

Abstract
Despite the recent malaria burden reduction in the Americas, focal transmission persists across the Amazon Basin. Timely analysis of surveillance data is crucial to characterize high-risk individuals and households for better targeting of regional elimination efforts. Here we analyzed 5,480 records of laboratory-confirmed clinical malaria episodes combined with demographic and socioeconomic information to identify risk factors for elevated malaria incidence in Mâncio Lima, the main urban transmission hotspot of Brazil. Overdispersed malaria count data clustered into households were fitted with random-effects zero-inflated negative binomial regression models. Random-effect predictors were used to characterize the spatial heterogeneity in malaria risk at the household level. Adult males were identified as the population stratum at greatest risk, likely due to increased occupational exposure away of the town. However, poor housing and residence in the less urbanized periphery of the town were also found to be key predictors of malaria risk, consistent with a substantial local transmission. Two thirds of the 8,878 urban residents remained uninfected after 23,975 person-years of follow-up. Importantly, we estimated that nearly 14% of them, mostly children and older adults living in the central urban hub, were free of malaria risk, being either unexposed, naturally unsusceptible, or immune to infection. We conclude that statistical modeling of routinely collected, but often neglected, malaria surveillance data can be explored to characterize drivers of transmission heterogeneity at the community level and provide evidence for the rational deployment of control interventions.
Introduction
Malaria continues to be a major cause of morbidity and mortality in sub-Saharan Africa, South and Southeast Asia, Oceania, and Latin America, with 219 million cases and 435,000 deaths worldwide in 2017 [1]. The disease typically affects the rural poor, since urbanization tends to reduce malaria risk through improved housing, greater access to health services, and environmental changes that may limit vector abundance [2]. Indeed, malaria rates are typically lower in cities, compared to their rural surroundings, in most [3,4], although not all [5], endemic settings. Despite this, the rapidly growing urban population in developing countries bears an increasingly larger proportion of the global malaria burden because of both local transmission and importation from rural sites [6,7].
Sprawling towns and cities are heterogeneous, and so is urban malaria risk. For example, in the early 1980s the number of infective bites per person was estimated to range between <1 every three years and >100 per year across the city of Brazzaville [8]. Risk heterogeneity translates into overdispersed frequency distributions of malaria episodes per person, with few subjects experiencing a disproportionately large disease burden due to frequent reinfection [9, 10]. Low socioeconomic status, poor housing quality, and proximity to larval habitats are among the household-level factors that fuel malaria transmission in urban Africa [11,12]. Travel to rural communities is another major risk factor that entails a different set of preventive measures [13].
Surprisingly, the epidemiology of urban malaria has been little investigated in Latin America, the most urbanized region of the developing world [7,14]. Imported cases from surrounding rural sites appear to be the main contributors to malaria infections diagnosed in the city of Quibdó, Colombia [15,16], but urban malaria transmission has been documented in coastal Peru [17] and in the outskirts of major cities in Amazonian Brazil [18,19]. Anopheles darlingi, the main malaria vector in the Amazon Basin, is typically found in forested areas [20], but urban environments are not necessarily unsuitable for this species. Indeed, the accelerated urbanization process in the Amazon over the past few decades originated a multitude of small cities and towns characterized by poor housing and little public infrastructure and interspersed with rural spaces. Unsurprisingly, immature stages of malaria vectors can develop in many types of natural and man-made water habitats in precarious urban and peri-urban settlements across the region, including the fish farming ponds recently opened for commercial aquaculture [21–23].
Statistical modeling of routinely collected malaria surveillance data can be particularly challenging. Poisson regression models are commonly used to analyze count-type data in epidemiology, but cannot adequately fit overdispersed malaria case distributions that are typically found in endemic settings [9,10]. A variety of alternative models have been used instead, e.g. the negative binomial (NB) [9]. However, as malaria rates decline, more subjects will remain uninfected over extended periods of time, increasing the proportion of zero counts in cohort studies. Zero-inflated statistical models, such as the zero-inflated negative binomial (ZINB), usually provide a better fitting to malaria count data [10,24] and household-level malaria vector densities [25] with an excess of zero counts. The ZINB model combines the NB distribution and the logit distribution. As a consequence, it can additionally estimate the fraction of unexposed or protected individuals in the population by allowing for a mixture of two latent classes: (i) at-risk individuals who contribute cases according to the NB distribution function and (ii) not-at-risk individuals with zero cases, described by the logit component, hence termed “structural zeros”. The not-at-risk fraction of the population described by the structural zero component of the model is intrinsically free of any malaria risk and will remain uninfected irrespective of any protective measure. Importantly, this subpopulation can bias estimates of the effect of interventions for controlling and eliminating malaria [10]. A further challenge for statistical modeling of malaria surveillance data is the clustering of observations into households, where key risk factors for infection such as poor housing quality and proximity to mosquito breeding sites are shared [11,12]. Random-effects (RE-) ZINB models can account for the dependency between observations [26] but, surprisingly, have not yet been used to analyze data from community-wide malaria surveys.
Despite the dramatic decrease in the burden of malaria in Brazil in recent decades, focal transmission persists across the Amazon Basin [27]. Transmission rates are greatest in Juruá Valley, next to the Brazil-Peru border. With 0.5% of the Amazon's population, the region accounts for 18.5% of the country's malaria burden, estimated at 157,000 cases in 2016 [1]. A large proportion of infections in Juruá Valley are reportedly acquired in urban settings–up to 45% in the municipality of Mâncio Lima, compared with the country's average of 17% in 2013 [28]. Here, we characterize high-risk individuals and households by applying RE-ZINB regression analysis to overdispersed and household-clustered surveillance data. Our findings may allow for better targeting of interventions in the main malaria hotspot of Brazil.
Material and methods
Ethics statement
The study protocol was approved by the Institutional Review Board of the Institute of Biomedical Sciences, University of São Paulo, Brazil (CEPH-ICB 1368/17); written informed consent and assent were obtained.
Study area and population
The municipality of Mâncio Lima covers a surface area of 4,672 km² in northwestern Brazil (S1 Fig) and comprises a single town next to the Japiim river, where nearly half of its 17,545 inhabitants reside. Streams, wetlands rich in moriche palm trees, and fish farming ponds are widespread in the town. With a typical equatorial humid climate, Mâncio Lima receives most rainfall between November and April, but malaria transmission occurs year-round. The annual parasite incidence, estimated at 436.4 cases per 1,000 inhabitants in 2016, is the highest for a municipality in Brazil [29]. Local distribution of long-lasting insecticidal bed nets (LLINs) and indoor residual spraying (IRS) with pyrethroids or propoxur are currently limited to high-risk households. The primary local malaria vector is An. darlingi, but An. albitarsis s.l. is also abundant across the town, especially in fish farming ponds [22,30].
The study population comprised all permanent residents in the town of Mâncio Lima enumerated by a census survey between November 2015 and April 2016. During the survey, dwellings were geo-localized and a questionnaire was applied to collect demographic, health, behavioral, and socioeconomic data. Principal component analysis was used to compute an assets-based wealth index for each household [31].
Malaria surveillance and treatment
The study outcome was laboratory-confirmed malaria, defined as any episode of parasitemia, irrespective of parasite density or symptoms, diagnosed through active or passive case detection from 1 January 2014 through 30 September 2016. We retrieved malaria case records from the electronic malaria notification system of the Ministry of Health of Brazil (http://200.214.130.44/sivep_malaria/). Because malaria is a notifiable disease in Brazil and diagnostic testing and treatment are not available outside the network of government-run health care facilities, the database comprises the vast majority of malaria episodes confirmed by thick-smear microscopy in Mâncio Lima residents over the study period (33 months). According to a recent estimate, the electronic malaria notification system comprises 99.6% of all clinical malaria cases diagnosed countrywide [32]. At least 100 fields are routinely examined for malaria parasites under 1000× magnification by experienced local microscopists before a slide is declared negative. Partially supervised chloroquine-primaquine and artemether-lumefantrine regimes were administered to treat Plasmodium vivax and P. falciparum malaria, respectively [33]. A minimal interval of 28 days between two consecutive episodes was required to count the second episode as a new malaria infection; when different species were diagnosed <28 days apart, a single mixed-species infection was counted.
Statistical methods
The R package gamlss [34] was used for statistical analysis (R Foundation for Statistical Computing, Vienna, Austria). The generalized additive models for location, scale and shape (GAMLSS) approach [35] was used to fit ZINB [10,24] distribution functions to malaria counts and to choose the best-fitting model. We note that the term “additive” refers to the option, provided by the GAMLSS approach but not applied here, to include nonparametric components into the linear predictors of the models. We used randomized normal quantile-quantile (Q-Q) plots and detrended normal Q-Q plots, known as worm plots, as diagnostic tools to analyze residuals [36].
Individual- and household-level explanatory variables were added to the count component of the first standard ZINB regression model. The individual-level variables entered in the multivariable models were: age (stratified as 0 [birth]-5, 6–15, 16–40, 41–60, and >60 years); sex (female vs. male); reported bed net use, either insecticide-impregnated or not, the previous night (no vs. yes); sleeping time (before 10 pm, between 10 and 11 pm, after 11 pm); and wake-up time (before 7 am, between 7 and 8 am, after 8 am). Household-level variables were: household size (<5 vs. ≥5 people); wealth index (stratified into terciles); LLIN available in the household (no, yes, unknown); IRS within the past three years (no, yes, unknown); and housing quality indicators such as incomplete walls and ceiling (no vs. yes), presence of screens in doors and windows (no vs. yes), and type of lavatory (indoors vs. outhouse). We used the R package GoodmanKruskal to identify significant pairwise associations between model covariates; none was found (S2 File). The multivariable model was adjusted for the covariate “follow-up duration”, the number of person-years at risk contributed by each study participant. This was calculated for the period between the date of birth or 1 January 2014, whichever was the most recent, and 30 September 2016, when the follow-up ended.
Next, to account for clustering of observations into households, household-level RE terms were also considered into the multivariable ZINB regression. Worm plot diagnostic of the RE-ZINB model indicated too large fitted variance, with many data points lying outside the 95% confidence intervals (CI) of the expected deviation. To reach satisfactory model diagnostics, we shrunk the random-effects distribution toward the overall mean [37] by decreasing the degrees of freedom originally estimated by the model to 150; further details are provided in S1 File.
We next used the random-effect predictors to characterize the spatial heterogeneity in malaria risk while controlling for potential confounders [26]. The high (low) magnitude of household random-effects predictors was used to select households with higher (lower) than average malaria incidence density. We examined the spatial distribution of households with the top 5% and bottom 5% random-effects predictors of the RE-ZINB models (here termed “hot houses” and “cold houses”, respectively) by mapping their GPS coordinates.
Given the results of the spatial analysis described above, we tested whether model fitting could be further improved by including a variable describing subjects' zone of residence, whether in the center (“urban hub”) or in the less-urbanized periphery of the town, close to the most vegetated areas. To this end, geo-localized houses were classified as centrally or peripherally situated using the computational approach described in S3 File. We next used the Akaike information criterion (AIC) to compare the quality of RE-ZINB models with and without the covariate “zone of residence”.
To further characterize study participants at no risk of malaria [10], we built additional RE-ZINB models with the following variables added to the structural zero component: zone of residence, age, sex, and follow-up duration. The following variables were initially entered in the count component: age, sex, bed net use, follow-up duration, zone of residence, household size, LLIN availability, recent IRS, presence of complete walls, and type of lavatory.
The best RE-ZINB models were selected using the strategy stepGAICALL.A() proposed by Stasinopoulos and colleagues [34] with the following steps: (a) an initial NB model was built for the count component (forward approach); (b) given this model, a model was built for the logit component (forward approach); (c) given the NB and logit models, we checked whether the terms for the logit model were needed using backward elimination; (d) given the NB and logit models, we checked whether the terms for the NB model were needed (backward elimination). The generalized AIC (GAIC) was used for model comparison.
Results
The study comprised 8,878 subjects with ages ranging between <1 month and 105 years (mean, 27.0 years) distributed into 2,329 households. They experienced a total of 5,480 laboratory-confirmed malaria episodes over 23,975.3 person-years of follow-up, with an overall malaria incidence density estimated at 22.6 cases per 100 person-years at risk. Plasmodium vivax accounted for 84.2% of the episodes (incidence density, 19.0 cases per 100 person-years at risk); 14.4% of the infections were due to P. falciparum, (incidence density, 3.2 cases per 100 person-years at risk), and 1.4% due to both species. The incidence densities were lowest among under-five children and over-sixty adults (Fig 1A), mostly due to the age-related variation in P. vivax incidence (Fig 1B). This age-incidence pattern likely reflects the combined effect of differential exposure and acquired immunity across age groups. Male adults aged 16–60 years were more often infected than their female counterparts (Fig 1A), consistent with increased occupational exposure.
Fig 1. Age-related malaria incidence distribution in urban Mâncio Lima, northwestern Brazil.

(A) Age- and gender-related malaria incidence density, regardless of the infecting parasite species. (B) Species-specific age-related malaria incidence density.
Statistical model fitting
The frequency distribution of malaria cases was overdispersed, with a mean of 0.62 (range, 0 to 12; variance, 1.4) episodes per person. The vast majority (67.4%) of study participants remained free of malaria and less than 1% of them had six or more repeated episodes during the follow-up. Empirical frequency distribution data were properly fitted with ZINB distributions (Fig 2).
Fig 2. Zero-inflated negative binomial (ZINB) model fit to malaria episode counts per person in urban Mâncio Lima, northwestern Brazil.
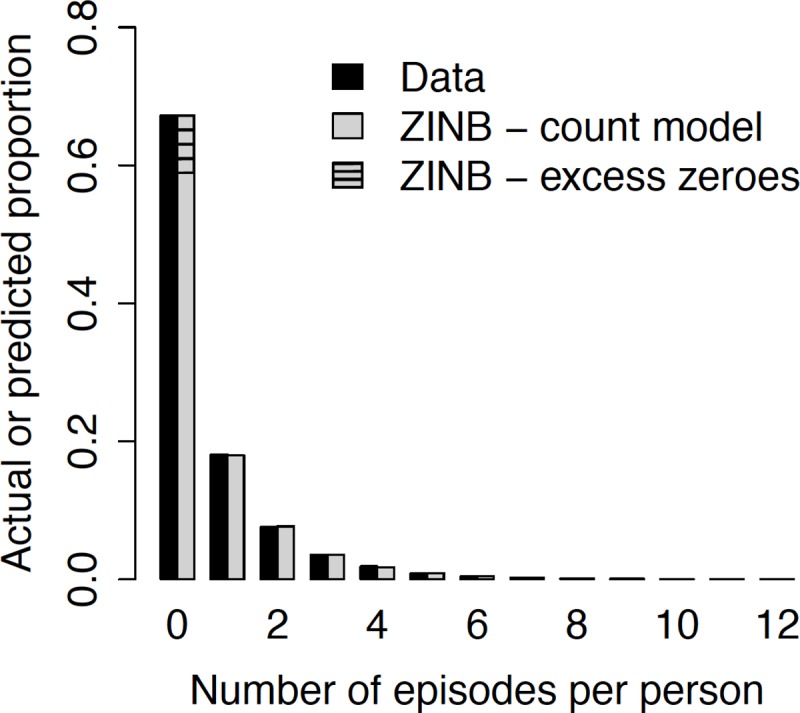
We analyzed data from 8,431 individuals (447 were excluded due to missing values in key variables) and the RE-ZINB count regression model obtained comprises the explanatory variables listed in S1 Table. RE-ZINB regression analysis estimated that 13.6% (95% CI, 5.1–31.3%) of the study participants (roughly 1,200 residents) were intrinsically free of malaria risk and accounted for the excess zero counts beyond the NB expectations.
We next examined the spatial distribution of “hot houses” and “cold houses”. These were defined as the households within the top 5% (hot houses) and the bottom 5% (cold houses) estimates of random-effects predictors for the count compartment of the RE-ZINB regression model, adjusted for all explanatory variables shown in S1 Table. We show that most hot houses are indeed situated in the periphery of the town (Fig 3) and, therefore, geo-localized houses were classified as centrally or peripherally situated using the computational method described in S3 File. The covariate indicating the zone of residence (whether in the center or in the less-urbanized periphery of the town) was introduced to the regression and the RE-ZINB model fitting was improved (Table 1). These results further indicate that households in the less-urbanized periphery of the town, surrounded by more densely vegetated areas, constitute the priority target for spatial interventions aimed to reduce local malaria transmission.
Fig 3. Spatial distribution of households and malaria incidence in urban Mâncio Lima, northwestern Brazil.

Table 1. Degrees of freedom and Akaike information criterion (AIC) values for the RE-ZINB regression models fitted to empirical data.
| Regression model | Zone of residence | Degrees of freedom | AIC | |
|---|---|---|---|---|
| Fixed effects | Random effects | |||
| Random-effects ZINB | No | 17 | 150 | 17319.74 |
| Yes | 18 | 150 | 17225.61 | |
Study households with lower-than-average (“cold houses”) and higher-than-average malaria incidence (“hot houses”) were identified using the random-effect predictors from the zero-inflated negative binomial (RE-ZINB) model. Red dots show “hot houses” with the top 5% random-effect predictors and blue dots show “cold houses” with the bottom 5% random-effect predictors of RE-ZINB model; all other households are represented as grey dots. Vegetated areas (data retrieved from Brazilian Institute for Space Research (2018) PRODES Project, http://www.inpe.br/cra/projetos_pesquisas/terraclass2014.php.) are shown in green and roads and streets (data retrieved under the Open Database License from the Open Street Map Foundation website at https://www.openstreetmap.org/#map=13/-7.6220/-72.8960&layers=HNas) are shown as thin black lines. Figure created with the QGIS software version 3.4.3, an open source Geographic Information System (GIS) licensed under the GNU General Public License (https://qgis.org/en/site/about/index.html).
Predictors of malaria incidence density
Table 2 shows independent associations between explanatory variables and malaria incidence density revealed by the best-fitting multivariable ZINB regression model with RE estimators, which include zone of residence as a covariate. We note that the count compartment of the ZINB model allows for identifying predictors of malaria incidence density in the at-risk fraction (86.4%) of the population. Age between 6 and 60 years, male sex, residence in the less-urbanized periphery, and indicators of poor housing quality were key predictors of increased malaria incidence density in the community (Table 2). It is not surprising that LLIN availability in the household, reported bed net use, and recent IRS were all positively associated with malaria incidence density, given that households perceived to be at increased malaria risk are selectively targeted for LLIN distribution and IRS.
Table 2. Independent predictors of malaria incidence density in urban Mâncio Lima, Brazil, identified by multivariable random-effects zero-inflated negative binomial (RE-ZINB) regression analysis.
| RE-ZINB model estimates, count compartment | |||||
|---|---|---|---|---|---|
| Variable | No. subjects | IRR | 95% CI | P-value | |
| Individual-level | |||||
| Age (years) | |||||
| 0–5 | 1002 | Ref. | |||
| 6–15 | 1973 | 1.29 | 1.10 | 1.51 | 0.0017 |
| 16–40 | 3545 | 1.51 | 1.30 | 1.75 | <0.0001 |
| 41–60 | 1262 | 1.35 | 1.14 | 1.60 | 0.0006 |
| >60 | 649 | 0.74 | 0.59 | 0.91 | 0.0053 |
| Gender | |||||
| Male | 4184 | Ref. | |||
| Female | 4247 | 0.79 | 0.74 | 0.85 | <0.0001 |
| Bed net use the previous night | |||||
| No | 2469 | Ref. | |||
| Yes | 5962 | 1.10 | 1.01 | 1.21 | 0.0328 |
| Follow-up duration | |||||
| 8431 | 3.83 | 2.71 | 5.40 | <0.0001 | |
| Household-level | |||||
| Zone of residence | |||||
| Center | 5296 | Ref. | |||
| Periphery | 3135 | 1.56 | 1.44 | 1.68 | <0.0001 |
| Household size | |||||
| ≤ 5 | 4524 | Ref. | |||
| > 5 | 3907 | 1.11 | 1.03 | 1.20 | 0.0084 |
| LLIN available | |||||
| No | 2870 | Ref. | |||
| Yes | 3057 | 1.11 | 1.02 | 1.22 | 0.0196 |
| Unknown | 2504 | 0.95 | 0.86 | 1.05 | 0.2857 |
| Recent IRS | |||||
| No | 1520 | Ref. | |||
| Yes | 1497 | 1.23 | 1.09 | 1.39 | 0.0012 |
| Unknown | 5414 | 1.00 | 0.90 | 1.11 | 0.9724 |
| Complete walls | |||||
| No | 22 | Ref. | |||
| Yes | 8409 | 0.28 | 0.16 | 0.49 | <0.0001 |
| Type of lavatory | |||||
| Outhouse | 4657 | Ref. | |||
| Indoors | 3774 | 0.86 | 0.79 | 0.93 | 0.0002 |
Abbreviations: RE-ZINB, random-effects zero-inflated negative binomial; IRR, incidence rate ratio; CI, confidence interval; LLIN, long-lasting insecticidal bed net; IRS, indoor residual spraying.
To further characterize high-risk study participants, we tested whether their increased malaria incidence density was due to larger proportions of subjects experiencing at least one malaria episode or to an increased number of repeated malaria episodes (that may include parasite recrudescences and relapses in addition to new infections) among those who had malaria episodes recorded during the study. We found that both factors contribute to the increased malaria incidence density observed in high-risk population strata. Indeed, 742 (42.5%) of 1,746 male study participants aged 16–40 years, but only 2,020 (30.2%) of the remaining 6,685 study participants, had at least one malaria episode during the 33-month follow-up (P < 0.0001, χ2 = 94.78, 1 degree of freedom). Moreover, 1,263 (40.3%) of 3,135 study participants living in the periphery of Mâncio Lima, compared to 1,499 (28.3%) of the 5,296 individuals living in the central area of the town, experienced at least one malaria episode during the follow-up (P < 0.0001, χ2 = 128.36). However, once infected high-risk subjects were also more likely to have repeated malaria episodes during the follow-up. In fact, the frequency distributions of malaria episodes in male study participants aged 16–40 years and those living in the periphery were significantly shifted to the right, compared to their respective counterparts (S2 Fig.).
Not-at-risk subjects
The not-at-risk fraction of the population described by the structural zero compartment of the RE-ZINB model may be either unexposed, naturally unsusceptible to infection, or may have acquired immunity over time. Because our explanatory variables did not directly measure natural susceptibility or acquired immunity, we focus further analyses on age, sex and zone of residence as proxies of exposure. These variables were added to the logistic (structural zero) component of the RE-ZINB model, which was further adjusted for follow-up duration (person-years at risk). The best-fitting RE-ZINB regression model revealed a negative association of age between 16 and 40 years (but not sex) and residence in the periphery of the town with the odds of being a structural zero. Interestingly age > 60 years (a proxy of cumulative exposure and acquired immunity) remained as a significant predictor of decreased malaria incidence density, but not of being a structural zero (Table 3). This indicates that age-related and spatial differences in malaria exposure, rather than acquired immunity, can account, at least in part, for the presence of not-at-risk subjects in the community. Overall, associations between covariates and malaria incidence density identified by the NB compartment of the RE-ZINB model that also included covariates in the logit compartment (Table 3) were similar to those identified by the RE-ZINB model with an empty (i.e., no covariates added) logit compartment (Table 2).
Table 3. Independent predictors of malaria incidence density and odds of being at no risk of malaria in urban Mâncio Lima, Brazil, identified by multivariable random-effects zero-inflated negative binomial (RE-ZINB) regression analysis with explanatory added to the structural zero component of the model.
| Count model | Structural zero | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | No. subjects | IRR | 95% CI | P-value | OR | 95% CI | P-value | ||
| Individual-level | |||||||||
| Age (years) | |||||||||
| 0–5 | 1002 | Ref. | Ref. | ||||||
| 6–15 | 1973 | 1.16 | 0.93 | 1.46 | 0.1892 | 0.68 | 0.39 | 1.20 | 0.1831 |
| 16–40 | 3545 | 1.16 | 0.94 | 1.44 | 0.1658 | 0.31 | 0.15 | 0.64 | 0.0015 |
| 41–60 | 1262 | 1.15 | 0.90 | 1.47 | 0.2567 | 0.55 | 0.28 | 1.06 | 0.0745 |
| >60 | 649 | 0.63 | 0.45 | 0.89 | 0.0084 | 0.56 | 0.19 | 1.69 | 0.3046 |
| Gender | |||||||||
| Male | 4184 | Ref. | |||||||
| Female | 4247 | 0.78 | 0.73 | 0.84 | <0.0001 | ||||
| Bed net use the previous night | |||||||||
| No | 2469 | Ref. | |||||||
| Yes | 5962 | 1.11 | 1.01 | 1.21 | 0.0265 | ||||
| Follow-up duration | |||||||||
| 8431 | - | - | - | - | 0.13 | 0.07 | 0.24 | <0.0001 | |
| Household-level | |||||||||
| Zone of residence | |||||||||
| Center | 5296 | Ref. | Ref. | ||||||
| Periphery | 3135 | 1.39 | 1.25 | 1.54 | <0.0001 | 0.56 | 0.38 | 0.82 | 0.0031 |
| Household size | |||||||||
| ≤ 5 | 4524 | Ref. | |||||||
| > 5 | 3907 | 1.10 | 1.02 | 1.19 | 0.0145 | ||||
| LLIN available | |||||||||
| No | 2870 | Ref. | |||||||
| Yes | 3057 | 1.11 | 1.01 | 1.21 | 0.0276 | ||||
| Unknown | 2504 | 0.96 | 0.87 | 1.06 | 0.4064 | ||||
| Recent IRS | |||||||||
| No | 1520 | Ref. | |||||||
| Yes | 1497 | 1.22 | 1.08 | 1.38 | 0.0015 | ||||
| Unknown | 5414 | 0.99 | 0.90 | 1.10 | 0.8864 | ||||
| Complete walls | |||||||||
| No | 22 | Ref. | |||||||
| Yes | 8409 | 0.29 | 0.16 | 0.51 | <0.0001 | ||||
| Type of lavatory | |||||||||
| Outhouse | 4657 | Ref. | |||||||
| Indoors | 3774 | 0.87 | 0.80 | 0.94 | 0.0003 | ||||
Abbreviations: RE-ZINB, random-effects zero-inflated negative binomial; IRR, incidence rate ratio; CI, confidence interval; OR, odds ratio; LLIN, long-lasting insecticidal bed net; IRS, indoor residual spraying.
Discussion
The long-standing consensus that malaria transmission is spatially heterogeneous provides the basis for targeting control interventions in elimination settings [38,39]. Residual malaria transmission clusters at different spatial scales, from regions to households [40–42], with specific high-risk groups termed “hot-pops” being disproportionally affected [40]. Identifying hot-pops is a top priority of malaria elimination programs.
Here, we examine the drivers of small-area variation in malaria rates in the main urban hotspot in Brazil by fitting multivariable RE-ZINB regression models to community-wide surveillance data. We show that RE-ZINB models can: (i) properly fit overdispersed malaria count data and identify hot pops, (ii) characterize spatial heterogeneity in malaria risk while controlling for potential confounders and identify hot houses, and (iii) characterize the not-at-risk fraction of the population.
Results suggest both imported and locally acquired infections contribute to the malaria burden in the study population. Each entails different malaria control interventions. We hypothesize that increased occupational exposure characterizes the main malaria hot-pop in the community, comprised of adult male residents often engaged in subsistence farming in nearby settlements [43]. These subjects may serve as a source of new parasite strains continuously introduced in the town, being the main targets of interventions to reduce malaria importation. Control measures may include deploying periodic malaria screening and treatment, as well as LLINs, to the most mobile subjects in the community. Conversely, the RE-ZINB model estimates that 14% of the study participants comprises the not-at-risk fraction of the population. This relatively large fraction of the urban population is mostly comprised of children and older adults living in the central urban hub who will remain uninfected regardless of any malaria control measure.
Local transmission also appears to contribute to malaria risk, especially in the less-urbanized periphery. We confirm that better housing is associated with reduced malaria incidence [44,45] even in an endemic setting dominated by vectors that feed and rest predominantly outdoors [46]. Interestingly, hot houses identified by the analysis of random-effects predictors of the RE-ZINB regression model tend to be peripherally located, but they do not form clear, easily detectable clusters. Importantly, the fraction of study participants residing along the town boundaries (37% of the total) appear to be at increased risk after controlling for potential confounders, indicating that the association between place of residence and malaria risk is mostly spatial, and is not severely confounded by age, sex, and housing quality differences among households. These findings are consistent with focal malaria transmission across the urban-rural transition in the periphery of the town [43]. Control measures to reduce local malaria transmission include, among others, IRS and LLIN distribution targeted at hot houses. Moreover, large-scale screening of windows and other house openings may represent a valuable measure to render high-risk hot houses mosquito-proof, as suggested by recent data from urban Africa [47].
The present study has some limitations. First, surveillance data were retrieved retrospectively from a case notification database and no blood samples were available for further confirmatory (e.g., molecular) diagnostic tests. We assume that nearly all malaria episodes diagnosed by microscopy and treated in study participants were retrieved [32], but routine surveillance overlooks transient sub-microscopic parasitemias that do not develop into detectable infections but remain infectious to mosquitoes [48]. Therefore, risk factors described for microscopy-positive malaria in the community are not necessarily the same for sub-microscopic, often asymptomatic infections. Next, surveillance data comprises cases diagnosed by both passive and active case detection, but our data set does not allow for distinguishing between case-finding strategies. Moreover, analyses of passively detected cases are prone to biases due to variation in access to health facilities and health-seeking behavior, even in relatively compact urban areas where health facilities are readily accessible and provide care at no cost. Finally, the infrequency of P. falciparum malaria precludes further between-species comparisons of risk factors in the study population.
Conclusion
We conclude that both local transmission and imported cases from rural and/or forest areas are responsible for the maintenance of malaria in the urban setting of Mâncio Lima. Large sets of routinely collected surveillance data linked to additional demographic and socioeconomic information can be explored for evidence-based planning and deployment of malaria control interventions.
Supporting information
Map of South America showing the location of the field site, the municipality of Mâncio Lima in the state of Acre, northwestern Brazil, next to the border with Peru. Figure created using data extracted from the GADM database (www.gadm.org), version 2.8, under the Creative Commons Attribution License (CCAL) CC BY 4.0 (http://creativecommons.org/licenses/by/4.0/).
(TIF)
Only study participants who had at least one malaria episode diagnosed during the follow-up are included in this analysis. The upper panel shows the frequency distributions of malaria episodes in males aged 16–40 years (A; n = 742 study participants) and in all other population strata in Mâncio Lima (B; n = 2,020). The frequency distributions are significantly different (Kolmogorov-Smirnov test, P = 0.0219). The lower panel shows the frequency distributions of malaria episodes in study participants living in the periphery (C; n = 1,263) and in the center (D; n = 1,499) of the town of Mâncio Lima. The frequency distributions are also significantly different (Kolmogorov-Smirnov test, P < 0.0001).
(TIF)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
Acknowledgments
We thank Odaílton A. Nery and Dr. Nathália F. Lima for logistic support, Maria José Menezes for administrative support, Dr. Igor C. Johansen (University of São Paulo, Brazil) for figure drawing, and Prof. Mikis D. Stasinopoulos and Prof. Robert A. Rigby (London Metropolitan University, UK) for helpful discussions.
Abbreviations
- AIC
Akaike information criterion; CI, confidence interval
- GAIC
generalized Akaike information criterion
- GAMLSS
generalized additive models for location, scale and shape
- IRR
incidence rate ratio
- IRS
indoor residual spraying
- LLIN
long-lasting insecticidal bed net
- OR
odds ratio
- Q-Q plot
quantile-quantile plot
- RE
random effects
- ZINB
zero-inflated negative binomial
- RE-ZINB
random-effects zero-inflated negative binomial
Data Availability
Original data files contain information that may potentially lead to the identification of study participants. Requests for access to original data should be submitted to the Institutional Review Board of the Institute of Biomedical Sciences, University of São Paulo, Brazil, at cep@icb.usp.br. The study identification number is CEPSH 1368.
Funding Statement
This work was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP; http://www.fapesp.br/en/), Brazil (grant 2016/18740-9), and by the National Institute of Allergy and Infectious Diseases, National Institutes of Health, (https://www.niaid.nih.gov/), United States of America (grant U19 AI089681). RMC receives a doctoral fellowship from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; http://cnpq.br/), which also provides senior research scholarships to GAP and MUF. FAPESP (2018/18557-5) provides a doctoral fellowship to AP. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.World Health Organization. World Malaria Report 2018. Geneva: World Health Organization, 2018. [Google Scholar]
- 2.Hay SI, Guerra CA, Tatem AJ, Atkinson PM, Snow RW. Urbanization, malaria transmission and disease burden in Africa. Nat Rev Microbiol. 2005;3:81–90. 10.1038/nrmicro1069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Trape JF. Malaria and urbanization in central Africa: the example of Brazzaville. Part IV. Parasitological and serological surveys in urban and surrounding rural areas. Trans R Soc Trop Med Hyg. 1987;81 Suppl 2:26–33. [DOI] [PubMed] [Google Scholar]
- 4.Robert V, Macintyre K, Keating J, Trape JF, Duchemin JB, Warren M, et al. Malaria transmission in urban sub-Saharan Africa. Am J Trop Med Hyg. 2003;68:169–76. [PubMed] [Google Scholar]
- 5.Qi Q, Guerra CA, Moyes CL, Elyazar IR, Gething PW, Hay SI, et al. The effects of urbanization on global Plasmodium vivax malaria transmission. Malar J. 2012;11:403 10.1186/1475-2875-11-403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Keiser J, Utzinger J, Caldas de Castro M, Smith TA, Tanner M, Singer BH. Urbanization in sub-Saharan Africa and implication for malaria control. Am J Trop Med Hyg. 2004;71(2 Suppl):118–27. [PubMed] [Google Scholar]
- 7.Wilson ML, Krogstad DJ, Arinaitwe E, Arevalo-Herrera M, Chery L, Ferreira MU et al. Urban malaria: Understanding its epidemiology, ecology, and transmission across seven diverse ICEMR network sites. Am J Trop Med Hyg. 2015;93(3 Suppl):110–23. 10.4269/ajtmh.14-0834 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Trape JF, Zoulani A. Malaria and urbanization in central Africa: the example of Brazzaville. Part II: Results of entomological surveys and epidemiological analysis. Trans R Soc Trop Med Hyg. 1987; 81 Suppl 2:10–8. [DOI] [PubMed] [Google Scholar]
- 9.Mwangi TW, Fegan G, Williams TN, Kinyanjui SM, Snow RW, Marsh K. Evidence for over-dispersion in the distribution of clinical malaria episodes in children. PLoS One. 2008;3:e2196 10.1371/journal.pone.0002196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cairns ME, Asante KP, Owusu-Agyei S, Chandramohan D, Greenwood BM, Milligan PJ. Analysis of partial and complete protection in malaria cohort studies. Malar J. 2013;12:355 10.1186/1475-2875-12-355 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Clark TD, Greenhouse B, Njama-Meya D, et al. Factors determining the heterogeneity of malaria incidence in children in Kampala, Uganda. J Infect Dis. 2008;198:393–400. 10.1086/589778 [DOI] [PubMed] [Google Scholar]
- 12.Alemu A, Tsegaye W, Golassa L, Abebe G. Urban malaria and associated risk factors in Jimma town, south-west Ethiopia. Malar J. 2011;10:173 10.1186/1475-2875-10-173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mathanga DP, Tembo AK, Mzilahowa T, Bauleni A, Mtimaukenena K, Taylor TE et al. Patterns and determinants of malaria risk in urban and peri-urban areas of Blantyre, Malawi. Malar J. 2016;15:590 10.1186/s12936-016-1623-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Padilla JC, Chaparro PE, Molina K, Arevalo-Herrera M, Herrera S. Is there malaria transmission in urban settings in Colombia? Malar J. 2015;14:453 10.1186/s12936-015-0956-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Osorio L, Todd J, Pearce R, Bradley DJ. The role of imported cases in the epidemiology of urban Plasmodium falciparum malaria in Quibdó, Colombia. Trop Med Int Health. 2007;12:331–41. 10.1111/j.1365-3156.2006.01791.x [DOI] [PubMed] [Google Scholar]
- 16.Molina Gómez K, Caicedo MA, Gaitán A, Herrera-Varela M, Arce MI, Vallejo AF et al. Characterizing the malaria rural-to-urban transmission interface: The importance of reactive case detection. PLoS Negl Trop Dis. 2017; 11:e0005780 10.1371/journal.pntd.0005780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rosas-Aguirre A, Ponce OJ, Carrasco-Escobar G, Speybroeck N, Contreras-Mancilla J, Gamboa D, et al. Plasmodium vivax malaria at households: spatial clustering and risk factors in a low endemicity urban area of the northwestern Peruvian coast. Malar J. 2015;14:176 10.1186/s12936-015-0670-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tada MS, Marques RP, Mesquita E, Dalla Martha RC, Rodrigues JA, Costa JD, et al. Urban malaria in the Brazilian Western Amazon Region I: high prevalence of asymptomatic carriers in an urban riverside district is associated with a high level of clinical malaria. Mem Inst Oswaldo Cruz. 2007;102:263–9. 10.1590/s0074-02762007005000012 [DOI] [PubMed] [Google Scholar]
- 19.Saraiva MGG, Amorim RDS, Moura MAS, Martinez-Espinosa FE, Barbosa MGV. Expansão urbana e distribuição espacial da malária no município de Manaus, Estado do Amazonas [in Portuguese]. Rev Soc Bras Med Trop. 2009;42:515–22. [DOI] [PubMed] [Google Scholar]
- 20.Hiwat H, Bretas G. Ecology of Anopheles darlingi Root with respect to vector importance: a review. Parasit Vectors. 2011;4:177 10.1186/1756-3305-4-177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Maheu-Giroux M, Casapía M, Soto-Calle VE, Ford LB, Buckeridge DL, Coomes OT, et al. Risk of malaria transmission from fish ponds in the Peruvian Amazon. Acta Trop. 2010;115:112–118. 10.1016/j.actatropica.2010.02.011 [DOI] [PubMed] [Google Scholar]
- 22.dos Reis IC, Codeço CT, Degener CM, Keppeler EC, Muniz MM, de Oliveira FG et al. Contribution of fish farming ponds to the production of immature Anopheles spp. in a malaria-endemic Amazonian town. Malar J. 2015;14:452 10.1186/s12936-015-0947-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Reis IC, Honório NA, Barros FS, Barcellos C, Kitron U, Camara DC et al. Epidemic and endemic malaria transmission related to fish farming ponds in the Amazon frontier. PLoS One. 2015; 10:e0137521 10.1371/journal.pone.0137521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ndungu FM, Marsh K, Fegan G, Wambua J, Nyangweso G, Ogada E, et al. Identifying children with excess malaria episodes after adjusting for a variation in exposure: identification from a longitudinal study using statistical count models. BMC Med. 2015;13:183 10.1186/s12916-015-0422-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kang SY, Battle KE, Gibson HS, Cooper LV, Maxwell K, Kamya M, et al. Heterogeneous exposure and hotspots for malaria vectors at three study sites in Uganda. Gates Open Res. 2018;2:32 10.12688/gatesopenres.12838.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yau KKW, Wang K, Lee AH. Zero-inflated negative binomial mixed regression modeling of over-dispersed count data with extra zeros. Biometr J. 2003;45:437–52. [Google Scholar]
- 27.Ferreira MU, Castro MC. Challenges for malaria elimination in Brazil. Malar J. 2016;15:284 10.1186/s12936-016-1335-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ministry of Health of Brazil. Malaria epidemiologic status in Brazil, 2012 and 2013 [in Portuguese]. Brasília, Brazil: Ministry of Health of Brazil, 2015. Available from: http://portalarquivos.saude.gov.br/images/pdf/2015/dezembro/16/2015-003—Mal—ria.pdf [Google Scholar]
- 29.Ministry of Health of Brazil. List of municipalities belonging to at risk or malaria-endemic areas [in Portuguese]. Brasília: Ministry of Health of Brazil, 2017. Available from: http://portalarquivos.saude.gov.br/images/pdf/2017/maio/19/Lista-de-municipios-pertencentes-as-areas-de-risco-ou-endemicas-para-malaria.pdf [Google Scholar]
- 30.Reis IC, Codeço CT, Câmara DCP, Carvajal JJ, Pereira GR, Keppeler EC, et al. Diversity of Anopheles spp. (Diptera: Cullicidae) in an Amazonian urban area. Neotrop Entomol. 2018;47:412–7. 10.1007/s13744-018-0595-6 [DOI] [PubMed] [Google Scholar]
- 31.Filmer D, Pritchett LH. Estimating wealth effects without expenditure data—or tears: an application to educational enrollments in states of India. Demography. 2001; 38:115–32. 10.1353/dem.2001.0003 [DOI] [PubMed] [Google Scholar]
- 32.Ministry of Health of Brazil. Malaria: Case Monitoring in Brazil in 2014 [in Portuguese]. Available from: http://portalarquivos2.saude.gov.br/images/pdf/2015/agosto/18/2015-009—Mal—ria-para-publica—-o.pdf
- 33.Ministry of Health of Brazil. Practical guidelines for malaria therapy [in Portuguese]. Brasília, Ministry of Health of Brazil; Brasília, Brazil: Ministry of Health of Brazil; 2010. Available from: http://bvsms.saude.gov.br/bvs/publicacoes/guia_pratico_malaria.pdf [Google Scholar]
- 34.Stasinopoulos MD, Rigby RA, Gillian ZH, Voudouris V, De Bastiani F. Flexible Regression and Smoothing Using GAMLSS in R. London: Chapman and Hall/CRC Press, 2017. 549 pp. [Google Scholar]
- 35.Rigby RA, Stasinopoulos DM. Generalized additive models for location, scale and shape. Appl Statist. 2005;54:507–54. [Google Scholar]
- 36.van Buuren S, Fredriks M. Worm plot: simple diagnostic device for modeling growth reference curves. Statist Med. 2001;20:1259–77. [DOI] [PubMed] [Google Scholar]
- 37.Pan J, Huang C. Random effects selection in generalized linear mixed models via shrinkage penalty function. Statist Comput. 2014;24:725–38. [Google Scholar]
- 38.Greenwood BM. The microepidemiology of malaria and its importance to malaria control. Trans R Soc Trop Med Hyg 1989;83 Suppl:25–29. [DOI] [PubMed] [Google Scholar]
- 39.Carter R, Mendis KN, Roberts D. Spatial targeting of interventions against malaria. Bull World Health Organ. 2000;78:1401–1411. [PMC free article] [PubMed] [Google Scholar]
- 40.Bousema T, Griffin JT, Sauerwein RW, Smith DL, Churcher TS, Takken W, et al. Hitting hotspots: spatial targeting of malaria for control and elimination. PLoS Med. 2012;9:e1001165 10.1371/journal.pmed.1001165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Cotter C, Sturrock HJW, Hsiang MS, Liu J, Phillips AA, Hwang J, et al. The changing epidemiology of malaria elimination: new strategies for new challenges. Lancet. 2013;382:P900–P911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bejon P, Williams TN, Nyundo C, Hay SI, Benz D, Gething PW, et al. A micro-epidemiological analysis of febrile malaria in Coastal Kenya showing hotspots within hotspots. eLife. 2014;3:e02130 10.7554/eLife.02130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lana RM, Riback TIS, Lima TFM, da Silva Nunes M, Cruz OG, Oliveira FGS et al. Socioeconomic and demographic characterization of an endemic malaria region in Brazil by multiple correspondence analysis. Malar J. 2017;16:397 10.1186/s12936-017-2045-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tusting LS, Bottomley C, Gibson H, Kleinschmidt I, Tatem AJ, Lindsay SW, et al. Housing improvements and malaria risk in sub-Saharan Africa: a multi-country analysis of survey data. PLoS Med. 2017;14:e1002234 10.1371/journal.pmed.1002234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Rek JC, Alegana V, Arinaitwe E, Cameron E, Kamya MR, Katureebe A, et al. Rapid improvements to rural Ugandan housing and their association with malaria from intense and reduced transmission: a cohort study. Lancet Planet Health. 2018;2:e83–94. 10.1016/S2542-5196(18)30010-X [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Martins LMO, David MR, Maciel-de-Freitas R, Silva-do-Nascimento TF. Diversity of Anopheles mosquitoes from four landscapes in the highest endemic region of malaria transmission in Brazil. J Vector Ecol. 2018;43:235–244. 10.1111/jvec.12307 [DOI] [PubMed] [Google Scholar]
- 47.Killeen GF, Govella NJ, Mlacha YP, Chaki PP. Suppression of malaria vector densities and human infection prevalence associated with scale-up of mosquito-proofed housing in Dar es Salaam, Tanzania: re-analysis of an observational series of parasitological and entomological surveys. Lancet Planet Health. 2019;3:e132–e143. 10.1016/S2542-5196(19)30035-X [DOI] [PubMed] [Google Scholar]
- 48.Barbosa S, Gozze AB, Lima NF, Batista CL, Bastos MS, Nicolete VC, et al. Epidemiology of disappearing Plasmodium vivax malaria: a case study in rural Amazonia. PLoS Negl Trop Dis. 2014;8:e3109 10.1371/journal.pntd.0003109 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Map of South America showing the location of the field site, the municipality of Mâncio Lima in the state of Acre, northwestern Brazil, next to the border with Peru. Figure created using data extracted from the GADM database (www.gadm.org), version 2.8, under the Creative Commons Attribution License (CCAL) CC BY 4.0 (http://creativecommons.org/licenses/by/4.0/).
(TIF)
Only study participants who had at least one malaria episode diagnosed during the follow-up are included in this analysis. The upper panel shows the frequency distributions of malaria episodes in males aged 16–40 years (A; n = 742 study participants) and in all other population strata in Mâncio Lima (B; n = 2,020). The frequency distributions are significantly different (Kolmogorov-Smirnov test, P = 0.0219). The lower panel shows the frequency distributions of malaria episodes in study participants living in the periphery (C; n = 1,263) and in the center (D; n = 1,499) of the town of Mâncio Lima. The frequency distributions are also significantly different (Kolmogorov-Smirnov test, P < 0.0001).
(TIF)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
Data Availability Statement
Original data files contain information that may potentially lead to the identification of study participants. Requests for access to original data should be submitted to the Institutional Review Board of the Institute of Biomedical Sciences, University of São Paulo, Brazil, at cep@icb.usp.br. The study identification number is CEPSH 1368.