

Abstract
The visual system in the central nervous system processes diverse visual signals. Although the overall structure has been characterized from the retina through the lateral geniculate nucleus to the visual cortex, the system is complex. Cellular and molecular studies have been conducted to elucidate the mechanisms underpinning visual processing and, by extension, disease mechanisms. These studies may contribute to the development of artificial visual systems. To validate the results of these studies, behavioral vision testing is necessary. Here, we show that the looming stimulation experiment is a reliable mouse vision test that requires a relatively simple setup. The looming experiment was conducted in a large enclosure with a shelter in one corner and a computer monitor located on the ceiling. A CCD camera positioned next to the computer monitor served to observe mouse behavior. A mouse was placed in the enclosure for 10 minutes and allowed to acclimate to and explore the surroundings. Then, the monitor projected a program-derived looming stimulus 10 times. The mouse responded to the stimuli either by freezing or by fleeing to the hiding place. The mouse’s behavior before and after the looming stimuli was recorded, and the video was analyzed using motion tracking software. The velocity of the mouse movement significantly changed after the looming stimuli. In contrast, no reaction was observed in blind mice. Our results demonstrate that the simple looming experiment is a reliable test of mouse vision.
Keywords: This Month in JoVE, Issue 148, mouse behavior, vision test, flight, freeze, motion track, fear
Introduction
The visual system starts at the retina, where visual signals are captured by photoreceptors, channeled to bipolar cells (2nd-order neurons), and finally passed to ganglion cells (3rd-order neurons). Retinal 2nd- and 3rd-order neurons are thought to form multiple neural pathways that convey particular aspects of visual signaling such as color, motion, or shape. These diverse visual features are relayed to the lateral geniculate nucleus and the visual cortex. In contrast, visual signals leading to eye movement are sent to the superior colliculus. Classically, two retino-cortical pathways have been identified: the magnocellular and the parvocellular pathways. These pathways encode moving and stationary objects», respectively, and their existence embodies the basic concept of parallel processing1,2,3,4,5,6. Recently, more than 15 types of bipolar cells7,8,9,10,11 and ganglion cells12,13,14,15,16 have been reported in the retina of many species, including the primate retina. These cells are distinguished not only by morphological aspects, but also by the expression of distinct markers and genes8,10,1, 18, suggesting that various features of visual signals are processed in parallel, which is more complicated than originally anticipated.
Cellular and molecular technologies have contributed to our understanding of visual processing and potential disease mechanisms that may arise from aberrant visual processing. Such an understanding may contribute to the development of artificial eyes. Although cellular examinations and analysis offer in-depth knowledge at a cellular level, a combination of behavioral experiments and cellular experiments would significantly augment our current understanding of minute visual processes. For example, Yoshida et al.19 found that starburst amacrine cells are the key neurons for motion detection in the mouse retina. Following cellular experiments, they performed the optokinetic nystagmus (OKN) behavioral experiment to show that mutant mice in which starburst amacrine cells were dysfunctional did not respond to moving objects, thereby confirming their cellular investigations. In addition, Pearson et al.20 conducted photoreceptor transplantation in the mouse retina to restore vision in diseased mice. They conducted not only cellular experiments, but also measured mouse behavior through the use of optomotor response recordings and water-maze tasks thus allowing Pearson et al. to verify that transplanted photoreceptors restored vision in the formerly blind mice. Taken together, behavioral experiments are strong tools to assess mouse vision.
Multiple methods are available for measuring mouse vision. These methods have advantages and limitations. In vivo ERG provides information on whether the mouse retina, particularly photoreceptors and ON bipolar cells, appropriately responds to light stimuli. ERG can be tested either under scotopic or photopic conditions21, 2. However, ERG requires anesthesia, which might affect the output measurement23. The optokinetic reflex (OKR) or optomotor response (OMR) is a robust method to assess contrast sensitivity and spatial resolution, both functional components of mouse vision. However, OKR requires surgery to attach a fixation device to the mouse skull24. OMR requires neither surgery nor mouse training; however, it requires training to allow an experimenter to subjectively detect subtle mouse head movements in response to a moving grating in an optic drum 25,26. Pupil light reflex measures pupil constriction in response to light stimuli, which does not require anesthesia and exhibits objective and robust responses 19. Although the pupil reflex simulates retinal light response in vivo, the reflex is mediated mainly by the intrinsically photosensitive retinal ganglion cells (ipRGCs)27. Because ipRGCs represent a small minority of RGCs and do not serve as conventional image-forming ganglion cells, this measurement does not provide information pertaining to the majority of ganglion cells.
The looming light experiment has not previously been considered a major test for measuring mouse vision. However, it is also a robust and reliable vision test across various species, such as mouse28,29, zebrafish30, locust31,32, and human33,34,35. Importantly, the looming experiment is one of only a few methods to test the image-forming pathway - it is not a reflex pathway - given the visual and the limbic systems in the central nervous system are involved in this circuit36,37,38‘.We have established a looming visual stimulus system and have demonstrated its ability to elicit motion detection in the mouse, which we use as a proxy to assess the intactness of the mouse visual system.
Protocol
All experiments and animal care were conducted in accordance with protocol approved by the Institutional Animal Care and Use Committees at Wayne State University (protocol no. 17-11-0399).
1. Preparation for the experiment
Build a rectangular open-lid enclosure to house the mouse during looming visual stimuli presentation. We constructed a 40 cm × 50 cm × 33 cm enclosure using aluminum framing and PVC panels (Figure 1A,B). Lay a sheet of paper to cover the entire floor of the enclosure to ensure easy cleanup between trials. Add an opaque shelter in a corner of the enclosure with an entrance facing the center of the arena for easy entrance and exit.
Set up a camera with a wide-angle lens for capturing the mouse’s behavior. Secure the camera to a table-mounted stand adjacent to the enclosure. For best quality video capturing, use a camera frame rate of 60 FPS or higher.
Set up a computer monitor on top of the enclosure. Because the monitor could not be seen from the outside, a second monitor was prepared, which duplicated the images shown on the primary monitor.
-
Prepare a looming pattern for projection. One way to do this is to use the PsychToolbox3 within MatLab software to code for an expanding black circle (Figure 1C). Set the stimulus to begin at a visual angle of 2° and expand to 50° over 250 ms; these parameters determine stimulus speed (see Figure 1D for visual angle calculation). Set the code to repeat the stimulus 10 times with an interval of 1 s.
NOTE: The stimulus began each repetition immediately upon termination of the previous presentation. For further information on stimulus presentation, please refer to section 3.
Select mice of interest for the looming stimuli. Here, use 32 healthy-eyed mice of a C57 background, male and female, 4 to 14 weeks old. Also, use 3 blind mice (severe cataracts in both eyes) to assess whether the response to looming stimulus was truly a visually guided behavior. These blind mice had no pupillary light reflex and no optomotor response.
Figure 1: Experimental system.
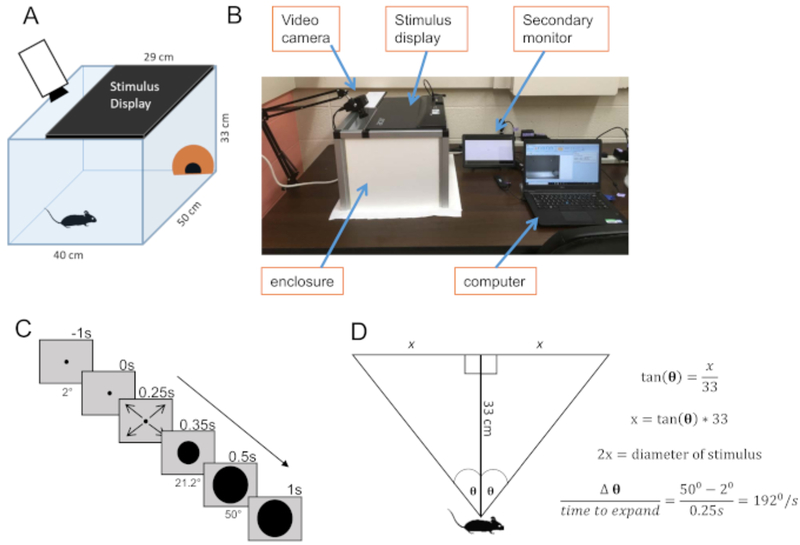
(A) Schematic of the looming stimuli enclosure. A computer monitor (21”) covers the ceiling. There is an opaque dome in one corner of the enclosure in which a mouse may take refuge. A video monitor with a wide-angle lens captures the mouse behavior. (B) Overall view of our entire setup. The secondary monitor duplicates the image showing on the stimulus display. (C) Diagram of the looming stimulus. The looming stimulus begins at 2° (1.15cm) and holds at this size for 250 ms. It then expands to 50° (30.8cm) over the course of 250 ms and remains 50° for an additional 500 ms. This 1s sequence then repeats 9 more times prior to terminating. (D) Diagram of stimulus calculations. The height of the cage dictates the necessary start and end size (in centimeters) of the stimulus in order to produce a stimulus that expands from 2° to 50° when directly above the mouse.
2. Mouse acclimation
Place a mouse in the enclosure and let it freely explore its surroundings. If possible, try to minimize stress during animal transfer by using the back of your free hand as a resting place for the mouse instead of letting it hang without support. The mouse should find the entire enclosure to be safe and should discover the hiding place. Drop a few food pellets in the corner opposite the refuge to encourage the mouse to remain outside the refuge.
Allow the mouse to acclimate anywhere from 7 to 15 min29,39 .We allowed 10 min of acclimation prior to stimulus onset. Furthermore, 10 min acclimation one day prior to the experiment may ease the mice.
3. Looming visual stimuli projection
Prior to inserting the mouse into the arena, make sure the stimulus code is ready to run to facilitate as few lighting changes as possible while the mouse is in the enclosure. Once the software is ready to run, gently place the mouse in the enclosure.
10 seconds prior to the stimulation, start the video capturing.
- Start the looming visual stimuli when the mouse is away from the shelter and moving freely in the open arena. Wait 10 seconds after the last stimulus presentation to terminate the recording.
- Begin the stimulus presentation when the mouse is in the corner farthest from the refuge. However, when mice seem unwilling to explore the far corner, present the stimulus when the mouse is in a different corner of the arena. This does not appear to make a difference in animal behavioral response.
Transfer the mouse back to its original cage. Clean the enclosure for the next mouse by spraying the walls and refuge with 70% ethanol and wiping it down. Replace the paper floor liner if soiled and reposition the refuge to the same initial location if moved during animal transfer and enclosure cleaning.
4. Video analysis
-
Save the video clip for each mouse in .avi format without file compression in order to ensure no data loss during transfer to the analysis software.
NOTE: Lack of compression will lead to larger file size; therefore, use external hard drive for storage.
Use analysis software to track the animal’s motion around the arena prior to, during, and after stimulus presentation. Use commercially available software (see Table of Materials) with a manual tracking ability to track the position of the mouse head in every video frame, which generated X and Y coordination every 1/60 ms. Other motion-tracking software includes FIJI (NIH)40 and EthoVision (Noldus).
Calculate the velocity and the distance of the mouse from the refuge. If the image of the arena is distorted due to the video angle, correct the X and Y coordination prior to the calculation (Figure 2).
Compare the parameters before and after looming stimulus onset to determine how the mouse responded to the stimuli, whether by freezing, fleeing, or demonstrating no change in behavior29. Define freezing as episodes where speed was less than 20 mm/s for 0.5 s or longer. Define flight as episodes where speed increased to 400 mm/s or greater and ended with the mouse in the refuge. Definitions for freezing and flight were based on those set by Franceschi et al.29
Figure 2: Analysis calculations.
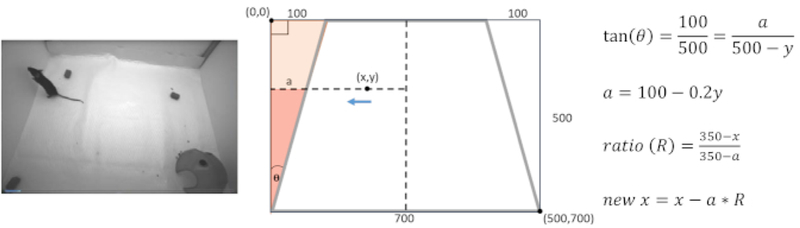
Calculations to correct skew from wide-angle lens. Due to the placement of the camera, the floor of the arena appears as a trapezoid instead of a rectangle (left). Therefore, the X and Y coordinates of the mouse must be corrected to accurately analyze mouse position (mid). Using the geometry of congruent triangles, it is possible to find how much the x-coordinate must shift in order to correctly represent the mouse’s movement in the 3-dimensional space (right).
Representative Results
A mouse with healthy eyes was placed in the enclosure and allowed to acclimate for 10 min. The arena with the monitor on the ceiling was kept under mesopic light conditions (7 × 105 photons/μm2/s). During the acclimation period, the mouse explored the space and found the opaque dome as a refuge. When the mouse moved away from the refuge, video capturing started, followed by initiation of the visual stimulus. In response to the looming stimulus, most mice ran into the dome (flight response), which was observed in 30 out of 31 mice (97%). Some of the mice exhibited freeze responses before they fled (19/31 mice, 61%). The looming stimulus reduced the light condition 1 log (6 × 105 photons/μm2/s).
Captured video clips were analyzed using either a commercial analytics software with a manual tracking function (Image Pro Plus) or FIJI (NIH). Using the tracking feature, the mouse’s position was identified in each frame of the video (60 frames/s) before, during, and after the looming stimuli. We analyzed the velocity changes over time as well as the distance to the shelter (Figure 3). When flight occurred, the velocity abruptly increased and the distance to the shelter reduced accordingly. In contrast, the velocity was near 0 mm/s when mice froze. The latency from the onset of the looming stimuli to flight ranged from 0.1 to 6.0 seconds (average 2.2 s, 30 mice). The range of maximum velocity for flight response was 500–3000 mm/s (30 mice).
Figure 3: Representative responses to looming stimuli.
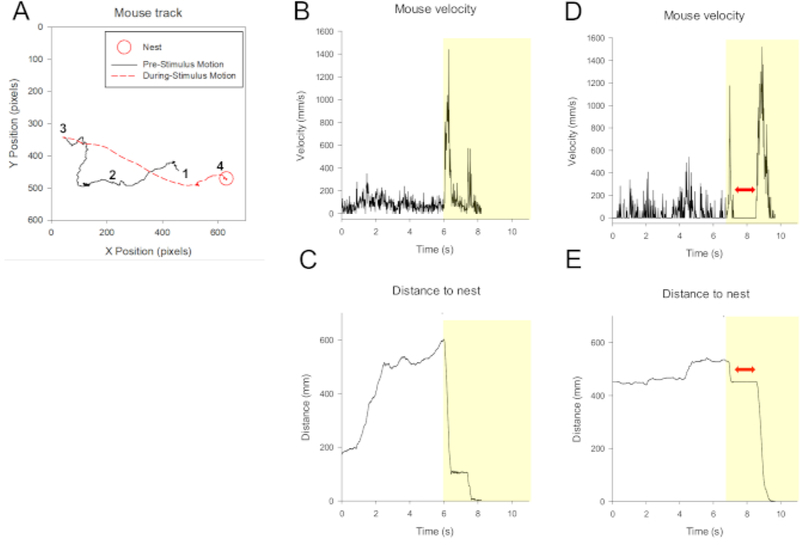
(A) An example of a mouse’s movement tracked within the arena. A red circle shows the dome where the mouse fled to and stayed until the looming disappeared. 1 = mouse position starting point when video capturing started. 2 = movement prior to stimulus onset when the mouse explored the arena. 3 = looming stimulus started. The mouse dashed to the dome (shown by a red dashed line). 4 = the mouse stayed in the dome until and after termination of the stimulus. (B) Velocity changes as a function of time for this mouse. The dotted line indicates when the looming stimulation began. Stimulus duration is indicated by the yellow background. The full 10 second cycle is not shown here since the mice remained stationary in the refuge for the entire stimulus duration. (C) Distance from the dome over time for the same mouse in (A) and (B). (D) and (E) The velocity and distance to the dome for a mouse that exhibited the freeze reaction (freezing duration shown by the red double sided arrow) prior to flight. The velocity was reduced compared to the control (before looming). The distance to the dome did not change during this period.
Discussion
With the looming visual stimuli system, a majority (97%) of healthy eye-mice showed flight response. One of 29 mice did not show an obvious flight response. However, the mouse walked toward the dome and remained near it until looming disappeared, indicating that the mouse was at least cautious when the looming stimuli occurred. Therefore, the looming stimuli consistently elicited innate fear responses in healthy-eyed mice. On the other hand, three blind mice did not show any responses to the looming (preliminary results). Taken together, we demonstrate that looming experiments are a useful and consistent vision test for mice.
We set the speed of the looming stimuli at 192 degree/s. Franceschi et al.29 examined the looming responses at varying speeds, 5 to 84 degree/ s and observed freeze responses preferentially at lower speed levels. Yilmaz and Meister28 observed flight responses at 35 to 350 degree/s; however, flight latency was longer at higher speeds. Therefore, to evoke consistent flight responses, looming should be at a speed of 50 degree/ s or above. Looming stimuli can be generated easily even with standard presentation software. However, such software cannot create higher speeds of looming stimuli. We instead used MatLab and PsychToolbox3 to create the visual stimuli at 192 degree/s.
We acclimated the mice for 10–15 min before the looming stimuli, which is the acclimation time previous researchers described28,29,39 .We furthermore tested whether acclimation the day before changed the looming behavior. We placed the mice in the enclosure for 10 min without looming stimuli the day before the looming stimuli. This acclimation significantly shortened the flight latency (p < 0.01, n = 7 mice, data not shown). Although 10 min of acclimation on the day of looming consistently caused flight responses, 1 day prior acclimation decreased the latency of flight responses.
There are some limitations for using looming stimuli as a vision test. First, it is hard to test one eye at a time. Unless suturing the one eye, both eyes are tested together. Second, the mechanisms of the behavioral looming response have not been fully established. In the retina, Yilmaz and Meister 28 suggested that ventral OFF-DSGCs (direction-selective ganglion cells), but not ON-DSGCs, convey the looming signals to cause responses. This conclusion arose from their results that mice responded to dark looming stimuli, but not to white looming. In the brain, Wei et al.36 and Shang et al.37 demonstrated that the pathways from the superior colliculus through the amygdala and the periaqueductal gray are responsible for the looming. However, more studies should be conducted to confirm these investigations.
Even though some limitations exist with respect to the looming experiment, the looming visual stimulus generates consistent and robust fear response in mice and should be a useful test of mouse vision which requires minimal training of the experimenter.
Supplementary Material
Acknowledgments
This work was supported by NIH R01 EY028915 (TI) and RPB grants.
Footnotes
Disclosures
The authors have nothing to disclose.
Video Link
The video component of this article can be found at https://www.jove.com/video/59766/
References
- 1.Enroth-Cugell C, Robson JG The contrast sensitivity of retinal ganglion cells of the cat. The Journal of Physiology. 187 (3), 517–552 (1966) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Boycott BB, Wassle H The morphological types of ganglion cells of the domestic cat’s retina. The Journal of Physiology. 240 (2), 397–419 (1974) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Livingstone MS, Hubel DH Segregation of form, color, movement, and depth: anatomy, physiology, and perception. Science. 240 (4853), 740–749 (1988) [DOI] [PubMed] [Google Scholar]
- 4.Livingstone MS, Hubel DH Psychophysical evidence for separate channels for the perception of form, color, movement, and depth. The Journal of Neuroscience. 7 (11), 3416–3468 (1987) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wassle H Parallel processing in the mammalian retina. Nature Reviews Neuroscience. 5 (10), 747–757 (2004) [DOI] [PubMed] [Google Scholar]
- 6.Awatramani GB, Slaughter MM Origin of transient and sustained responses in ganglion cells of the retina. The Journal of Neuroscience. 20 (18), 7087–7095 (2000) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ghosh KK, Bujan S, Haverkamp S, Feigenspan A, Wassle H Types of bipolar cells in the mouse retina. The Journal of Comparative Neuroscience. 469 (1), 70–82 (2004) [DOI] [PubMed] [Google Scholar]
- 8.Wässle H, Puller C, Muller F, Haverkamp S Cone contacts, mosaics, and territories of bipolar cells in the mouse retina. The Journal of Neuroscience. 29 (1), 106–117 (2009) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Helmstaedter M, et al. Connectomic reconstruction of the inner plexiform layer in the mouse retina. Nature. 500 (7461), 168–174, (2013) [DOI] [PubMed] [Google Scholar]
- 10.Shekhar K, et al. Comprehensive Classification of Retinal Bipolar Neurons by Single-Cell Transcriptomics. Cell. 166 (5), 1308–1323 e1330, (2016) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wu SM, Gao F, Maple BR Functional architecture of synapses in the inner retina: segregation of visual signals by stratification of bipolar cell axon terminals. The Journal of Neuroscience. 20 (12), 4462–4470 (2000) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sun W, Li N, He S Large-scale morphological survey of mouse retinal ganglion cells. The Journal of Comparative Neuroscience. 451 (2), 115–126 (2002) [DOI] [PubMed] [Google Scholar]
- 13.Volgyi B, Chheda S, Bloomfield SA Tracer coupling patterns of the ganglion cell subtypes in the mouse retina. The Journal of Comparative Neuroscience. 512 (5), 664–687, (2009) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kong JH, Fish DR, Rockhill RL, Masland RH Diversity of ganglion cells in the mouse retina: Unsupervised morphological classification and its limits. The Journal of Comparative Neuroscience. 489 (3), 293–310 (2005) [DOI] [PubMed] [Google Scholar]
- 15.Sumbul U et al. A genetic and computational approach to structurally classify neuronal types. Nature Communications. 5, 3512, (2014) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Baden T et al. The functional diversity of retinal ganglion cells in the mouse. Nature. 529 (7586), 345–350, (2016) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lindstrom SH, Ryan DG, Shi J, DeVries SH Kainate receptor subunit diversity underlying response diversity in retinal Off bipolar cells. The Journal of Physiology. 592 (Pt 7), 1457–1477, (2014) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Euler T, Haverkamp S, Schubert T, Baden T Retinal bipolar cells: elementary building blocks of vision. Nature Reviews Neuroscience. 15 (8), 507–519 (2014) [DOI] [PubMed] [Google Scholar]
- 19.Yoshida K et al. A key role of starburst amacrine cells in originating retinal directional selectivity and optokinetic eye movement. Neuron. 30 (3), 771–780 (2001) [DOI] [PubMed] [Google Scholar]
- 20.Pearson RA et al. Restoration of vision after transplantation of photoreceptors. Nature. 485 (7396), 99–103, (2012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Saszik SM, Robson JG, Frishman LJ The scotopic threshold response of the dark-adapted electroretinogram of the mouse. The Journal of Physiology. 543 (Pt 3), 899–916 (2002) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Reuter JH, Sanyal S Development and degeneration of retina in rds mutant mice: the electroretinogram. Neuroscience Letters. 48 (2), 231–237 (1984) [DOI] [PubMed] [Google Scholar]
- 23.Woodward WR et al. Isoflurane is an effective alternative to ketamine/xylazine/acepromazine as an anesthetic agent for the mouse electroretinogram. Documenta Ophthalmologica. 115 (3), 187–201, (2007) [DOI] [PubMed] [Google Scholar]
- 24.Cahill H, Nathans J The optokinetic reflex as a tool for quantitative analyses of nervous system function in mice: application to genetic and drug-induced variation. PLoS One. 3 (4), e2055, (2008) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Prusky GT, Alam NM, Beekman S, Douglas RM Rapid quantification of adult and developing mouse spatial vision using a virtual optomotor system. Investigative Ophthalmology & Visual Science. 45 (12), 4611–4616, (2004) [DOI] [PubMed] [Google Scholar]
- 26.Lu Q, Ganjawala TH, Hattar S, Abrams GW, Pan ZH A Robust Optomotor Assay for Assessing the Efficacy of Optogenetic Tools for Vision Restoration. Investigative Ophthalmology & Visual Science. 59 (3), 1288–1294, (2018) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Xue T et al. Melanopsin signalling in mammalian iris and retina. Nature. 479 (7371), 67–73, (2011) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yilmaz M, Meister M Rapid innate defensive responses of mice to looming visual stimuli. Current Biology. 23 (20), 2011–2015, (2013) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.De Franceschi G, Vivattanasarn T, Saleem AB, Solomon SG Vision Guides Selection of Freeze or Flight Defense Strategies in Mice. Current Biology. 26 (16), 2150–2154, (2016) [DOI] [PubMed] [Google Scholar]
- 30.Temizer I, Donovan JC, Baier H, Semmelhack JL A Visual Pathway for Looming-Evoked Escape in Larval Zebrafish. Current Biology. 25 (14), 1823–1834, (2015) [DOI] [PubMed] [Google Scholar]
- 31.Guest BB, Gray JR Responses of a looming-sensitive neuron to compound and paired object approaches. Journal of Neurophysiology. 95 (3), 1428–1441, (2006) [DOI] [PubMed] [Google Scholar]
- 32.McMillan GA, Gray JR A looming-sensitive pathway responds to changes in the trajectory of object motion. Journal of Neurophysiology. 108 (4), 1052–1068, (2012) [DOI] [PubMed] [Google Scholar]
- 33.Vagnoni E, Lourenco SF, Longo MR Threat modulates neural responses to looming visual stimuli. Eur The Journal of Neuroscience. 42 (5), 2190–2202, (2015) [DOI] [PubMed] [Google Scholar]
- 34.Coker-Appiah DS et al. Looming animate and inanimate threats: the response of the amygdala and periaqueductal gray. Social Neuroscience. 8 (6), 621–630, (2013) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tyll S et al. Neural basis of multisensory looming signals. Neuroimage. 65, 13–22, (2013) [DOI] [PubMed] [Google Scholar]
- 36.Wei P et al. Processing of visually evoked innate fear by a non-canonical thalamic pathway. Nature Communications. 6, 6756, (2015) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Shang C et al. Divergent midbrain circuits orchestrate escape and freezing responses to looming stimuli in mice. Nature Communications. 9 (1), 1232, (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Salay LD, Ishiko N, Huberman AD A midline thalamic circuit determines reactions to visual threat. Nature. 557 (7704), 183–189,(2018) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Vale R, Evans D, Branco T A Behavioral Assay for Investigating the Role of Spatial Memory During Instinctive Defense in Mice. Journal of Visualized Experiments. (137), (2018) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tungtur SK, Nishimune N, Radel J, Nishimune H Mouse Behavior Tracker: An economical method for tracking behavior in home cages. Biotechniques. 63 (5), 215–220, (2017) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.