

Abstract
This study was performed to investigate the effects of dietary inclusion of 20% rapeseed meal (RSM) as an alternative to soybean meal (SBM) in a three-month feeding experiment with growing finishing pigs. Dietary alteration affected growth performance, several carcass traits and transcriptional responses in the skeletal muscle, but did not affect measured meat quality traits. In general, pigs fed the RSM test diet exhibited reduced growth performance compared to pigs on SBM control diet. Significant transcriptional changes in the skeletal muscle of growing pigs fed RSM diet were likely the consequence of an increased amount of fiber and higher polyunsaturated fatty acids, and presence of bioactive phytochemicals, such as glucosinolates. RNAseq pipeline using Tophat2-Cuffdiff identified 57 upregulated and 63 downregulated genes in RSM compared to SBM pigs. Significantly enriched among downregulated pathways was p53–mediated signalling involved in cellular proliferation, while activation of negative growth regulators (IER5, KLF10, BTG2, KLF11, RETREG1, PRUNE2) in RSM fed pigs provided further evidence for reduced proliferation and increased cellular death, in accordance with the observed reduction in performance traits. Upregulation of well-known metabolic controllers (PDK4, UCP3, ESRRG and ESRRB), involved in energy homeostasis (glucose and lipid metabolism, and mitochondrial function), suggested less available energy and nutrients in RSM pigs. Furthermore, several genes supported more pronounced proteolysis (ABTB1, OTUD1, PADI2, SPP1) and reduced protein synthesis (THBS1, HSF4, AP1S2) in RSM muscle tissue. In parallel, higher levels of NR4A3, PDK4 and FGF21, and a drop in adropin, ELOVL6 and CIDEC/FSP27 indicated increased lipolysis and fatty acid oxidation, reflective of lower dressing percentage. Finally, pigs exposed to RSM showed greater expression level of genes responsive to oxidative stress, indicated by upregulation of GPX1, GPX2, and TXNIP.
Introduction
European pig production, especially in the Nordic countries, is to a large extent based on imported protein feedstuffs such as soybean meal (SBM). To improve sustainability and self-sufficiency of pig production, alternative indigenous protein feedstuffs are needed. Rapeseed is a viable alternative crop that can be grown in cold climates, and its cultivation has increased during recent years due to a growing demand for oil for biofuel production [1]. Rapeseed meal (RSM) is a co-product in oil and biofuel production, it is abundant and cheap and can serve as an alternative to SBM in diets for monogastric animals. Rapeseed meal has been used in animal diets over an extended time period, but due to a high fiber content, and anti-nutritional factors such as glucosinolates, sinapine, tannins, and erucic acid, the inclusion of RSM in pig diets usually has been kept low [2, 3]. Nevertheless, beneficial effects of RSM dietary fiber on intestinal health and overall well-being of pigs have been demonstrated [4–6].
When aiming at increasing the sustainability of pig production, feed efficiency (FE) is an essential factor both for reducing feed costs and environmental impact. Modern pig genotypes have mostly been selected for growth performance and FE under intensive feeding conditions, and Noblet et al. [7] showed that within a pig line, there was individual variation in the ability to digest organic matter, nitrogen and energy when growing finishing pigs were fed high fiber diets. The Norwegian Landrace pig used in this study is an example of such fast growing, highly efficient breed [8]. Yet, identifying the best genotypes that perform well when fed high fiber diets will be of considerable value for further improvements in sustainable pig production. For better utilization of local fiber-rich feedstuffs, more knowledge on the molecular mechanisms causing growth performance differences under those dietary regimes is required. A recent study showed that Norwegian Landrace piglets fed high dietary levels of RSM perform quite differently due to large individual variations in utilization of crude protein (CP) and neutral detergent fiber (NDF), supporting the existence of superior genotypes within the same breed that are better at digesting CP and NDF [9].
Genetic background has a large influence on animal performance, therefore, a synergy between genetics and nutrition is needed to facilitate our understanding of what constitutes optimal responses to fiber-rich diets. Only recently, it became possible to combine high-throughput omics technologies with nutrition to better understand complex biology behind economically important traits. Novel network-based approaches are being used to discover and expand gene networks associated with particular phenotypes. Recently, Vincent et al. [10] linked differences in performance between pigs selected for improved efficiency and gene expression variations in muscle tissue related to protein synthesis and mitochondrial energy metabolism. Similarly, Jing et al. [11] performed transcriptome analysis of mRNA and miRNA in skeletal muscle of Yorkshire boars and found association between gene networks controlling mitochondrial energy metabolism and muscle growth and different FE phenotypes. Another recent work compared muscle transcriptome of different pig genotypes and identified genes and regulatory networks associated with growth, fatness and metabolism [12].
The present study was conducted to evaluate the influence of two different diets: a typical commercial-like diet containing imported SBM as a control, and a locally produced diet with 20% inclusion of RSM, on animal performance, carcass and meat quality traits. The second objective was to characterize diet effects on gene expression profiles in skeletal muscle (Longissimus dorsi) and to identify molecular players with the largest impact on observed phenotypic differences. Hence, we employed an integrated approach combining RNAseq technology and quantitative real-time PCR (qRT-PCR) to investigate effects of a dietary alteration on global transcriptome changes in the muscle.
Materials and methods
Ethics statement
The experiment was conducted at the Center for Animal Research, Norwegian University of Life Sciences, Aas, Norway and the experimental protocol for the study was approved by the Norwegian Food Safety Authority (ID: 8217). All animals were cared for according to laws and regulations controlling experiments with live animals in Norway (the Animal Protection Act of December 20th, 1974, and the Animal Protection Ordinance concerning experiments with animals of January 15th, 1996).
Animals, allotment and housing
A total of 84 purebred Norwegian Landrace pigs from 16 litters were used. Average initial weight was 24.9 kg ± 1.98 standard deviation (SD) and average final weight was 109.7 kg ± 5.44 SD. The experiment was conducted as a randomized complete block design. Pigs were blocked by litter and sex, and allotted by initial weight to two dietary treatments with 42 replicates per treatment with equal number of males and females. The experimental period lasted on average 88 days. At feeding time, each pig was restrained in an individual feeding stall until the feed was consumed in order to obtain individual feed intake. Thus, each pig was the experimental unit. The experiment was split into a growing period from start until 60 kg live weight, a finishing period from 60 kg live weight until slaughter, and the overall period. Pigs were housed in an environmentally controlled barn with partially slotted concrete floor. Fourteen 8.2 m2 pens designed for individual feeding were used. Due to the experimental goals, no bedding material was used, the pens were cleaned regularly, and they were equipped with rubber mats and activity enrichment tools. Average ambient daily temperature was 18.2°C (range 15.9–20.5°C). The clinical health status of the pigs was monitored daily during the feeding time. Overall, all animals were healthy throughout the experimental period and no mortalities were recorded.
Diets and feeding
The dietary treatments were: 1) a control diet based on barley, wheat, oat and soybean meal (SBM-diet), and 2) an experimental diet based on barley, wheat, oat and 20% inclusion of commercial expeller pressed rapeseed (Mestilla UAB, Klaipeda, Lithuania) (RSM-diet). Levels of standardized ileal digestible (SID) lysine on net energy (NE) basis was adjusted to be the same in both diets by using crystalline amino acids. The diets were also designed to be isonitrogenous and isoenergetic and to contain equal levels of methionine + cysteine, and threonine. The diets were pelleted with a 3-mm die and produced by the Center for Feed Technology (FôrTek) at the Norwegian University of Life Sciences. The content of digestible lysine, threonine, methionine and cystine of the ingredients was estimated using analysed values, multiplied by the standardized ileal digestibility coefficients for nitrogen and amino acids [13]. Diets were formulated to meet or exceed the requirements for indispensable amino acids and all other nutrients [14]. A cumulative feed sample from each dietary treatment was taken for chemical analysis. Composition and analyses of diets are shown in Table 1.
Table 1. Ingredient and chemical composition (g kg-1, unless otherwise stated) of diets based on soybean meal (SBM) and rapeseed meal (RSM).
| Diet | ||
|---|---|---|
| Ingredients | SBM | RSM |
| Barley | 380.2 | 340.4 |
| Wheat | 240.0 | 233.4 |
| Oats | 140.0 | 140.0 |
| Soybean meal (SBM) (45% CP) | 150.0 | 0.0 |
| Rapeseed meal (RSM) | 0.0 | 200.0 |
| Rendered fat (tallow) | 50.4 | 50.0 |
| Limestone | 11.3 | 8.0 |
| Monocalcium phosphate | 16.4 | 16.4 |
| Salt | 4.0 | 4.0 |
| L-lysine. HCl (98%) | 2.9 | 3.8 |
| Threonine | 1.5 | 1.5 |
| DL-methionine | 0.9 | 0.0 |
| Tryptophan | 0.1 | 0.2 |
| Premix1 | 2.2 | 2.2 |
| Yttrium oxide | 0.1 | 0.1 |
| Calculated contents, g kg-1 | ||
| Net energy, MJ kg-1 | 9.3 | 9.2 |
| SID2 lysine | 8.2 | 8.2 |
| SID methionine + cysteine | 4.9 | 5.0 |
| SID threonine | 5.4 | 5.4 |
| SID tryptophan | 1.6 | 1.6 |
| Calcium | 8.7 | 8.5 |
| ATTD3 phosphorus | 3.9 | 4.3 |
| Analysed content, g kg-1 | ||
| DM | 907.0 | 881.0 |
| Gross energy, MJ kg-1 | 17.6 | 16.6 |
| Crude protein | 147.0 | 142.0 |
| Crude fat | 63.0 | 43.0 |
| Ash | 54.0 | 50.0 |
| Neutral detergent fiber (NDF) | 144.0 | 159.0 |
| Acid detergent fiber (ADF) | 43.0 | 61.0 |
| Calcium | 9.5 | 9.6 |
| Total phosphorus | 8.3 | 8.5 |
| Lysine | 9.0 | 8.8 |
| Threonine | 7.2 | 7.3 |
| Methionine | 2.7 | 2.4 |
| Cysteine | 2.6 | 2.8 |
| Tryptophan Total glucosinolates, mmol kg-1 |
2.0 - |
1.9 2.3 |
1Provided per kilogram of feed: 105 mg of Zn (ZnO); 75 mg of Fe (FeSO4. H20); 60 mg of Mn (MnO); 15 mg of Cu (CuSO4 x 5H2O); 0.75 mg of I (Ca(IO3)2; 0.3 mg of Se (Na2SeO3); 9000 IU of vitamin A; 1125 IU of cholecalciferol; 112.5 mg of dl-a-tocopheryl acetate; 2.25 mg of menadione; 5.625 mg of riboflavin, 18.73 mg of d-pantothenic acid; 22.5 mg of cyanocobalamine; 22.5 mg of niacin; 0.225 mg of biotin; 1.69 mg of folic acid; 364 mg of choline.
2 Standardized ileal digestible
3 Apparent total tract digestible.
All pigs were individually fed twice per day according to a semi-ad libitum feeding regime providing a moderate feeding intensity during the growing period followed by an increasing feeding intensity during the finishing period [15]. Feed refusals for each pig were recorded and subtracted from the total feed intake. All pigs were given free access to water from nipple drinkers. Water was also provided directly in the trough during meals. Feed consumption and pigs’ weight were recorded weekly to determine average daily gain (ADG), average daily feed intake (ADFI), and feed conversion ratio (FCR).
Slaughter procedure and tissue sampling
Pigs were slaughtered at the Center for Animal Research using a mobile slaughterhouse (Mobilslakt AS) under the supervision of the Norwegian Food Safety Authority. Pigs were fixed and anesthetized using head-to-heart electrical immobilization followed by exsanguination. Samples of the muscle (L. dorsi), liver, ileum and colon tissues were snap frozen in liquid nitrogen for gene expression analyses.
Chemical analyses
Samples of dietary ingredients and the two diets were analysed for CP by Kjeldahl-N x 6.25 [16], fat using ASE 350 Accelerated Solvent Extractor, dry matter (DM) by drying to constant weight at 104°C [16], ash by incineration at 550°C [16], acid detergent fiber (ADF) and NDF using a fiber analyzer system (Ankom200, ANKOM Technologies, Fairport, NY, USA) with filter bags (Ankom F58, ANKOM Technologies). Gross energy (GE) content was determined by a Parr 1281 Adiabatic Bomb Calorimeter (Parr Instruments, Moline, IL, USA) according to [17]. Phosphorus and calcium content of the diets were analysed by atomic absorption spectrophotometry according to methods described by [18]. Analysis of amino acids in the diets was carried out according to [16] using Biochrom 30 Amino Acid Analyzer. Tryptophan in the diets was determined according to [16] using high-performance liquid chromatography system (Dionex UltiMate 3000, Dionex Softron GmbH, Germering, Germany) equipped with fluorescent detector (Shimadzu RF- 535, Shimadzu Corp, Kyoto, Japan). Glucosinolate level in the experimental RSM-diet was determined by Eurofins Food & Feed Testing Norway AS (Kambo, Moss, Norway) using liquid chromatography coupled with diode array detector- method (LC-DAD).
Carcass characteristics
Dressing percentage was determined by the following equation: (hot carcass weight/final live weight) x 100. Live weight was monitored at the time of slaughter. Carcass lean percentage was determined commercially on the slaughter line using a GP7Q pistol (Hennessy System Ltd, Auckland, New Zealand) to measure the depth of the L. dorsi and the backfat thickness at two sites (behind the last rib, 6 cm from the midline) [15]. The prediction of carcass lean percentage was done according to an equation from the Norwegian Meat Research Centre, Oslo, Norway.
Forty-one pigs (20 from SBM and 21 from RSM group) were selected for carcass and meat quality assessment at Animalia–The Norwegian Meat and Poultry Research Centre. Backfat thickness and backfat+loin thickness including loin and fat were measured behind the last rib, 6 cm from the midline. Total fat was measured according to the method described by O’Farell et al. [19]. The carcasses were subjected to commercial cutting procedure to determine primal cut yields (backpart, midpart and frontpart), and to measure percent carcass lean and percent carcass fat. In addition, muscle pH and meat colour measures were determined from the L. dorsi muscle. Meat colour was measured after one hour blooming at 4°C using a Minolta CR400 (Minolta Co. Ltd., Osaka, Japan). Three measurements were taken from each muscle, and mean values for CIELAB L*, a*, b*, Hue and Chroma (saturation) were calculated. Values for Hue was calculated as Hue = tan-1 (b*/a*), and Chroma as Croma = “squareroot”(A*2 +b*2). Drip loss was measured in the L. dorsi and the loin, according to the EZ-driploss metod [20]. Collagen, fat, protein and water content in the L. dorsi was analysed using a FOSS FoodScan [21].
RNA extraction
Total RNA from muscle, liver, ileum and colon was extracted from 6 SBM and 6 RSM pigs using TRIzol protocol (Invitrogen) followed by RNeasy Plus Mini protocol (Qiagen). After the first washing step, on-column DNAse treatment was performed using PureLink DNase kit (Invitrogen). RNA purity and quality was measured using NanoDrop 8000 spectrophotometer (Thermo Fisher Scientific, Wilmington, USA) and Agilent 2100 Bioanalyzer (Agilent Technologies, Waldbronn, Germany). Only high quality (RIN ≥ 7) samples were sent for sequencing at the Norwegian Sequencing Centre (http://www.sequencing.uio.no) and were used for validation of sequencing data by qRT-PCR. Six pigs from each dietary treatment were selected for RNA sequencing based on their FCR (three pigs with FCR higher than average and three pigs with FCR lower than average), while aiming at equal gender distribution. Pigs that were selected for sequencing originated from different litters in order to minimize the effect of single genotypes on the outcome of this study.
Library construction and RNA sequencing
In total 48 RNA sequencing libraries were prepared with TruSeq stranded mRNA prep (Illumina, San Diego, USA) using unique barcodes and 150 bp paired end sequencing was performed across 4 lanes in HiSeq 4000 System (Illumina, San Diego, USA). Of those 48 libraries, 12 libraries contained muscle samples and the remaining 36 consisted of liver, ileum and colon samples, which will be analysed and presented in another study (data not shown). Profiling of transcriptomic responses from liver and gut tissues will provide additional understanding of effects of RSM-based diets on development and function of those organs in growing pigs. In this study, only data from 12 muscle samples was analysed and presented. Sequence data was handled using RTA v 2.7.6 and demultiplexed and converted into fastq files employing the unique barcodes using bcl2fastq v2.18.0.12.
Raw data from four lanes for each sample were concatenated together before proceeding with the analysis. Sequenced data were pre-processed by trimming/removing reads containing adapter sequences, low quality reads and PhiX genome (Illumina spike-in) using BBDuK v34.56 part of BBtools software package [22].
Data pre-processing and differential expression analysis
Cleaned data was aligned against the Sus scrofa11.1 transcriptome and genome (Genbank assembly accession GCA_000003025.6) using tophat v2 [23] using—library-type fr-firststrand—no-mixed—no-novel-junc as options. Average insert fragment size for each sample was calculated by aligning 1 million cleaned paired end reads against the Sus scrofa cDNA using bowtie2 V2.2.3 [24] and Picard v1.112 CollectInsertSizeMetrics tool and were provided as parameters for tophat2 alignment. Differential expression of known genes (ensembl release 95) was calculated using the tophat2 aligned bam files in cuffdiff v2.2.1 using default parameters and visualised using cummeRbund v2.18.0 package [25] in R.
cDNA synthesis and validation by qRT-PCR
To validate RNAseq data, the expression levels of activating transcription factor 3 (ATF3), thioredoxin-interacting protein (TXNIP), mitochondrial uncoupling protein 3 (UCP3), parvalbumin (PVALB), and myostatin (MSTN) were measured by qRT-PCR. Primers were designed using the PRIMER3 program (http://frodo.wi.mit.edu) and their full list is given in S1 Table. Prior to cDNA synthesis all samples were normalized to 300 ng/μL. cDNA synthesis was performed using AffinityScript QPCR cDNA Synthesis kit (Agilent Technologies). The qRT- PCR was performed in a total volume of 20 μL using 10 μL LightCycler 480 SYBR Green I Master, 2 μL primers, 3 μL Milli-Q water and 5 μL cDNA diluted 1:50. The PCR conditions were: 95°C for 10 min, 95°C for 10 sec, 60–64°C for 10 sec depending on the primers, 72°C for 10 sec, in a total of 40 cycles. Samples were analysed using LightCycler 480 System (Roche Diagnostics, Mannheim, Germany). All reactions were performed in duplicates and gene expression levels were quantified relative to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and β-actin (ACTB) using a mean –ΔΔCt value.
Statistical and functional annotation analyses
Statistical analyses on animal performance and carcass characteristics were performed using the GLM procedure of SAS [26] for a complete randomized block design with individual pig as the experimental unit. Results are presented as the least square mean for each treatment, and variance is expressed as standard error of the mean (SEM). Means were separated according to the LSmeans procedure. Significant difference among treatments was shown at p<0.05.
Heatmap showing the log2(fpkm +1) values of differentially expressed genes (DEGs) was constructed based on Euclidean correlation and average linkage clustering in software Multi Experimental Viewer (MeV) (http://www.mev.tm4.org). For the functional interpretation of transcriptomic data, the identified pig DEGs were converted to human orthologs using the g:Orth tool in the g:Profiler web server [27] and then used for Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG) [28] and Reactome [29] enrichment analyses. To distinguish significant results from random matches, the Benjamini-Hochberg FDR method was chosen and p< = 0.05 was set as a cutoff.
Results
Growth performance
During the growing (p = 0.119), finishing (p = 0.002) and overall (p = 0.004) period, ADFI was lower in pigs fed the RSM diet than those fed the SBM diet. Pigs fed the RSM diet also had a lower ADG during the growing (p = 0.002), finishing (p = 0.002) and overall period (p = 0.001). The FCR was poorer in pigs fed the RSM diet in the growing (p = 0.007) and in the overall period (p = 0.012), whereas there were no differences in FCR between groups in the finishing period (p = 0.223). Similarly, the NE intake per kg gain was higher in RSM pigs during the growing and overall periods, while there was no difference between treatment groups during the finishing period (Table 2).
Table 2. Growth performance and carcass traits of growing-finishing pigs fed diets based on soybean meal (SBM) or rapeseed meal (RSM).
| Diet | SEM | p-value; effect of diet | ||
|---|---|---|---|---|
| SBM | RSM | |||
| Number of pigs | 42 | 42 | ||
| Live weight, kg | ||||
| Initial | 29.2 | 28.8 | 0.19 | 0.139 |
| End of growing period | 64.4 | 62.2 | 0.46 | 0.002 |
| At slaughter | 112.2 | 107.5 | 0.66 | 0.001 |
| Days in experiment | 88.1 | 88.1 | 0.45 | 0.777 |
| Growing period | ||||
| ADG1, g | 837.6 | 795.2 | 9.16 | 0.002 |
| ADFI2, g | 1707.0 | 1678.0 | 13.00 | 0.119 |
| FCR3, g feed/g gain | 2.0 | 2.1 | 0.02 | 0.007 |
| NE:gain, MJ/kg gain | 19.0 | 19.5 | 0.17 | 0.017 |
| Finishing period | ||||
| ADG, g | 1039.4 | 986.9 | 11.04 | 0.002 |
| ADFI, g | 2557.0 | 2470.0 | 17.70 | 0.002 |
| FCR, g feed/g gain | 2.5 | 2.5 | 0.02 | 0.223 |
| NE:gain, MJ/kg gain | 22.9 | 23.2 | 0.21 | 0.390 |
| Overall period | ||||
| ADG, g | 942.9 | 895.0 | 7.69 | 0.001 |
| ADFI, g | 2151.0 | 2088.0 | 13.40 | 0.004 |
| FCR, g feed/g gain | 2.29 | 2.34 | 0.01 | 0.012 |
| NE:gain, MJ/kg gain | 21.2 | 21.6 | 0.12 | 0.047 |
| Carcass traits4 | ||||
| Carcass weight, kg | 78.7 | 73.8 | 0.51 | 0.001 |
| Dressing percentage | 70.1 | 68.7 | 0.26 | 0.001 |
| Carcass lean percentage | 61.9 | 62.1 | 0.25 | 0.845 |
| Kg feed: kg carcass | 2.4 | 2.5 | 0.01 | 0.001 |
1Average daily gain
2Average daily feed intake
3Feed conversion ratio
4 SBM diet: N = 41 and RSM diet: N = 42
Carcass and meat quality traits
Carcasses of RSM pigs were lighter than those of SBM pigs (p = 0.001), dressing percentage was lower (p = 0.001), but carcass lean percentage was similar among dietary treatments (p = 0.845). Feed intake per kg carcass was higher in RSM than SBM pigs (p = 0.001) (Table 2). The carcass quality traits reported in Table 3 were generally very similar among dietary treatments. However, the backfat+loin thickness was significantly (p = <0.031) smaller in RSM than SBM carcasses. There were also tendencies for differences in the conformation of the carcasses with the backpart making up similar amounts in the two groups while the midpart was slightly smaller (p = 0.097) and the frontpart slightly larger (p = 0.059) in the RSM carcasses.
Table 3. Carcass and meat quality traits of pigs fed diets based on either soybean meal (SBM) or rapeseed meal (RSM).
| Diet | SEM | p-value; effect of diet | ||
|---|---|---|---|---|
| SBM | RSM | |||
| Number of pigs1 | 20 | 21 | ||
| Carcass weight, kg | 76.6 | 74.8 | 1.01 | 0.199 |
| Carcass lean percent, GP7 | 62.3 | 62.0 | 0.73 | 0.780 |
| Backfat (mm) 2 | 13.8 | 12.7 | 0.58 | 0.188 |
| Backfat+loin thickness (mm) 2 | 54.1 | 50.1 | 1.24 | 0.031 |
| Loin (LD), fat (mm) | 10.0 | 9.2 | 0.85 | 0.493 |
| Total carcass fat (%)3 | 18.0 | 17.7 | 0.93 | 0.847 |
| Primal cuts method | ||||
| Percent carcass lean | 63.0 | 63.0 | 0.73 | 0.968 |
| Percent carcass fat | 19.5 | 19.5 | 0.93 | 0.951 |
| Backpart, % | 34.9 | 34.8 | 0.30 | 0.899 |
| Midpart, % | 35.9 | 35.3 | 0.25 | 0.097 |
| Frontpart, % | 29.2 | 29.9 | 0.24 | 0.059 |
| Drip loss, loin, % | 5.4 | 5.2 | 0.41 | 0.702 |
| Drip loss, belly, % | 4.7 | 4.7 | 0.54 | 0.948 |
| pH, LD-muscle | 5.5 | 5.4 | 0.02 | 0.473 |
| Collagen, Foodscan (%) | 1.0 | 1.1 | 0.03 | 0.039 |
| Fat, Foodscan (%) | 1.2 | 1.2 | 0.05 | 0.606 |
| Protein, Foodscan (%) | 23.3 | 23.3 | 0.14 | 0.967 |
| Water, Foodscan (%) | 74.4 | 74.3 | 0.11 | 0.604 |
| Minolta Chrome | 9.1 | 9.8 | 0.35 | 0.170 |
| Minolta Hue | 20.6 | 21.8 | 1.17 | 0.424 |
Meat quality traits were measured on 20 SBM and 21 RSM carcasses, and there were generally no differences between treatment groups, the only significant difference being a slightly higher content of collagen in RSM carcasses (p = 0.039) (Table 3).
Transcriptomic analysis
RNA sequencing was performed to identify changes in muscle gene expression profiles of 12 Norwegian Landrace pigs belonging to the two groups (SBM and RSM). On average sequencing resulted in 30 M 150bp paired end reads per sample, of which more than 98% were retained after pre-processing for alignment and further analyses (S2 Table). Up to 71% of these cleaned reads aligned to the reference genome/transcriptome using the tophat2 pipeline and nearly 99% of these were of concordant alignment. The sequence reads have been submitted to the NCBI Sequence Read Archive (SRA) under accession number PRJNA506343.
The expression of 25,878 genes was tested in the muscle tissue of pigs subjected to RNAseq. The criteria for DEGs was selected by comparing the test RSM group to the control SBM, with the log2-fold change = >0.5; p< = 0.05 and q< = 0.05 adjusted for FDR, which resulted in 57 upregulated and 63 downregulated DEGs. Previous studies investigating muscle gene expression patterns in pigs reported that alterations in expression profiles could be small [30] [31], hence, in this study, all transcripts with p< = 0.05 and q< = 0.05 were considered DEGs. Volcano plot for the DEGs is shown in S1 Fig. Heat map of gene expression profiles in control SBM and experimental RSM group is shown in Fig 1 and the complete info of all identified DEGs is shown in S3 Table.
Fig 1. Heat map of gene expression profiles in control SBM and experimental RSM group.

Heatmap is showing the log2(fpkm +1) values. The genes are clustered using Euclidean correlation and average linkage clustering in MeV.
Due to the incomplete annotation of the reference pig genome (Sus scrofa11.1), the DEGs were converted to human orthologs and the web server g:Profiler was used for functional analyses of enriched biological processes and pathways. The analysis of 49 upregulated genes that entered GO analysis revealed 21 enriched biological processes, most of which were related to lipid and carbohydrate metabolic process, skeletal muscle tissue development and function, response to nutrient levels and energy homeostasis and oxidative stress. Reactome identified nuclear receptor transcription and RET signaling pathways as enriched among the upregulated genes. A pie chart with an overview of enriched biological processes represented by at least four or more upregulated genes is shown in Fig 2. The group of 63 downregulated genes, of which 60 were used for analysis, were enriched for 2 biological processes, skeletal muscle adaptation and cell aggregation. Enrichment analysis of biological pathways among downregulated genes revealed two enriched pathways, p53 signaling pathway and diseases of glycosylation (pie chart not shown). The complete list of enriched GO processes and KEGG and Reactome pathways with corresponding p-values is provided in S4 Table.
Fig 2. Gene Ontology analysis showing significantly enriched biological processes among upregulated DEGs in the muscle of RSM pigs in comparison to SBM pigs.

Biological processes represented by less than 4 genes were excluded.
Differentially expressed genes between RSM and SBM fed pigs
A number of DEGs orchestrating muscle metabolism and development were regulated: a growth factor FGF21 that functions as a key metabolic regulator induced by different kinds of stress; FOS encoding leucine zipper protein implicated in skeletal muscle signal transduction, cell proliferation and differentiation; LHX6 developmental protein implicated in differentiation, neurogenesis and transcription regulation; IFRD1/TIS7 controlling growth, differentiation and muscle tissue regeneration; ESRRG and ESRRB expressed in tissues with high metabolic demand like skeletal muscle where they play important roles in fiber-type determination, muscle vascularization, oxidative and lipid metabolism; FGF1 implicated in myogenesis and muscle regeneration and negative regulator of skeletal muscle growth MSTN. In addition, dietary treatment affected the levels of transcripts mediating calcium signaling and muscle contraction, with STC1 and RRAD showing increased expression, while PVALB and STAC exhibited lower mRNA levels (Table 4).
Table 4. Differentially expressed genes in the muscle of the RSM pigs in comparison to the SBM pigs.
Data are log2-fold change (FC).
| Muscle metabolism and development | Short gene name | log2-FC | Functional description | References |
|---|---|---|---|---|
| Fibroblast growth factor 21 | FGF21 | 3.52 | energy homeostasis, mitochondrial function, oxidative stress | [32] |
| Fos proto-oncogene | FOS | 1.70 | muscle metabolism, signaling, apoptosis | [33] |
| LIM homeobox 6 | LHX6 | 1.20 | transcription regulator, differentiation, neurogenesis, development | [34] |
| Estrogen related receptor gamma | ESRRG | 0.91 | transcription regulator, muscle development, oxidative metabolism | [35] [36] |
| Estrogen related receptor beta | ESRRB | 0.72 | transcription regulator, muscle development, oxidative metabolism | [37] |
| Fibroblast growth factor 1 | FGF1 | 0.60 | growth factor, muscle development | [38] |
| Interferon related developmental regulator 1 | IFRD1/TIS7 | 0.57 | muscle cell differentiation, skeletal muscle regeneration | [39] |
| Myostatin | MSTN | -0.84 | regulator of myogenesis, muscle weight | [40] |
| Calcium and potassium signaling | ||||
| Stanniocalcin 1 | STC1 | 0.89 | calcium ion homeostasis | [41] |
| Ras related glycolysis inhibitor and calcium channel regulator | RRAD | 0.63 | calcium and insulin signaling, muscle fiber type determination | [42] |
| SH3 and cysteine rich domain | STAC | -1.14 | muscle contraction, ion transport | [43] |
| Parvalbumin | PVALB | -0.98 | muscle protein, calcium ion binding, contraction | [33] |
| Negative regulation of proliferation, apoptosis, stress response | ||||
| Ankyrin repeat and BTB domain containing 1 | ABTB1/BPOZ | 2.59 | growth-suppressive signaling, proteolysis | [44] |
| Activating transcription factor 3 | ATF3 | 1.54 | transcription regulator, stress response | [45] [46] |
| Immediate early response 5 | IER5 | 1.20 | transcription regulator, growth-suppressive signaling | [47] |
| BTG anti-proliferation factor 2 | BTG2 | 1.16 | growth-suppressive signaling | [48] |
| Kruppel like factor 10 | KLF10/TIEG1 | 1.06 | proliferation repressor, transcription repressor, muscle weight | [49] [50] |
| Reticulophagy regulator 1 | RETREG1 | 0.95 | autophagy, ER-phagy, apoptosis | [51] |
| Protein prune homolog 2 | PRUNE2 | 0.84 | pro-apoptotic | [52] |
| Kruppel like factor 11 | KLF11/TIEG3 | 0.72 | anti-proliferative, apoptosis | [53] |
| Asparagine synthetase (glutamine-hydrolyzing) | ASNS | -1.03 | cell cycle, cell growth | [54] |
| Centrosomal protein 135 | CEP135 | -0.78 | centriole formation, cell cycle | [55] |
| Insulin like growth factor binding protein 5 | IGFBP5 | -0.68 | myogenesis activator, growth | [56] |
| Nuclear protein 1, transcriptional regulator | NUPR1 | -0.68 | stress response, negative regulation of cell cycle | [57] |
| Structural maintenance of chromosomes 5 | SMC5 | -0.63 | cell cycle, cell division, DNA damage, stress response | [58] [59] |
| Positive regulation of proliferation | ||||
| Histamine receptor H1 | HRH1 | 1.09 | proliferation, angiogenesis, development, growth | [60] [61] |
| Glial cell derived neurotrophic factor | GDNF | 0.94 | growth factor, positive regulation of proliferation | [62] |
| ROS proto-oncogene 1. receptor tyrosine kinase | ROS1 | 0.94 | differentiation, proliferation, vascular remodeling, stress response | [63] |
| Zinc finger MIZ-type containing 1 | ZMIZ1 | 0.71 | artery development, vasculogenesis, fibroblast proliferation | [64] [65] |
| GRB2 associated binding protein 2 | GAB2 | 0.57 | signalling, positive regulation of cell proliferation, neurogenesis | [66] |
| Phorbol-12-myristate-13-acetate-induced protein 1 | PMAIP1/NOXA | -0.88 | DNA damage, apoptosis, response to oxidative stress | [67] |
| Base methyltransferase of 25S rRNA 2 homolog | SAMTOR | -0.68 | mTORC1 signaling, cell growth | [68] |
| HAUS augmin like complex subunit 6 | HAUS6 | -0.66 | cell cycle, cell division | [69] |
| Protein metabolism/protein turnover | ||||
| OTU deubiquitinase 1 | OTUD1 | 1.69 | proteolysis, protein transport | [70] |
| Phenylalanine—tRNA ligase beta subunit-like | FARSB | 1.06 | protein biosynthesis | |
| Peptidyl arginine deiminase 2 | PADI2 | 0.73 | citrullination of vimentin, degradation | [71] |
| Adaptor related protein complex 1 subunit sigma 2 | AP1S2 | -1.34 | protein transport | [72] |
| Heat shock factor protein 4 | HSF4 | -0.90 | unfolded protein response, stress response | [73] |
| Fragile X mental retardation 1 | FMR1 | -0.71 | translation repressor, proteasomal protein catabolism | [74] |
| WD repeat and SOCS box containing 1 | WSB1/SWIP1 | -0.63 | protein ubiquitination, protein modification | [75] |
| Calpain 5 | CAPN5 | -0.61 | proteolysis, neuronal differentiation, response to stress | [76] |
| Carbohydrate, lipid and energy metabolism | ||||
| Nuclear receptor subfamily 4 group A member 3 | NR4A3 | 2.03 | metabolic function, energy homeostasis, oxidative metabolism | [77] [78] |
| Alkaline ceramidase 1 | ACER1 | 1.18 | ceramide degradation, sphingosine-1-phosphate signaling | [79] |
| Pyruvate dehydrogenase kinase 4 | PDK4 | 1.11 | glucose and fatty acid metabolism | [80] [81] |
| Insulin receptor substrate 2 | IRS2 | 0.76 | insulin signaling in skeletal muscle | [82] |
| 1.4-alpha-glucan branching enzyme 1 | GBE1 | 0.61 | glycogen metabolism | [83] |
| ST8 alpha-N-acetyl-neuraminide alpha-2.8-sialyltransferase 5 | ST8SIA5 | 0.57 | sphingolipid biosynthesis, protein glycosylation | [84] |
| Retinol dehydrogenase 16 (all-trans) | RDH16 | -0.84 | electron transport, vitamin A metabolism, lipid metabolism | [85] |
| Energy homeostasis associated | ENHO/adropin | -0.79 | glucose homeostasis, lipid and energy metabolism | [86] [87] |
| Adiponectin | ADIPOQ | -0.75 | adipokine, β-oxidation | [88] |
| Cell death inducing DFFA like effector c | CIDEC/FSP27 | -0.68 | lipid droplet organization, lypolysis | [89] |
| ELOVL fatty acid elongase 6 | ELOVL6 | -0.63 | fatty acid elongation, lipogenesis | [90] |
| Biotransformation/xenobiotic metabolism | ||||
| Glutathione peroxidase 1 | GPX1 | 1.30 | antioxidant enzyme | [91] [92] |
| Glutathione peroxidase 2 | GPX2 | 0.93 | antioxidant enzyme | [91] [93] |
| Uncoupling protein 3 | UCP3 | 0.78 | response to ROS, mitochondrial FA transport | [94] |
| Thioredoxin interacting protein | TXNIP | 0.62 | response to ROS, energy metabolism | [95] [96] |
| Endothelial PAS domain protein 1 | EPAS1 | 0.62 | oxygen sensor, response to hypoxia, proliferation, angiogenesis | [97] |
| ATP-binding cassette sub-family A member 8 | ABCA8 | -0.88 | detoxification, transport | [98] |
| Thrombospondin 1 | THBS1/TSP1 | -0.62 | cell adhesion, angiogenesis, oxidative stress, unfolded protein response | [99] |
| Sestrin 3 | SESN3 | -0.55 | metabolic regulator, response to oxidative stress | [100] |
| Extra cellular matrix and cell adhesion | ||||
| Secreted phosphoprotein 1 | SPP1/OPN | 1.79 | cytokine, signaling, ECM remodelling; muscular dystrophy | [101] [102] [103] |
| Thrombospondin type 1 domain containing 4 | THSD4 | 0.77 | matrix assembly | [104] |
| Otoraplin | OTOR | -1.27 | connective tissue development | [105] |
| Carcinoembryonic antigen related cell adhesion molecule 16 | CEACAM16 | -1.20 | cell adhesion, cell growth, angiogenesis | [106] |
| Tenomodulin | TNMD | -1.13 | angiogenesis inhibitor, marker of tendons and ligaments | [107] |
| ADAM metallopeptidase with thrombospondin type 1 motif 6 | ADAMTS6 | -1.05 | ECM proteolysis, biosynthesis of collagen | [108] |
| Collagen type XI alpha 1 chain | COL11A1 | -0.99 | connective tissue, ECM | [109] |
| Fibromodulin | FMOD | -0.81 | ECM, collagen fibril organization | [110] |
| Osteoglycin | OGN | -0.78 | ECM, collagen fibrillogenesis, growth factor | [111] |
| Cytoskeleton and cell motility | ||||
| CAP-Gly domain containing linker protein family member 4 | CLIP4 | 0.72 | microtubule binding, intracellular transport | [112] |
| Tubulin alpha-1D chain | TUBA1D | 0.59 | assembly of the cytoskeleton | [113] |
| Myocilin | MYOC | -0.56 | cytoskeletal function | [114] |
One of the most highly expressed transcripts in the RSM fed pigs was the mRNA coding for ABTB1 elongation factor involved in PTEN growth-suppressive signaling and protein degradation via ubiquitin- proteasome pathway. Similarly, the amount of OTUD1 mRNA was more than 3 fold greater in RSM pigs implying higher level of proteasomal activity. Further, the RSM pigs exhibited higher levels of transcripts implicated in growth-suppressive signaling and negative regulation of proliferation and apoptosis: IER5, BTG2, KLF10, KLF11, RETREG1 and PRUNE2. The cell cycle regulators ASNS, CEP135, SMC5 and NUPR1 and myogenesis promoter IGFBP5, were all downregulated in RSM group, further suggesting reduced cell proliferation. Nevertheless, observed regulation of transcripts involved in growth suppression and apoptosis in RSM fed pigs was accompanied with downregulation of negative regulators of skeletal muscle mass MSTN, CAPN5 and SAMTOR possibly promoting protein synthesis and muscle growth. In addition, RSM fed pigs had higher levels of several positive regulators of proliferation including HRH1, GDNF, ZMIZ1 and GAB2.
IRS2 transcript encoding a cytoplasmic signaling molecule regulating insulin-mediated glucose transport was upregulated in RSM compared to SBM pigs, while expression of ENHO involved in insulin signaling, metabolic adaptation to fasting and glucose homeostasis decreased. Transcript involved in glycogen metabolism, GBE1, exhibited higher expression in pigs fed RSM compared to the SBM diet.
Majority of lipid metabolism regulators were downregulated by RSM dietary treatment including RDH16, ADIPOQ, FSP27 and ELOVL6, while ACER1 and ST8SIA5 were found upregulated. Expression of NR4A3 transcript involved in the regulation of glucose and fatty acid (FA) utilization genes in skeletal muscle and modulation of feeding behavior and energy balance was more than 4 fold higher in pigs receiving the RSM diet. Moreover, the fiber containing RSM diet increased expression of FGF21, PDK4, ESRRB and ESRRG, which are all known metabolic regulators activated under the conditions of deprived energy or oxidative stress (Table 4).
Dietary treatment affected a number of genes with functions in cellular detoxification pathways and response to reactive oxygen species (ROS) (Table 4). Interestingly, several transcripts with beneficial roles in reducing oxidative stress and enhancing free radical scavenging were more abundant in RSM pigs: GPX1, GPX2, TXNIP, and UCP3. The RSM pigs also had higher levels of an adaptive-response gene ATF3, involved in ER stress response, and EPAS1, which is responsive to hypoxic stress. ABCA8, downregulated by RSM, is a member of the xenobiotic transporter ABC-subfamily, which facilitates cholesterol efflux and mediates transport of xenobiotics, while THBS1, also downregulated by RSM, mediates ER stress response by inducing protective antioxidants.
Two transcripts involved in cytoskeleton organization and cell motility, CLIP4 and TUBA1D, were upregulated in RSM pigs, while MYOC exhibited lower levels compared to control. The majority of genes involved in the organization of the extra cellular matrix (ECM) and cell-matrix adhesion were reduced in RSM pigs, evidenced by the expression of TNMD, CEACAM16, COL11A1, ADAMTS6, FMOD and OGN (Table 4).
Validation of sequencing data by quantitative RT-PCR
qRT-PCR of 5 genes was performed to validate RNAseq data. The majority of genes selected for validation included those involved in the regulation of cellular redox systems (TXNIP, UCP3, ATF3) and energy and muscle metabolism (PVALB and MSTN). Validation revealed high correspondence between RNAseq and qRT-PCR for 4 genes, while higher number of animals are likely needed to confirm PVALB expression (Fig 3).
Fig 3. Validation of RNAseq data by qRT-PCR for 5 selected genes.
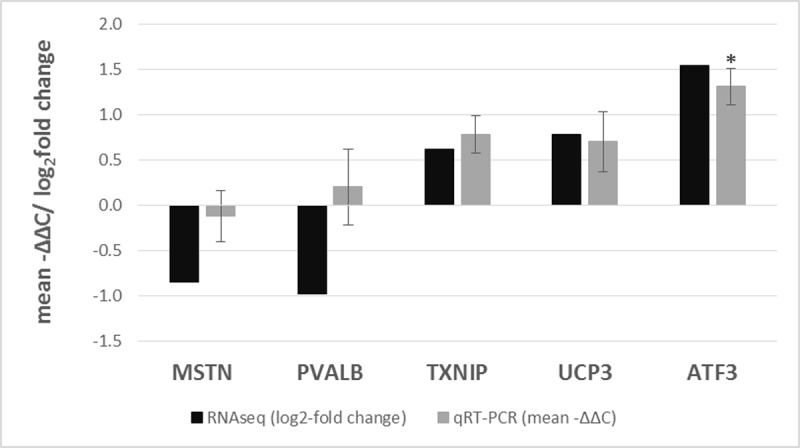
RNAseq data is shown as log2-fold change, while qPCR data is shown as mean –ΔΔCt ± SD, so that they are comparable, (n = 6). All RNAseq results are significant; significant difference detected by qRT-PCR is marked with *. The significance level was set to p<0.05. The expression of selected genes was normalized to GAPDH- glyceraldehyde-3-phosphate dehydrogenase and ACTB- β-actin in the qRT-PCR analysis. TXNIP- thioredoxin-interacting protein; UCP3- uncoupling protein 3 (mitochondrial, proton carrier); ATF3- Activating transcription factor 3; PVALB- parvalbumin; MSTN- myostatin.
Discussion
A number of studies show that increased use of RSM in pig diets holds great potential to improve sustainability and self-sufficiency, while simultaneously reducing environmental burden [115–117]. Here, we report effects of a three-month dietary replacement of SBM with RSM on growth performance, carcass and meat quality traits and skeletal muscle gene expression in growing finishing Norwegian Landrace pigs.
Despite equal calculated net energy and nitrogen contents, and balanced amino acid profiles of the two diets, feeding RSM reduced growth performance. Multiple factors in the RSM diet could contribute to this observation, including lower feed intake, higher content and composition of fiber, and content of bioactive compounds, mainly glucosinolates. Increased use of RSM in diets for pigs is limited by the properties of the low degradable fiber fraction in the RSM, which reduces the availability of energy and nutrients [118–120]. The physicochemical properties of the RSM fiber, including the swelling, water-holding capacity, solubility, degradability and fermentability, have been shown to influence pig performance by affecting feed intake, satiety, digesta passage rate, digestion efficiency and absorption of nutrients [121, 122]. Young pigs fed a similar RSM based diet, using the same RSM type (Mestilla UAB) as in the present experiment, had lower digestibility of nutrients and energy compared to the SBM based control [123]. Detailed profiling of the fiber component revealed that replacement of SBM with RSM resulted in increased content of non-starch polysaccharides, NDF, cellulose and lignin, demonstrating the presence of more indigestible fiber in the RSM compared to the SBM. A similar difference in the fiber fraction between the RSM and the SBM diet can be anticipated in the present study.
The RSM diet resulted in reduced carcass weight and dressing percentage of the pigs. The negative effect on carcass yield is commonly observed after an extended exposure of fibrous diets to pigs, which could at least partially be attributed to viscera enlargement [124]. It has also been shown that larger viscera not only reduce carcass dressing percentage, but at the same time increase energy and nutrient requirements, additionally affecting FCR [125, 126]. In a recent review, Agyekum and Nyachoti [127] provided a detailed description of how feeding fiber-rich diets to pigs leads to increase in maintenance requirements due to enlarged viscera and liver. Signaling transducers, growth factors, hormones and receptors involved in pathways that regulate muscle tissue development and metabolism were found regulated.
At slaughter, the SBM group weighed on average 5 kg more than the RSM group, a difference close to 5%. However, the lower cost of the RSM diet can at least partly compensate for reduction in performance. There were large individual differences in ADG and ADFI in both dietary treatments suggesting that performance traits are to a great extent affected by genetics. Higher levels of NDF and ADF in the RSM diet likely had the largest effect on growth performance, but anti-proliferative effects of glucosinolates could also be a contributing factor [3]. Reduced palatability of glucosinolate-rich diets with consequent negative effects on feed intake is well-documented [128–130]. The concentration of glucosinolates with 20% RSM inclusion slightly exceeded the recommended limit of 2.1 mmol/kg feed for pigs [131], and some of the observed responses could therefore be attributed to biological properties of glucosinolates. Harmful effects of glucosinolates are mainly associated with their breakdown products, goitrogenic substances, which when present in higher amounts interfere with iodine metabolism and disturb the function of the thyroid gland consequently affecting animal performance [3, 128].
DEGs significantly affected by dietary treatments were classified according to their biological function and associated pathways. In general, dietary treatments caused moderate skeletal muscle transcriptional changes. However, RNAseq profiling revealed potentially important differences that could be related to RSM-mediated reduction in growth performance and carcass traits, metabolic adaptations, and heightened response to oxidative stress.
Regulation of cellular proliferation, ECM remodeling and muscle development
In response to the lower available energy and nutrients in the RSM diet, pigs fed this diet showed higher levels of numerous transcripts encoding factors involved in negative regulation of cell growth and apoptosis (IER5, BTG2, KLF10/TIEG1, KLF11/TIEG3, RETREG1, PRUNE2). Downregulation of three essential negative regulators of skeletal muscle mass (MSTN, CAPN5 and FMR1) suggested that cellular growth in RSM pigs was low and did not require additional inhibitory regulation. RET signaling pathway, which is essential for development of neuronal lineages [132], was activated in RSM pigs, and likely had a role during muscle tissue remodeling, as previously described in developing zebrafish muscle [133]. RET signaling is activated by the secreted neurotrophic factor GDNF that modulates connections between nerve and muscle, which showed higher mRNA levels in RSM pigs [134]; [135]. LHX6 was another transcript implied in neural and muscle development that showed the same pattern of expression as GDNF. Upregulated genes involved in positive regulation of proliferation (EPAS1, HRH1, ROS1, ZMIZ1) could be involved in the proliferation of cells other than skeletal muscle cells. Overall, the net effect of genes promoting apoptosis on one hand, versus those stimulating proliferation suggests reduced proliferation and tissue growth in RSM fed pigs. The use of fiber and polyunsaturated fatty acid rich diets that contain bioactive phytochemicals such as glucosinolates from cruciferous vegetables, provides a number of beneficial health effects for humans, including prevention of obesity and chemoprevention mediated by anti-proliferative, as well as anti-oxidant and anti-lipogenic properties of these diets [136].
Genes involved in pathways controlling protein breakdown and synthesis and regulation of muscle mass (ABTB1, OTUD1, PADI2, SPP1) might play a role in hypotrophy in RSM pigs. In addition to the lower availability of nutrients and energy of the RSM diet, lower feed intake in this group could also affect expression of these genes. Two of the most highly expressed transcripts, ABTB1 and OTUD1, suggested increased rate of protein breakdown via ubiquitin proteasome system. Similarly, RSM group showed high expression of SPP1, whose elevated levels were reported in muscle dystrophy accompanied by loss of muscle mass [102]. Gene profiles with roles in skeletal muscle development may be associated with accelerated proteolysis, but they may also play homeostatic functions during normal muscle growth, which requires both protein synthesis and degradation.
RNAseq analysis showed downregulation of numerous genes regulating ECM and connective tissue development in RSM pigs, including the mRNA coding for procollagen COL11A1, and genes important for collagen assembly and ECM structure and function (TNMD, OTOR, CEACAM16, FMOD, ADAMTS6, OGN). Signaling transducers, growth factors, hormones and receptors also likely involved in pathways that regulate muscle tissue development and metabolism were among the top regulated genes (Table 4). Upregulation of SPP1/OPN in RSM further supported need for tissue remodeling and reorganization of ECM [101]. Changes of ECM could affect overall tissue architecture and homeostasis [137, 138], as well as the meat quality traits [137, 139]. However, despite the tissue remodeling that occurred in response to RSM diet, no differences in meat quality traits between the two groups were observed. Slightly higher content of collagen in RSM carcasses (Table 3) can be considered negligible.
Metabolic adaptations to RSM diet
Growing skeletal muscle has large metabolic plasticity to withstand various environmental cues, but variations in dietary composition can pose a metabolic challenge during development affecting muscle mass [140]. Studies have shown that changes in factors mediating metabolic processes contribute to the variation in FE traits [141, 142]. In our study, dietary treatment affected genes and pathways with roles in metabolic activity regulation (FGF21, FGF1, FOS, PDK4, NR4A3, ESRRB, ESRRG and nuclear receptor transcription pathway), some of which were previously reported to affect FE traits in mammals [10, 143, 144]. Molecular regulation of skeletal muscle metabolism is primarily regulated by nuclear receptors, which function as transcription factors that bind DNA and mediate gene expression regulation orchestrating FA metabolism, oxidative phosphorylation and mitochondrial biogenesis [145]. Hence, upregulation of nuclear receptor transcription pathway in the muscle of RSM pigs could be responsible for major differences in skeletal muscle transcription profiles between dietary treatments. In this respect, activation of nuclear receptors ESRRB and ESRRG might have stimulated the upregulation of PDK4 expression [146], a kinase that decreases glucose utilization and increases fat metabolism in response to lower energy levels [80]. Moreover, a downregulation of ENHO/adropin transcript, which plays a critical role in carbohydrate and lipid metabolism and metabolic adaptation to fasting and upregulation of IRS2, FGF21, NR4A3, RRAD and TXNIP involved in insulin and glucose transport suggested that RSM pigs could have enhanced need for glucose uptake compared to control pigs, in order to compensate for the lower energy status. Gao et al. [87] proposed that adropin regulates the preference for fuel selection in skeletal muscle in the feeding and fasting cycle, and plays a role in governing glucose and lipid homeostasis. Hence, reduced levels of adropin mRNA in muscle of RSM fed pigs might have induced a shift to fat use rather than glucose resulting in increased FA uptake and oxidation. This was further supported by marked upregulation of FGF21, a key mediator of energy metabolism that activates lipolysis, increases FA oxidation and enhances glucose uptake in response to nutritional status [147] [148].
In line with this, functional annotation of upregulated DEGs highlighted three biological processes in the RSM muscle: ‘multicellular organismal homeostasis’, ‘response to nutrient levels’ and ‘energy reserve metabolic process’ (Fig 2 and S4 Table)–a set of adaptive biochemical and physiological changes to reduce metabolism and activate pathways to derive energy from stored compounds such as lipids or glycogen. Additional evidence for reduced anabolic processes in RSM fed pigs included regulation of several key genes involved in lipid catabolism and FA oxidation; lower levels of ELOVL6 encoding the only elongase involved in de novo lipogenesis [149]; downregulation of CIDEC/FSP27, which controls triglycerides accumulation and formation of lipid droplets [89]; higher level of UCP3 mitochondrial transporter, which enhances muscle FA oxidation [150].
In accordance to our findings, two recent studies in pigs reported that dietary supplementation of polyunsaturated fatty acids and bioactive polyphenols stimulated the expression of genes involved in lipogenesis and oxidative processes in skeletal muscle [90] [151]. Rapeseed contains high amounts of oleic, linoleic and linolenic long chain fatty acids, which also have the ability to affect expression of lipid metabolic genes [152, 153]. Recent RNAseq studies in pigs investigated effects of dietary supplementation of rapeseed and linseed oils with higher concentration of polyunsaturated fatty acids on transcription profiles in liver and muscle tissues [154] [155]. In addition to anti-adipogenic effects of dietary long chain fatty acids, addition of dietary fiber have also been reported to reduce lipogenesis in pigs [156]. In agreement with this, our RNAseq data revealed a number of genes possibly affected by different composition of dietary long chain fatty acids between the two diets (IRS2, FGF21, FGF1, PDK4, HRH1, UCP3, NR4A3). Genes coding for FGF21, NR4A3, PDK4, UCP3 and IRS2 that were more abundant in the RSM group, were reported to promote muscle lipid oxidation and leaner growth by regulating genes involved in mitochondrial and peroxisomal FA oxidation. FGF21 has been described as a marker of mitochondrial dysfunction and stress, which is induced in muscle to enhance mitochondrial function and efficiency and help in stress adaptation by activating the mTOR-YY1-PGC1α pathway [32]. Similarly, ESRRG transcription factor which had higher expression in RSM pigs has been reported to activate oxidative potential and is considered a key regulator of oxidative metabolism that increases mitochondrial activity in skeletal muscle. Rangwala et al. [35] reported higher levels of ESRRG mRNA in oxidative, slow-twitch muscle fibers and low levels in glycolytic fibers. In this study, regulation of key genes associated with muscle mitochondrial activity and energy metabolism control (FGF21, ESRRG, ESRRB, NR4A3, PDK4) indicated a shift toward the oxidative muscle phenotype, characterized by enhanced mitochondrial function and increased fuel uptake in the skeletal muscle exposed to RSM in comparison to SBM diet.
Scheffler and Gerrard [157] showed that muscle fibers with greater oxidative capacity usually exhibit increased protein turnover which can lead to decrease in efficiency and growth potential. Jing et al. [11] reported downregulation of ESRRB transcript in the skeletal muscle of more efficient pigs showing a negative correlation between improved FE and muscle remodeling toward oxidative phenotype and increased mitochondrial metabolism. In view of these findings, similar adverse effects on performance traits due to increased muscle oxidative metabolism in RSM fed pigs could be anticipated.
Response to oxidative stress
The GO analysis of upregulated genes revealed enriched biological functions associated with modulation of cellular redox state in RSM pigs, namely, response to ROS and oxidative stress (Fig 2 and S4 Table). One of the well-recognized beneficial properties of bioactive phytochemicals from RSM is their potent antioxidant activity and modulation of cellular redox state by the induction of glutathione-based Phase-2 detoxifying enzymes across vertebrates [158, 159], including fish [160]. This is in agreement with observed higher levels of transcripts involved in maintenance of antioxidant defenses and repression of ROS in the RSM group, including GPX1 and GPX2 from the glutathione pathway, TXNIP and UCP3.
Interestingly, several studies have reported metabolic effects of dietary phytochemicals on signaling networks controlling energy metabolism through the regulation of cellular redox state [136, 161, 162]. Lower energy level provided by the RSM diet with the resultant tissue downsizing (by increased proteolysis, decreased cellular proliferation and increased apoptosis), and metabolic adaptations (stimulation of mitochondrial biogenesis, and switch to better utilization of lipid metabolism) strongly suggested lower level of anabolic processes, and consequently lower threat of oxidative stress. Therefore, activation of Phase-2 GPX1 and GPX2 enzymes in RSM pigs can be considered beneficial. The findings of this study could be extrapolated to other farmed animals and humans. Functional consequences of RSM-based and similar diets need to be further investigated to ensure optimal use with no negative effects.
Conclusion
Pigs fed a diet containing 20% RSM with higher content of fiber and bioactive phytochemicals, including glucosinolates, showed somewhat reduced growth performance and dressing percentage compared to pigs fed a control SBM diet. Yet, dietary treatments had no effect on meat quality traits.
The differential gene expression profile reflected growth performance differences between pigs receiving the two diets, revealing underlying molecular mechanisms of observed phenotypes. The analysis of global changes in expression of genes regulating protein degradation, tissue remodeling, carbohydrate and lipid metabolism, mitochondrial adaptation and ROS metabolism improved our understanding of the effects of RSM-based diets on growth performance and skeletal muscle of growing-finishing Norwegian Landrace pigs. The research will help facilitate a more sustainable Norwegian pig production characterized by increased use of local ingredients and reduced environmental impact.
Supporting information
(DOCX)
(DOCX)
(XLSX)
(XLSX)
X-axis shows the fold change (log2) vs p value (-log10). Significantly differentially expressed genes (log2-fold change = > 0.5; p value < = 0.05) area marked in red.
(PNG)
{kind=link}
Acknowledgments
The sequencing service was provided by the Norwegian Sequencing Centre (www.sequencing.uio.no), a national technology platform hosted by the Oslo University Hospital and the University of Oslo and supported by the "Functional Genomics" and "Infrastructure" programs of the Research Council of Norway and the Southeastern Regional Health Authorities.
Data Availability
All relevant data is available in the NCBI Sequence Read Archive under accession number PRJNA506343.
Funding Statement
Funding for this research was provided by the FeedMileage-Efficient use of Feed Resources for a Sustainable Norwegian Food Production (Research Council of Norway, Oslo, Norway; grant no. 233685/E50), Foods of Norway, Centre for Research-based Innovation (Research Council of Norway; grant no. 237841/030), and SusPig ERA-Net (Research Council of Norway, Oslo, Norway; grant no. 272334/E50) projects.
References
- 1.Messerschmidt U, Eklund M, Sauer N, Rist VTS, Rosenfelder P, Spindler HK. Chemical composition and standardized ileal amino acid digestibility in rapeseed meals sourced from German oil mills for growing pigs. Animal Feed Science and Technology. 2014. 187:68–76. [Google Scholar]
- 2.Schone F, Jahreis G, Lange R, Seffner W, Groppel B, Hennig A. Effect of varying glucosinolate and iodine intake via rapeseed meal diets on serum thyroid hormone level and total iodine in the thyroid in growing pigs. Endocrinologia experimentalis. 1990. 24:415–27. [PubMed] [Google Scholar]
- 3.Mejicanos G, Sanjayan N, Kim IH, Nyachoti CM. Recent advances in canola meal utilization in swine nutrition. Journal of animal science and technology. 2016. 58:7 10.1186/s40781-016-0085-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Montagne L, Loisel F, Le Naou T, Gondret F, Gilbert H, Le Gall M. Difference in short-term responses to a high-fiber diet in pigs divergently selected for residual feed intake. Journal of animal science. 2014. 92:1512–23. 10.2527/jas.2013-6623 [DOI] [PubMed] [Google Scholar]
- 5.Schedle K. Sustainable pig and poultry nutrition by improvement of nutrient utilisation–A review. Die Bodenkultur: Journal of Land Management, Food and Environment. 2016. 67:45–60. [Google Scholar]
- 6.Noblet J, Le Goff GI. Effect of dietary fibre on the energy value of feeds for pigs. 2001. 35–52 p. [Google Scholar]
- 7.Noblet J, Gilbert H, Jaguelin-Peyraud Y, Lebrun T. Evidence of genetic variability for digestive efficiency in the growing pig fed a fibrous diet. Animal: an international journal of animal bioscience. 2013. 7:1259–64. [DOI] [PubMed] [Google Scholar]
- 8.Kolstad K, Vangen O. Breed differences in maintenance requirements of growing pigs when accounting for changes in body composition. Livestock Production Science. 1996. 47:23–32. [Google Scholar]
- 9.Perez de Nanclares M, Trudeau MP, Hansen JØ, Mydland LT, Urriola PE, Shurson G. High-fiber rapeseed co-product diet for Norwegian Landrace pigs: Effect on digestibility. 2017. 203: 1–9. [Google Scholar]
- 10.Vincent A, Louveau I, Gondret F, Trefeu C, Gilbert H, Lefaucheur L. Divergent selection for residual feed intake affects the transcriptomic and proteomic profiles of pig skeletal muscle. Journal of animal science. 2015. 93:2745–58. 10.2527/jas.2015-8928 [DOI] [PubMed] [Google Scholar]
- 11.Jing L, Hou Y, Wu H, Miao Y, Li X, Cao J. Transcriptome analysis of mRNA and miRNA in skeletal muscle indicates an important network for differential Residual Feed Intake in pigs. Scientific reports. 2015. 5:11953 10.1038/srep11953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ayuso M, Fernandez A, Nunez Y, Benitez R, Isabel B, Barragan C. Comparative Analysis of Muscle Transcriptome between Pig Genotypes Identifies Genes and Regulatory Mechanisms Associated to Growth, Fatness and Metabolism. PloS one. 2015. 10:e0145162 10.1371/journal.pone.0145162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sauvant D, Perez JM, Tran G. Tables of composition and nutritional value of feed materials. Amstelveen, Netherlands: Wageningen Academic Publishers; 2004. 304 p. [Google Scholar]
- 14.National Research Council NRC. Nutrient Requirements of Swine: Eleventh Revised Edition. The National Academies Press: Washington, DC: 2012. 420 p. [Google Scholar]
- 15.Øverland M, Granli T, Kjos NP, Fjetland O, Steien SH, Stokstad M. Effect of dietary formates on growth performance, carcass traits, sensory quality, intestinal microflora, and stomach alterations in growing-finishing pigs. Journal of animal science. 2000. 78:1875–84. 10.2527/2000.7871875x [DOI] [PubMed] [Google Scholar]
- 16.European Commission EC. Commission Regulation 152/2009 of 26 February 2009 laying down the methods of sampling and analysis for the official control of feed. Off J Eur Comm. 2009. L54:1–130. [Google Scholar]
- 17.International organization for standardization ISO. Animal Feeding Stuffs, Animal Products, and Faeces or urine. Determination of Gross Calorific Value Bomb Calorimeter Method. ISO 9831. International Organization for Standardization, Geneva, Switzerland. 1998.
- 18.Association of Official Analytical Chemists AOAC. Official methods of analysis of the association of official analytical chemists. 15th edition Arlington, Virginia, USA: 1990. p. 1102. [Google Scholar]
- 19.O'Farrell M, Wold JP, Høy M, Tschudi J, Schulerud H. On-Line Fat Content Classification of Inhomogeneous Pork Trimmings Using Multispectral near Infrared Interactance Imaging. Journal of Near Infrared Spectroscopy. 2010. 18:135–45. [Google Scholar]
- 20.Christensen LB. Drip loss sampling in porcine m. longissimus dorsi. Meat science. 2003. 63:469–77. [DOI] [PubMed] [Google Scholar]
- 21.Anderson S. Determination of fat, moisture, and protein in meat and meat products by using the FOSS FoodScan Near-Infrared Spectrophotometer with FOSS Artificial Neural Network Calibration Model and Associated Database: collaborative study. Journal of AOAC International. 2007. 90:1073–83. [PubMed] [Google Scholar]
- 22.Bushnell B. BBTools software package. BBTools—DOE Joint Genome Institute. https://jgi.doe.gov/data-and-tools/bbtools2014
- 23.Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome biology. 2013. 14:R36 10.1186/gb-2013-14-4-r36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nature methods. 2012. 9:357–9. 10.1038/nmeth.1923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Goff L, Trapnell C, Kelley D. cummeRbund: Analysis, exploration, manipulation, and visualization of Cufflinks high-throughput sequencing data. R package version. 2013. 2(0). [Google Scholar]
- 26.SAS. Users Guide. Statistical Analysis System Institute: Cary, NC, United States: 1990. [Google Scholar]
- 27.Reimand J, Arak T, Adler P, Kolberg L, Reisberg S, Peterson H. g:Profiler-a web server for functional interpretation of gene lists (2016 update). Nucleic acids research. 2016. 44:W83–9. 10.1093/nar/gkw199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic acids research. 2000. 28:27–30. 10.1093/nar/28.1.27 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Croft D, O'Kelly G, Wu G, Haw R, Gillespie M, Matthews L. Reactome: a database of reactions, pathways and biological processes. Nucleic acids research. 2011. 39:D691–7. 10.1093/nar/gkq1018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Liu J, Damon M, Guitton N, Guisle I, Ecolan P, Vincent A. Differentially-expressed genes in pig Longissimus muscles with contrasting levels of fat, as identified by combined transcriptomic, reverse transcription PCR, and proteomic analyses. Journal of agricultural and food chemistry. 2009. 57:3808–17. 10.1021/jf8033144 [DOI] [PubMed] [Google Scholar]
- 31.Hamill RM, Aslan O, Mullen AM, O’Doherty JV, McBryan J, Morris DG. Transcriptome analysis of porcine M. semimembranosus divergent in intramuscular fat as a consequence of dietary protein restriction. BMC Genomics. 2013. 14:453 10.1186/1471-2164-14-453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ji K, Zheng J, Lv J, Xu J, Ji X, Luo YB. Skeletal muscle increases FGF21 expression in mitochondrial disorders to compensate for energy metabolic insufficiency by activating the mTOR-YY1-PGC1alpha pathway. Free radical biology & medicine. 2015. 84:161–70. [DOI] [PubMed] [Google Scholar]
- 33.Ayuso M, Fernandez A, Nunez Y, Benitez R, Isabel B, Fernandez AI. Developmental Stage, Muscle and Genetic Type Modify Muscle Transcriptome in Pigs: Effects on Gene Expression and Regulatory Factors Involved in Growth and Metabolism. PloS one. 2016. 11:e0167858 10.1371/journal.pone.0167858 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zhou C, Yang G, Chen M, He L, Xiang L, Ricupero C. Lhx6 and Lhx8: cell fate regulators and beyond. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2015. 29:4083–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Rangwala SM, Wang X, Calvo JA, Lindsley L, Zhang Y, Deyneko G. Estrogen-related receptor gamma is a key regulator of muscle mitochondrial activity and oxidative capacity. The Journal of biological chemistry. 2010. 285:22619–29. 10.1074/jbc.M110.125401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fan W, He N, Lin CS, Wei Z, Hah N, Waizenegger W. ERRgamma Promotes Angiogenesis, Mitochondrial Biogenesis, and Oxidative Remodeling in PGC1alpha/beta-Deficient Muscle. Cell reports. 2018. 22:2521–9. 10.1016/j.celrep.2018.02.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fan W, Atkins AR, Yu RT, Downes M, Evans RM. Road to exercise mimetics: targeting nuclear receptors in skeletal muscle. Journal of molecular endocrinology. 2013. 51:T87–t100. 10.1530/JME-13-0258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Conte C, Ainaoui N, Delluc-Clavieres A, Khoury MP, Azar R, Pujol F. Fibroblast growth factor 1 induced during myogenesis by a transcription-translation coupling mechanism. Nucleic acids research. 2009. 37:5267–78. 10.1093/nar/gkp550 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Micheli L, Leonardi L, Conti F, Maresca G, Colazingari S, Mattei E. PC4/Tis7/IFRD1 stimulates skeletal muscle regeneration and is involved in myoblast differentiation as a regulator of MyoD and NF-kappaB. J Biol Chem. 2011. 286:5691–707. 10.1074/jbc.M110.162842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Stinckens A, Luyten T, Bijttebier J, Van den Maagdenberg K, Dieltiens D, Janssens S. Characterization of the complete porcine MSTN gene and expression levels in pig breeds differing in muscularity. Animal genetics. 2008. 39:586–96. 10.1111/j.1365-2052.2008.01774.x [DOI] [PubMed] [Google Scholar]
- 41.Wan X, Wang D, Xiong Q, Xiang H, Li H, Wang H. Elucidating a molecular mechanism that the deterioration of porcine meat quality responds to increased cortisol based on transcriptome sequencing. Scientific reports. 2016. 6:36589 10.1038/srep36589 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Coletta DK, Balas B, Chavez AO, Baig M, Abdul-Ghani M, Kashyap SR. Effect of acute physiological hyperinsulinemia on gene expression in human skeletal muscle in vivo. American journal of physiology Endocrinology and metabolism. 2008. 294:E910–7. 10.1152/ajpendo.00607.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Weiss N. Stac gets the skeletal L-type calcium channel unstuck. General physiology and biophysics. 2015. 34:101–3. 10.4149/gpb_2015011 [DOI] [PubMed] [Google Scholar]
- 44.Unoki M, Nakamura Y. Growth-suppressive effects of BPOZ and EGR2, two genes involved in the PTEN signaling pathway. Oncogene. 2001. 20:4457–65. 10.1038/sj.onc.1204608 [DOI] [PubMed] [Google Scholar]
- 45.Hai T, Wolfgang CD, Marsee DK, Allen AE, Sivaprasad U. ATF3 and stress responses. Gene expression. 1999. 7:321–35. [PMC free article] [PubMed] [Google Scholar]
- 46.Lu D, Chen J, Hai T. The regulation of ATF3 gene expression by mitogen-activated protein kinases. The Biochemical journal. 2007. 401:559–67. 10.1042/BJ20061081 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ding KK, Shang ZF, Hao C, Xu QZ, Shen JJ, Yang CJ. Induced expression of the IER5 gene by gamma-ray irradiation and its involvement in cell cycle checkpoint control and survival. Radiation and environmental biophysics. 2009. 48:205–13. 10.1007/s00411-009-0213-4 [DOI] [PubMed] [Google Scholar]
- 48.Feng Z, Tang ZL, Li K, Liu B, Yu M, Zhao SH. Molecular characterization of the BTG2 and BTG3 genes in fetal muscle development of pigs. Gene. 2007. 403:170–7. 10.1016/j.gene.2007.08.009 [DOI] [PubMed] [Google Scholar]
- 49.Ribeiro A, Bronk SF, Roberts PJ, Urrutia R, Gores GJ. The transforming growth factor beta(1)-inducible transcription factor TIEG1, mediates apoptosis through oxidative stress. Hepatology (Baltimore, Md). 1999. 30:1490–7. [DOI] [PubMed] [Google Scholar]
- 50.Miyake M, Hayashi S, Iwasaki S, Uchida T, Watanabe K, Ohwada S. TIEG1 negatively controls the myoblast pool indispensable for fusion during myogenic differentiation of C2C12 cells. Journal of cellular physiology. 2011. 226:1128–36. 10.1002/jcp.22434 [DOI] [PubMed] [Google Scholar]
- 51.Grumati P, Dikic I, Stolz A. ER-phagy at a glance. Journal of cell science. 2018. 131:jcs217364. [DOI] [PubMed] [Google Scholar]
- 52.Tatsumi Y, Takano R, Islam MS, Yokochi T, Itami M, Nakamura Y. BMCC1, which is an interacting partner of BCL2, attenuates AKT activity, accompanied by apoptosis. Cell death & disease. 2015. 6:e1607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Spittau B, Krieglstein K. Klf10 and Klf11 as mediators of TGF-beta superfamily signaling. Cell and tissue research. 2012. 347:65–72. 10.1007/s00441-011-1186-6 [DOI] [PubMed] [Google Scholar]
- 54.Balasubramanian MN, Butterworth EA, Kilberg MS. Asparagine synthetase: regulation by cell stress and involvement in tumor biology. American journal of physiology Endocrinology and metabolism. 2013. 304:E789–99. 10.1152/ajpendo.00015.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Inanç B, Pütz M, Lalor P, Dockery P, Kuriyama R, Gergely F. Abnormal centrosomal structure and duplication in Cep135-deficient vertebrate cells. Molecular biology of the cell. 2013. 24:2645–54. 10.1091/mbc.E13-03-0149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ren H, Yin P, Duan C. IGFBP-5 regulates muscle cell differentiation by binding to IGF-II and switching on the IGF-II auto-regulation loop. The Journal of cell biology. 2008. 182:979–91. 10.1083/jcb.200712110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hamidi T, Cano CE, Grasso D, Garcia MN, Sandi MJ, Calvo EL. Nupr1-aurora kinase A pathway provides protection against metabolic stress-mediated autophagic-associated cell death. Clin Cancer Res. 2012. 18:5234–46. 10.1158/1078-0432.CCR-12-0026 [DOI] [PubMed] [Google Scholar]
- 58.Ampatzidou E, Irmisch A, O'Connell MJ, Murray JM. Smc5/6 is required for repair at collapsed replication forks. Molecular and cellular biology. 2006. 26:9387–401. 10.1128/MCB.01335-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Behlke-Steinert S, Touat-Todeschini L, Skoufias DA, Margolis RL. SMC5 and MMS21 are required for chromosome cohesion and mitotic progression. Cell cycle (Georgetown, Tex). 2009. 8:2211–8. [DOI] [PubMed] [Google Scholar]
- 60.Masaki T, Chiba S, Yasuda T, Noguchi H, Kakuma T, Watanabe T. Involvement of hypothalamic histamine H1 receptor in the regulation of feeding rhythm and obesity. Diabetes. 2004. 53:2250–60. 10.2337/diabetes.53.9.2250 [DOI] [PubMed] [Google Scholar]
- 61.Medina VA, Rivera ES. Histamine receptors and cancer pharmacology. British Journal of Pharmacology. 2010. 161:755–67. 10.1111/j.1476-5381.2010.00961.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.McCullough MJ, Peplinski NG, Kinnell KR, Spitsbergen JM. Glial cell line-derived neurotrophic factor protein content in rat skeletal muscle is altered by increased physical activity in vivo and in vitro. Neuroscience. 2011. 174:234–44. 10.1016/j.neuroscience.2010.11.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ali ZA, de Jesus Perez V, Yuan K, Orcholski M, Pan S, Qi W. Oxido-reductive regulation of vascular remodeling by receptor tyrosine kinase ROS1. The Journal of clinical investigation. 2014. 124:5159–74. 10.1172/JCI77484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Napierala JS, Li Y, Lu Y, Lin K, Hauser LA, Lynch DR. Comprehensive analysis of gene expression patterns in Friedreich's ataxia fibroblasts by RNA sequencing reveals altered levels of protein synthesis factors and solute carriers. Disease models & mechanisms. 2017. 10:1353–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Turner DC, Seaborne RA, Sharples AP. Comparative Transcriptome and Methylome Analysis in Human Skeletal Muscle Anabolism, Hypertrophy and Epigenetic Memory. Scientific reports. 2019. 9:4251 10.1038/s41598-019-40787-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wohrle FU, Daly RJ, Brummer T. Function, regulation and pathological roles of the Gab/DOS docking proteins. Cell communication and signaling: CCS. 2009. 7:22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gameiro PA, Struhl K. Nutrient Deprivation Elicits a Transcriptional and Translational Inflammatory Response Coupled to Decreased Protein Synthesis. Cell reports. 2018. 24:1415–24. 10.1016/j.celrep.2018.07.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Gu X, Orozco JM, Saxton RA, Condon KJ, Liu GY, Krawczyk PA. SAMTOR is an S-adenosylmethionine sensor for the mTORC1 pathway. Science. 2017. 358:813–8. 10.1126/science.aao3265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Larsson VJ, Jafferali MH, Vijayaraghavan B, Figueroa RA, Hallberg E. Mitotic spindle assembly and γ-tubulin localisation depend on the integral nuclear membrane protein, Samp1. Journal of cell science. 2018. jcs.211664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Mevissen TE, Hospenthal MK, Geurink PP, Elliott PR, Akutsu M, Arnaudo N. OTU deubiquitinases reveal mechanisms of linkage specificity and enable ubiquitin chain restriction analysis. Cell. 2013. 154:169–84. 10.1016/j.cell.2013.05.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Vossenaar ER, Radstake TR, van der Heijden A, van Mansum MA, Dieteren C, de Rooij DJ. Expression and activity of citrullinating peptidylarginine deiminase enzymes in monocytes and macrophages. Annals of the rheumatic diseases. 2004. 63:373–81. 10.1136/ard.2003.012211 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Edeling MA, Smith C, Owen D. Life of a clathrin coat: insights from clathrin and AP structures. Nature reviews Molecular cell biology. 2006. 7:32–44. 10.1038/nrm1786 [DOI] [PubMed] [Google Scholar]
- 73.Fujimoto M, Nakai A. The heat shock factor family and adaptation to proteotoxic stress. The FEBS journal. 2010. 277:4112–25. [DOI] [PubMed] [Google Scholar]
- 74.Santini E, Huynh TN, Longo F, Koo SY, Mojica E, D'Andrea L. Reducing eIF4E-eIF4G interactions restores the balance between protein synthesis and actin dynamics in fragile X syndrome model mice. Science signaling. 2017. 10(504). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Choi DW, Seo YM, Kim EA, Sung KS, Ahn JW, Park SJ. Ubiquitination and degradation of homeodomain-interacting protein kinase 2 by WD40 repeat/SOCS box protein WSB-1. The Journal of biological chemistry. 2008. 283:4682–9. 10.1074/jbc.M708873200 [DOI] [PubMed] [Google Scholar]
- 76.Singh R, Brewer MK, Mashburn CB, Lou D, Bondada V, Graham B. Calpain 5 is highly expressed in the central nervous system (CNS), carries dual nuclear localization signals, and is associated with nuclear promyelocytic leukemia protein bodies. The Journal of biological chemistry. 2014. 289:19383–94. 10.1074/jbc.M114.575159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Kawasaki E, Hokari F, Sasaki M, Sakai A, Koshinaka K, Kawanaka K. The effects of beta-adrenergic stimulation and exercise on NR4A3 protein expression in rat skeletal muscle. The journal of physiological sciences: JPS. 2011. 61:1–11. 10.1007/s12576-010-0114-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Pearen MA, Goode JM, Fitzsimmons RL, Eriksson NA, Thomas GP, Cowin GJ. Transgenic muscle-specific Nor-1 expression regulates multiple pathways that effect adiposity, metabolism, and endurance. Molecular endocrinology (Baltimore, Md). 2013. 27:1897–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Mao C, Obeid LM. Ceramidases: regulators of cellular responses mediated by ceramide, sphingosine, and sphingosine-1-phosphate. Biochimica et biophysica acta. 2008. 1781:424–34. 10.1016/j.bbalip.2008.06.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wu P, Blair PV, Sato J, Jaskiewicz J, Popov KM, Harris RA. Starvation increases the amount of pyruvate dehydrogenase kinase in several mammalian tissues. Archives of biochemistry and biophysics. 2000. 381:1–7. 10.1006/abbi.2000.1946 [DOI] [PubMed] [Google Scholar]
- 81.Zhang S, Hulver MW, McMillan RP, Cline MA, Gilbert ER. The pivotal role of pyruvate dehydrogenase kinases in metabolic flexibility. Nutrition & metabolism. 2014. 11:10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Chibalin AV, Yu M, Ryder JW, Song XM, Galuska D, Krook A. Exercise-induced changes in expression and activity of proteins involved in insulin signal transduction in skeletal muscle: Differential effects on insulin-receptor substrates 1 and 2. Proceedings of the National Academy of Sciences. 2000. 97:38–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Greenberg CC, Jurczak MJ, Danos AM, Brady MJ. Glycogen branches out: new perspectives on the role of glycogen metabolism in the integration of metabolic pathways. American journal of physiology Endocrinology and metabolism. 2006. 291:E1–8. 10.1152/ajpendo.00652.2005 [DOI] [PubMed] [Google Scholar]
- 84.Narasimhan A, Greiner R, Bathe OF, Baracos V, Damaraju S. Differentially expressed alternatively spliced genes in skeletal muscle from cancer patients with cachexia. Journal of cachexia, sarcopenia and muscle. 2018. 1:60–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kojima M, Nakajima I, Arakawa A, Mikawa S, Matsumoto T, Uenishi H. Differences in gene expression profiles for subcutaneous adipose, liver, and skeletal muscle tissues between Meishan and Landrace pigs with different backfat thicknesses. PloS one. 2018. 13:e0204135-e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Kumar KG, Trevaskis JL, Lam DD, Sutton GM, Koza RA, Chouljenko VN. Identification of adropin as a secreted factor linking dietary macronutrient intake with energy homeostasis and lipid metabolism. Cell metabolism. 2008. 8:468–81. 10.1016/j.cmet.2008.10.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Gao S, McMillan RP, Zhu Q, Lopaschuk GD, Hulver MW, Butler AA. Therapeutic effects of adropin on glucose tolerance and substrate utilization in diet-induced obese mice with insulin resistance. Molecular metabolism. 2015. 4:310–24. 10.1016/j.molmet.2015.01.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Tao X, Liang Y, Yang X, Pang J, Zhong Z, Chen X. Transcriptomic profiling in muscle and adipose tissue identifies genes related to growth and lipid deposition. PloS one. 2017. 12:e0184120-e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Puri V, Ranjit S, Konda S, Nicoloro SM, Straubhaar J, Chawla A. Cidea is associated with lipid droplets and insulin sensitivity in humans. Proceedings of the National Academy of Sciences of the United States of America. 2008. 105:7833–8. 10.1073/pnas.0802063105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Vitali M, Dimauro C, Sirri R, Zappaterra M, Zambonelli P, Manca E. Effect of dietary polyunsaturated fatty acid and antioxidant supplementation on the transcriptional level of genes involved in lipid and energy metabolism in swine. PloS one. 2018. 13:e0204869 10.1371/journal.pone.0204869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Chan KM, Decker EA, Feustman C. Endogenous skeletal muscle antioxidants. Critical Reviews in Food Science and Nutrition. 1994. 34:403–26. 10.1080/10408399409527669 [DOI] [PubMed] [Google Scholar]
- 92.Halliwell B, Murcia MA, Chirico S, Aruoma OI. Free radicals and antioxidants in food and in vivo: what they do and how they work. Crit Rev Food Sci Nutr. 1995. 35:7–20. 10.1080/10408399509527682 [DOI] [PubMed] [Google Scholar]
- 93.Brigelius-Flohe R. Glutathione peroxidases and redox-regulated transcription factors. Biological chemistry. 2006. 387:1329–35. 10.1515/BC.2006.166 [DOI] [PubMed] [Google Scholar]
- 94.Cardoso S, Santos RX, Carvalho C, Correia S, Santos MS, Moreira PI. Mitochondrial Uncoupling Proteins and Oxidative Stress: Implications for Diabetes and Neurodegeneration. Free Radicals and Antioxidants 2011. p. 4–14. [Google Scholar]
- 95.Hand LE, Saer BR, Hui ST, Jinnah HA, Steinlechner S, Loudon AS. Induction of the metabolic regulator Txnip in fasting-induced and natural torpor. Endocrinology. 2013. 154:2081–91. 10.1210/en.2012-2051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Alhawiti NM, Al Mahri S, Aziz MA, Malik SS, Mohammad S. TXNIP in Metabolic Regulation: Physiological Role and Therapeutic Outlook. Current drug targets. 2017. 18:1095–103. 10.2174/1389450118666170130145514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kaelin WG Jr. , Ratcliffe PJ. Oxygen sensing by metazoans: the central role of the HIF hydroxylase pathway. Molecular cell. 2008. 30:393–402. 10.1016/j.molcel.2008.04.009 [DOI] [PubMed] [Google Scholar]
- 98.Cascorbi I. Role of pharmacogenetics of ATP-binding cassette transporters in the pharmacokinetics of drugs. Pharmacology & therapeutics. 2006. 112:457–73. [DOI] [PubMed] [Google Scholar]
- 99.Cardoso TF, Quintanilla R, Tibau J, Gil M, Mármol-Sánchez E, González-Rodríguez O. Nutrient supply affects the mRNA expression profile of the porcine skeletal muscle. BMC Genomics. 2017. 18:603 10.1186/s12864-017-3986-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Lee JH, Budanov AV, Talukdar S, Park EJ, Park HL, Park HW. Maintenance of metabolic homeostasis by Sestrin2 and Sestrin3. Cell metabolism. 2012. 16:311–21. 10.1016/j.cmet.2012.08.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Liaw L, Birk DE, Ballas CB, Whitsitt JS, Davidson JM, Hogan BL. Altered wound healing in mice lacking a functional osteopontin gene (spp1). The Journal of clinical investigation. 1998. 101:1468–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Vetrone SA, Montecino-Rodriguez E, Kudryashova E, Kramerova I, Hoffman EP, Liu SD. Osteopontin promotes fibrosis in dystrophic mouse muscle by modulating immune cell subsets and intramuscular TGF-beta. The Journal of clinical investigation. 2009. 119:1583–94. 10.1172/JCI37662 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Pagel CN, Wasgewatte Wijesinghe DK, Taghavi Esfandouni N, Mackie EJ. Osteopontin, inflammation and myogenesis: influencing regeneration, fibrosis and size of skeletal muscle. Journal of cell communication and signaling. 2014. 8:95–103. 10.1007/s12079-013-0217-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Tsutsui K, Manabe R, Yamada T, Nakano I, Oguri Y, Keene DR. ADAMTSL-6 is a novel extracellular matrix protein that binds to fibrillin-1 and promotes fibrillin-1 fibril formation. The Journal of biological chemistry. 2010. 285:4870–82. 10.1074/jbc.M109.076919 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Robertson NG, Heller S, Lin JS, Resendes BL, Weremowicz S, Denis CS. A novel conserved cochlear gene, OTOR: identification, expression analysis, and chromosomal mapping. Genomics. 2000. 66:242–8. 10.1006/geno.2000.6224 [DOI] [PubMed] [Google Scholar]
- 106.Kuespert K, Pils S, Hauck CR. CEACAMs: their role in physiology and pathophysiology. Current opinion in cell biology. 2006. 18:565–71. 10.1016/j.ceb.2006.08.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Shukunami C, Yoshimoto Y, Takimoto A, Yamashita H, Hiraki Y. Molecular characterization and function of tenomodulin, a marker of tendons and ligaments that integrate musculoskeletal components. The Japanese dental science review. 2016. 52:84–92. 10.1016/j.jdsr.2016.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Kelwick R, Desanlis I, Wheeler GN, Edwards DR. The ADAMTS (A Disintegrin and Metalloproteinase with Thrombospondin motifs) family. Genome biology. 2015. 16:113 10.1186/s13059-015-0676-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Li Y, Lacerda DA, Warman ML, Beier DR, Yoshioka H, Ninomiya Y. A fibrillar collagen gene, Col11a1, is essential for skeletal morphogenesis. Cell. 1995. 80:423–30. 10.1016/0092-8674(95)90492-1 [DOI] [PubMed] [Google Scholar]
- 110.Jan AT, Lee EJ, Choi I. Fibromodulin: A regulatory molecule maintaining cellular architecture for normal cellular function. The international journal of biochemistry & cell biology. 2016. 80:66–70. [DOI] [PubMed] [Google Scholar]
- 111.Deckx S, Heymans S, Papageorgiou AP. The diverse functions of osteoglycin: a deceitful dwarf, or a master regulator of disease? FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2016. 30:2651–61. [DOI] [PubMed] [Google Scholar]
- 112.Ghahramani Seno MM, Trollet C, Athanasopoulos T, Graham IR, Hu P, Dickson G. Transcriptomic analysis of dystrophin RNAi knockdown reveals a central role for dystrophin in muscle differentiation and contractile apparatus organization. BMC genomics. 2010. 11:345 10.1186/1471-2164-11-345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Sidyakin AA, Kaysheva AL, Kopylov AT, Lobanov AV, Morozov SG. Proteomic Analysis of Cerebral Cortex Extracts from Sus scrofa with Induced Hemorrhagic Stroke. Journal of molecular neuroscience: MN. 2018. 65:28–34. 10.1007/s12031-018-1064-8 [DOI] [PubMed] [Google Scholar]
- 114.Anderssohn AM, Cox K, O'Malley K, Dees S, Hosseini M, Boren L. Molecular chaperone function for myocilin. Investigative ophthalmology & visual science. 2011. 52:7548–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Eriksson IS, Elmquist H, Stern S, Nybrant T. Environmental Systems Analysis of Pig Production—The Impact of Feed Choice The International Journal of Life Cycle Assessment. 2005. 10:143–54. [Google Scholar]
- 116.Weightman R, Garland P, Phelps E, Clarke S, Hazzledine M, Berry P. Nutritional value of oilseed rape and its co-products for pig and poultry feed: potential improvements and implications for plant breeders. HGCA Research Review. 2014. 80:58. [Google Scholar]
- 117.Skoufos I, Tzora A, Giannenas I, Bonos E, Papagiannis N, Tsinas A. Dietary Inclusion of Rapeseed Meal as Soybean Meal Substitute on Growth Performance, Gut Microbiota, Oxidative Stability and Fatty Acid Profile in Growing-Fattening Pigs. Asian Journal of Animal and Veterinary Advances. 2016. 11:89–97. [Google Scholar]
- 118.Landero JL, Beltranena E, Cervantes M, Morales A, Zijlstra RT. The effect of feeding solvent-extracted canola meal on growth performance and diet nutrient digestibility in weaned pigs. Animal Feed Science and Technology. 2011. 170:136–40. [Google Scholar]
- 119.Lindberg JE. Fiber effects in nutrition and gut health in pigs. Journal of animal science and biotechnology. 2014. 5:15 10.1186/2049-1891-5-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Knudsen KE. Fiber and nonstarch polysaccharide content and variation in common crops used in broiler diets. Poultry science. 2014. 93:2380–93. 10.3382/ps.2014-03902 [DOI] [PubMed] [Google Scholar]
- 121.Potkins ZV, Lawrence TL, Thomlinson JR. Effects of structural and non-structural polysaccharides in the diet of the growing pig on gastric emptying rate and rate of passage of digesta to the terminal ileum and through the total gastrointestinal tract. The British journal of nutrition. 1991. 65:391–413. 10.1079/bjn19910100 [DOI] [PubMed] [Google Scholar]
- 122.Bach Knudsen KE, Hansen I. Gastrointestinal implications in pigs of wheat and oat fractions. 1. Digestibility and bulking properties of polysaccharides and other major constituents. The British journal of nutrition. 1991. 65:217–32. 10.1079/bjn19910082 [DOI] [PubMed] [Google Scholar]
- 123.Perez de Nanclares M, Marcussen C, Tauson AH, Hansen JØ, Kjos NP, Mydland LT. Increasing levels of rapeseed expeller meal in diets for pigs: effects on protein and energy metabolism. Animal: an international journal of animal bioscience. 2018:1–10. [DOI] [PubMed] [Google Scholar]
- 124.Jorgensen H, Zhao XQ, Eggum BO. The influence of dietary fibre and environmental temperature on the development of the gastrointestinal tract, digestibility, degree of fermentation in the hind-gut and energy metabolism in pigs. The British journal of nutrition. 1996. 75:365–78. 10.1079/bjn19960140 [DOI] [PubMed] [Google Scholar]
- 125.Nyachoti CM, de Lange CFM, McBride BW, Leeson S, Schulze H. Dietary influence on organ size and in vitro oxygen consumption by visceral organs of growing pigs. Livestock Production Science. 2000. 65:229–37. [Google Scholar]
- 126.Patience JF, Rossoni-Serao MC, Gutierrez NA. A review of feed efficiency in swine: biology and application. Journal of animal science and biotechnology. 2015. 6:33 10.1186/s40104-015-0031-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Agyekum AK, Nyachoti CM. Nutritional and Metabolic Consequences of Feeding High-Fiber Diets to Swine: A Review. Engineering. 2017. 3:716–25. [Google Scholar]
- 128.Mawson R, Heaney RK, Zdunczyk Z, Kozlowska H. Rapeseed meal-glucosinolates and their antinutritional effects. Part 3. Animal growth and performance. Die Nahrung. 1994. 38:167–77. [DOI] [PubMed] [Google Scholar]
- 129.Tripathi MK, Mishra AS. Glucosinolates in animal nutrition: A review. Animal Feed Science and Technology. 2007. 132:1–27. [Google Scholar]
- 130.Tripathi MK, Mishra AS. Prospects and Problems of Dietary Glucosinolates in Animal Feeding. J Adv Dairy Res. 2017. 5:180. [Google Scholar]
- 131.European Food Safety Authority EFSA. Glucosinolates as undesirable substances in animal feed—Scientific Opinion of the Panel on Contaminants in the Food Chain. EFSA Journal. 2008. 6:590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Moyle LA, Blanc E, Jaka O, Prueller J, Banerji CR, Tedesco FS. Ret function in muscle stem cells points to tyrosine kinase inhibitor therapy for facioscapulohumeral muscular dystrophy. eLife. 2016. 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Knight RD, Mebus K, d'Angelo A, Yokoya K, Heanue T, Roehl H. Ret signalling integrates a craniofacial muscle module during development. Development (Cambridge, England). 2011. 138:2015–24. [DOI] [PubMed] [Google Scholar]
- 134.Yang LX, Nelson PG. Glia cell line-derived neurotrophic factor regulates the distribution of acetylcholine receptors in mouse primary skeletal muscle cells. Neuroscience. 2004. 128:497–509. 10.1016/j.neuroscience.2004.06.067 [DOI] [PubMed] [Google Scholar]
- 135.Baudet C, Pozas E, Adameyko I, Andersson E, Ericson J, Ernfors P. Retrograde signaling onto Ret during motor nerve terminal maturation. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2008. 28:963–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Tosetti F, Noonan DM, Albini A. Metabolic regulation and redox activity as mechanisms for angioprevention by dietary phytochemicals. International journal of cancer. 2009. 125:1997–2003. 10.1002/ijc.24677 [DOI] [PubMed] [Google Scholar]
- 137.Velleman SG. Meat Science and Muscle Biology Symposium: extracellular matrix regulation of skeletal muscle formation. Journal of animal science. 2012. 90:936–41. 10.2527/jas.2011-4497 [DOI] [PubMed] [Google Scholar]
- 138.Thorsteinsdottir S, Deries M, Cachaco AS, Bajanca F. The extracellular matrix dimension of skeletal muscle development. Developmental biology. 2011. 354:191–207. 10.1016/j.ydbio.2011.03.015 [DOI] [PubMed] [Google Scholar]
- 139.Du M, Carlin KM. Meat Science and Muscle Biology Symposium: extracellular matrix in skeletal muscle development and meat quality. Journal of animal science. 2012. 90:922–3. 10.2527/jas.2011-4937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.de Lange P, Moreno M, Silvestri E, Lombardi A, Goglia F, Lanni A. Fuel economy in food-deprived skeletal muscle: signaling pathways and regulatory mechanisms. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2007. 21:3431–41. [DOI] [PubMed] [Google Scholar]
- 141.Dekkers JC, Gilbert H, editors. Genetic and biological aspect of residual feed intake in pigs. 9 World Congress on Genetics Applied to Livestock Production; 2010: Leipzig, Deutschland.
- 142.Do DN, Strathe AB, Ostersen T, Pant SD, Kadarmideen HN. Genome-wide association and pathway analysis of feed efficiency in pigs reveal candidate genes and pathways for residual feed intake. Frontiers in genetics. 2014. 5:307 10.3389/fgene.2014.00307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Faure J, Lefaucheur L, Bonhomme N, Ecolan P, Meteau K, Coustard SM, et al. Consequences of divergent selection for residual feed intake in pigs on muscle energy metabolism and meat quality. Meat science. 2013. 93:37–45. 10.1016/j.meatsci.2012.07.006 [DOI] [PubMed] [Google Scholar]
- 144.Tizioto PC, Coutinho LL, Oliveira PS, Cesar AS, Diniz WJ, Lima AO, et al. Gene expression differences in Longissimus muscle of Nelore steers genetically divergent for residual feed intake. Scientific reports. 2016. 6:39493 10.1038/srep39493 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Sfyri P, Narkar V, Matsakas A. Regulation of skeletal muscle metabolic and angiogenic properties by nuclear hormone receptors: Implications for skeletal muscle regeneration. Neuromuscular Disorders. 2015. 25:S185. [Google Scholar]
- 146.Zhang Y, Ma K, Sadana P, Chowdhury F, Gaillard S, Wang F. Estrogen-related receptors stimulate pyruvate dehydrogenase kinase isoform 4 gene expression. The Journal of biological chemistry. 2006. 281:39897–906. 10.1074/jbc.M608657200 [DOI] [PubMed] [Google Scholar]
- 147.Kharitonenkov A, Shiyanova TL, Koester A, Ford AM, Micanovic R, Galbreath EJ, et al. FGF-21 as a novel metabolic regulator. The Journal of clinical investigation. 2005. 115:1627–35. 10.1172/JCI23606 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Potthoff MJ, Inagaki T, Satapati S, Ding X, He T, Goetz R. FGF21 induces PGC-1α and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response. Proceedings of the National Academy of Sciences. 2009. 106:10853–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Corominas J, Ramayo-Caldas Y, Puig-Oliveras A, Perez-Montarelo D, Noguera JL, Folch JM. Polymorphism in the ELOVL6 gene is associated with a major QTL effect on fatty acid composition in pigs. PloS one. 2013. 8:e53687 10.1371/journal.pone.0053687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Costford SR, Seifert EL, Bezaire V, M FG, Bevilacqua L, Gowing A. The energetic implications of uncoupling protein-3 in skeletal muscle. Applied physiology, nutrition, and metabolism = Physiologie appliquee, nutrition et metabolisme. 2007. 32:884–94. 10.1139/H07-063 [DOI] [PubMed] [Google Scholar]
- 151.Sirri R, Vitali M, Zambonelli P, Giannini G, Zappaterra M, Lo Fiego DP. Effect of diets supplemented with linseed alone or combined with vitamin E and selenium or with plant extracts, on Longissimus thoracis transcriptome in growing-finishing Italian Large White pigs. Journal of animal science and biotechnology. 2018. 9:81 10.1186/s40104-018-0297-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Kaur N, Chugh V, Gupta AK. Essential fatty acids as functional components of foods- a review. Journal of food science and technology. 2014. 51:2289–303. 10.1007/s13197-012-0677-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Figueiredo PS, Inada AC, Marcelino G, Cardozo CML, de Cassia KF, de Cassia RAG. Fatty Acids Consumption: The Role Metabolic Aspects Involved in Obesity and Its Associated Disorders. Nutrients. 2017. 9:1158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Oczkowicz M, Świątkiewicz M, Ropka-Molik K, Gurgul A, Żukowski K. Effects of Different Sources of Fat in the Diet of Pigs on the Liver Transcriptome Estimated by RNA-Seq. 2016. 16:1073. [Google Scholar]
- 155.Ogluszka M, Szostak A, Te Pas MFW, Polawska E, Urbanski P, Blicharski T, et al. A porcine gluteus medius muscle genome-wide transcriptome analysis: dietary effects of omega-6 and omega-3 fatty acids on biological mechanisms. Genes & nutrition. 2017. 12:4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Oczkowicz M, Szmatoła T, Świątkiewicz M, Pawlina-Tyszko K, Gurgul A, Ząbek T. Corn dried distillers grains with solubles (cDDGS) in the diet of pigs change the expression of adipose genes that are potential therapeutic targets in metabolic and cardiovascular diseases. BMC Genomics. 2018. 19:864 10.1186/s12864-018-5265-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Scheffler TL, Gerrard DE. GROWTH AND DEVELOPMENT SYMPOSIUM: Adenosine monophosphate-activated protein kinase and mitochondria in Rendement Napole pig growth. Journal of animal science. 2016. 94:3601–12. 10.2527/jas.2015-0055 [DOI] [PubMed] [Google Scholar]
- 158.Valgimigli L, Iori R. Antioxidant and pro-oxidant capacities of ITCs. Environmental and molecular mutagenesis. 2009. 50:222–37. 10.1002/em.20468 [DOI] [PubMed] [Google Scholar]
- 159.Vig AP, Rampal G, Thind TS, Arora S. Bio-protective effects of glucosinolates–A review. LWT—Food Science and Technology. 2009. 42:1561–72. [Google Scholar]
- 160.Skugor S, Jodaa Holm H, Bjelland AK, Pino J, Evensen O, Krasnov A. Nutrigenomic effects of glucosinolates on liver, muscle and distal kidney in parasite-free and salmon louse infected Atlantic salmon. Parasites & vectors. 2016. 9:639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Orzechowski A, Ostaszewski P, Jank M, Berwid SJ. Bioactive substances of plant origin in food—impact on genomics. Reproduction, nutrition, development. 2002. 42:461–77. [DOI] [PubMed] [Google Scholar]
- 162.Qin S, Hou D-X. The Biofunctions of Phytochemicals and Their Applications in Farm Animals: The Nrf2/Keap1 System as a Target. Engineering. 2017. 3:738–52. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(DOCX)
(XLSX)
(XLSX)
X-axis shows the fold change (log2) vs p value (-log10). Significantly differentially expressed genes (log2-fold change = > 0.5; p value < = 0.05) area marked in red.
(PNG)
Data Availability Statement
All relevant data is available in the NCBI Sequence Read Archive under accession number PRJNA506343.