels, and did not alter the LH/FSH ratio (F=0.7304, p=0.5426) (Fig. 5A-E). It is unlikely, therefore, that altered sexual function is secondary to hypogonadism.
Fig. 5.
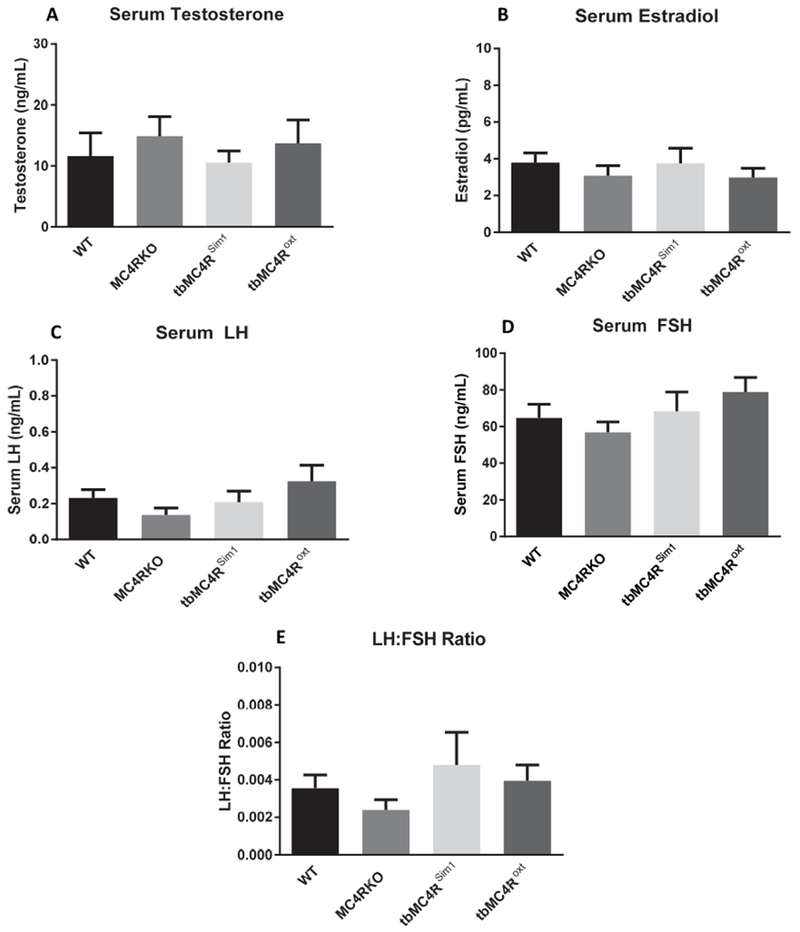
Reproductive hormone levels in MC4RKO mice and re-expression models. (A) Serum testosterone, (B) estradiol, (C) LH, (D) FSH, and (E) the LH:FSH ratio concentrations were not different using a one-way ANOVA (N=7-9)
Discussion:
In humans, a multitude of studies have found a link between sexual dysfunction and obesity [56–60]. Physical activity has been shown to reduce the risk of sexual dysfunction, while hypertension, diabetes, and cardiovascular disease increase the risk [60,61]. Men who lost weight through diet and exercise were able to improve erectile function [62,63]. These studies were primarily done in men older than 40 years of age. One study investigated men younger than 50 and found reduced erectile function associated with obesity due to reduced testosterone levels [64]. After the age of 50, men have been found to have more impairments in desire, erection, and ability to reach ejaculation [57]. Neuronal degeneration and vascular disease may contribute to the effects of age on sexual function; with age, the incidence of obesity [65], hypertension [66], diabetes [67], and heart disease [68] all increase. It is unclear whether human sexual dysfunction is primarily related to obesity directly or through its comorbidities.
Rodent studies can help to elucidate the mechanisms involved in obesity-related sexual dysfunction. Diet-induced obese mice have been shown to have deficits in erectile function, as shown by decreased intracavernosal pressure at roughly 4 months of age [69]. Similarly, obese MC4R null mice show evidence of erectile dysfunction [24,29]. Since the MC4R is known to play a role in both metabolic regulation [22,70–72] and sexual behavior [24,13,73,14,74], the present studies used young male MC4RKO mice to elucidate the role of the MC4R in sexual function independent of metabolic factors. We specifically examined whether these defects are secondary to increased body weight, activity levels, or glucose intolerance. We found that the MC4R was directly involved in ejaculation latency in male mice, and furthermore, that expressing MC4R exclusively in Sim1 or oxytocin neurons corrected this deficit.
At two months of age, MC4R disruption showed some degree of adiposity, although to a lesser degree than older mice. No glucose intolerance was seen at this age. Notably, the weight of MC4RK0 mice at this age was equivalent to the weight of six-month-old WT mice, which do not show impaired sexual function [29]. At two months, tbMC4Roxt mouse weights were equivalent to those of MC4RKO mice. In contrast, tbMC4RSim1 mice had somewhat normalized body weights, which is consistent with previously published findings [28]. Strikingly, both of these mouse models had normal sexual function compared to MC4RKO mice. Our results also demonstrated that the sexual function of age-matched, weight-matched WT-HFD mice was significantly better than MC4RKO mice. From these data, we conclude that the sexual deficits exhibited by two-month-old MC4RKO mice cannot be explained by metabolic disruption.
Because sexual function was intact in all groups that expressed MC4R, it is possible that the melanocortin system plays a neuroprotective role against the metabolic effects of obesity on copulation. In fact, the melanocortin system has been shown to have neuroprotective effects such as inhibiting mediators of inflammation such as NFKB [75], iNOS [76,77], COX-2 [76], and TNFα [77]. Upregulation of the MC4R has been found in surviving tissue following ischemic injury [78]. However, any neuroprotection that may be conferred by melanocortins is likely limited, as obese mice with intact melanocortin systems have been known to exhibit sexual dysfunction [79].
In these studies, we found that the MC4R, when isolated from its effects on body weight, is particularly involved in ejaculation in mice. Melanocortins and oxytocin have been implicated in central ejaculatory control [80–83]. In human studies involving PET imaging, the midbrain, thalamic nuclei, and cerebellum are associated with ejaculation [84,85]. Animal studies have also implicated the MPOA, the nucleus paragigantocellular, and the PVH in ejaculatory function [86,87]. Parasympathetic control drives penile erection [88], and the emission phase of ejaculation requires coordinated control of both parasympathetic and sympathetic nerves [86]. Neurons in the PVH, including oxytocin neurons, project to the brain stem and spinal cord autonomic systems [89–91], so it is possible for these neurons to be involved in ejaculatory processes.
The fact that penile erection and sexual motivation were unaffected in our study was a surprising finding as previous studies have implicated the MC4R in these aspects of sexual function [24,73,14,74]. Besides overlooking the potential confound of metabolic dysfunction, many of these other studies examined penile erection outside of a copulatory setting. Given that the current experiments focused on how the MC4R affected erection during copulation, they may not generalize to non-copulatory settings.
Because of the importance of erectile dysfunction in humans, ejaculation is sometimes underemphasized in rodent studies of sexual function. However, we previously reported that MC4RKO male mice at six months of age are unable to reach ejaculation [29]. Van der Ploeg and colleagues found reduced ejaculatory efficiency in MC4RKO mice at three to six months of age [14]. In both these studies of older mice, the impaired ejaculation was found in conjunction with impaired erectile function, while here we found only ejaculatory impairment at two months of age. These data suggest a critical role of melanocortin signaling in ejaculation which may merge with age and body weight-dependent effects on erectile function. Melanocortins may therefore have unexplored potential as a treatment for ejaculatory disorders.
MC4R expression by Sim1 neurons prevented sexual deficits. Sim1 and MC4R expression overlap in the PVH, supraoptic nucleus, anterior hypothalamic nucleus, and MeA [92,25]. To narrow down the location and neuronal subtype involved in this effect, we chose to examine the role of oxytocin neurons. Sim1 haploinsufficiency leads to an 80% reduction in oxytocin expression in mice [32]. Oxytocin neurons are found in the PVH and SON [93]. Not all Sim1 neurons in the PVH or SON also express oxytocin, however [32,49]; this fact may explain the weight changes that were seen in tbMC4RSim1 but not in tbMC4Roxt mice.
Oxytocin’s role in regulating erection and ejaculation has been extensively studied in rodents [94]. Oxytocinergic projections from the PVH to the hippocampus, medulla oblongata and spinal cord facilitate penile erection [95]. Our results reveal MC4R signaling in oxytocin neurons permits ejaculation but argue against this pathway directly mediating erectile function. Administration of both systemic and ICV oxytocin has been found to facilitate ejaculatory function [96], while IV, ICV, and intrathecal administration of an antagonist of oxytocin receptor (GSK557296) has been found to inhibit ejaculation [97]. Similar studies have found a decreased latency to ejaculate in rabbits and rats after administration of oxytocin [98,96]. The effect of oxytocin receptor ligands on the ejaculatory response may be due to modulation of serotonin and dopamine neurotransmission [99]. The injection of a specific D4 agonist into the PVN induced penile erection in rats, which was blocked by the intraventricular injection of an oxytocin inhibitor [100], suggesting some interaction between dopamine and oxytocin in the regulation of sexual function. Studies have also implicated the involvement of serotonin in this neurocircuitry. Rats that do not express the serotonin transporter (SERT) exhibit delayed ejaculation compared to controls [101]. Using pharmacological studies with serotonin agonists and antagonists, 5HT1A and 5HT7 receptors were implicated in ejaculatory function [101]. The MC4R has been found to be downstream of the action of serotonin agonism in a study of hypophagia [102]. Similar studies using serotonin and dopamine agonists in MC4R null mice could elucidate interactions between serotonin, dopamine, and melanocortin neurocircuitry in sexual behavior.
Delayed ejaculation is a poorly understood, but distressing condition. Pharmaceutical therapy for this disorder is under investigation [103]. One study found that intranasal oxytocin resulted in an increased latency to ejaculation compared to placebo [38], but another study failed to replicate this finding [104]. Both studies involved healthy men, rather than men with ejaculatory dysfunction. Our results suggest that targeted oxytocin treatment may warrant further exploration.
Given that our studies used re-expression models, they address the sufficiency but not the necessity of the pathways investigated. Interestingly, another study using an oxytocin knock-out mouse model found that oxytocin was not necessary for sexual behavior such as anogenital sniffing, mounting, intromission, and ejaculation [105]. Redundant pathways supporting ejaculatory function likely exist. For example, oxytocin knock-out males may exhibit normal sexual behavior due to parallel dopamine neurocircuitry [105]. It should also be recognized that when gene ablation occurs prenatally, parallel pathways may develop abnormal strength to compensate for the loss of a neuropeptide such as oxytocin. The use of such technology as DREADD and optogenetics may allow for temporally controlled suppression of these pathways to improve understanding of how the melanocortin system works physiologically to impact sexual function.
In conclusion, we found that melanocortin signaling via the MC4R on Sim1-expressing neurons and oxytocin neurons enables ejaculation in young male mice. Furthermore, we confirmed that this effect, unlike melanocortin effects on sexual motivation and erectile function, was not related to the metabolic effects of MC4R deletion. These results emphasize the importance of considering the action of age and age-related obesity on sexual behavior. These data provide new insight into the neurocircuitry underlying ejaculation and help to reconcile conflicting literature on sexual dysfunction in the MC4RKO mouse. Elucidating this neurocircuitry will lead to better treatment options for men with sexual dysfunction.
Materials and Methods:
Animal Production and Care:
To generate the MC4RKO mice, heterozygous mice were purchased from The Jackson Laboratory (loxTB Mc4r, 006414). In these mice, a transcription blocker in the promoter of the MC4R gene prevents MC4R expression. This transcription blocker is flanked by loxp sites, such that cre recombinase exposure results in its removal, allowing expression of MC4R. Therefore, mice expressing cre on Sim1 neurons, and homozygous for the floxed MC4R gene express MC4R only on Sim1-expressing neurons (tbMC4RSim1). To generate tbMC4RSim1 mice, a Sim1-cre-expressing mouse line (The Jackson Laboratory, 006395) was bred with the loxTB MC4R mice. These models have been published previously [28] and expression patterns were verified in our lab using immunohistochemistry [29]. Similarly, oxytocin-ires cre mice (Oxt-cre; The Jackson Laboratory, 024234) were used to generate mice that expressed MC4R only on oxytocin neurons (tbMC4Roxt). Breeding pairs were designed so that one parent would be heterozygous for the loxTB MC4R and the other would be both heterozygous for the loxTB MC4R as well as hemizygous for either Sim1-cre or oxytocin-cre. Littermates therefore included wild-type (WT), cre controls, MC4RKO, and mice with targeted MC4R re-expression. For detection of cre expressing neurons, mice were crossed with a loxp flanked GFP reporter line (The Jackson Laboratory, 024750). All mice were on a mixed C57Bl6/129S background. Genotyping for the mouse lines was performed by Transnetyx, Inc. using real-time PCR.
All mice used in this study were kept in accordance with the University’s IACUC guidelines. Mice were housed in animal facilities where they were given ad libidum food and water on a 12:12 light dark cycle. Except for the mice placed in calorimetric cages, the same cohorts used for sexual behavior testing were used for all tests. All tests of MC4RKO, control groups, tbMC4RSim1, and tbMC4Roxt mice were performed concurrently.
Metabolic Studies:
The morning after sexual behavior testing, nuclear magnetic resonance (NMR) was used to assess fat content of the mice (BrukerOptics). At 9-12 weeks old, the mice were given a glucose tolerance test (GTT). The morning of the GTT, mice were fasted for six hours on ALPHA-dri bedding (Shepherd) starting at 8am. At 2pm, baseline glucose levels are obtained by measuring tail-vein blood. A 2g/kg dose of dextrose was administered intraperitoneally and subsequent blood glucose levels were measured at 15, 30, 45, 60, 90, and 120 minutes post-injection using a veterinary glucometer (AlphaTRAK 2, ADW Diabetes).
Mice were placed in calorimetric cages (Columbus Instruments’ Comprehensive Lab Animal Monitoring System) at two months of age in order to assess food intake, water intake, physical activity, and energy expenditure. Mice were allowed to acclimate to the cages for over 24 hours before data was collected. Data was collected every 20 minutes for 72 hours before returning each mouse to their home cage (Oxymax for Windows).
Diet Induced Obesity:
A subset of WT littermate mice were given a high-fat diet (HFD; OpenSource Diets, 60% fat content) instead of standard chow starting at 3 weeks of age. Mice were weighed weekly until their weights were comparable to MC4RKO mice at the time of their sexual behavior testing.
Sexual Behavior Studies:
Male mice at the age of 7-8 weeks were exposed to hormone primed, ovariectomized female mice three separate times to gain sexual experience. To prime the females, a subcutaneous 100 μL dose of β-estradiol-3-benzoate in sesame oil (200μg/mL) was given 48 hours prior to sexual behavior, and then an intraperitoneal 125 μL dose of progesterone (4mg/mL) was administered 7 hours prior to pairing [106]. Pairing occurred from 8pm-9am in the male’s cage, during the normal period of activity for mice. The fourth pairing was videotaped (DVR Swann 4500 and T850 Day and Night Security Camera security system) and analyzed for sexual behaviors between 8pm-2am; this time period was determined during preliminary experiments to include the majority of relevant sexual behaviors.
Sexual behaviors measured included anogenital sniffing, mounting, intromission, and ejaculation. Anogenital sniffing was defined as the period in which the male sniffed the female’s genitals prior to the first mounting attempt. Mounting attempts were defined as any time the male placed his forepaws onto the female’s back and attempted to thrust. Successful intromission was defined as deep, slow rhythmic thrusting during a mounting attempt. The intromission to mount ratio was defined as the number of times intromission was reached per total mounting attempts. Ejaculation was identified by observing the male freeze and fall over following intromission.
Hormone Assays:
Submandibular blood samples were taken at week 10-13. Serum was obtained by placing samples in the centrifuge for 10 minutes at 4°C at 4472 RCF and then collecting the serum layer. Analysis was done using testosterone and estradiol ELISA kits (CalBiotech). LH and FSH was measured by the University of Virginia Ligand Assay and Analysis Core using a multiplex assay.
Free-Floating Brain Section Immunohistochemistry:
Upon euthanasia, each mouse was anesthetized with a ketamine/xylazine mixture (100mg/kg ketamine, 10mg/kg xylazine) and perfused with 10% formaldehyde. The brain was removed and stored in 10% formaldehyde for 24 hours, followed by increasing concentrations of sucrose in PBS up to 30% sucrose until sectioned with a freezing microtome. The brain was sectioned in 35-40μm slices and then stored in cryoprotectant (20% ethylene glycol with 20% glycerol in PBS at pH 7.4) at −20°C. To verify that oxytocin neurons expressed MC4Rs in targeted nuclei, brain sections were blocked with 5% donkey antisera and incubated overnight with rabbit anti-MC4R (Abcam ab24233) and chicken anti-GFP (Aves Labs Inc, IgY0511FP12). Sections were exposed to goat anti-chicken Alexa Fluor 488 (A11039, life technologies) and donkey anti-rabbit Alexa Fluor 594 (A21207, Life Technologies) secondary antisera as previously described [29] and observed using a confocal microscope (TCS SP5 Laser Scanning Confocal Microscope, Leica Microsystems).
Data Analysis:
All data in figures are represented as mean ± SEM. One-way and two-way ANOVA tests were used to compare more than two groups followed by Bonferroni’s post-tests. Correlation tests were used to determine the relationship between two continuous variables, with results reported as r values. Statistical significance was defined as p<0.05. In figure legends, *, p<0.05; **, p<0.01; ***p<0.001; ****p<0.0001.
Acknowledgements:
This work was supported by NIH award R01 HD081792 to JWH. We would like to acknowledge the University of Toledo Advanced Microscopy & Imaging Center and the University of Virginia Ligand Assay and Analysis Core for the analysis of LH/FSH serum concentrations. The University of Virginia Center for Research in Reproduction Ligand Assay and Analysis Core is supported by the Eunice Kennedy Shriver NICHD/NIH (NCTRI) Grant P50-HD28934.
Footnotes
Conflict of interest: The authors declare that they have no conflict of interest.
Compliance with Ethical Standards:
Research involving Human Participants and/or Animals: All procedures performed in studies involving animals were in accordance with the ethical standards of the institution at which the studies were conducted. This article does not contain any studies with human participants performed by any of the authors.
References:
- 1.Lewis RW, Fugl-Meyer KS, Corona G, Hayes RD, Laumann EO, Moreira ED Jr., Rellini AH, Segraves T (2010) Definitions/epidemiology/risk factors for sexual dysfunction. J Sex Med 7 (4 Pt 2):1598–1607. doi: 10.1m/j.1743-6109.2010.01778.x [DOI] [PubMed] [Google Scholar]
- 2.Hatzimouratidis K, Amar E, Eardley I, Giuliano F, Hatzichristou D, Montorsi F, Vardi Y, Wespes E (2010) Guidelines on male sexual dysfunction: erectile dysfunction and premature ejaculation. European urology 57 (5):804–814. doi: 10.1016/j.eururo.2010.02.020 [DOI] [PubMed] [Google Scholar]
- 3.McCabe MP, Sharlip ID, Lewis R, Atalla E, Balon R, Fisher AD, Laumann E, Lee SW, Segraves RT (2016) Incidence and Prevalence of Sexual Dysfunction in Women and Men: A Consensus Statement from the Fourth International Consultation on Sexual Medicine 2015. The journal of sexual medicine 13 (2):144–152. doi: 10.1016/j.jsxm.2015.12.034 [DOI] [PubMed] [Google Scholar]
- 4.Nicolosi A, Moreira ED Jr, Shirai M, Bin Mohd Tambi MI, Glasser DB (2003) Epidemiology of erectile dysfunction in four countries: cross-national study of the prevalence and correlates of erectile dysfunction. Urology 61 (1):201–206 [DOI] [PubMed] [Google Scholar]
- 5.Hatzimouratidis K (2007) Epidemiology of male sexual dysfunction. American journal of men’s health 1 (2):103–125. doi: 10.1177/1557988306298006 [DOI] [PubMed] [Google Scholar]
- 6.Mitchell KR, Mercer CH, Ploubidis GB, Jones KG, Datta J, Field N, Copas AJ, Tanton C, Erens B, Sonnenberg P, Clifton S, Macdowall W, Phelps A, Johnson AM, Wellings K (2013) Sexual function in Britain: findings from the third National Survey of Sexual Attitudes and Lifestyles (Natsal-3). Lancet (London, England) 382 (9907):1817–1829. doi: 10.1016/s0140-6736(13)62366-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gades NM, Jacobson DJ, McGree ME, St Sauver JL, Lieber MM, Nehra A, Girman CJ, Jacobsen SJ (2009) Longitudinal evaluation of sexual function in a male cohort: the Olmsted county study of urinary symptoms and health status among men. The journal of sexual medicine 6 (9):2455–2466. doi: 10.1111/j.1743-6109.2009.01374.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Thorve VS, Kshirsagar AD, Vyawahare NS, Joshi VS, Ingale KG, Mohite RJ (2011) Diabetes-induced erectile dysfunction: epidemiology, pathophysiology and management. Journal of diabetes and its complications 25 (2):129–136. doi: 10.1016/j.jdiacomp.2010.03.003 [DOI] [PubMed] [Google Scholar]
- 9.Giuliano F, Allard J (2001) Dopamine and male sexual function. European urology 40 (6):601–608 [DOI] [PubMed] [Google Scholar]
- 10.Wessells H, Gralnek D, Dorr R, Hruby VJ, Hadley ME, Levine N (2000) Effect of an alpha-melanocyte stimulating hormone analog on penile erection and sexual desire in men with organic erectile dysfunction. Urology 56 (4):641–646 [DOI] [PubMed] [Google Scholar]
- 11.Safarinejad MR, Hosseini SY (2008) Salvage of sildenafil failures with bremelanotide: a randomized, double-blind, placebo controlled study. The Journal of urology 179 (3): 1066–1071. doi: 10.1016/j.juro.2007.10.063 [DOI] [PubMed] [Google Scholar]
- 12.Shadiack AM, Sharma SD, Earle DC, Spana C, Hallam TJ (2007) Melanocortins in the treatment of male and female sexual dysfunction. Current topics in medicinal chemistry 7 (11): 1137–1144 [DOI] [PubMed] [Google Scholar]
- 13.Hadley ME (2005) Discovery that a melanocortin regulates sexual functions in male and female humans. Peptides 26 (10):1687–1689. doi: 10.1016/j.peptides.2005.01.023 [DOI] [PubMed] [Google Scholar]
- 14.Van der Ploeg LH, Martin WJ, Howard AD, Nargund RP, Austin CP, Guan X, Drisko J, Cashen D, Sebhat I, Patchett AA, Figueroa DJ, DiLella AG, Connolly BM, Weinberg DH, Tan CP, Palyha OC, Pong SS, MacNeil T, Rosenblum C, Vongs A, Tang R, Yu H, Sailer AW, Fong TM, Huang C, Tota MR, Chang RS, Stearns R, Tamvakopoulos C, Christ G, Drazen DL, Spar BD, Nelson RJ, MacIntyre DE (2002) A role for the melanocortin 4 receptor in sexual function. Proc Natl Acad Sci U S A 99 (17): 11381–11386. doi: 10.1073/pnas.172378699 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shadiack AM, Althof S (2008) Preclinical effects of melanocortins in male sexual dysfunction. International journal of impotence research 20 Suppl 1:S11–16. doi: 10.1038/ijir.2008.17 [DOI] [PubMed] [Google Scholar]
- 16.Cone RD (2005) Anatomy and regulation of the central melanocortin system. Nature neuroscience 8 (5):571–578. doi: 10.1038/nn1455 [DOI] [PubMed] [Google Scholar]
- 17.Yang Y (2011) Structure, function and regulation of the melanocortin receptors. European journal of pharmacology 660 (1): 125–130. doi: 10.1016/j.ejphar.2010.12.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schioth HB, Chhajlani V, Muceniece R, Klusa V, Wikberg JE (1996) Major pharmacological distinction of the ACTH receptor from other melanocortin receptors. Life sciences 59 (10):797–801 [DOI] [PubMed] [Google Scholar]
- 19.Mountjoy KG (2010) Distribution and function of melanocortin receptors within the brain. Advances in experimental medicine and biology 681:29–48. doi: 10.1007/978-1-4419-6354-3_3 [DOI] [PubMed] [Google Scholar]
- 20.Rodrigues AR, Almeida H, Gouveia AM (2015) Intracellular signaling mechanisms of the melanocortin receptors: current state of the art. Cellular and molecular life sciences : CMLS 72 (7): 1331–1345. doi: 10.1007/s00018-014-1800-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Adan RA, Tiesjema B, Hillebrand JJ, la Fleur SE, Kas MJ, de Krom M (2006) The MC4 receptor and control of appetite. British journal of pharmacology 149 (7):815–827. doi: 10.1038/sj.bjp.0706929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Huszar D, Lynch CA, Fairchild-Huntress V, Dunmore JH, Fang Q, Berkemeier LR, Gu W, Kesterson RA, Boston BA, Cone RD, Smith FJ, Campfield LA, Burn P, Lee F (1997) Targeted Disruption of the Melanocortin-4 Receptor Results in Obesity in Mice. Cell 88 (1): 131–141. doi: 10.1016/S0092-8674(00)81865-6 [DOI] [PubMed] [Google Scholar]
- 23.Argiolas A, Melis MR, Murgia S, Schioth HB (2000) ACTH- and alpha-MSH-induced grooming, stretching, yawning and penile erection in male rats: site of action in the brain and role of melanocortin receptors. Brain Res Bull 51 (5):425–431 [DOI] [PubMed] [Google Scholar]
- 24.Faulkner LD, Dowling AR, Stuart RC, Nillni EA, Hill JW (2015) Reduced Melanocortin Production Causes Sexual Dysfunction in Male Mice With POMC Neuronal Insulin and Leptin Insensitivity. Endocrinology 156 (4): 1372–1385. doi: 10.1210/en.2014-1788 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tao YX (2010) The melanocortin-4 receptor: physiology, pharmacology, and pathophysiology. Endocrine reviews 31 (4):506–543. doi: 10.1210/er.2009-0037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tolson KP, Gemelli T, Gautron L, Elmquist JK, Zinn AR, Kublaoui BM (2010) Postnatal Sim1 deficiency causes hyperphagic obesity and reduced Mc4r and oxytocin expression. The Journal of neuroscience : the official journal of the Society for Neuroscience 30 (10):3803–3812. doi: 10.1523/jneurosci.5444-09.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Michaud JL, Rosenquist T, May NR, Fan CM (1998) Development of neuroendocrine lineages requires the bHLH-PAS transcription factor SIM1. Genes & development 12 (20):3264–3275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T, Ferreira M, Tang V, McGovern RA, Kenny CD, Christiansen LM, Edelstein E, Choi B, Boss O, Aschkenasi C, Zhang CY, Mountjoy K, Kishi T, Elmquist JK, Lowell BB (2005) Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell 123 (3):493–505. doi: 10.1016/j.cell.2005.08.035 [DOI] [PubMed] [Google Scholar]
- 29.Semple E, Hill JW (2018) Sim1 Neurons Are Sufficient for MC4R-Mediated Sexual Function in Male Mice. Endocrinology 159 (1):439–449. doi: 10.1210/en.2017-00488 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Shah BP, Vong L, Olson DP, Koda S, Krashes MJ, Ye C, Yang Z, Fuller PM, Elmquist JK, Lowell BB (2014) MC4R-expressing glutamatergic neurons in the paraventricular hypothalamus regulate feeding and are synaptically connected to the parabrachial nucleus. Proceedings of the National Academy of Sciences of the United States of America 111 (36):13193–13198. doi: 10.1073/pnas.1407843111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Xu Y, Wu Z, Sun H, Zhu Y, Kim ER, Lowell BB, Arenkiel BR, Xu Y, Tong Q (2013) Glutamate mediates the function of melanocortin receptor 4 on Sim1 neurons in body weight regulation. Cell metabolism 18 (6):860–870. doi: 10.1016/j.cmet.2013.11.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kublaoui BM, Gemelli T, Tolson KP, Wang Y, Zinn AR (2008) Oxytocin deficiency mediates hyperphagic obesity of Sim1 haploinsufficient mice. Molecular endocrinology (Baltimore, Md) 22 (7):1723–1734. doi: 10.1210/me.2008-0067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Barberis C, Mouillac B, Durroux T (1998) Structural bases of vasopressin/oxytocin receptor function. The Journal of endocrinology 156 (2):223–229 [DOI] [PubMed] [Google Scholar]
- 34.Argiolas A, Melis MR, Gessa GL (1988) Yawning and penile erection: central dopamine-oxytocin-adrenocorticotropin connection. Annals of the New York Academy of Sciences 525:330–337 [DOI] [PubMed] [Google Scholar]
- 35.Melis MR, Melis T, Cocco C, Succu S, Sanna F, Pillolla G, Boi A, Ferri GL, Argiolas A (2007) Oxytocin injected into the ventral tegmental area induces penile erection and increases extracellular dopamine in the nucleus accumbens and paraventricular nucleus of the hypothalamus of male rats. The European journal of neuroscience 26 (4):1026–1035. doi: 10.1111/j.1460-9568.2007.05721.x [DOI] [PubMed] [Google Scholar]
- 36.Succu S, Sanna F, Cocco C, Melis T, Boi A, Ferri GL, Argiolas A, Melis MR (2008) Oxytocin induces penile erection when injected into the ventral tegmental area of male rats: role of nitric oxide and cyclic GMP. The European journal of neuroscience 28 (4):813–821. doi: 10.1111/j.1460-9568.2008.06385.x [DOI] [PubMed] [Google Scholar]
- 37.Behnia B, Heinrichs M, Bergmann W, Jung S, Germann J, Schedlowski M, Hartmann U, Kruger TH (2014) Differential effects of intranasal oxytocin on sexual experiences and partner interactions in couples. Hormones and behavior 65 (3):308–318. doi: 10.1016/j.yhbeh.2014.01.009 [DOI] [PubMed] [Google Scholar]
- 38.Burri A, Heinrichs M, Schedlowski M, Kruger TH (2008) The acute effects of intranasal oxytocin administration on endocrine and sexual function in males. Psychoneuroendocrinology 33 (5):591–600. doi: 10.1016/j.psyneuen.2008.01.014 [DOI] [PubMed] [Google Scholar]
- 39.Modi ME, Inoue K, Barrett CE, Kittelberger KA, Smith DG, Landgraf R, Young LJ (2015) Melanocortin Receptor Agonists Facilitate Oxytocin-Dependent Partner Preference Formation in the Prairie Vole. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology 40 (8):1856–1865. doi: 10.1038/npp.2015.35 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gelez H, Poirier S, Facchinetti P, Allers KA, Wayman C, Alexandre L, Giuliano F (2010) Neuroanatomical evidence for a role of central melanocortin-4 receptors and oxytocin in the efferent control of the rodent clitoris and vagina. The journal of sexual medicine 7 (6):2056–2067. doi: 10.1111/j.1743-6109.2010.01760.x [DOI] [PubMed] [Google Scholar]
- 41.Chen AS, Metzger JM, Trumbauer ME, Guan XM, Yu H, Frazier EG, Marsh DJ, Forrest MJ, Gopal-Truter S, Fisher J, Camacho RE, Strack AM, Mellin TN, MacIntyre DE, Chen HY, Van der Ploeg LH (2000) Role of the melanocortin-4 receptor in metabolic rate and food intake in mice. Transgenic research 9 (2): 145–154 [DOI] [PubMed] [Google Scholar]
- 42.Garfield AS, Li C, Madara JC, Shah BP, Webber E, Steger JS, Campbell JN, Gavrilova O, Lee CE, Olson DP, Elmquist JK, Tannous BA, Krashes MJ, Lowell BB (2015) A neural basis for melanocortin-4 receptor-regulated appetite. Nature neuroscience 18 (6):863–871. doi : 10.1038/nn.4011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Glavas MM, Joachim SE, Draper SJ, Smith MS, Grove KL (2007) Melanocortinergic Activation by Melanotan II Inhibits Feeding and Increases Uncoupling Protein 1 Messenger Ribonucleic Acid in the Developing Rat. Endocrinology 148 (7):3279–3287. doi: 10.1210/en.2007-0184 [DOI] [PubMed] [Google Scholar]
- 44.Huszar D, Lynch CA, Fairchild-Huntress V, Dunmore JH, Fang Q, Berkemeier LR, Gu W, Kesterson RA, Boston BA, Cone RD, Smith FJ, Campfield LA, Burn P, Lee F (1997) Targeted disruption of the melanocortin-4 receptor results in obesity in mice. Cell 88 (1): 131–141 [DOI] [PubMed] [Google Scholar]
- 45.Ste Marie L, Miura GI, Marsh DJ, Yagaloff K, Palmiter RD (2000) A metabolic defect promotes obesity in mice lacking melanocortin-4 receptors. Proceedings of the National Academy of Sciences of the United States of America 97 (22): 12339–12344. doi : 10.1073/pnas.220409497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tallam LS, Stec DE, Willis MA, da Silva AA, Hall JE (2005) Melanocortin-4 receptor-deficient mice are not hypertensive or salt-sensitive despite obesity, hyperinsulinemia, and hyperleptinemia. Hypertension (Dallas, Tex : 1979) 46 (2):326–332. doi: 10.1161/01.HYP.0000175474.99326.bf [DOI] [PubMed] [Google Scholar]
- 47.Surwit RS, Kuhn CM, Cochrane C, McCubbin JA, Feinglos MN (1988) Diet-induced type II diabetes in C57BL/6J mice. Diabetes 37 (9): 1163–1167 [DOI] [PubMed] [Google Scholar]
- 48.Butler AA, Marks DL, Fan W, Kuhn CM, Bartolome M, Cone RD (2001) Melanocortin-4 receptor is required for acute homeostatic responses to increased dietary fat. Nature neuroscience 4 (6):605–611. doi: 10.1038/88423 [DOI] [PubMed] [Google Scholar]
- 49.Xi D, Gandhi N, Lai M, Kublaoui BM (2012) Ablation of Sim1 neurons causes obesity through hyperphagia and reduced energy expenditure. PloS one 7 (4):e36453. doi: 10.1371/journal.pone.0036453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hull EM, Dominguez JM (2007) Sexual behavior in male rodents. Hormones and behavior 52 (1):45–55. doi: 10.1016/j.yhbeh.2007.03.030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dewsbury DA (1969) Copulatory behaviour of rats (Rattus norvegicus) as a function of prior copulatory experience. Animal behaviour 17 (2):217–223 [DOI] [PubMed] [Google Scholar]
- 52.Swaney WT, Dubose BN, Curley JP, Champagne FA (2012) Sexual experience affects reproductive behavior and preoptic androgen receptors in male mice. Hormones and behavior 61 (4):472–478. doi: 10.1016/j.yhbeh.2012.01.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Allouh MZ (2015) Effects of swimming activity on the copulatory behavior of sexually active male rats. International journal of impotence research 27 (3): 113–117. doi: 10.1038/ijir.2014.42 [DOI] [PubMed] [Google Scholar]
- 54.Gajbhiye SV, Jadhav KS, Marathe PA, Pawar DB (2015) Animal models of erectile dysfunction. Indian Journal of Urology : IJU : Journal of the Urological Society of India 31 (1):15–21. doi: 10.4103/0970-1591.128496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.McMurray G, Casey JH, Naylor AM (2006) Animal models in urological disease and sexual dysfunction. British journal of pharmacology 147 (Suppl 2):S62–79. doi: 10.1038/sj.bjp.0706630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hackett G (2009) The burden and extent of comorbid conditions in patients with erectile dysfunction. International journal of clinical practice 63 (8): 1205–1213. doi: 10.1111/j.1742-1241.2009.02088.x [DOI] [PubMed] [Google Scholar]
- 57.Bacon CG, Mittleman MA, Kawachi I, Giovannucci E, Glasser DB, Rimm EB (2003) Sexual function in men older than 50 years of age: results from the health professionals follow-up study. Annals of internal medicine 139 (3):161–168 [DOI] [PubMed] [Google Scholar]
- 58.Han TS, Tajar A, O’Neill TW, Jiang M, Bartfai G, Boonen S, Casanueva F, Finn JD, Forti G, Giwercman A, Huhtaniemi IT, Kula K, Pendleton N, Punab M, Silman AJ, Vanderschueren D, Lean ME, Wu FC (2011) Impaired quality of life and sexual function in overweight and obese men: the European Male Ageing Study. European journal of endocrinology 164 (6): 1003–1011. doi: 10.1530/eje-10-1129 [DOI] [PubMed] [Google Scholar]
- 59.Kolotkin RL, Zunker C, Ostbye T (2012) Sexual functioning and obesity: a review. Obesity (Silver Spring, Md) 20 (12):2325–2333. doi: 10.1038/oby.2012.104 [DOI] [PubMed] [Google Scholar]
- 60.Bacon CG, Mittleman MA, Kawachi I, Giovannucci E, Glasser DB, Rimm EB (2006) A prospective study of risk factors for erectile dysfunction. The Journal of urology 176 (1):217–221. doi: 10.1016/s0022-5347(06)00589-1 [DOI] [PubMed] [Google Scholar]
- 61.Dong JY, Zhang YH, Qin LQ (2011) Erectile dysfunction and risk of cardiovascular disease: meta-analysis of prospective cohort studies. Journal of the American College of Cardiology 58 (13):1378–1385. doi: 10.1016/j.jacc.2011.06.024 [DOI] [PubMed] [Google Scholar]
- 62.Esposito K, Giugliano F, Di Palo C, Giugliano G, Marfella R, D’Andrea F, D’Armiento M, Giugliano D (2004) Effect of lifestyle changes on erectile dysfunction in obese men: a randomized controlled trial. Jama 291 (24):2978–2984. doi: 10.1001/jama.291.24.2978 [DOI] [PubMed] [Google Scholar]
- 63.Horasanli K, Boylu U, Kendirci M, Miroglu C (2008) Do lifestyle changes work for improving erectile dysfunction? Asian journal of andrology 10 (1):28–35. doi: 10.1111/j.1745-7262.2008.00363.x [DOI] [PubMed] [Google Scholar]
- 64.Jastrzebska S, Walczak-Jedrzejowska R, Kramek E, Marchlewska K, Oszukowska E, Filipiak E, Kula K, Slowikowska-Hilczer J (2014) Relationship between sexual function, body mass index and levels of sex steroid hormones in young men. Endokrynologia Polska 65 (3):203–209. doi: 10.5603/ep.2014.0028 [DOI] [PubMed] [Google Scholar]
- 65.Jura M, Kozak LP (2016) Obesity and related consequences to ageing. Age 38 (1). doi: 10.1007/s11357-016-9884-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Pinto E (2007) Blood pressure and ageing. Postgraduate Medical Journal 83 (976):109–114. doi: 10.1136/pgmj.2006.048371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gambert SR, Pinkstaff S (2006) Emerging Epidemic: Diabetes in Older Adults: Demography, Economic Impact, and Pathophysiology. Diabetes Spectrum 19 (4):221 [Google Scholar]
- 68.Jousilahti P, Vartiainen E, Tuomilehto J, Puska P (1999) Sex, Age, Cardiovascular Risk Factors, and Coronary Heart Disease. Circulation 99 (9): 1165. [DOI] [PubMed] [Google Scholar]
- 69.Silva FH, Alexandre EC, Calmasini FB, Calixto MC, Antunes E (2015) Treatment With Metformin Improves Erectile Dysfunction in a Murine Model of Obesity Associated With Insulin Resistance. Urology 86 (2):423.e421–426. doi: 10.1016/j.urology.2015.04.035 [DOI] [PubMed] [Google Scholar]
- 70.Almundarij TI, Smyers ME, Spriggs A, Heemstra LA, Beltz L, Dyne E, Ridenour C, Novak CM (2016) Physical Activity, Energy Expenditure, and Defense of Body Weight in Melanocortin 4 Receptor-Deficient Male Rats. Scientific reports 6:37435. doi: 10.1038/srep37435 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hill JW (2008) Hypothalamic pathways linking energy balance and reproduction. 294 (5):E827–832. doi: 10.1152/ajpendo.00670.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hill JW, Elias CF (2018) Neuroanatomical Framework of the Metabolic Control of Reproduction. Physiological reviews 98 (4):2349–2380. doi: 10.1152/physrev.00033.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Martin WJ, McGowan E, Cashen DE, Gantert LT, Drisko JE, Hom GJ, Nargund R, Sebhat I, Howard AD, Van der Ploeg LHT, MacIntyre DE (2002) Activation of melanocortin MC4 receptors increases erectile activity in rats ex copula. European journal of pharmacology 454 (1):71–79. doi: 10.1016/S0014-2999(02)02479-2 [DOI] [PubMed] [Google Scholar]
- 74.Wessells H, Levine N, Hadley ME, Dorr R, Hruby V (2000) Melanocortin receptor agonists, penile erection, and sexual motivation: human studies with Melanotan II. International journal of impotence research 12 Suppl 4:S74–79 [DOI] [PubMed] [Google Scholar]
- 75.Muceniece R, Zvejniece L, Kirjanova O, Liepinsh E, Krigere L, Vilskersts R, Baumane L, Gordjusina V, Kalvinsh I, Wikberg JE, Dambrova M (2005) Beta-MSH inhibits brain inflammation via MC(3)/(4) receptors and impaired NF-kappaB signaling. Journal of neuroimmunology 169 (1-2): 13–19. doi: 10.1016/j.jneuroim.2005.07.024 [DOI] [PubMed] [Google Scholar]
- 76.Caruso C, Mohn C, Karara AL, Rettori V, Watanobe H, Schioth HB, Seilicovich A, Lasaga M (2004) Alpha-melanocyte-stimulating hormone through melanocortin-4 receptor inhibits nitric oxide synthase and cyclooxygenase expression in the hypothalamus of male rats. Neuroendocrinology 79 (5):278–286. doi: 10.1159/000079321 [DOI] [PubMed] [Google Scholar]
- 77.Galimberti D, Baron P, Meda L, Prat E, Scarpini E, Delgado R, Catania A, Lipton JM, Scarlato G (1999) Alpha-MSH peptides inhibit production of nitric oxide and tumor necrosis factor-alpha by microglial cells activated with beta-amyloid and interferon gamma. Biochem Biophys Res Commun 263 (1):251–256. doi: 10.1006/bbrc.1999.1276 [DOI] [PubMed] [Google Scholar]
- 78.Mountjoy KG, Guan J, Elia CJ, Sirimanne ES, Williams CE (1999) Melanocortin-4 receptor messenger RNA expression is up-regulated in the non-damaged striatum following unilateral hypoxic-ischaemic brain injury. Neuroscience 89 (1):183–190 [DOI] [PubMed] [Google Scholar]
- 79.Toque HA, da Silva FH, Calixto MC, Lintomen L, Schenka AA, Saad MJ, Zanesco A, Antunes E (2011) High-fat diet associated with obesity induces impairment of mouse corpus cavernosum responses. BJU international 107 (10):1628–1634. doi: 10.1111/j.1464-410X.2010.09704.x [DOI] [PubMed] [Google Scholar]
- 80.Courtois F, Carrier S, Charvier K, Guertin PA, Journel NM (2013) The control of male sexual responses. Current pharmaceutical design 19 (24):4341–4356 [DOI] [PubMed] [Google Scholar]
- 81.Alwaal A, Breyer BN, Lue TF (2015) Normal male sexual function: emphasis on orgasm and ejaculation. Fertility and sterility 104 (5):1051–1060. doi: 10.1016/j.fertnstert.2015.08.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Arletti R, Benelli A, Bertolini A (1992) Oxytocin involvement in male and female sexual behavior. Annals of the New York Academy of Sciences 652:180–193 [DOI] [PubMed] [Google Scholar]
- 83.Clement P, Bernabe J, Denys P, Alexandre L, Giuliano F (2007) Ejaculation induced by i.c.v. injection of the preferential dopamine D(3) receptor agonist 7-hydroxy-2-(di-N-propylamino)tetralin in anesthetized rats. Neuroscience 145 (2):605–610. doi: 10.1016/j.neuroscience.2006.12.003 [DOI] [PubMed] [Google Scholar]
- 84.Holstege G, Georgiadis JR, Paans AM, Meiners LC, van der Graaf FH, Reinders AA (2003) Brain activation during human male ejaculation. The Journal of neuroscience : the official journal of the Society for Neuroscience 23 (27):9185–9193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Georgiadis JR, Reinders AA, Van der Graaf FH, Paans AM, Kortekaas R (2007) Brain activation during human male ejaculation revisited. Neuroreport 18 (6):553–557. doi: 10.1097/WNR.0b013e3280b10bfe [DOI] [PubMed] [Google Scholar]
- 86.Coolen LM, Allard J, Truitt WA, McKenna KE (2004) Central regulation of ejaculation. Physiology & behavior 83 (2):203–215. doi: 10.1016/j.physbeh.2004.08.023 [DOI] [PubMed] [Google Scholar]
- 87.Marson L, Platt KB, McKenna KE (1993) Central nervous system innervation of the penis as revealed by the transneuronal transport of pseudorabies virus. Neuroscience 55 (1):263–280 [DOI] [PubMed] [Google Scholar]
- 88.Dean RC, Lue TF (2005) Physiology of Penile Erection and Pathophysiology of Erectile Dysfunction. The Urologic clinics of North America 32 (4):379–v. doi: 10.1016/j.ucl.2005.08.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Sawchenko PE, Swanson LW (1982) Immunohistochemical identification of neurons in the paraventricular nucleus of the hypothalamus that project to the medulla or to the spinal cord in the rat. The Journal of comparative neurology 205 (3):260–272. doi: 10.1002/cne.902050306 [DOI] [PubMed] [Google Scholar]
- 90.Sawchenko PE, Swanson LW (1982) The organization of noradrenergic pathways from the brainstem to the paraventricular and supraoptic nuclei in the rat. Brain research 257 (3):275–325 [DOI] [PubMed] [Google Scholar]
- 91.Sofroniew MV (1980) Projections from vasopressin, oxytocin, and neurophysin neurons to neural targets in the rat and human. The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 28 (5):475–478. doi: 10.1177/28.5.7381192 [DOI] [PubMed] [Google Scholar]
- 92.Mountjoy KG (2010) Distribution and Function of Melanocortin Receptors within the Brain. Adv Exp Med Biol 681:29–48. doi:Book_Doi 10.1007/978-1-4419-6354-3 [DOI] [PubMed] [Google Scholar]
- 93.Gimpl G, Fahrenholz F (2001) The oxytocin receptor system: structure, function, and regulation. Physiological reviews 81 (2):629–683. doi: 10.1152/physrev.2001.81.2.629 [DOI] [PubMed] [Google Scholar]
- 94.Clement P, Giuliano F (2016) Physiology and Pharmacology of Ejaculation. Basic & Clinical Pharmacology & Toxicology 119:18–25. doi: 10.1111/bcpt.12546 [DOI] [PubMed] [Google Scholar]
- 95.Argiolas A, Melis MR (2005) Central control of penile erection: role of the paraventricular nucleus of the hypothalamus. Progress in neurobiology 76 (1): 1–21. doi: 10.1016/j.pneurobio.2005.06.002 [DOI] [PubMed] [Google Scholar]
- 96.Arletti R, Bazzani C, Castelli M, Bertolini A (1985) Oxytocin improves male copulatory performance in rats. Hormones and behavior 19 (1):14–20 [DOI] [PubMed] [Google Scholar]
- 97.Clement P, Bernabe J, Compagnie S, Alexandre L, McCallum S, Giuliano F (2013) Inhibition of ejaculation by the non-peptide oxytocin receptor antagonist GSK557296: a multilevel site of action. British journal of pharmacology 169 (7):1477–1485. doi: 10.1111/bph.12198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Fjellstrom D, Kihlstrom JE, Melin P (1968) The effect of synthetic oxytocin upon seminal characteristics and sexual behaviour in male rabbits. Journal of reproduction and fertility 17 (1):207–209 [DOI] [PubMed] [Google Scholar]
- 99.de Jong TR, Veening JG, Olivier B, Waldinger MD (2007) Oxytocin involvement in SSRI-induced delayed ejaculation: a review of animal studies. The journal of sexual medicine 4 (1): 14–28. doi: 10.1111/j.1743-6109.2006.00394.x [DOI] [PubMed] [Google Scholar]
- 100.Melis MR, Succu S, Mascia MS, Argiolas A (2005) PD-168077, a selective dopamine D4 receptor agonist, induces penile erection when injected into the paraventricular nucleus of male rats. Neuroscience letters 379 (1):59–62. doi: 10.1016/j.neulet.2004.12.053 [DOI] [PubMed] [Google Scholar]
- 101.Chan JS, Snoeren EM, Cuppen E, Waldinger MD, Olivier B, Oosting RS (2011) The serotonin transporter plays an important role in male sexual behavior: a study in serotonin transporter knockout rats. The journal of sexual medicine 8 (1):97–108. doi: 10.1111/j.1743-6109.2010.01961.x [DOI] [PubMed] [Google Scholar]
- 102.Heisler LK, Jobst EE, Sutton GM, Zhou L, Borok E, Thornton-Jones Z, Liu HY, Zigman JM, Balthasar N, Kishi T, Lee CE, Aschkenasi CJ, Zhang CY, Yu J, Boss O, Mountjoy KG, Clifton PG, Lowell BB, Friedman JM, Horvath T, Butler AA, Elmquist JK, Cowley MA (2006) Serotonin reciprocally regulates melanocortin neurons to modulate food intake. Neuron 51 (2):239–249. doi: 10.1016/j.neuron.2006.06.004 [DOI] [PubMed] [Google Scholar]
- 103.Abdel-Hamid IA, Elsaied MA, Mostafa T (2016) The drug treatment of delayed ejaculation. Translational andrology and urology 5 (4):576–591. doi: 10.21037/tau.2016.05.05 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Walch K, Eder R, Schindler A, Feichtinger W (2001) The effect of single-dose oxytocin application on time to ejaculation and seminal parameters in men. Journal of assisted reproduction and genetics 18 (12):655–659 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Lazzari VM, Becker RO, de Azevedo MS, Morris M, Rigatto K, Almeida S, Lucion AB, Giovenardi M (2013) Oxytocin modulates social interaction but is not essential for sexual behavior in male mice. Behav Brain Res 244:130–136. doi: 10.1016/j.bbr.2013.01.025 [DOI] [PubMed] [Google Scholar]
- 106.Frye CA, Vongher JM (1999) Progesterone has rapid and membrane effects in the facilitation of female mouse sexual behavior. Brain research 815 (2):259–269 [DOI] [PubMed] [Google Scholar]