

Abstract
Δ9‐tetrahydrocannabinol, the principal active component in Cannabis sativa extracts such as marijuana, participates in cell signalling by binding to cannabinoid CB1 and CB2 receptors on the cell surface. The CB1 receptors are present in both inhibitory and excitatory presynaptic terminals and the CB2 receptors are found in neuronal subpopulations in addition to microglial cells and astrocytes and are present in both presynaptic and postsynaptic terminals. Subsequent to the discovery of the endocannabinoid (eCB) system, studies have suggested that alcohol alters the eCB system and that this system plays a major role in the motivation to abuse alcohol. Preclinical studies have provided evidence that chronic alcohol consumption modulates eCBs and expression of CB1 receptors in brain addiction circuits. In addition, studies have further established the distinct function of the eCB system in the development of fetal alcohol spectrum disorders. This review provides a recent and comprehensive assessment of the literature related to the function of the eCB system in alcohol abuse disorders.
Abbreviations
- 2‐AG
2‐arachidonoylglycerol
- AEA
arachidonoyl ethanolamide, anandamide
- Arc
activity‐regulated cytoskeleton‐associated protein
- AUD
alcohol abuse disorder
- BLA
basolateral amygdala
- CB
cannabinoid
- CBD
cannabidiol
- CDK5
cyclin‐dependent kinase 5
- CeA
central nucleus
- CREB
cAMP response element‐binding protein
- DAGL
DAG lipase
- DNMT
DNA methyltransferase
- eCB
endocannabinoid
- FAAH
fatty acid amide hydrolase
- FABPs
fatty acid binding proteins
- FASDs
fetal alcohol spectrum disorders
- GDE1
glycerophosphodiesterase
- HP
hippocampus
- IPSCs
inhibitory postsynaptic currents
- MAGL
monoacylglycerol lipase
- MeCP2
methyl‐CpG‐binding protein 2
- msP
Marchigian Sardinian alcohol‐preferring
- NAc
nucleus accumbens
- NAPE‐PLD
N‐acylphosphatidylethanolamine‐specific PLD
- PFC
prefrontal cortex
- Rac1
Ras‐related C3 botulinum toxin substrate 1
- VTA
ventral tegmental area
- Δ9‐THC
Δ9‐tetrahydrocannabinol
1. INTRODUCTION
Cannabinoids (CBs) are naturally found in the plant Cannabis sativa. Among the over 500 different compounds in Cannabis, only approximately 85 are named CBs (Brenneisen, 2007). The understanding of the endogenous cannabinoid (eCB) system as an essential neuromodulatory system emerged more than two decades ago. The appreciation of the role of eCB came long after the discovery of the bioactive and psychoactive components of Cannabis, such as cannabidiol (CBD) and Δ9‐tetrahydrocannabinol (Δ9‐THC; see Pava & Woodward, 2012). Later, Δ9‐THC was found to elicit its psychoactive effects by binding to “orphan” GPCRs called CB receptors (Howlett et al., 1990) on the cell membrane and is mainly responsible for the psychotropic effects of cannabis plant preparations. CB receptors were identified (Howlett et al., 1990) 27 years after the discovery of Δ9‐THC, and 3 years later, the cloning of a second peripheral receptor for CBs (see Onaivi, Ishiguro, Gu, & Liu, 2012; CB 2) was achieved. CB receptor signalling is primarily involved in a range of physiological functions, as well as in several pathophysiological conditions of the CNS.
Discovery of both N‐arachidonoyl ethanolamide (anandamide [AEA]) and 2‐arachidonoylglycerol (2‐AG) in the brain emphasized the significance of CB receptors and their signalling through endogenous ligands in the regulation of a majority of physiological functions. AEA and 2‐AG act on CB receptors to elicit their biological function; thus, they are termed eCBs (see Basavarajappa, Shivakumar, Joshi, & Subbanna, 2017; Lu & Anderson, 2017). eCBs are lipophilic and are biosynthesized on demand from membrane phospholipids mainly via N‐acylphosphatidylethanolamine‐specific PLD (NAPE‐PLD), but other relevant enzymes include glycerophosphodiesterase (GDE1), abhydrolase domain 4, and the phosphatase protein tyrosine phosphatase, non‐receptor type 22. The biosynthesis of 2‐AG occurs in neurons via two possible pathways. DAG lipase‐α and ‐β (DAGLα and DAGLβ) both contribute, to a large degree, to the regulation of the steady contents of 2‐AG in the brain and other tissues (see Basavarajappa, 2015; Lu & Anderson, 2017). The eCB system functions in neural development, immune function, metabolism and energy homeostasis, pain, emotional states, arousal, sleep, stress reactivity, synaptic plasticity, learning, and the reward processing of many drugs of abuse, including alcohol, and they can readily pass through and diffuse into cellular membranes without being stored in vesicles. Both AEA and 2‐AG are derivatives of arachidonic acid, and several pathways contribute to their biosynthesis (see Basavarajappa et al., 2017). Notably, there is strong evidence for the contribution of calcium in both of these biosynthetic pathways, which may underlie the requirement for postsynaptic Ca2+ in specific forms of depolarization‐induced synaptic plasticity (see Basavarajappa & Arancio, 2008).
eCBs elicit their function principally via CB1 receptors, which are chiefly confined to the CNS, and CB2 receptors, which are widely expressed in peripheral systems and are expressed at lower concentrations in the CNS (Lu & Anderson, 2017). After being released by postsynaptic neurons, eCBs bind to CB1 receptors located on the presynaptic membrane to inhibit neurotransmitter release (Lu & Anderson, 2017). eCBs are removed from the synaptic area after activation ov CB1 receptors by transport into cells and are then hydrolysed. Fatty acid binding protein (FABP) is an intracellular carrier that transports AEA to fatty acid amide hydrolase (FAAH), an enzyme involved in AEA hydrolysis in neurons. Compounds that bind to FABP block AEA hydrolysis, increasing AEA levels (Deutsch, 2016). The pharmacological inhibition or genetic ablation of FABP5 eliminates both phasic and tonic eCB‐mediated excitatory synaptic transmission in the dorsal raphe nucleus without affecting CB1 receptors or eCB biosynthesis (Haj‐Dahmane et al., 2018). Thus, AEA action may be terminated by the coordinated function of FABP and FAAH (Deutsch, 2016). After activation of CB1 receptors, 2‐AG is hydrolysed in neurons by the action of monoacylglycerol lipase (MAGL; Lu & Anderson, 2017). Neuronal activity has been shown to stimulate eCB synthesis and release from postsynaptic neurons. The eCBs, which are lipid mediators, traverse the synapse to bind presynaptic CB1s in a retrograde manner. As retrograde messengers, eCBs provide feedback inhibition via the suppression of neurotransmitter release at both excitatory and inhibitory synapses. They serve a critical function in the regulation of both short‐ and long‐term synaptic plasticity, which functions in adaptive reward‐motivated learning (Basavarajappa et al., 2017; Lu & Anderson, 2017). In addition, eCBs mediate somatodendritic slow self‐inhibition and the long‐term modulation of inhibitory connections in layer 2/3 pyramidal neurons, allowing inhibitory and excitatory neurons to regulate their own activity in cortical circuits (Marinelli, Pacioni, Cannich, Marsicano, & Bacci, 2009). These unique functions of eCBs have provided a strong rationale for their examination as therapeutic targets for many pathological conditions.
AEA and 2‐AG activate CB receptors with differential efficacies. Both CB1 and CB2 receptors are GPCRs and, once they are activated, they are primarily positively coupled to Gi/Go proteins, leading to the inhibition of AC and calcium channels but activation of potassium channels and the regulation of many different cellular functions. The CB1 receptors are considered as the most abundant metabotropic receptor in the brain and are highly expressed in the cortex, the basal ganglia, the hippocampus (HP), and cerebellar regions. Subcellularly, CB1 receptors are present on presynaptic terminals and, therefore, these receptors are often referred to as the “brain cannabinoid receptors.” CB1 receptor densities are similar to those of GABAA‐ and glutamate‐gated ion channels (Herkenham et al., 1991). The CB2 receptors have been identified in distinct locations of the CNS in many animal species, including humans, at moderate levels, and are confined to microglia and vascular elements (Onaivi et al., 2012). However, the understanding of the specific role of this receptor in the CNS is evolving slowly (Onaivi et al., 2012; Zhang et al., 2016). Recently, the presence of Cnr2 mRNA in hippocampal neuronal cells (Li & Kim, 2015) and of dopamine‐expressing neurons in the ventral tegmental area (VTA) have been demonstrated (Zhang et al., 2016). The CB2 receptor‐mediated regulation of cell type‐specific synaptic plasticity has been shown in the hippocampus (Li & Kim, 2016). Additionally, enhanced neuronal levels of CB2 receptors have been observed under pathological conditions (Viscomi et al., 2009). We and others have reviewed the functions of the eCB system in the normal brain (Basavarajappa, 2015; Lu & Anderson, 2017), and the current review aims to expand the new understanding of the role and functions of the eCB system in alcohol abuse disorders (AUDs).
2. THE SYNERGISTIC INTERACTION BETWEEN ALCOHOL AND CANNABIS USE
The first scientific proof of the relationship between alcohol and cannabis use was noted in a study in which participants with a history of heavy cannabis use became less intoxicated from alcohol and exhibited fewer alcohol‐induced neuropsychological defects than those without a history of heavy cannabis use (Jones & Stone, 1970). However, this study presented no data from non‐cannabis‐using control participants. The results revealed that a previous history of heavy cannabis abuse might cause cross‐tolerance to the acute effects of alcohol. Additionally, a synergetic interaction between acute alcohol and cannabis use was observed in rodents in a study in which the coadministration of alcohol with Δ9‐THC increased sleep time, compared to that noted after the injection of either drug alone (Friedman & Gershon, 1974). In a double‐blinded placebo controlled investigation, alcohol and cannabis interacted synergistically to produce cognitive, psychomotor, and attention abnormalities (Marks & MacAvoy, 1989) after their coadministration. Consistent with these findings, a more recent study demonstrated that a single dose of alcohol confers tolerance to subsequent treatment with cannabis (da Silva, Morato, & Takahashi, 2001). Additionally, the pre‐administration of a single dose of cannabis enables the acute effects of alcohol (da Silva et al., 2001).
More recent studies have proposed that the acute tolerance effects of alcohol are mediated through CB1 receptors (Lemos, Takahashi, & Morato, 2007). Another study also established similar cross‐tolerance in mice subjected to the constant administration of alcohol chronically for 10 days. The mice displayed considerably diminished sensitivity to CB‐induced hypomotility, hypothermia, and antinociception through decreased CB1 receptor levels in the periaqueductal grey, hypothalamus, and VTA (Pava et al., 2012). As discussed before, most of the neurophysiological outcomes of intoxication with CBs are exerted through the eCB system by altering neurotransmission, primarily at glutamatergic and GABAergic synapses. These observations suggest that, although alcohol and cannabis have various specific effects, these two drugs produce similar cognitive deficits after both acute and chronic exposure. Nevertheless, the molecular mechanisms by which alcohol and cannabis interact are numerous and differ significantly based on the neurochemical pathways involved.
3. THE FUNCTION OF eCBs IN AUDs
Despite the early surge of studies concerning the synergistic interaction between alcohol and cannabis, there was an apparent absence of studies assessing the interaction between alcohol and CB compounds until the 1990s. Our laboratory was the first to report alcohol as a modulator of eCB biosynthetic enzymes and establish that chronic alcohol exposure causes the specific up‐regulation of a Ca2+‐dependent arachidonic acid‐specific isoform of PLA 2 in mouse brains (Basavarajappa, Cooper, & Hungund, 1998b). Shortly after that, we established that chronic alcohol vapour exposure decreases the number of CB1 receptors and their function in the mouse brain (Basavarajappa & Hungund, 1999b). Later, we showed that AEA and 2‐AG levels are increased through Ca2+‐mediated activation of PLA2 followed by the enhancement of NAPE‐PLD in cultured cells exposed to chronic alcohol (Basavarajappa & Hungund, 1999a; Basavarajappa, Saito, Cooper, & Hungund, 2000). During this time, studies indicated that CB1 receptor agonists (Gallate, Saharov, Mallet, & McGregor, 1999) and antagonists (inverse agonists; Arnone et al., 1997; Colombo et al., 1998; Gallate & McGregor, 1999; Rodriguez de Fonseca, Roberts, Bilbao, Koob, & Navarro, 1999) can increase or inhibit alcohol intake, respectively, and demonstrated that alcohol intake can be controlled via CB1 receptors. These seminal reports and advances in the biochemistry and physiology of the eCB system have aided considerably in the firmly recognized role of the eCB system in controlling the reinforcing properties of alcohol.
In other studies, the short‐term exposure of hippocampal neurons (Mironov & Hermann, 1996) to alcohol resulted in an increase in intracellular Ca2+ due to its release from intracellular stores. The exposure of hippocampal neurons in culture to acute alcohol (approximately 2 mg·ml−1) was sufficient to enhance both AEA and 2‐AG in a Ca2+‐dependent manner and to inhibit presynaptic glutamate release (Basavarajappa, Ninan, & Arancio, 2008). In another study, chronic pretreatment with WIN 55,212‐2 (WIN) rescued acute alcohol‐induced spontaneous firing in basolateral amygdala (BLA) and VTA area (Perra, Pillolla, Luchicchi, & Pistis, 2008) projection neurons. Similar results were also produced by evoked activity in nucleus accumbens (NAc) neurons (Perra et al., 2005). These results together suggest that the rewarding properties of alcohol may be decreased after the chronic activation of CB1 receptors.
In contrast to these findings, there is also data that acute alcohol treatment obstructs eCB signalling in a brain region‐specific manner. For instance, acute exposure to alcohol reduces eCB content in the hippocampus, striatum, prefrontal cortex (PFC), amygdala, and cerebellum (Ferrer et al., 2007) without altering FAAH activity (Rubio et al., 2009), signifying that the effects of acute alcohol exposure are not due to enhanced eCB metabolism. Additionally, acutely administered alcohol was shown to inhibit the CB1 receptor‐mediated presynaptic facilitation of GABAergic transmission in pyramidal neurons in the central amygdala (Roberto et al., 2010). Analyses of cerebellar Purkinje neurons have shown that the activation of CB1 receptors inhibits alcohol‐facilitated GABA release from presynaptic terminals (as demonstrated by enhanced inhibitory postsynaptic current [IPSC] frequency) through a PKA‐dependent pathway that leads to the release of Ca2+ from internal stores independent of eCB biosynthesis (Kelm, Criswell, & Breese, 2008). The administration of alcohol at concentrations relevant to intoxication enhances the frequency of spontaneous and miniature GABAergic IPSCs, a process that is blocked by eCB/ CB1 receptor modulation, suggesting that these receptors play a role in the presynaptic effects of alcohol seeking and drinking (Talani & Lovinger, 2015). Acute alcohol exposure inhibits eCB release from medium spiny neurons in the dorsomedial striatum and blocks the long‐lasting disinhibition of these neurons, and this role is independent of eCB biosynthesis and CB1 receptors (Clarke & Adermark, 2010). Our in vivo experiments with FAAH−/− mice also suggested that AEA opposes some of the acute effects of alcohol, including the loss of the righting reflex and hypothermia, while aggravating others (Basavarajappa, Yalamanchili, Cravatt, Cooper, & Hungund, 2006).
Collectively, these findings demonstrate that there is crosstalk between the eCB system and some acute effects of alcohol in a brain region‐dependent manner. The general anaesthetic propofol increases AEA via inhibiting FAAH and significantly enhances the righting reflex via CB1 receptors (Patel et al., 2003), implying that the use of anaesthetics during in vivo studies may influence eCB signalling. However, there are no studies on the influence of urethane on the eCB system. Therefore, future studies on eCBs should consider the possible exposure of the organism under evaluation to anaesthesia and alcohol and the influence of such exposure on the eCB system. Together, these studies indicate that the effects of acute alcohol exposure are mediated in part by the release of eCBs from neural tissue and their consequent actions on neurotransmission. Identifying the contribution of other eCB components to the effects of acute alcohol exposure on neurotransmission is an important next step in appreciating how the molecular effect of alcohol translates into altered neuronal and circuit function via an eCB‐mediated mechanism.
4. THE ROLE OF THE eCB SYSTEM IN THE REINFORCING PROPERTIES OF ALCOHOL
The considerable body of data collected over the past 20 years has proposed that the eCB system contributes an important but intricate role in regulating the function of reward circuitry for the rewarding properties of both non‐drugs and drugs of abuse, including alcohol (Serrano & Parsons, 2011). The action of mesolimbic dopaminergic neurons in the VTA on the NAc has been demonstrated to control reward and learning behaviour, leading to compulsive drug‐seeking behaviour (see Zhang et al., 2016). The functional modification of rostromedial tegmental nucleus projections to dopaminergic neurons via regulating 2‐AG metabolism influences the rewarding/aversive properties of alcohol, which may be a factor in the innate preference for and the enhanced intake of alcohol observed in Sardinian alcohol‐preferring rats (Melis et al., 2014). The study suggests that inhibition of GABA A receptors in VTA dopaminergic neurons is controlled by the presynaptic actions of eCBs and that long‐term withdrawal from chronic intermittent alcohol vapour exposure (CIE) treatment enhances eCB‐mediated inhibition, thereby suppressing GABA release (Harlan, Becker, Woodward, & Riegel, 2018). It has been shown that acute alcohol exposure increases dopamine release in the NAc in a CB1 receptor‐dependent manner, demonstrating that these receptors regulate the alcohol‐induced activation of VTA‐ dopaminergic neurons (Cheer et al., 2007).
As described earlier, another study demonstrated that acute alcohol treatment causes an increased firing rate of VTA‐ dopaminergic neurons in a CB1 receptor‐dependent manner (Perra et al., 2005). In our previous studies, acute alcohol‐enhanced NAc‐dopamine release was blocked by the pharmacological blockade or the genetic ablation of CB1 receptors (CB1 −/−; Hungund, Szakall, Adam, Basavarajappa, & Vadasz, 2003), and CB1 −/− mice displayed diminished alcohol‐induced conditioned place preference (CPP; Houchi et al., 2005). Collectively, these observations suggest that alcohol reward is regulated in part by the eCB‐mediated facilitation of the VTA‐dopaminergic system. Therefore, while changes in dopamine release and dopaminergic neuron firing are some of the most well‐studied and consistent effects of alcohol, more work is warranted to examine the role of the other components of the eCB system in the VTA‐dopaminergic system on alcohol reinforcing properties.
5. THE ROLE OF THE eCB SYSTEM IN ALCOHOL INTAKE/SELF‐ADMINISTRATION BEHAVIOUR
Studies have shown that genetic variability in CB1 receptor expression and signalling may influence some individuals to abuse and become dependent on alcohol. In line with this notion, we have demonstrated that C57BL/6J mice, which show a higher preference for alcohol than that of DBA/2 mice, exhibit lower levels of CB1 receptors (Hungund & Basavarajappa, 2000) and signalling (Basavarajappa & Hungund, 2001) than that of DBA/2 mice. However, it is not yet clear how much eCBs and CB1 receptors contribute to the difference in alcohol preference, and it is also possible that the observed differences other than the reduction in CB1 receptor levels may contribute to reduced alcohol drinking in the DBA/2 line. In other investigations, the pharmacological blockade of FAAH or the genetic ablation of FAAH (FAAH−/−) caused a higher preference for alcohol (Basavarajappa et al., 2006; Blednov, Cravatt, Boehm, Walker, & Harris, 2007). Similarly, AA (Alko, Alcohol; alcohol‐preferring) rats exhibit reduced FAAH activity and decreased CB1 receptor levels in the PFC compared with those of alcohol avoiding or alcohol‐non‐preferring rats (Hansson et al., 2007). In the same study, the pharmacological blockade of FAAH (in the PFC) increased alcohol preference in non‐selected Wistar rats (Hansson et al., 2007). These findings are consistent with the earlier discussion of the ability of acute alcohol exposure to enable eCB biosynthesis and release (Basavarajappa et al., 2008; Perra et al., 2008), and the reduced CB1 receptor levels observed in these studies may be due to the β‐arrestin‐mediated endocytosis of CB1 receptors (D'Souza et al., 2016) because of enhanced AEA tone. Collectively, these findings establish that impaired FAAH activity is accompanied by reduced CB1 receptor levels and enhanced alcohol preference. It is not clear how alcohol‐enhanced AEA tone activates β‐arrestin signalling to lead to the endocytosis of CB1 receptors, and this mechanism warrants further investigation.
In other investigations, the self‐administration of alcohol was demonstrated to increase the extracellular level of 2‐AG in the NAc, which is linked to the amount of alcohol consumed, but not of AEA in the same region (Caille, Alvarez‐Jaimes, Polis, Stouffer, & Parsons, 2007). Furthermore, a mouse model of methamphetamine‐induced neurotoxic lesioning of nigrostriatal dopaminergic projections exhibited increased alcohol intake and enhanced 2‐AG levels in the limbic forebrain tissues comprising the anterior cingulate and the NAc (Gutierrez‐Lopez et al., 2010). The pharmacological blockade of MAGL also enhanced alcohol consumption and preference (Serrano et al., 2018; Figure 1). However, future investigations that utilize more specific inhibitors and knockout mice to dissect the roles of AEA and 2‐AG in mediating the effects of alcohol should be employed.
Figure 1.
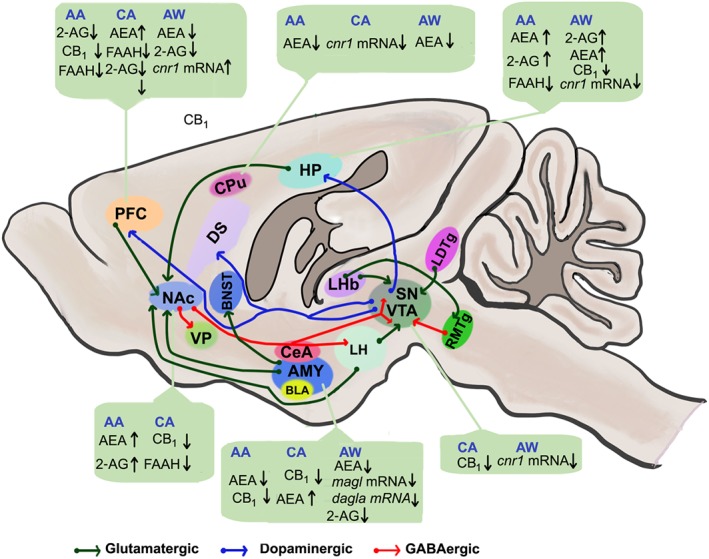
Representative sagittal cross section of a rodent brain showing the reward circuitry affected by alcohol‐induced alterations in eCB functions and highlighting signalling to and from the nucleus accumbens (NAc) and ventral tegmental area (VTA). Glutamatergic transmission drives signalling via the reward and reward‐related circuitry. GABAergic transmission from NAc and other regions suppresses neuronal activity in target regions. The release of dopamine from the VTA and substantia nigra (SN) regulates synaptic output in other target regions (dopaminergic transmission). 2‐AG, 2‐arachidonyl glycerol; AA, acute alcohol; AEA, anandamide; AMY, amygdala; AW, alcohol withdrawal; BNST, bed nucleus of the stria terminalis; CA, chronic alcohol; CB1, CB1 receptor; CeA, central nucleus of the amygdala; CPu, caudate putamen; dagla, DAG lipase‐α; DS, dorsal striatum; FAAH, fatty acid amide hydrolase; HP, hippocampus; LDTg, laterodorsal tegmentum; LHb, lateral habenula; LH, lateral hypothalamus; magl, monoacylglycerol lipase; PFC, prefrontal cortex; RMTg, rostromedial tegmental nucleus; SN, substantia nigra; VP, ventral pallidum
The activation of CB1 receptors is likely to mediate the influence of increased AEA on alcohol consumption, as several studies have demonstrated that the direct activation or blockade of CB1 receptors modifies alcohol consumption. Preadministration with a CB1 receptor agonist increases the motivation of rats to self‐administer beer in spite of increased responses for both beer and sucrose (Gallate et al., 1999). In another study, the microinjection of WIN into the posterior VTA was shown to increase binge‐like alcohol consumption in the second half of the drinking‐in‐the‐dark model, indicating that the activation of CB1 receptors in VTA neurons contributes to the motivation to consume alcohol (Linsenbardt & Boehm, 2009). Previously, the pharmacological inhibition of CB1 receptors reduced alcohol intake in C57BL/6 mice (Arnone et al., 1997). In a later study, the injection of a CB1 receptor antagonist or inverse agonist rimonabant (SR141716A or SR) was shown to reduce the self‐administration of beer (Gallate & McGregor, 1999), indicating that the pharmacological inhibition of CB1 receptors decreases the rewarding properties of alcohol. Moreover, SR administration has been reported to decrease operant responses to alcohol and sucrose through sipper tube access (Freedland, Sharpe, Samson, & Porrino, 2001). Similarly, in Sardinian alcohol‐preferring rats, SR administration reduces both alcohol and food intake (Colombo et al., 1998), and similar results have been found in AA (alcohol‐preferring) rats trained to self‐administer alcohol in an operant chamber (Hansson et al., 2007).
Consistent results were also demonstrated in a study using a self‐administration model in Wistar and Marchigian Sardinian alcohol‐preferring (msP) rats, in which the administration of SR was shown to decrease alcohol, saccharin, and sucrose intake without affecting food intake (Cippitelli et al., 2005). Additionally, decreased Cnr1 mRNA levels have been found in several brain regions of msP rats after alcohol consumption. Furthermore, SR has also been demonstrated to decrease alcohol self‐administration in alcohol‐dependent rats without affecting control rats (Rodriguez de Fonseca et al., 1999). Although the neuroanatomical region responsible for CB1‐regulated alcohol self‐administration is not well studied, the existing results suggest that the brain regions typically associated with addiction circuits may be involved. A study revealed that the microinjection of SR into the medial PFC but not into the dorsal striatum decreases alcohol self‐administration (Hansson et al., 2007). Similarly, the microinjection of SR in the NAc decreased alcohol self‐administration in another study (Caille et al., 2007). Further studies are warranted to further appreciate the participation of other brain regions, such as the VTA and amygdala, in regulating CB1 receptors and alcohol consumption.
After initial reports demonstrating that CB1 −/− mice display decreased alcohol intake and preference (Hungund et al., 2003; Wang, Liu, Harvey‐white, Zimmer, & Kunos, 2003), many studies have replicated these data (see Naassila, Pierrefiche, Ledent, & Daoust, 2004). Furthermore, in another study, the loss of glutamate clearance via genetic ablation of the astrocytic glutamate transporter GLAST (EAAT1) led to the loss of retrograde eCB signalling and reduced alcohol consumption and the rewarding properties of alcohol in mice (Karlsson et al., 2012). S426A/S430A mutant mice, which express a desensitization‐resistant form of the CB1 receptor and exhibit an increased response to eCBs and ∆9‐THC, display a modestly enhanced intake of and preference for 6% alcohol but not higher concentrations of alcohol. While CB1 receptors enhance alcohol intake, the reward response, tolerance, and acute sensitivity to alcohol and other drugs (morphine) remain normal (Marcus et al., 2017). CBD, a non‐psychoactive constituent of marijuana, binds to many receptors, such as opioid, 5‐HT and CB1. Within the eCB system, CBD is a non‐competitive antagonist of CB1 receptors. CBD acts through negative allosteric modulation and has a low affinity for the primary ligand site of CB1 receptors (for a recent review, see Chye, Christensen, Solowij, & Yucel, 2019). The administration of CBD decreases alcohol intake, alcohol‐induced hypothermia, and handling‐induced convulsions without altering blood alcohol concentrations in C57BL/6J mice. Additionally, CBD significantly reduces tyrosine hydroxylase (TH) gene expression in the VTA and reduces Oprm1, Cnr1, and Gpr55 gene expression and increases Cnr2 gene expression in the NAc. Together, these findings suggest that CBD augments alcohol‐motivated behaviours. These results strongly demonstrate that CBD may be beneficial for the treatment of alcohol use disorders. Thus, preclinical studies have proposed that the activation of CB1 receptors promotes and the inhibition of CB1 receptors blocks alcohol self‐administration, suggesting that the eCB system plays a vital function in alcohol intake. Future clinical studies using clinically safe tools to manipulate the eCB system will facilitate the use of the eCB system as a likely target for AUD treatment.
6. THE eCB SYSTEM AND ALCOHOL TOLERANCE AND DEPENDENCE
The two main characteristics of AUDs are alcohol dependence and tolerance. Tolerance is a lack of response to the repeated use of alcohol and leads to the need for higher volumes to experience the familiar effects. Dependence is a physical condition in which the body has adapted to the continued presence of alcohol and drives craving. When chronic alcohol abuse is stopped, the signs of withdrawal are initiated and cause withdrawal syndrome, leading to the consumption of more alcohol to avoid the withdrawal effects. Findings from several researchers have demonstrated that the eCB system plays a role in alcohol tolerance and dependence. Moreover, most of the previous work proposing an interaction between alcohol and CB drugs corroborates the view that the eCB system participates in mediating these two characteristics of addiction. However, at the time these studies were performed, the existence of the eCB system and the mechanisms by which alcohol and CBs produce their effects were not clear. In addition to the results of the findings discussed in the earlier sections, the symmetrical cross‐tolerance that develops from the ataxic effects of CBs and alcohol are CB1 receptor‐dependent (Lemos et al., 2007), and this cross‐tolerance seems to be consistent with changes in CB1 receptor expression (see Pava & Woodward, 2014).
After chronic alcohol exposure for 3 days, which causes alcohol tolerance and dependence, the levels and function of CB1 receptors are decreased (Basavarajappa & Hungund, 1999b). These initial results have been replicated by several researchers using different chronic and subchronic alcohol exposure models. For example, the subchronic administration of alcohol for 7 days decreases sensitivity to WIN‐induced alterations in monoamine synthesis in many brain regions (Moranta, Esteban, & Garcia‐Sevilla, 2006). In another study using rats that exhibited alcohol dependency after exposure to 52 days of forced access to a 10% alcohol solution, Cnr1 gene expression was found to be reduced in the striatum, hippocampus and hypothalamus (Ortiz, Oliva, Perez, Palomo, & Manzanares, 2004). In another dependence study using rats that were made alcohol dependent using a chronic intermittent alcohol exposure paradigm, Cnr1 gene expression and CB1 receptor protein levels in hippocampal tissues were decreased, and the CB1 receptor‐mediated inhibition of GABAergic synaptic transmission was impaired (Mitrirattanakul et al., 2007). Remarkably, alcohol withdrawal for 40 days caused recovery of CB1 receptors to above control levels. Similar recovery of CB1 receptors was found after alcohol withdrawal for 3 weeks in chronic alcohol‐administered animals (Rimondini, Arlinde, Sommer, & Heilig, 2002). In another study, alcohol administration for 10 days followed by 3 hr of withdrawal was also demonstrated to decrease CB1 receptor levels (Rubio et al., 2009). Furthermore, the chronic alcohol‐induced alterations in cortical, hippocampal, and cerebellar NMDA and GABAA receptor expression that are observed in wild‐type (WT) mice were not observed in CB1 −/− mice (Warnault et al., 2007).
Alcohol withdrawal reduces the density of CB1 receptors, eCBs, and related N‐acylethanolamines in the globus pallidus, and the administration of SR inhibits alcohol‐induced anxiogenic behaviour in animals (Rubio et al., 2008). Additionally, SR prevents alcohol‐induced dopamine deficits in the amygdala (Amy) and VTA. These findings suggest that SR may prevent alcohol withdrawal symptoms through the normalization of GABA, glutamate, and dopamine transmission in emotion‐ and motor‐related brain areas. Collectively, these data establish that alcohol exposure models that cause tolerance and dependence decrease levels and function of CB1 receptors and that alcohol withdrawal results in the up‐regulation of CB1 receptor levels as acute withdrawal symptoms lessen. Although the downstream influence of reduced CB1 receptor signalling in alcohol tolerance and dependence is not well examined, data from CB1 −/− animals suggest that reduced CB1 receptor signalling may ensure the stabilization of neural adaptations to impaired NMDA and GABAA receptors after chronic alcohol exposure. A plausible explanation for the decreased levels of CB1 receptors in chronic alcohol‐exposed animals was described in our original experiments conducted using cultured cells. The chronic exposure of cells to intoxicating concentrations of alcohol was found to increase both AEA and 2‐AG content (Basavarajappa et al., 2000; Basavarajappa & Hungund, 1999a) through the activation of PLA2 followed by eCB biosynthesis (see Basavarajappa & Hungund, 2002).
In another study in which rats were allowed to consume alcohol (7.2%) via a liquid diet, AEA content was enhanced in the limbic forebrain but was decreased in the midbrain (Gonzalez et al., 2002), amygdala, and striatum (Rubio et al., 2008). Alcohol‐dependent rats show enhanced AEA and 2‐AG content that persists 40 days into withdrawal in the hippocampus (Mitrirattanakul et al., 2007). In cultured cerebellar neurons exposed to chronic alcohol, increased levels of AEA in the media are accompanied by decreased FAAH and AEA transport mechanisms (Basavarajappa, Saito, Cooper, & Hungund, 2003). Data from human post‐mortem tissue have also established enhanced AEA levels and decreased levels of CB1 receptors and FAAH expression and activity in the ventral striatum of alcoholic patients (Vinod et al., 2010). Collectively, these data reveal that the increase in eCB levels that follows chronic alcohol administration may be due to enhanced eCB synthesis and reduced inactivation mechanisms. The further availability of methods to selectively block AEA formation will be beneficial in determining the mechanisms of CB1 receptor reduction and will improve our knowledge of the molecular pathways responsible for the distinct function of the eCB system in alcohol tolerance and dependence.
Additionally, studies have suggested that alcohol dependence causes a reduced baseline 2‐AG dialysate content and an enhanced baseline content of glutamate and GABA. Acute alcohol abstinence induces the augmentation of these dependence‐induced effects, and the levels of 2‐AG and GABA are reinstated upon alcohol re‐exposure. Moreover, alcohol self‐administration enhances the central nucleus (CeA) 2‐AG content in alcohol‐dependent rats. Enhanced anxiety‐like behaviour and alcohol intake are augmented mainly by MAGL inhibitors (Serrano et al., 2018). These findings propose a key function for eCB signalling in motivational neuroadaptations during alcohol dependence, in which a loss of CeA 2‐AG signalling in alcohol‐dependent animals is associated with stress and excessive alcohol intake behaviour. In another study, acute alcohol exposure was shown to reduce EPSP amplitudes in Wistar and male msP rats but not in female msPs. The activation of CB1 receptors by WIN reduces EPSP amplitudes in msPs and in male but not female Wistar rats. The coapplication of WIN and alcohol causes strain‐specific effects in female rats. Tonic CB1 receptor signalling was not present at glutamatergic synapses in the CeA of any of the groups, and no interaction with alcohol was observed. Collectively, these data establish sex–strain‐specific alterations in the effects of alcohol and eCB on CeA glutamatergic signalling.
The acute administration of alcohol increases relative CB1 receptor binding, as well as AEA levels in the NAc, but not after chronic alcohol consumption or after a 14‐day withdrawal period (Ceccarini, Casteels, Koole, Bormans, & Van Laere, 2013). In contrast, chronic alcohol intake reduces relative CB1 receptor binding in the hippocampus and caudate‐putamen, although these brain regions display enhanced relative CB1 receptor binding after 7 and 14 days of withdrawal. In addition, a similar alcohol withdrawal paradigm was shown to reduce relative CB1 receptor binding in the orbitofrontal cortex (Ceccarini et al., 2013). These findings suggest that alcohol affects the eCB system differentially in different brain regions. Furthermore, prolonged acute alcohol administration (0–80 mM of alcohol up to 40‐min exposure) causes the dose‐dependent inhibition of glutamatergic synaptic activity in a CB1 receptor‐dependent manner. Notably, this inhibition by acute alcohol exposure is mitigated after 10 days of CIE. Interestingly, CIE significantly reduces CB1 receptor‐mediated presynaptic inhibition at glutamatergic synapses but spares the CB1 receptor‐mediated inhibition of GABAergic synapses. CIE also significantly elevates BLA AEA content and decreases CB1 receptor protein levels (Robinson, Alexander, Bluett, Patel, & McCool, 2016). Additionally, CIE prevents the inhibitory effects of WIN on mIPSC but not sIPSC frequency, an effect that is rescued by AM251. However, acute alcohol exposure increases CeA GABA release in both naïve and alcohol‐exposed rats, and AM251 prevents these effects, suggesting the indirect participation of CB1 receptors (Varodayan et al., 2016). Furthermore, CIE in rats has been shown to inhibit retrograde tonic eCB/CB1 signalling in the BLA. However, acute alcohol exposure enhances GABAergic transmission equally in naïve and chronic alcohol‐exposed rats through both presynaptic and postsynaptic mechanisms (Varodayan et al., 2017). Together, these findings demonstrate the dynamic regulation of the CeA and BLA eCB systems by acute and chronic alcohol exposure.
The repeated 10‐day exposure of slice cultures of the PFC to alcohol enhances the duration of the up state of the activity of neurons for 4 days. The administration of WIN enhanced the amplitude of the up state in control cultures but not in those treated previously with alcohol. No significant changes in CB1 receptor protein expression has been found. Chronic alcohol treatment and withdrawal also prevent the inhibition of electrically evoked GABA IPSCs in layer II/III pyramidal neurons by WIN. However, alcohol treatment and withdrawal fail to influence WIN‐inhibited electrically evoked NMDA EPSCs in both layer II/III and V/VI neurons (Pava & Woodward, 2014). Collectively, these findings indicate that the reduction in CB1 receptor signalling that results from alcohol exposure causes altered network activity in the PFC. CIE causes the down‐regulation of CB1 receptor signalling, the loss of CB1 receptor‐dependent LTD, and the expansion of dendritic material in dorsal striatum neurons (DePoy et al., 2013). Together, these results suggest that the down‐regulation of CB1 receptors induced by chronic alcohol exposure may be a key step in the progression of alcoholism.
7. THE eCB SYSTEM IN ALCOHOL RELAPSE BEHAVIOUR
As addiction is a complex chronic disease, the goal of all addiction treatments, including treatments for alcohol addiction, is to prevent relapse, substitute the abused drug with less abusive drugs, or maintain moderate use. It is conceivable that the eCB system, which plays an indispensable role in the rewarding effects of alcohol, alcohol consumption, and alcohol withdrawal processes, may also contribute to the mechanisms related to relapse. Thus, non‐contingent exposure to WIN during a period of alcohol withdrawal enhanced relapse‐like drinking in rats. Moreover, subchronic exposure to WIN decreases dopamine release in the NAc shell in response to subsequent doses of alcohol (see Lopez‐Moreno et al., 2008). Along the same line, many investigations have established the influence of CB1 receptor inhibition on reinstatement of alcohol self‐administration. Furthermore, the combined administration of subthreshold doses of the CB1 receptor antagonist SR with either an adenosine A 2A or mGlu 5 receptor antagonist was also found to prevent relapse‐like alcohol intake (Adams, Short, & Lawrence, 2010). This latter study is exciting and may be beneficial for clinical purposes to reduce or prevent the unfavourable psychiatric side effects of higher doses of SR. Furthermore, SR exposure fails to affect foot‐shock‐elicited relapse, suggesting that CB1 receptors play no significant role in stress‐induced relapse (Economidou et al., 2007). Together, these findings demonstrate that CB1 receptors play a vital role in alcohol relapse behaviour. However, future investigations are warranted to explore the neuroanatomical location of CB1 and the function of the other components of the eCB system in preventing the reinstatement of alcohol‐seeking behaviour.
8. THE eCB SYSTEM AND SUSCEPTIBILITY TO AUDs
Despite the large number of animal studies that have been conducted on the role of the eCB system in chronic alcohol abuse and withdrawal, few studies have explored whether CNR1 gene variation contributes to the inherent susceptibility to alcohol dependence. It has been demonstrated that being homozygous for the CNR1 allele and having five or more repeats of a microsatellite polymorphism are linked to a reduced amplitude of the P300 wave of evoked related potentials in the frontal lobe (Johnson et al., 1997). Furthermore, the decreased amplitude of the P300 wave of evoked related potentials has been recognized as a physiological marker that is associated with a family history of alcohol dependence and attentional processing disorders (Begleiter, Porjesz, Bihari, & Kissin, 1984). Additionally, single nucleotide polymorphisms, such as 1359G/A (rs1049353), have been shown to facilitate the withdrawal severity experienced by chronic alcoholic patients, and those who are homozygous for the A allele exhibit more severe symptoms than those with other genotypes (Schmidt et al., 2002). In another study, it was shown that individuals with at least one copy of the C allele (rs202323) display increased craving for and salivary response to an alcohol‐associated cue (van den Wildenberg, Janssen, Hutchison, van Breukelen, & Wiers, 2007). Furthermore, the C allele of rs2023239 in the above study was accompanied by increased CB1 receptor expression in post‐mortem tissues of the human PFC. Additionally, alcohol‐dependent patients with the C allele exhibit enhanced PFC, orbitofrontal cortex, and NAc activation in response to alcohol‐associated cues and report larger subjective reward following the consumption of several alcoholic beverages (Hutchison et al., 2008). In addition, the CNR1 C allele (Marcos et al., 2012) and the FAAH C385A single nucleotide polymorphism (Buhler et al., 2014; Spagnolo et al., 2016) have been identified as likely indicators of individuals who are at higher risk for alcohol abuse. Furthermore, the FAAH Pro129Thr missense variant (rs324420) has also been shown to be associated with alcohol dependence severity in European Americans (Sloan et al., 2018). These observations together suggest that polymorphisms of eCB‐related genes may confer more susceptibility to alcohol use and indicate that a genetic polymorphism in the eCB system gene may contribute to the development of AUDs.
9. THE FUNCTION OF CB2 RECEPTORS IN AUDs
To date, most research involving the eCB system in AUDs has focused on eCB transmitters, their related synthetic and inactivating enzymes, and CB1 receptors. This is pobably due to the deep‐seated view in the CB field that the CB1 receptors represent the CB receptor in the CNS (Matsuda, Lolait, Brownstein, Young, & Bonner, 1990) and that the CB2 receptor is the peripheral CB receptor (Bayewitch et al., 1995). Additionally, the existence of CB2 receptors in the brain is controversial. However, the existence and function of central CB2 receptors are beginning to expand (Onaivi et al., 2012), and a recent behavioural study has revealed that CB2 receptors are implicated in anxiogenic, pneumonic, and motor processes (Ortega‐Alvaro, Aracil‐Fernandez, Garcia‐Gutierrez, Navarrete, & Manzanares, 2011). Additionally, alcohol exposure and consumption have been shown to alter Cnr2 gene expression in the brain (Ishiguro et al., 2007). Mice treated chronically with alcohol for 21 days exhibit reduced CB2 receptor levels in the PFC and the hippocampus at the end of the chronic alcohol treatment. However, on the fifth day of withdrawal, CB2 receptors were found to be enhanced, and alcohol challenge was observed to counteract CB2 receptor up‐regulation in the PFC, VTA, amygdala, striatum, and hippocampus (Al Mansouri et al., 2014). Acute alcohol administration increases Th and Oprm1 gene expression in CB2 −/− mice, while a lower alcohol dose decreases Th gene expression in WT mice. CB2 −/− mice exhibit increased handling‐induced convulsion scores, alcohol‐induced CPP, voluntary alcohol intake, and preference compared with those of WT mice (Al Mansouri et al., 2014). The CB2 receptor agonist β‐caryophyllene dose dependently reduces alcohol intake and preference in a two‐bottle choice paradigm. Most importantly, β‐caryophyllene repressed alcohol‐induced CPP acquisition and exacerbates the loss of righting reflex duration. Remarkably, these effects are augmented in mice preadministered with a selective CB2 receptor antagonist (AM‐630; Al Mansouri et al., 2014). Together, these findings suggest that CB2 receptors play a role in alcohol dependence and sensitivity.
A lack of CB2 receptors leads to increased alcohol consumption in an intermittent forced drinking paradigm under group‐housing conditions (Pradier, Erxlebe, Markert, & Racz, 2015). The infusion of a selective CB2 receptor agonist (JWH‐133) in the dorsal hippocampus (DH) has been shown to decrease glutamate release in the DH in alcohol‐naïve rats and is rescued by AM‐630 (Zheng, Wu, Dong, Ding, & Song, 2015). Intra‐DH infusions of JWH‐133 inhibit ischaemia‐induced glutamate release in the DH after 30 days of withdrawal. The administration of JWH‐133 failed to increase cumulative alcohol intake. JWH‐133 specifically augments the harmful effect of alcohol on NPC proliferation in the subventricular zone of the lateral ventricles and subgranular zone of the dentate gyrus (Rivera et al., 2015). These findings suggest that the specific activation of CB2 receptors may provide neuroprotection against neural damage in alcohol dependence. CB2 −/− mice display an increased magnitude of alcohol‐induced CPP compared to that of WT mice. Furthermore, neither agonists nor antagonists of CB2 receptors influence alcohol consumption or the induction of CPP, and CB2 receptor antagonist treatment during CPP acquisition trials also does not affect CPP (Powers, Breit, & Chester, 2015).
Studies have indicated that the genetic ablation of the Cnr2 gene enhances the preference for and vulnerability to alcohol intake partly through the enhanced alcohol‐induced sensitivity of Th and Oprm1 gene expression in mesolimbic neurons (Navarrete, Garcia‐Gutierrez, & Manzanares, 2018). The activation of CB2 receptors by JWH‐133 significantly decreases the number of reinforced responses, 8% alcohol consumption, breaking point, Th gene expression in the VTA, and Oprm1 gene expression in the NAc. Furthermore, the inhibition of CB2 receptors by AM‐630 produces a significantly contrary effect (Navarrete et al., 2018). These data suggest that the activation of CB2 receptors significantly reduces alcohol consumption, and further studies are required to dissect the mechanism(s) by which CB2 receptors influence the development of AUDs.
10. THE ROLE OF THE eCB SYSTEM DURING DEVELOPMENT AND ITS FUNCTION IN FETAL ALCOHOL SPECTRUM DISORDER
The developing brain exhibits a wide distribution of CB1 receptors, and the pattern of expression of these receptors parallels that of neuronal differentiation in the embryo from the most primitive stages. Numerous studies have established the Cnr1 mRNA expression pattern and distribution of CB1 receptors in the fetal and neonatal rat brain (Berrendero, Sepe, Ramos, Di Marzo, & Fernandez‐Ruiz, 1999). Cnr1 mRNA expression and receptor binding have been reported from gestational day 14 in rats, corresponding to the phenotypic expression pattern of most components of neurotransmitter systems (see Insel, 1995). At this age, of CB1 receptors are already coupled to Gi/Go proteins, indicating that they are functional (Berrendero et al., 1999). Developing human and rat brains express higher levels of CB1 receptors (see Glass, Dragunow, & Faull, 1997). The presence of CB1 receptors during early brain development suggests the possible participation of these receptors in cell proliferation, migration, and axonal elongation and later in synaptogenesis and myelinogenesis (see Basavarajappa et al., 2017). Hence, CB1 receptors contribute to generating neuronal divergence in different brain regions throughout early brain development. CB1 receptors are expressed in the presynaptic area of brain regions that are central to the regulation of learning and memory (hippocampus), fear, anxiety (amygdala), stress (hypothalamic nuclei), depression (PFC), and addiction (striatum; see Basavarajappa et al., 2017). We still have limited knowledge of the developmental role of the eCB system with respect to brain maturation and circuit formation, and future studies in this direction are warranted.
CB exposure during the early developmental period has been shown to cause delays in the maturation of neurotransmitter systems and impair their activities (Fernandez‐Ruiz, Berrendero, Hernandez, & Ramos, 2000). These negative consequences are due to the activation of CB1 receptors, which are expressed early in the developing brain (Berrendero et al., 1999; Fernandez‐Ruiz et al., 2000). Exposure to CBs at doses similar to those observed in cannabis users has been shown to delay neurotransmitter maturation and trigger neurobehavioural defects (de Salas‐Quiroga et al., 2015). The acute administration of Δ9‐THC markedly enhances the pro‐apoptotic properties of alcohol in the neonatal rat brain (Hansen et al., 2008). However, Δ9‐THC does not induce neurodegeneration by itself, even though neuronal loss becomes widespread and severe when Δ9‐THC is combined with a mildly intoxicating alcohol dose. The effects of this combination of Δ9‐THC and a moderate dose of alcohol dose resemble the massive neurodegeneration observed when alcohol is administered alone at much higher doses (Hansen et al., 2008). Additionally, Δ9‐THC and the coadministration of a low dose of alcohol increase expression of CB1 receptors without affecting expression of CB2 receptors in the thalamus and dorsal subiculum. The influence of Δ9‐THC on neuronal cell death is mirrored by the effects of WIN (1–10 mg·kg−1) in a CB1 receptor‐dependent manner (Hansen et al., 2008). Additionally, neonatal CB1 −/− mice are less susceptible to the neurotoxic effects of a low dose of alcohol. Moreover, the CB1 receptor antagonist SR prevents the apoptotic effects of alcohol (Hansen et al., 2008).
The function of the CB1 receptor signalling pathway during brain development has not been well characterized. The available data suggest the role of ERK1/2 via a mechanism comprising the upstream inhibition of Rap1 and B‐Raf. The activation of CB1 receptors also inhibits the recruitment of new synapses by inhibiting the formation of cAMP (see Harkany et al., 2007). While intracellular signalling events involving MAPK coupled with the activation of CB1 receptors have been examined in the embryonic developmental stage (Berghuis et al., 2007), they have not been well described during postnatal development. Many studies using different cell lines in culture have demonstrated that MAPK is both up‐ and down‐regulated during Δ9‐THC‐mediated apoptosis (see Galve‐Roperh et al., 2000). Additionally, cannabis exposure during brain development also causes a variety of defects, which are perhaps facilitated by the activation of CB1 receptors, that are similar to what is observed in several specific human developmental disorders (see Stefanis et al., 2004) and may well overlap with those found in fetal alcohol syndrome (Wu, Jew, & Lu, 2011), which is probably mediated through enhanced function of CB1 receptors.
In addition to increased AEA and associated biosynthetic enzymes, the alcohol‐induced transcriptional activation of the Cnr1 gene results in increased levels of Cnr1 mRNA and CB1 receptor protein expression in cortical and hippocampal brain regions (Subbanna, Shivakumar, Psychoyos, Xie, & Basavarajappa, 2013). Remarkably, we found that postnatal alcohol exposure in mice enhances the acetylation of histone (H4) on Lys8 (H4K8ace) in exon 1 of Cnr1 and CB1 receptor binding and CB1 receptor agonist‐stimulated GTPγS binding in cortical and hippocampal brain regions (Subbanna, Nagre, Umapathy, Pace, & Basavarajappa, 2015). The administration of SR or the genetic ablation of CB1 receptors (CB1 −/−) before alcohol exposure prevents neuronal cell death (Subbanna et al., 2013; Subbanna et al., 2015). Interestingly, synaptic plasticity, learning, and memory are disrupted by early alcohol exposure and are then restored by the pharmacological blockade or genetic deletion of CB1 receptors. The enhanced AEA/ CB1 receptor signalling pathway may be directly responsible for the neurobehavioural defects accompanying fetal alcohol spectrum disorder (FASD).
Studies using a postnatal alcohol exposure model have established the specific roles of CB1 receptor‐mediated pERK1/2, phosphorylated cAMP response element‐binding protein (pCREB), pAkt, and activity‐regulated cytoskeleton‐associated protein (Arc) in alcohol‐induced neurodegeneration. P7 alcohol treatment significantly reduces the activation of ERK1/2, Akt, and CREB, which is followed by the inhibition of Arc protein expression in the hippocampus and neocortex (Subbanna et al., 2013). Furthermore, the inhibition of ERK1/2, CREB, and Arc protein expression by ethanol is prevented by SR pretreatment, but Akt activation is not affected. Likewise, CB1 −/− mice, which do not show alcohol‐induced neurodegeneration, are protected against the P7 alcohol‐induced inhibition of ERK1/2 and CREB activation and Arc protein expression, but they fail to induce the inhibition of Akt phosphorylation. Therefore, alcohol‐activated, CB1 receptor‐induced neurodegeneration is regulated by the CB1/pERK1/2/pCREB/Arc pathway but not by PI3K/Akt signalling in the developing brain (Subbanna et al., 2013; Subbanna et al., 2015; Figure 2). CB1 receptor‐mediated Arc regulation via the MAPK pathway is an essential physiological mechanism by which CBs and eCBs can modulate synaptic plasticity.
Figure 2.
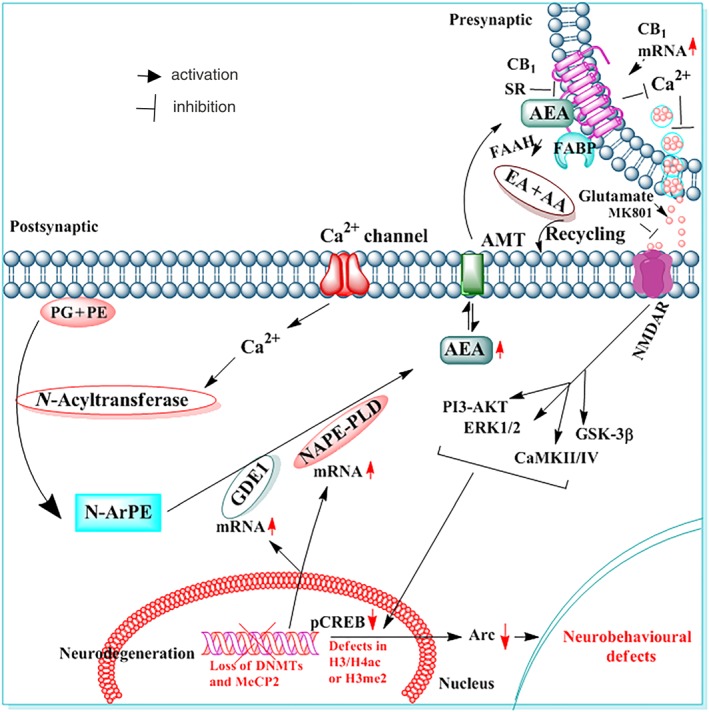
Graphic representation of CB1 receptor function in the development of neurobehavioural deficits induced by developmental alcohol exposure. Postnatal alcohol exposure enhances AEA levels in postsynaptic neurons through the transcription activation of the genes encoding the enzymes NAPE‐PLD and GDE1. AEA, acting through CB1 receptors on presynaptic neurons, results in decreased glutamate release, which causes NMDA receptor (NMDAR) hypofunction and CDK5, ERK1/2, and CREB hypophosphorylation, leading to inhibition of Arc and Rac1 expression followed by neonatal neurodegeneration. Earlier studies have shown that activation of CB1 receptors inhibits NMDAR function in several experimental models (Twitchell, Brown, & Mackie, 1997) and alcohol has been shown to reduce glutamatergic neurotransmission via activation of CB1 receptors (Basavarajappa et al., 2008). These CB1 receptor events during postnatal development may disrupt the refinement of neuronal circuits (Wilson, Peterson, Basavaraj, & Saito, 2011) and cause long‐lasting deficits in synaptic plasticity and memory in adult animals. The inhibition of CB1 receptors (AEA tone) prevents CDK5 activation, pERK1/2 and CREB hypophosphorylation, the loss of MeCP2, DNMT1/2 and DNA methylation, deficits in Arc and Rac1 expression, and neonatal neurodegeneration (through tau and caspase‐3 cleavage), leading to normal neurobehavioural function in adult mice. The genetic ablation of CB1 receptors does not affect NMDAR antagonist‐induced apoptosis but does protect against alcohol‐induced neonatal neurodegeneration and synaptic and memory deficits in adult mice. Thus, the putative AEA/CB1/CDK5/pERK1/2/pCREB/Arc/Rac1 signalling mechanism may have a possible regulatory role in neuronal function in the developing brain and may be a valuable therapeutic target for FASD. The effects of alcohol are shown in red or with red arrows
The pharmacological blockade of the NMDA receptor for a few hours during the synaptogenesis period has been shown to trigger massive and widespread neuronal apoptosis in the rodent brain (Ikonomidou et al., 1999). Therefore, at this developmental stage, the survival of neurons is dependent on glutamatergic input that is controlled within narrow periods (Ikonomidou et al., 1999). eCBs and CBs are known to inhibit glutamatergic signalling (Gerdeman & Lovinger, 2001), and therefore, alcohol‐induced eCBs (Basavarajappa et al., 2008; Subbanna et al., 2013) may contribute to the neonatal apoptosis and lasting behavioural defects (see Joshi, Subbanna, Shivakumar, & Basavarajappa, 2019) observed after binge‐like alcohol exposure during this specific susceptible period of brain development. Additionally, the inhibition or genetic deletion of CB1 receptors rescues the eCB‐mediated blockade of glutamate release by alcohol, resulting in a reduction in alcohol‐elicited neuronal apoptosis. Thus, CB1s serve as good candidate targets for regulating NMDA receptor function in developmental disorders. Interestingly, earlier studies have shown that the apoptotic effects of alcohol are mediated by inhibiting NMDA receptors (Ikonomidou, Stefovska, & Turski, 2000). In our studies, an NMDA receptor antagonist was found to induce apoptosis in CB1 −/− mice but not by alcohol, indicating that the CB1 receptor‐mediated glutamate release is responsible for the apoptotic action of alcohol through NMDA receptors. Collectively, these data indicate that alcohol‐induced activation of CB1 receptors negatively regulates NMDA receptor function (Basavarajappa et al., 2008), causing apoptosis in the developing brain (Subbanna et al., 2013) and further proving the mechanism by which postnatal alcohol induces its harmful effects in the developing brain.
The data obtained from neonatal rats demonstrate that alcohol may affect CA3 pyramidal neurons through the inhibition of postsynaptic AMPA receptors, which results in the blockade of glutamatergic function (Mameli, Zamudio, Carta, & Valenzuela, 2005). Furthermore, it has been observed that exogenous CBs block glutamatergic release by activating the CB1 receptor‐mediated inhibition of N‐type (Cav2.2) and P/Q‐type calcium (Cav2.1) channels (Twitchell et al., 1997) and may be responsible for the increased vulnerability of the immature brain to alcohol neurotoxicity (Hansen et al., 2008) and persistent neurobehavioural defects (for references, see Joshi et al., 2019). Collectively, these findings suggest that a heightened CB1 receptor signalling pathway may delay the maturation of synaptic circuits, and future studies are required to elucidate the underlying molecular mechanisms.
Although the molecular events are still being revealed, alcohol exposure during early postnatal development prompts persistent synaptic defects in adulthood (for references, see Joshi et al., 2019). These defects are due to alcohol‐enhanced AEA‐ CB1 receptor signalling, which delays the maturation of neuronal circuits and causes long‐lasting neurobehavioural defects. This could clarify why some cortical maps and olfactory‐hippocampal networks (Wilson et al., 2011) are changed in FASD models. Consistent with these data, the inhibition of CB1 receptor activity completely rescues postnatal alcohol‐induced LTP defects (Subbanna et al., 2013). Similarly, the genetic ablation of CB1 receptors offers complete protection against postnatal alcohol‐induced LTP deficits. However, CB1 −/− mice display a greater LTP magnitude compared to that of WT or C57BL/6J saline‐exposed mice (Subbanna et al., 2013; Subbanna et al., 2015; Subbanna & Basavarajappa, 2014), as found in other studies (Bohme, Laville, Ledent, Parmentier, & Imperato, 2000; Reibaud et al., 1999). Additionally, postnatal alcohol exposure produces object recognition and spatial and social interaction memory deficits, which are blocked in mice by treatment with a CB1 receptor antagonist (Subbanna et al., 2013). Additionally, CB1 −/− mice are protected against postnatal alcohol‐induced memory and social interaction defects, as observed by LTP. It is also likely that AEA/ CB1 receptor signalling during the critical period of brain development can interrupt the maturation of several neurotransmitter systems, including the glutamatergic, catecholaminergic, serotonergic, GABAergic, and opioid systems (Fernandez‐Ruiz et al., 2000), subsequently contributing to a diminished hippocampal network and long‐term behavioural defects (Schneider, 2009). Although more investigations are warranted, enhanced CB1 receptor activity during postnatal development can lead to long‐term behavioural deficits (Campolongo, Trezza, Ratano, Palmery, & Cuomo, 2011) that are controlled by NMDA receptor activity (Subbanna et al., 2013). Moreover, more research is warranted to determine the influence of enhanced CB1 receptor activity during brain development on the maturation of multiple neurotransmitters, which may also instigate lasting morphological alterations underlying synaptic and memory defects.
Furthermore, postnatal alcohol exposure activates caspase‐3 through CB1 receptors and leads to the loss of DNA methyltransferases (DNMT1 and DNMT3A; Nagre, Subbanna, Shivakumar, Psychoyos, & Basavarajappa, 2015), a methylated DNA binding protein (methyl‐CpG‐binding protein 2 [MeCP2]) and DNA methylation in neonatal mice (Nagre et al., 2015). CB1 −/− or the injection of SR before alcohol exposure not only prevents caspase‐3 activation but also augments the loss of DNMT1, DNMT3A, MeCP2, pCREB, and Arc expression. Collectively, these data indicate that the alcohol‐induced, CB1 receptor‐controlled activation of caspase‐3 facilitates the degradation of DNMT1, DNMT3A, and MeCP2 in the P7 mouse brain and triggers long‐lasting neurobehavioural defects in adult mice. This CB1 receptor‐regulated instability of MeCP2 during active synaptic maturation may delay synaptic circuit maturation and contribute to neurobehavioural defects, as observed in this animal model of FASD.
Additionally, postnatal alcohol exposure also generates p25, a cyclin‐dependent kinase 5 (CDK5)‐activating peptide, and silences Ras‐related C3 botulinum toxin substrate 1 (Rac1) expression through an epigenetic mechanism in a CB1 receptor‐dependent manner (Joshi et al., 2019). The inhibition of CDK5 activity augments the alcohol‐induced loss of Rac1 expression in neonatal mice. Rac1 expression is regulated by the presence of H3K9me2 and G9a, which repress chromatin, in the Rac1 gene promoter region, causing the persistent loss of Rac1 expression in adulthood. The inhibition of CDK5 activity by roscovitine (seliciclib) in P7 mice also rescues neurodegeneration in neonatal mice and augments pERK1/2, pCREB, and Arc signalling deficits and the loss of Rac1 gene expression, synaptic plasticity, and behavioural defects in adult mice exposed to alcohol at P7. These data indicate that the CB1 receptor‐mediated (Subbanna et al., 2013; Subbanna et al., 2014) up‐regulation of CDK5/p25 activity followed by the inhibition of pERK, pCREB, and the epigenetic suppression of Arc (Subbanna et al., 2018) and Rac1 expression is responsible for the long‐lasting neurobehavioural defects observed in adult mice exposed to alcohol at P7.
In a recent investigation, it was noted that the administration of SR before alcohol treatment in P7 mice rescues activity‐dependent (Y‐maze behaviour) signalling defects, such as signalling defects in phosphorylated calcium/calmodulin‐dependent PK IV, pCREB, and phosphorylated calcium/calmodulin‐dependent PK II, in adult mice exposed postnatally to alcohol. The administration of SR prior to alcohol exposure also rescues impaired activity‐dependent global epigenetic marks such as H4K8 acetylation (ac), H3K14ac, and H3K9 dimethylation (me2) on the Arc gene promoter in adult mice exposed postnatally to alcohol (Subbanna, Joshi, & Basavarajappa, 2018). Collectively, these findings highlight the significance of the AEA‐CB1/CDK5/pERK/pCREB/Arc/Rac1 signalling mechanism in the development of FASD. The regulation of excitatory synaptic plasticity in VTA dopaminergic neurons is substantially altered in PE‐exposed adult animals (Hausknecht, Shen, Wang, Haj‐Dahmane, & Shen, 2017). Both moderate and high doses of alcohol reduce CB1 receptor function and persistently impair the low‐frequency stimulation‐induced eCB‐LTD of VTA dopaminergic neurons. These processes may contribute to increased LTP and the maintenance of better excitatory synaptic strength in VTA dopaminergic neurons, leading to increased addiction vulnerability after gestational alcohol exposure. Together, these findings suggest that alcohol exposure during development impairs synaptic events differently in different brain regions, causing learning and memory defects and increasing the susceptibility to addiction.
Recently, there have been many advances in the treatment of addiction using compounds that target the eCB system. Based on the preclinical studies already mentioned, it is clear that many components of the eCB system may be a target for treating human alcohol consumption and AUDs, although clinical studies on rimonabant have not been very successful and limited evidence suggests a non‐significant reduction in relapse to heavy drinking in the rimonabant group compared with the placebo group (George et al., 2010; Soyka et al., 2008). Thus, future investigations with larger sample sizes are necessary before the conclusion that rimonabant is not a suitable treatment can be made. New better CB1 receptor antagonists and neutral CB1 receptor antagonists may have the potential to treat human alcohol consumption. Further extensive studies are required to examine the potential clinical use of FAAH/MAGL activators, NAPE‐PLD inhibitors, and CBD to treat human AUDs.
11. SUMMARY
The past literature related to the interaction between marijuana and alcohol undoubtedly suggests the significant role of the eCB system in the acute reinforcing properties of alcohol and the neuroadaptations that occur with its chronic use. By the end of the 1990s, the molecular components of the eCB system were well defined. In the past decade, many investigations have shown the direct relationship between alcohol and the eCB system. Moreover, acute alcohol (Table 1) consumption inhibits glutamate release via enhanced eCB release in hippocampal neurons. If a similar mechanism exists in cortical neurons, one would assume that alcohol‐enhanced eCB release would inhibit cortical output and hence produce a synergistic pathway with that of the mesolimbic dopaminergic system. Several reports have demonstrated that alcohol enhances the tissue content of eCBs, such as 2‐AG, in the NAc of rats after alcohol self‐administration (Table 2). Additionally, the infusion of CB1 receptor agonists into the posterior VTA increases alcohol intake, indicating a common pathway.
Table 1.
Acute ethanol effects on the EC system
| Measure | Ethanol exposure | System | Species | Genetic background | Method of analysis | Brain region | Effect | Reference |
|---|---|---|---|---|---|---|---|---|
| AEA | 50‐mM ethanol, 30 min | In vitro | Mouse | C57BL/6J | Cell and medium extract | HP | ↑ | Basavarajappa et al., 2008 |
| 8% v/v ethanol in liquid diet, 24‐hr access | Ex vivo | Rat | Sprague–Dawley | Tissue content | AMY | ↓ | Rubio, McHugh, Fernandez‐Ruiz, Bradshaw, & Walker, 2007 | |
| CPu | ↓ | |||||||
| HyTh | ↓ | |||||||
| PFC | None | |||||||
| 4 g·kg−1 ethanol, i.p. injection | Ex vivo | Rat | Wistar | Tissue content | Cereb | ↓ | Ferrer et al., 2007 | |
| HP | ↓ | |||||||
| NAc | ↓ | |||||||
| Microdialysis | NAc | None | ||||||
| 10% (w/v) ethanol, self‐administration (30 min) | In vivo | Rat | Wistar | Microdialysis | NAc | None | Caille et al., 2007 | |
| 4 g·kg−1 ethanol, i.p. injection | In vivo | Rat | Wistar | Microdialysis | NAc | ↑ | Ceccarini et al., 2013 | |
| 2‐AG | 50‐mM ethanol, 30 min | In vitro | Mouse | C57BL/6J | Cell and medium extract | HP | ↑ | Basavarajappa et al., 2008 |
| 8% v/v ethanol in liquid diet, 24‐hr access | Ex vivo | Rat | Sprague–Dawley | Tissue content |
AMY CPu HyTh PFC |
None None None ↓ |
Rubio et al., 2007 | |
| 10% (w/v) ethanol, self‐administration (30 min) | In vivo | Rat | Wistar | Microdialysis | NAc | ↑ | Caille et al., 2007 | |
| CB1 | 50‐mM ethanol, 30–60 min | In vitro | Mouse | C57BL/6J | Protein | HP | None | Basavarajappa et al., 2008 |
|
4 g·kg−1 ethanol, i.p. injection 4 g·kg−1 ethanol, i.p. injection |
Ex vivo In vivo |
Rat Rat |
Wistar Wistar |
mRNA PET imaging |
Cereb NAc |
None ↑ binding |
Ferrer et al., 2007
Ceccarini et al., 2013 |
|
| 8% v/v ethanol in liquid diet, 24‐hr access | Ex vivo | Rat | Sprague–Dawley | Protein | AMY | ↓ | Rubio et al., 2009 | |
| HyTh | None | |||||||
| PFC | ↓ | |||||||
| CPu | None | |||||||
| FAAH | 8% v/v ethanol in liquid diet, 24‐hr access | Ex vivo | Rat | Sprague–Dawley | Activity/protein | AMY | None | Rubio et al., 2009 |
| CPu | None | |||||||
| HyTh | Activity (↓); protein (↑) | |||||||
| PFC | Activity (↓); protein (none) | |||||||
| 4 g·kg−1 ethanol, i.p. injection | Ex vivo | Rat | Wistar | Activity/mRNA | Cereb | None | Ferrer et al., 2007 | |
| HP | Activity (↓); mRNA (none) | |||||||
| NAc | None |
Abbreviations: 2‐AG, 2‐arachidonyl glycerol; AEA, anandamide; AMY, amygdala; CB1, cannabinoid receptor 1; Cereb, cerebellum; CPu, caudate putamen; FAAH, fatty acid amide hydrolase; HP, hippocampus; HyTh, hypothalamus; NAc, nucleus accumbens; PFC, prefrontal cortex.
Table 2.
Influence of EC system activity on alcohol abuse behaviours
| Mode of modulation | Method | Behavioural paradigm | Drug administration | Genus | Strain | Outcome | Reference |
|---|---|---|---|---|---|---|---|
| CB1 receptor activation | CP‐55,940 (30 μg·kg−1) | Lick‐based progressive ratio operant responding for beer | Systemic | Rat | Wistar | ↑ | Gallate et al., 1999 |
| WIN (10 mg·kg−1) | Deprivation‐induced escalation of operant response for ethanol | Systemic | Rat | Wistar | ↑ | Lopez‐Moreno, Gonzalez‐Cuevas, Rodriguez de Fonseca, & Navarro, 2004 | |
| CP‐55,940 (30 μg·kg−1) | Two‐bottle choice | Systemic | Mouse | C57BL/6J and DBA | ↑ | Vinod et al., 2008 | |
| WIN (10 mg·kg−1) | Deprivation‐induced escalation of operant response for ethanol | Systemic | Rat | Wistar | ↑ | Alen et al., 2009 | |
| WIN (0.5 mg·kg−1) | Drinking in the dark | Systemic | Mouse | C57BL/6J | ↑ | Linsenbardt & Boehm, 2009 | |
| WIN (0.5 μg per side) | VTA microinjection | Mouse | C57BL/6J | ↑ | Linsenbardt & Boehm, 2009 | ||
| WIN (3 mg·kg−1) | Two‐bottle choice | Systemic | Mouse | CD1 | ↑ | Frontera, Gonzalez Pini, Messore, & Brusco, 2018 | |
| CB1 inactivation | SR141716 (3 mg·kg−1) | Two‐bottle choice | Systemic | Mouse | C57BL/6J | ↓ | Arnone et al., 1997 |
| SR141716 (5 mg·kg−1) | Two‐bottle choice | Systemic | Rat | sP | ↓ | Colombo et al., 1998 | |
| SR141716 (1.5 and 3 mg·kg−1) | Lick‐based progressive ratio operant responding for beer | Systemic | Rat | Wistar | ↓ | Gallate et al., 1999; Gallate & McGregor, 1999 | |
| SR141716 (3 mg·kg−1) | Operant ethanol self‐administration in dependent animals | Systemic | Rat | Wistar | ↓ | Rodriguez de Fonseca et al., 1999 | |
| SR141716 (3 mg·kg−1) | Operant ethanol self‐administration | Systemic | Rat | Long‐Evans | ↓ | Freedland et al., 2001 | |
| SR141716 (3 mg·kg−1) | Operant ethanol self‐administration | Systemic | Rat | msP | ↓ | Cippitelli et al., 2005 | |
| SR141716 (3 mg·kg−1) | Operant ethanol self‐administration | Systemic | Rat | Wistar | ↓ | Cippitelli et al., 2005; Economidou et al., 2006 | |
| SR141716 (3 μg per side) | Operant ethanol self‐administration | NAc microinjection | Rat | Wistar | ↓ | Caille et al., 2007 | |
| SR141716 (3 μg per side) | Operant ethanol self‐administration | PFC microinjection | Rat | AA | ↓ | Hansson et al., 2007 | |
| AM251 (6 mg·kg−1) | Two‐bottle choice | Systemic | Rat | Fawn‐hooded | ↓ | Femenia, Garcia‐Gutierrez, & Manzanares, 2010 | |
| SR141716 (3 mg·kg−1) | Cue‐induced reinstatement to operant ethanol self‐administration | Systemic | Rat | Wistar | ↓ | Cippitelli et al., 2005 | |
| SR141716 (0.3 and 1 mg·kg−1) | Operant ethanol self‐administration | Systemic | Rat | iP | ↓ | Adams et al., 2010 | |
| SR141716 (3 mg·kg−1) | Two‐bottle choice | Systemic | Mouse | C57BL/6J | ↓ | Wang et al., 2003 | |
| CBD (30, 60, and 120 mg·kg−1) | Two‐bottle choice | Systemic | Mouse | C57BL/6J | ↓ | Viudez‐Martinez et al., 2018 | |
| CB1 null mice | Two‐bottle choice | — | Mouse | C57BL/6J | ↓ | Wang et al., 2003; Hungund et al., 2003 | |
| CD1 | ↓ | Naassila et al., 2004 | |||||
| FAAH inactivation | FAAH null mice | Two‐bottle choice | — | Mouse | 129/SvJxC57BL/6J | ↑ | Basavarajappa et al., 2006; Blednov et al., 2007 |
| URB597 (4.0 μg or 1 mg·kg−1) | Operant ethanol self‐administration | PFC microinjection | Rat | Wistar | ↑ | Hansson et al., 2007 | |
| Systemic | |||||||
| URB597 (1 mg·kg−1) | Two‐bottle choice | Systemic | Mouse | 129/SvJxC57BL/6J | ↑ | Blednov et al., 2007 | |
| URB597 (0.5–1 mg·kg−1) after 1 day of withdrawal from 3‐week chronic intermittent alcohol drinking | Two‐bottle choice | Systemic | Mouse | C57BL/6J | ↓ | Zhou et al., 2017 | |
| URB597 (1 μg) | Operant ethanol self‐administration | CeA and BLA microinjection | Rat | msP | ↓ | Stopponi et al., 2017 | |
| MAGL inhibition | MJN110 (10 and 20 mg·kg−1) | Operant ethanol self‐administration | Systemic | Rat | Male Wistar | ↓ | Serrano et al., 2018 |
| JZL184 (1 and 3 mg·kg−1) | Two‐bottle choice | Systemic | Mouse | Male C57BL/6J | ↓ |
Abbreviations: BLA, basolateral amygdala; CB1, cannabinoid receptor 1; CeA, central amygdala; FAAH, fatty acid amide hydrolase; iP, alcohol preferring; MAGL, monoacylglycerol lipase; msP, Marchigian Sardinian alcohol‐preferring; NAc, nucleus accumbens; PFC, prefrontal cortex; VTA, ventral tegmental area.
Furthermore, the broad variety of treatment paradigms applied by many of these investigations provides a strong, comprehensive view of the timescale of modifications to the eCB system, mainly changes in CB1 receptors. Experiments using a 3‐day alcohol exposure paradigm have consistently demonstrated an increase in eCB content that is associated with reduced FAAH and CB1 receptor activity, but this activity is reversed to basal levels after only 24 hr of withdrawal. In experiments where the duration of alcohol treatment is somewhat longer and the blood ethanol concentration varies because the subjects are not under a chronic exposure paradigm, CB1 receptor expression appears to be much more inconsistent and is brain region specific. In long‐term treatment paradigms followed by the immediate examination of CB1 receptor expression, the findings appear to consistently report decreased levels of CB1 receptors with enhanced eCB levels (Table 3). From these findings, it is clear that eCB release in response to alcohol promotes the reinforcing effects of alcohol and that chronic alcohol treatment that leads to tolerance and dependence significantly alters eCB‐mediated signalling.
Table 3.
Repeated ethanol and withdrawal effects on the EC system
| Measure | Ethanol exposure | Duration | System | Genus | Genetic background | Method of analysis | Brain region | Effect | Duration of abstinence | Withdrawal or abstinence effects | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AEA | 50–150 mM of ethanol | 24–72 hr | In vitro | Human | Neuroblastoma SK‐N‐SH cells | Cell and medium extract | — | ↑ | — | — | Basavarajappa & Hungund, 1999a |
| 100–150 mM of ethanol | 72 hr | In vitro | Rat | Wistar | Cell and medium extract | Cereb | ↑ | — | — | Basavarajappa et al., 2003 | |
| 7.2% v/v ethanol in liquid diet, 24‐hr access | 10–15 days | Ex vivo | Rat | Sprague–Dawley | Tissue content | AMY | — | 3 hr | ↓ | Gonzalez et al., 2002; Rubio et al., 2008 | |
| CPu | — | 3 hr | ↓ | ||||||||
| HyTh | — | 3 hr | None | ||||||||
| Cereb | None | — | — | ||||||||
| HP | None | — | — | ||||||||
| Limbic forebrain | ↓ | — | — | ||||||||
| Midbrain | ↓ | — | — | ||||||||
| Striatum | None | — | — | ||||||||
| Forced vapour inhalation | 72 hr | Ex vivo | Mouse | Swiss‐Wistar | Tissue content | Cortex | ↑ | 24 hr | None | Vinod, Yalamanchili, Xie, Cooper, & Hungund, 2006 | |
| Oral chronic intermittent 5–6 g·kg−1 alternate days | 120 days | Ex vivo | Mouse | Swiss‐Wistar | Tissue content | HP | None | 40 days | ↑ | Mitrirattanakul et al., 2007 | |
| Forced chronic ethanol consumption | 7 days | In vivo | Rat | Wistar | Microdialysis | NAc | None | 7–14 days | None | Ceccarini et al., 2013 | |
| Alcohol‐dependent patients | Lifetime | — | Human | — | Plasma content | Plasma | ↑ | 28 days | — | Garcia‐Marchena et al., 2017 | |
| Chronic intermittent ethanol vapour | 21 days every day | In vivo | Rat | Male Wistar | AEA dialysate | CeA | None | 12 hr | ↓ | Serrano et al., 2018 | |
| 7 days | None | ||||||||||
| Chronic intermittent ethanol vapour | 10 days every day | Ex vivo | Rat | Male Sprague–Dawley | AEA | BLA | ↑ | — | — | Robinson et al., 2016 | |
| NAPE‐PLD protein | BLA | None | — | — | |||||||
| FAAH protein | BLA | None | — | — | |||||||
| Chronic intermittent ethanol vapour | 48 days every day | Ex vivo | Rat | Male Wistar | AEA | BLA | ↓ | 3 days | ↓ | Henricks et al., 2017 | |
| Female Wistar | AEA | vmPFC | — | 3 days | — | ||||||
| Male Wistar | Napeld mRNA | BLA | — | 3 days | — | ||||||
| Female Wistar | Napeld mRNA | vmPFC | ↓ | 3 days | ↓ | ||||||
| Male Wistar | Faah mRNA | BLA | — | 3 days | — | ||||||
| Female Wistar | Faah mRNA | vmPFC | — | 3 days | — | ||||||
| 2‐AG | 100–150 mM of ethanol | 48–72 hr | In vitro | Rat | Sprague–Dawley | Cell and medium extract | Cereb | ↑ | — | — | Basavarajappa et al., 2000 |
| Oral chronic intermittent 5–6 g·kg−1 alternate days | 120 days + 2 days of withdrawal | Ex vivo | Mouse | Swiss‐Wistar | Tissue content | HP | ↑ | 40 days | ↑ | Mitrirattanakul et al., 2007 | |
| 7.2% v/v ethanol in liquid diet | 10–15 days | Ex vivo | Rat | Wistar | Tissue content | AMY | — | 3 hr | None | Rubio et al., 2007 | |
| CPu | — | 3 hr | None | ||||||||
| HyTh | — | 3 hr | |||||||||
| Cereb | None | — | — | ||||||||
| Cortex | None | — | — | ||||||||
| HP | None | — | |||||||||
| Limbic forebrain | None | — | — | ||||||||
| Midbrain | ↓ | — | — | ||||||||
| Striatum | None | — | — | ||||||||
| Chronic intermittent ethanol vapour | 48 days every day | Ex vivo | Rat | Male Wistar | Mgll mRNA | BLA | ↓ | 3 days | ↓ | Henricks et al., 2017 | |
| Female Wistar | Mgll mRNA | vmPFC | — | 3 days | — | ||||||
| Male Wistar | Dagla mRNA | BLA | ↓ | 3 days | ↓ | ||||||
| Female Wistar | Dagla mRNA | vmPFC | — | 3 days | — | ||||||
| Male Wistar | 2‐AG | BLA | — | 3 days | — | ||||||
| Female Wistar | 2‐AG | vmPFC | ↓ | 3 days | ↓ | ||||||
| 10 days every day | 10 days every day | Ex vivo | Rat | Male Sprague–Dawley | 2‐AG | BLA | None | — | — | Robinson et al., 2016 | |
| DAGL‐α protein | BLA | None | — | — | |||||||
| MAGL protein | BLA | None | — | — | |||||||
| Chronic intermittent ethanol vapour | 21 days every day | In vivo | Rat | Male Wistar | Mgll mRNA | CeA | — | 12 hr | ↑ | Serrano et al., 2018 | |
| Dagl mRNA | CeA | — | 12 hr | ↑ | |||||||
| 2‐AG dialysate | CeA | ↓ | 12 hr | ↓ | |||||||
| 7 days | ↓ | ||||||||||
| FAAH | Chronic ethanol treatment (100–150 mM) | 72 hr | In vitro | Rat | Sprague–Dawley | Activity and EC transport | Cereb | ↓ | — | — | Basavarajappa et al., 2003 |
| Forced vapour inhalation | 72 hr | Ex vivo | Mouse | Swiss‐Webster | Activity | Cortex | ↓ | — | — | Vinod et al., 2006 | |
| Alcohol dependent (patients/control) | Lifetime | Post‐mortem | Human | — | Protein and activity | NAc | ↓ | — | — | Vinod et al., 2010 | |
| CB1 | Continuous forced vapour inhalation | 72–96 hr | Ex vivo | Mouse | Swiss‐Webster | Binding and GTPγS binding | Whole brain | ↓ | 24 hr | None | Basavarajappa, Cooper, & Hungund, 1998a; Basavarajappa & Hungund, 1999b |
| Cortex | ↓ | 24 hr | None | Vinod et al., 2006 | |||||||
| HP | ↓ | ||||||||||
| Striatum | ↓ | ||||||||||
| Whole brain | ↓ | ||||||||||
| Chronic forced vapour inhalation (intermittent) | 49 days | Ex vivo | Rat | Wistar | mRNA | Cortex | — | 3 weeks | ↑ | Rimondini et al., 2002 | |
| 7.2% v/v ethanol in liquid diet | 10–15 days | Ex vivo | Rat | Wistar | mRNA and binding | AMY | — | 3 hr | None | Gonzalez et al., 2002; Rubio et al., 2008 | |
| None | — | — | |||||||||
| CPu | — | 3 hr | None | ||||||||
| None | — | — | |||||||||
| HyTh | None | — | — | ||||||||
| Cereb | None | — | — | ||||||||
| Cortex | — | 3 hr | None | ||||||||
| None | — | — | |||||||||
| HP | None | — | — | ||||||||
| NAc | None | — | — | ||||||||
| GP | — | 3 hr | ↓ | ||||||||
| SN | — | 3 hr | ↓ | ||||||||
| Forced consumption of ethanol (10%, v/v) | 52 days | Ex vivo | Rat | Wistar | mRNA | CPu | ↓ | — | — | Ortiz et al., 2004 | |
| Cortex | None | ||||||||||
| HP | Region dependent | ||||||||||
| HyTh | ↓ | — | — | ||||||||
| Operant ethanol self‐administration | 10 days | Ex vivo | Rat | msP | mRNA | AMY | None | — | — | Cippitelli et al., 2005 | |
| CPu | ↓ | ||||||||||
| Cortex | None | ||||||||||
| HP | None | ||||||||||
| Alcohol dependent (patients/control) | Lifetime | Post‐mortem | Human | — | Protein and GTPγS | NAc | ↓ | — | — | Vinod et al., 2010 | |
| Chronic, 5–6 g·kg−1 every other day (intermittent) | 120 days | Ex vivo | Rat | Sprague–Dawley | Protein and mRNA | HP | ↓ | 40 days | ↑ | Mitrirattanakul et al., 2007 | |
| Twice daily 4 g·kg−1 i.p. | 10 days | Ex vivo | Mouse | C57BL/6J | Protein | Cereb | None | — | — | Pava et al., 2012 | |
| Cortex | None | — | — | ||||||||
| HP | None | — | — | ||||||||
| HyTh | ↓ | — | — | ||||||||
| NAc | None | — | — | ||||||||
| Striatum | None | — | — | ||||||||
| VTA | ↓ | — | — | ||||||||
| Forced chronic ethanol consumption | 7 days | In vivo | Rat | Wistar | PET imaging | HP | ↓ binding | 7–14 days | ↑ binding | Ceccarini et al., 2013 | |
| CPu | ↓ binding | ↑ binding | |||||||||
| OFC | — | ↓ binding | |||||||||
| Chronic intermittent ethanol vapour | 10 days every day | Ex vivo | Rat | Male Sprague–Dawley | Protein | BLA | ↓ | — | — | Robinson et al., 2016 |
Abbreviations: 2‐AG, 2‐arachidonyl glycerol; AEA, anandamide; AMY, amygdala; BLA, basolateral amygdala; CB1, cannabinoid receptor 1; CeA, central amygdala; Cereb, cerebellum; CPu, caudate putamen; DAGL, DAG lipase; FAAH, fatty acid amide hydrolase; GP, globus pallidus; HP, hippocampus; HyTh, hypothalamus; MAGL, monoacylglycerol lipase; msP, marchigian Sardinian alcohol‐preferring; NAc, nucleus accumbens; NAPE‐PLD, N‐acylphosphatidylethanolamine‐specific PLD; OFC, orbitofrontal cortex; PFC, prefrontal cortex; SN, substantia nigra; vmPFC, ventromedial prefrontal cortex; VTA, ventral tegmental area.
Studies showing the participation of CB2 receptors in AUD have also emerged. However, our understanding of the pathways responsible for the changes in the eCB system in response to chronic alcohol is incomplete, and the data on the distinct function of the eCB system in regulating specific addiction circuits are also inadequate. Further experiments with more standardized methodologies are fundamental to better appreciate the complexity underlying the interaction between the eCB system and alcohol dependence. Furthermore, substantial evidence has emerged from developmental studies in which AEA‐CB1/CDK5/pERK/pCREB/Arc/Rac1 signalling and impaired eCB‐LTD of VTA dopaminergic neurons significantly contribute to alcohol‐elicited developmental disorders such as FASD. Additionally, novel areas of studies continue to emerge with basic findings surrounding the molecular components of the eCB system, and the extensive characterization of inhibitors (CB1 receptor antagonists, inverse agonists, CB2 receptor agonists, and CBD, NAPE‐PLD, and DAGL inhibitors) and activators (FAAH and MAGL) of the eCB system may provide a promising therapeutic value for the treatment of AUD.
11.1. Nomenclature of targets and ligands
Key protein targets and ligands in this article are hyperlinked to corresponding entries in http://www.guidetopharmacology.org, the common portal for data from the IUPHAR/BPS Guide to PHARMACOLOGY (Harding et al., 2018), and are permanently archived in the Concise Guide to PHARMACOLOGY 2017/18 (Alexander, Christopoulos et al., 2017; Alexander, Fabbro et al., 2017; Alexander, Peters et al., 2017; Alexander, Striessnig et al., 2017).
CONFLICT OF INTEREST
The authors declare no conflicts of interest.
ACKNOWLEDGEMENTS
This work was supported by National Institute of Health/NIAAA Grant AA019443 (B.S.B.). We thank Neha Balapal for editing the final version of the manuscript.
Basavarajappa BS, Joshi V, Shivakumar M, Subbanna S. Distinct functions of endogenous cannabinoid system in alcohol abuse disorders. Br J Pharmacol. 2019;176:3085–3109. 10.1111/bph.14780
REFERENCES
- Adams, C. L. , Short, J. L. , & Lawrence, A. J. (2010). Cue‐conditioned alcohol seeking in rats following abstinence: Involvement of metabotropic glutamate 5 receptors. British Journal of Pharmacology, 159(3), 534–542. 10.1111/j.1476-5381.2009.00562.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al Mansouri, S. , Ojha, S. , Al Maamari, E. , Al Ameri, M. , Nurulain, S. M. , & Bahi, A. (2014). The cannabinoid receptor 2 agonist, β‐caryophyllene, reduced voluntary alcohol intake and attenuated ethanol‐induced place preference and sensitivity in mice. Pharmacology, Biochemistry, and Behavior, 124, 260–268. 10.1016/j.pbb.2014.06.025 [DOI] [PubMed] [Google Scholar]
- Alen, F. , Santos, A. , Moreno‐Sanz, G. , Gonzalez‐Cuevas, G. , Gine, E. , Franco‐Ruiz, L. , … López‐Moreno, J. A. (2009). Cannabinoid‐induced increase in relapse‐like drinking is prevented by the blockade of the glycine‐binding site of N‐methyl‐D‐aspartate receptors. Neuroscience, 158(2), 465–473. 10.1016/j.neuroscience.2008.10.002 [DOI] [PubMed] [Google Scholar]
- Alexander, S. P. , Christopoulos, A. , Davenport, A. P. , Kelly, E. , Marrion, N. V. , Peters, J. A. , … CGTP Collaborators . (2017). The Concise Guide to PHARMACOLOGY 2017/18: G protein‐coupled receptors. British Journal of Pharmacology, 174(Suppl 1), S17–S129. 10.1111/bph.13878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Fabbro, D. , Kelly, E. , Marrion, N. V. , Peters, J. A. , Faccenda, E. , … CGTP Collaborators . (2017). The Concise Guide to PHARMACOLOGY 2017/18: Enzymes. British Journal of Pharmacology, 174, S272–S359. 10.1111/bph.13877 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Peters, J. A. , Kelly, E. , Marrion, N. V. , Faccenda, E. , Harding, S. D. , … CGTP Collaborators . (2017). The Concise Guide to PHARMACOLOGY 2017/18: Ligand‐gated ion channels. British Journal of Pharmacology, 174, S130–S159. 10.1111/bph.13879 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Striessnig, J. , Kelly, E. , Marrion, N. V. , Peters, J. A. , Faccenda, E. , … CGTP Collaborators . (2017). The Concise Guide to PHARMACOLOGY 2017/18: Voltage‐gated ion channels. British Journal of Pharmacology, 174, S160–S194. 10.1111/bph.13884 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnone, M. , Maruani, J. , Chaperon, F. , Thiebot, M. , Poncelet, M. , Soubrie, P. , & Le Fur, G. (1997). Selective inhibition of sucrose and ethanol intake by SR 141716, an antagonist of central cannabinoid (CB1) receptors. Psychopharmacology, 132, 104–106. [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. (2015). Fetal alcohol spectrum disorder: Potential role of endocannabinoids signaling. Brain Sciences, 5(4), 456–493. 10.3390/brainsci5040456 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , & Arancio, O. (2008). Synaptic plasticity: Emerging role for endocannabinoid system In Kaiser T. F., & Peters F. J. (Eds.), Synaptic plasticity: New research (pp. 77–112). NY, USA: Nova Science Publishers, Inc. [Google Scholar]
- Basavarajappa, B. S. , Cooper, T. B. , & Hungund, B. L. (1998a). Chronic ethanol administration down‐regulates cannabinoid receptors in mouse brain synaptic plasma membrane. Brain Research, 793, 212–218. 10.1016/S0006-8993(98)00175-9 [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , Cooper, T. B. , & Hungund, B. L. (1998b). Effect of chronic ethanol exposure on mouse brain arachidonic acid specific phospholipase A2 . Biochemical Pharmacology, 55, 515–521. 10.1016/S0006-2952(97)00501-7 [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , & Hungund, B. L. (1999a). Chronic ethanol increases the cannabinoid receptor agonist, anandamide and its precursor N‐arachidonyl phosphatidyl ethanolamine in SK‐N‐SH cells. Journal of Neurochemistry, 72, 522–528. 10.1046/j.1471-4159.1999.0720522.x [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , & Hungund, B. L. (1999b). Down‐regulation of cannabinoid receptor agonist‐stimulated [35S] GTPgS binding in synaptic plasma membrane from chronic ethanol exposed mouse. Brain Research, 815, 89–97. 10.1016/S0006-8993(98)01072-5 [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , & Hungund, B. L. (2001). Cannabinoid receptor agonist‐stimulated [35S]Guanosine TriphosphategS binding in the brain of C57BL/6 and DBA/2 mice. Journal of Neuroscience Research, 64, 429–436. 10.1002/jnr.1094 [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , & Hungund, B. L. (2002). Neuromodulatory role of the endocannabinoid signaling system in alcoholism: An overview. Prostaglandins, Leukotrienes, and Essential Fatty Acids, 66, 287–299. 10.1054/plef.2001.0352 [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , Ninan, I. , & Arancio, O. (2008). Acute ethanol suppresses glutamatergic neurotransmission through endocannabinoids in hippocampal neurons. Journal of Neurochemistry, 107(4), 1001–1013. 10.1111/j.1471-4159.2008.05685.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , Saito, M. , Cooper, T. B. , & Hungund, B. L. (2000). Stimulation of cannabinoid receptor agonist 2‐arachidonylglycerol by chronic ethanol and its modulation by specific neuromodulators in cerebellar granule neurons. Biochemica Biophysica Acta, 1535, 78–86. 10.1016/S0925-4439(00)00085-5 [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , Saito, M. , Cooper, T. B. , & Hungund, B. L. (2003). Chronic ethanol inhibits the anandamide transport and increases extracellular anandamide levels in cerebellar granule neurons. European Journal of Pharmacology, 466, 73–83. 10.1016/S0014-2999(03)01557-7 [DOI] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , Shivakumar, M. , Joshi, V. , & Subbanna, S. (2017). Endocannabinoid system in neurodegenerative disorders. Journal of Neurochemistry, 142(5), 624–648. 10.1111/jnc.14098 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basavarajappa, B. S. , Yalamanchili, R. , Cravatt, B. F. , Cooper, T. B. , & Hungund, B. L. (2006). Increased ethanol consumption and preference and decreased ethanol sensitivity in female FAAH knockout mice. Neuropharmacology, 50, 834–844. 10.1016/j.neuropharm.2005.12.005 [DOI] [PubMed] [Google Scholar]
- Bayewitch, M. , Avidor‐Reiss, T. , Levy, R. , Barg, J. , Mechoulam, R. , & Vogel, Z. (1995). The peripheral cannabinoid receptor: Adenylate cyclase inhibition and G protein coupling. FEBS Letters, 375(1–2), 143–147. 10.1016/0014-5793(95)01207-U [DOI] [PubMed] [Google Scholar]
- Begleiter, H. , Porjesz, B. , Bihari, B. , & Kissin, B. (1984). Event‐related brain potentials in boys at risk for alcoholism. Science, 225(4669), 1493–1496. 10.1126/science.6474187 [DOI] [PubMed] [Google Scholar]
- Berghuis, P. , Rajnicek, A. M. , Morozov, Y. M. , Ross, R. A. , Mulder, J. , Urban, G. M. , … Harkany, T. (2007). Hardwiring the brain: Endocannabinoids shape neuronal connectivity. Science, 316(5828), 1212–1216. 10.1126/science.1137406 [DOI] [PubMed] [Google Scholar]
- Berrendero, F. , Sepe, N. , Ramos, J. A. , Di Marzo, V. , & Fernandez‐Ruiz, J. J. (1999). Analysis of cannabinoid receptor binding and mRNA expression and endogenous cannabinoid contents in the developing rat brain during late gestation and early postnatal period. Synapse, 33(3), 181–191. [DOI] [PubMed] [Google Scholar]
- Blednov, Y. A. , Cravatt, B. F. , Boehm, S. L. II , Walker, D. , & Harris, R. A. (2007). Role of endocannabinoids in alcohol consumption and intoxication: Studies of mice lacking fatty acid amide hydrolase. Neuropsychopharmacology, 32, 1570–1582. 10.1038/sj.npp.1301274 [DOI] [PubMed] [Google Scholar]
- Bohme, G. A. , Laville, M. , Ledent, C. , Parmentier, M. , & Imperato, A. (2000). Enhanced long‐term potentiation in mice lacking cannabinoid CB1 receptors. Neuroscience, 95, 5–7. [DOI] [PubMed] [Google Scholar]
- Brenneisen, R. (2007). Chemistry and analysis of phytocannabinoids and other Cannabis constituents In ElSohly M. A. (Ed.), Marijuana and the cannabinoids (pp. 17–49). Totowa, NJ: Humana Press. [Google Scholar]
- Buhler, K. M. , Huertas, E. , Echeverry‐Alzate, V. , Gine, E. , Molto, E. , Montoliu, L. , & López‐Moreno, J. A. (2014). Risky alcohol consumption in young people is associated with the fatty acid amide hydrolase gene polymorphism C385A and affective rating of drug pictures. Molecular Genetics and Genomics: MGG, 289(3), 279–289. 10.1007/s00438-013-0809-x [DOI] [PubMed] [Google Scholar]
- Caille, S. , Alvarez‐Jaimes, L. , Polis, I. , Stouffer, D. G. , & Parsons, L. H. (2007). Specific alterations of extracellular endocannabinoid levels in the nucleus accumbens by ethanol, heroin, and cocaine self‐administration. The Journal of Neuroscience, 27(14), 3695–3702. 10.1523/JNEUROSCI.4403-06.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campolongo, P. , Trezza, V. , Ratano, P. , Palmery, M. , & Cuomo, V. (2011). Developmental consequences of perinatal cannabis exposure: Behavioral and neuroendocrine effects in adult rodents. Psychopharmacology, 214(1), 5–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceccarini, J. , Casteels, C. , Koole, M. , Bormans, G. , & Van Laere, K. (2013). Transient changes in the endocannabinoid system after acute and chronic ethanol exposure and abstinence in the rat: A combined PET and microdialysis study. European Journal of Nuclear Medicine and Molecular Imaging, 40(10), 1582–1594. 10.1007/s00259-013-2456-1 [DOI] [PubMed] [Google Scholar]
- Cheer, J. F. , Wassum, K. M. , Sombers, L. A. , Heien, M. L. , Ariansen, J. L. , Aragona, B. J. , … Wightman, R. M. (2007). Phasic dopamine release evoked by abused substances requires cannabinoid receptor activation. The Journal of Neuroscience, 27(4), 791–795. 10.1523/JNEUROSCI.4152-06.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chye, Y. , Christensen, E. , Solowij, N. , & Yucel, M. (2019). The endocannabinoid system and cannabidiol's promise for the treatment of substance use disorder. Frontiers in Psychiatry, 10, 63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cippitelli, A. , Bilbao, A. , Hansson, A. C. , del Arco, I. , Sommer, W. , Heilig, M. , … European TARGALC Consortium (2005). Cannabinoid CB1 receptor antagonism reduces conditioned reinstatement of ethanol‐seeking behavior in rats. The European Journal of Neuroscience, 21(8), 2243–2251. [DOI] [PubMed] [Google Scholar]
- Clarke, R. B. , & Adermark, L. (2010). Acute ethanol treatment prevents endocannabinoid‐mediated long‐lasting disinhibition of striatal output. Neuropharmacology, 58(4–5), 799–805. 10.1016/j.neuropharm.2009.12.006 [DOI] [PubMed] [Google Scholar]
- Colombo, G. , Agabio, R. , Fa, M. , Guano, L. , Lobina, C. , Loche, A. , … Gessa, G. L. (1998). Reduction of voluntary ethanol intake in ethanol‐preferring sP rats by the cannabinoid antagonist SR‐141716. Alcohol and Alcoholism, 33, 126–130. [DOI] [PubMed] [Google Scholar]
- da Silva, G. E. , Morato, G. S. , & Takahashi, R. N. (2001). Rapid tolerance to Δ9‐tetrahydrocannabinol and cross‐tolerance between ethanol and Δ9‐tetrahydrocannabinol in mice. European Journal of Pharmacology, 431(2), 201–207. 10.1016/S0014-2999(01)01449-2 [DOI] [PubMed] [Google Scholar]
- DePoy, L. , Daut, R. , Brigman, J. L. , MacPherson, K. , Crowley, N. , Gunduz‐Cinar, O. , … Holmes, A. (2013). Chronic alcohol produces neuroadaptations to prime dorsal striatal learning. Proceedings of the National Academy of Sciences of the United States of America, 110(36), 14783–14788. 10.1073/pnas.1308198110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Salas‐Quiroga, A. , Diaz‐Alonso, J. , Garcia‐Rincon, D. , Remmers, F. , Vega, D. , Gomez‐Canas, M. , … Galve‐Roperh, I. (2015). Prenatal exposure to cannabinoids evokes long‐lasting functional alterations by targeting CB1 receptors on developing cortical neurons. Proceedings of the National Academy of Sciences of the United States of America, 112(44), 13693–13698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deutsch, D. G. (2016). A personal retrospective: Elevating anandamide (AEA) by targeting fatty acid amide hydrolase (FAAH) and the fatty acid binding proteins (FABPs). Frontiers in Pharmacology, 7, 370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Souza, D. C. , Cortes‐Briones, J. A. , Ranganathan, M. , Thurnauer, H. , Creatura, G. , Surti, T. , … Skosnik, P. D. (2016). Rapid changes in cannabinoid 1 receptor availability in cannabis‐dependent male subjects after abstinence from cannabis. Biological Psychiatry. Cognitive Neuroscience and Neuroimaging, 1(1), 60–67. [DOI] [PubMed] [Google Scholar]
- Economidou, D. , Mattioli, L. , Cifani, C. , Perfumi, M. , Massi, M. , Cuomo, V. , … Ciccocioppo, R. (2006). Effect of the cannabinoid CB1 receptor antagonist SR‐141716A on ethanol self‐administration and ethanol‐seeking behaviour in rats. Psychopharmacology, 183(4), 394–403. 10.1007/s00213-005-0199-9 [DOI] [PubMed] [Google Scholar]
- Economidou, D. , Mattioli, L. , Ubaldi, M. , Lourdusamy, A. , Soverchia, L. , Hardiman, G. , … Ciccocioppo, R. (2007). Role of cannabinoidergic mechanisms in ethanol self‐administration and ethanol seeking in rat adult offspring following perinatal exposure to Δ9‐tetrahydrocannabinol. Toxicology and Applied Pharmacology, 223(1), 73–85. 10.1016/j.taap.2007.05.008 [DOI] [PubMed] [Google Scholar]
- Femenia, T. , Garcia‐Gutierrez, M. S. , & Manzanares, J. (2010). CB1 receptor blockade decreases ethanol intake and associated neurochemical changes in fawn‐hooded rats. Alcoholism, Clinical and Experimental Research, 34(1), 131–141. 10.1111/j.1530-0277.2009.01074.x [DOI] [PubMed] [Google Scholar]
- Fernandez‐Ruiz, J. , Berrendero, F. , Hernandez, M. L. , & Ramos, J. A. (2000). The endogenous cannabinoid system and brain development. Trends in Neurosciences, 23(1), 14–20. 10.1016/S0166-2236(99)01491-5 [DOI] [PubMed] [Google Scholar]
- Ferrer, B. , Bermudez‐Silva, F. J. , Bilbao, A. , Alvarez‐Jaimes, L. , Sanchez‐Vera, I. , Giuffrida, A. , et al. (2007). Regulation of brain anandamide by acute administration of ethanol. The Biochemical Journal, 404(1), 97–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freedland, C. S. , Sharpe, A. L. , Samson, H. H. , & Porrino, L. J. (2001). Effects of SR141716A on ethanol and sucrose self‐administration. Alcoholism, Clinical and Experimental Research, 25, 277–282. 10.1111/j.1530-0277.2001.tb02209.x [DOI] [PubMed] [Google Scholar]
- Friedman, E. , & Gershon, S. (1974). Effect of δ8‐THC on alcohol‐induced sleeping time in the rat. Psychopharmacologia, 39(3), 193–198. 10.1007/BF00421026 [DOI] [PubMed] [Google Scholar]
- Frontera, J. L. , Gonzalez Pini, V. M. , Messore, F. L. , & Brusco, A. (2018). Exposure to cannabinoid agonist WIN 55,212‐2 during early adolescence increases alcohol preference and anxiety in CD1 mice. Neuropharmacology, 137, 268–274. 10.1016/j.neuropharm.2018.05.018 [DOI] [PubMed] [Google Scholar]
- Gallate, J. E. , & McGregor, I. S. (1999). The motivation for beer in rats: Effects of ritanserin, naloxone and SR 141716. Psychopharmacology, 142, 302–308. 10.1007/s002130050893 [DOI] [PubMed] [Google Scholar]
- Gallate, J. E. , Saharov, T. , Mallet, P. E. , & McGregor, I. S. (1999). Increased motivation for beer in rats following administration of a cannabinoid CB1 receptor agonist. European Journal of Pharmacology, 370(3), 233–240. 10.1016/S0014-2999(99)00170-3 [DOI] [PubMed] [Google Scholar]
- Galve‐Roperh, I. , Sanchez, C. , Cortes, M. L. , Gomez del Pulgar, T. , Izquierdo, M. , & Guzman, M. (2000). Anti‐tumoral action of cannabinoids: Involvement of sustained ceramide accumulation and extracellular signal‐regulated kinase activation. Nature Medicine, 6(3), 313–319. 10.1038/73171 [DOI] [PubMed] [Google Scholar]
- Garcia‐Marchena, N. , Pavon, F. J. , Pastor, A. , Araos, P. , Pedraz, M. , Romero‐Sanchiz, P. , … Serrano, A. (2017). Plasma concentrations of oleoylethanolamide and other acylethanolamides are altered in alcohol‐dependent patients: Effect of length of abstinence. Addiction Biology, 22(5), 1366–1377. 10.1111/adb.12408 [DOI] [PubMed] [Google Scholar]
- George, D. T. , Herion, D. W. , Jones, C. L. , Phillips, M. J. , Hersh, J. , Hill, D. , … Kunos, G. (2010). Rimonabant (SR141716) has no effect on alcohol self‐administration or endocrine measures in nontreatment‐seeking heavy alcohol drinkers. Psychopharmacology, 208(1), 37–44. 10.1007/s00213-009-1704-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerdeman, G. , & Lovinger, D. M. (2001). CB1 cannabinoid receptor inhibits synaptic release of glutamate in rat dorsolateral striatum. Journal of Neurophysiology, 85, 468–471. 10.1152/jn.2001.85.1.468 [DOI] [PubMed] [Google Scholar]
- Glass, M. , Dragunow, M. , & Faull, R. L. (1997). Cannabinoid receptors in the human brain: A detailed anatomical and quantitative autoradiographic study in the fetal, neonatal and adult human brain. Neuroscience, 77, 299–318. 10.1016/S0306-4522(96)00428-9 [DOI] [PubMed] [Google Scholar]
- Gonzalez, S. , Grazia Cascio, M. , Fernandez‐Ruiz, J. , Fezza, F. , Di Marzo, V. , & Ramos, J. A. (2002). Changes in endocannabinoid contents in the brain of rats chronically exposed to nicotine, ethanol or cocaine. Brain Research, 954, 73. [DOI] [PubMed] [Google Scholar]
- Gutierrez‐Lopez, M. D. , Llopis, N. , Feng, S. , Barrett, D. A. , O'Shea, E. , & Colado, M. I. (2010). Involvement of 2‐arachidonoyl glycerol in the increased consumption of and preference for ethanol of mice treated with neurotoxic doses of methamphetamine. British Journal of Pharmacology, 160(3), 772–783. 10.1111/j.1476-5381.2010.00720.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haj‐Dahmane, S. , Shen, R. Y. , Elmes, M. W. , Studholme, K. , Kanjiya, M. P. , Bogdan, D. , … Kaczocha, M. (2018). Fatty‐acid‐binding protein 5 controls retrograde endocannabinoid signaling at central glutamate synapses. Proceedings of the National Academy of Sciences of the United States of America, 115(13), 3482–3487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen, H. H. , Krutz, B. , Sifringer, M. , Stefovska, V. , Bittigau, P. , Pragst, F. , … Ikonomidou, C. (2008). Cannabinoids enhance susceptibility of immature brain to ethanol neurotoxicity. Annals of Neurology, 64(1), 42–52. 10.1002/ana.21287 [DOI] [PubMed] [Google Scholar]
- Hansson, A. C. , Bermudez‐Silva, F. J. , Malinen, H. , Hyytia, P. , Sanchez‐Vera, I. , Rimondini, R. , … Heilig, M. (2007). Genetic impairment of frontocortical endocannabinoid degradation and high alcohol preference. Neuropsychopharmacology, 32, 117–126. 10.1038/sj.npp.1301034 [DOI] [PubMed] [Google Scholar]
- Harding, S. D. , Sharman, J. L. , Faccenda, E. , Southan, C. , Pawson, A. J. , Ireland, S. , … NC‐IUPHAR (2018). The IUPHAR/BPS Guide to PHARMACOLOGY in 2018: Updates and expansion to encompass the new guide to IMMUNOPHARMACOLOGY. Nucleic Acids Research, 46(D1), D1091–D1106. 10.1093/nar/gkx1121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harkany, T. , Guzman, M. , Galve‐Roperh, I. , Berghuis, P. , Devi, L. A. , & Mackie, K. (2007). The emerging functions of endocannabinoid signaling during CNS development. Trends in Pharmacological Sciences, 28(2), 83–92. 10.1016/j.tips.2006.12.004 [DOI] [PubMed] [Google Scholar]
- Harlan, B. A. , Becker, H. C. , Woodward, J. J. , & Riegel, A. C. (2018). Opposing actions of CRF‐R1 and CB1 receptors on VTA‐GABAergic plasticity following chronic exposure to ethanol. Neuropsychopharmacology, 43(10), 2064–2074. 10.1038/s41386-018-0106-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hausknecht, K. , Shen, Y. L. , Wang, R. X. , Haj‐Dahmane, S. , & Shen, R. Y. (2017). Prenatal ethanol exposure persistently alters endocannabinoid signaling and endocannabinoid‐mediated excitatory synaptic plasticity in ventral tegmental area dopamine neurons. The Journal of Neuroscience, 37(24), 5798–5808. 10.1523/JNEUROSCI.3894-16.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henricks, A. M. , Berger, A. L. , Lugo, J. M. , Baxter‐Potter, L. N. , Bieniasz, K. V. , Petrie, G. , … McLaughlin, R. J. (2017). Sex‐ and hormone‐dependent alterations in alcohol withdrawal‐induced anxiety and corticolimbic endocannabinoid signaling. Neuropharmacology, 124, 121–133. 10.1016/j.neuropharm.2017.05.023 [DOI] [PubMed] [Google Scholar]
- Herkenham, M. , Lynn, A. B. , Johnson, M. R. , Melvin, L. S. , de Cost, B. R. , & Rice, K. C. (1991). Characterization and localization of cannabinoid receptors in rat brain: A quantitative in vitro autoradiographic study. The Journal of Neuroscience, 16, 8057–8066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Houchi, H. , Babovic, D. , Pierrefiche, O. , Ledent, C. , Daoust, M. , & Naassila, M. (2005). CB1 receptor knockout mice display reduced ethanol‐induced conditioned place preference and increased striatal dopamine D2 receptors. Neuropsychopharmacology, 30(2), 339–349. 10.1038/sj.npp.1300568 [DOI] [PubMed] [Google Scholar]
- Howlett, A. C. , Bidaut‐Russell, M. , Devane, W. A. , Melvin, L. S. , Johnson, M. R. , & Herkenham, M. (1990). The cannabinoid receptor: Biochemical, anatomical and behavioral characterization. Trends in Neurosciences, 13(10), 420–423. 10.1016/0166-2236(90)90124-S [DOI] [PubMed] [Google Scholar]
- Hungund, B. L. , & Basavarajappa, B. S. (2000). Distinct differences in the cannabinoid receptor binding in the brain of C57BL/6 and DBA/2 mice, selected for their differences in voluntary ethanol consumption. Journal of Neuroscience Research, 60, 122–128. [DOI] [PubMed] [Google Scholar]
- Hungund, B. L. , Szakall, I. , Adam, A. , Basavarajappa, B. S. , & Vadasz, C. (2003). Cannabinoid CB1 receptor knockout mice exhibit markedly reduced voluntary alcohol consumption and lack alcohol‐induced dopamine release in the nucleus accumbens. Journal of Neurochemistry, 84(4), 698–704. 10.1046/j.1471-4159.2003.01576.x [DOI] [PubMed] [Google Scholar]
- Hutchison, K. E. , Haughey, H. , Niculescu, M. , Schacht, J. , Kaiser, A. , Stitzel, J. , … Filbey, F. (2008). The incentive salience of alcohol: Translating the effects of genetic variant in CNR1. Archives of General Psychiatry, 65(7), 841–850. 10.1001/archpsyc.65.7.841 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikonomidou, C. , Bosch, F. , Miksa, M. , Bittigau, P. , Vöckler, J. , Dikranian, K. , … Olney, J. W. (1999). Blockade of NMDA receptors and apoptotic neurodegeneration in the developing brain. Science, 283(5398), 70–74. 10.1126/science.283.5398.70 [DOI] [PubMed] [Google Scholar]
- Ikonomidou, C. , Stefovska, V. , & Turski, L. (2000). Neuronal death enhanced by N‐methyl‐D‐aspartate antagonists. Proceedings of the National Academy of Sciences of the United States of America, 97(23), 12885–12890. 10.1073/pnas.220412197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Insel, T. R. (1995). The development of brain and behavior In Bloom F. E., & Kupfer D. J. (Eds.), Psychopharmacology: The four generation of progress (pp. 683–694). New York: Raven Press. [Google Scholar]
- Ishiguro, H. , Iwasaki, S. , Teasenfitz, L. , Higuchi, S. , Horiuchi, Y. , Saito, T. , … Onaivi, E. S. (2007). Involvement of cannabinoid CB2 receptor in alcohol preference in mice and alcoholism in humans. The Pharmacogenomics Journal, 7(6), 380–385. 10.1038/sj.tpj.6500431 [DOI] [PubMed] [Google Scholar]
- Johnson, J. P. , Muhleman, D. , MacMurray, J. , Gade, R. , Verde, R. , Ask, M. , … Comings, D. E. (1997). Association between the cannabinoid receptor gene (CNR1) and the P300 event‐related potential. Molecular Psychiatry, 2(2), 169–171. 10.1038/sj.mp.4000246 [DOI] [PubMed] [Google Scholar]
- Jones, R. T. , & Stone, G. C. (1970). Psychological studies of marijuana and alcohol in man. Psychopharmacologia, 18(1), 108–117. 10.1007/BF00402390 [DOI] [PubMed] [Google Scholar]
- Joshi, V. , Subbanna, S. , Shivakumar, M. , & Basavarajappa, B. S. (2019). CB1R regulates CDK5 signaling and epigenetically controls Rac1 expression contributing to neurobehavioral abnormalities in mice postnatally exposed to ethanol. Neuropsychopharmacology, 44(3), 514–525. 10.1038/s41386-018-0181-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlsson, R. M. , Adermark, L. , Molander, A. , Perreau‐Lenz, S. , Singley, E. , Solomon, M. , … Heilig, M. (2012). Reduced alcohol intake and reward associated with impaired endocannabinoid signaling in mice with a deletion of the glutamate transporter GLAST. Neuropharmacology, 63(2), 181–189. 10.1016/j.neuropharm.2012.01.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelm, M. K. , Criswell, H. E. , & Breese, G. R. (2008). The role of protein kinase A in the ethanol‐induced increase in spontaneous GABA release onto cerebellar Purkinje neurons. Journal of Neurophysiology, 100(6), 3417–3428. 10.1152/jn.90970.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemos, J. I. , Takahashi, R. N. , & Morato, G. S. (2007). Effects of SR141716 and WIN 55,212‐2 on tolerance to ethanol in rats using the acute and rapid procedures. Psychopharmacology, 194(2), 139–149. 10.1007/s00213-007-0804-1 [DOI] [PubMed] [Google Scholar]
- Li, Y. , & Kim, J. (2015). Neuronal expression of CB2 cannabinoid receptor mRNAs in the mouse hippocampus. Neuroscience, 311, 253–267. 10.1016/j.neuroscience.2015.10.041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Y. , & Kim, J. (2016). Deletion of CB2 cannabinoid receptors reduces synaptic transmission and long‐term potentiation in the mouse hippocampus. Hippocampus, 26(3), 275–281. 10.1002/hipo.22558 [DOI] [PubMed] [Google Scholar]
- Linsenbardt, D. N. , & Boehm, S. L. 2nd (2009). Agonism of the endocannabinoid system modulates binge‐like alcohol intake in male C57BL/6J mice: Involvement of the posterior ventral tegmental area. Neuroscience, 164(2), 424–434. 10.1016/j.neuroscience.2009.08.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez‐Moreno, J. A. , Gonzalez‐Cuevas, G. , Rodriguez de Fonseca, F. , & Navarro, M. (2004). Long‐lasting increase of alcohol relapse by the cannabinoid receptor agonist WIN 55,212‐2 during alcohol deprivation. The Journal of Neuroscience, 24(38), 8245–8252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez‐Moreno, J. A. , Scherma, M. , Rodriguez de Fonseca, F. , Gonzalez‐Cuevas, G. , Fratta, W. , & Navarro, M. (2008). Changed accumbal responsiveness to alcohol in rats pre‐treated with nicotine or the cannabinoid receptor agonist WIN 55,212‐2. Neuroscience Letters, 433(1), 1–5. 10.1016/j.neulet.2007.11.074 [DOI] [PubMed] [Google Scholar]
- Lu, Y. , & Anderson, H. D. (2017). Cannabinoid signaling in health and disease. Canadian Journal of Physiology and Pharmacology, 95(4), 311–327. 10.1139/cjpp-2016-0346 [DOI] [PubMed] [Google Scholar]
- Mameli, M. , Zamudio, P. A. , Carta, M. , & Valenzuela, C. F. (2005). Developmentally regulated actions of alcohol on hippocampal glutamatergic transmission. The Journal of Neuroscience, 25(35), 8027–8036. 10.1523/JNEUROSCI.2434-05.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcos, M. , Pastor, I. , de la Calle, C. , Barrio‐Real, L. , Laso, F. J. , & Gonzalez‐Sarmiento, R. (2012). Cannabinoid receptor 1 gene is associated with alcohol dependence. Alcoholism, Clinical and Experimental Research, 36(2), 267–271. 10.1111/j.1530-0277.2011.01623.x [DOI] [PubMed] [Google Scholar]
- Marcus, D. J. , Henderson‐Redmond, A. N. , Gonek, M. , Zee, M. L. , Farnsworth, J. C. , Amin, R. A. , … Morgan, D. J. (2017). Mice expressing a “hyper‐sensitive” form of the CB1 cannabinoid receptor (CB1) show modestly enhanced alcohol preference and consumption. PLoS ONE, 12(4), e0174826 10.1371/journal.pone.0174826 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marinelli, S. , Pacioni, S. , Cannich, A. , Marsicano, G. , & Bacci, A. (2009). Self‐modulation of neocortical pyramidal neurons by endocannabinoids. Nature Neuroscience, 12(12), 1488–1490. 10.1038/nn.2430 [DOI] [PubMed] [Google Scholar]
- Marks, D. F. , & MacAvoy, M. G. (1989). Divided attention performance in cannabis users and non‐users following alcohol and cannabis separately and in combination. Psychopharmacology, 99(3), 397–401. 10.1007/BF00445566 [DOI] [PubMed] [Google Scholar]
- Matsuda, L. A. , Lolait, S. J. , Brownstein, M. J. , Young, A. C. , & Bonner, T. I. (1990). Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature, 346, 561–564. [DOI] [PubMed] [Google Scholar]
- Melis, M. , Sagheddu, C. , De Felice, M. , Casti, A. , Madeddu, C. , Spiga, S. , … Pistis, M. (2014). Enhanced endocannabinoid‐mediated modulation of rostromedial tegmental nucleus drive onto dopamine neurons in Sardinian alcohol‐preferring rats. The Journal of Neuroscience, 34(38), 12716–12724. 10.1523/JNEUROSCI.1844-14.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mironov, S. L. , & Hermann, A. (1996). Ethanol actions on the mechanisms of Ca2+ mobilization in rat hippocampal cells are mediated by protein kinase C. Brain Research, 714(1–2), 27–37. 10.1016/0006-8993(95)01456-X [DOI] [PubMed] [Google Scholar]
- Mitrirattanakul, S. , Lopez‐Valdes, H. E. , Liang, J. , Matsuka, Y. , Mackie, K. , Faull, K. F. , & Spigelman, I. (2007). Bidirectional alterations of hippocampal cannabinoid 1 receptors and their endogenous ligands in a rat model of alcohol withdrawal and dependence. Alcoholism, Clinical and Experimental Research, 31(5), 855–867. 10.1111/j.1530-0277.2007.00366.x [DOI] [PubMed] [Google Scholar]
- Moranta, D. , Esteban, S. , & Garcia‐Sevilla, J. A. (2006). Ethanol desensitizes cannabinoid CB1 receptors modulating monoamine synthesis in the rat brain in vivo. Neuroscience Letters, 392(1–2), 58–61. 10.1016/j.neulet.2005.08.061 [DOI] [PubMed] [Google Scholar]
- Naassila, M. , Pierrefiche, O. , Ledent, C. , & Daoust, M. (2004). Decreased alcohol self‐administration and increased alcohol sensitivity and withdrawal in CB1 receptor knockout mice. Neuropharmacology, 46, 243–253. [DOI] [PubMed] [Google Scholar]
- Nagre, N. N. , Subbanna, S. , Shivakumar, M. , Psychoyos, D. , & Basavarajappa, B. S. (2015). CB1‐receptor knockout neonatal mice are protected against ethanol‐induced impairments of DNMT1, DNMT3A, and DNA methylation. Journal of Neurochemistry, 132(4), 429–442. 10.1111/jnc.13006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navarrete, F. , Garcia‐Gutierrez, M. S. , & Manzanares, J. (2018). Pharmacological regulation of cannabinoid CB2 receptor modulates the reinforcing and motivational actions of ethanol. Biochemical Pharmacology, 157, 227–234. [DOI] [PubMed] [Google Scholar]
- Onaivi, E. S. , Ishiguro, H. , Gu, S. , & Liu, Q. R. (2012). CNS effects of CB2 cannabinoid receptors: Beyond neuro‐immuno‐cannabinoid activity. Journal of Psychopharmacology, 26(1), 92–103. 10.1177/0269881111400652 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ortega‐Alvaro, A. , Aracil‐Fernandez, A. , Garcia‐Gutierrez, M. S. , Navarrete, F. , & Manzanares, J. (2011). Deletion of CB2 cannabinoid receptor induces schizophrenia‐related behaviors in mice. Neuropsychopharmacology, 36(7), 1489–1504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ortiz, S. , Oliva, J. M. , Perez, S. , Palomo, T. , & Manzanares, J. (2004). Chronic ethanol consumption regulates cannabinoid CB1 receptor gene expression in selected regions of rat brain. Alcohol and Alcoholism, 39, 88–92. [DOI] [PubMed] [Google Scholar]
- Patel, S. , Wohlfeil, E. R. , Rademacher, D. J. , Carrier, E. J. , Perry, L. J. , Kundu, A. , … Hillard, C. J. (2003). The general anesthetic propofol increases brain N‐arachidonylethanolamine (anandamide) content and inhibits fatty acid amide hydrolase. British Journal of Pharmacology, 139(5), 1005–1013. 10.1038/sj.bjp.0705334 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pava, M. J. , Blake, E. M. , Green, S. T. , Mizroch, B. J. , Mulholland, P. J. , & Woodward, J. J. (2012). Tolerance to cannabinoid‐induced behaviors in mice treated chronically with ethanol. Psychopharmacology, 219(1), 137–147. 10.1007/s00213-011-2387-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pava, M. J. , & Woodward, J. J. (2012). A review of the interactions between alcohol and the endocannabinoid system: Implications for alcohol dependence and future directions for research. Alcohol, 46(3), 185–204. 10.1016/j.alcohol.2012.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pava, M. J. , & Woodward, J. J. (2014). Chronic ethanol alters network activity and endocannabinoid signaling in the prefrontal cortex. Frontiers in Integrative Neuroscience, 8, 58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perra, S. , Pillolla, G. , Luchicchi, A. , & Pistis, M. (2008). Alcohol inhibits spontaneous activity of basolateral amygdala projection neurons in the rat: Involvement of the endocannabinoid system. Alcoholism, Clinical and Experimental Research, 32(3), 443–449. [DOI] [PubMed] [Google Scholar]
- Perra, S. , Pillolla, G. , Melis, M. , Muntoni, A. L. , Gessa, G. L. , & Pistis, M. (2005). Involvement of the endogenous cannabinoid system in the effects of alcohol in the mesolimbic reward circuit: Electrophysiological evidence in vivo. Psychopharmacology, 183(3), 368–377. 10.1007/s00213-005-0195-0 [DOI] [PubMed] [Google Scholar]
- Powers, M. S. , Breit, K. R. , & Chester, J. A. (2015). Genetic versus pharmacological assessment of the role of cannabinoid type 2 receptors in alcohol reward‐related behaviors. Alcoholism, Clinical and Experimental Research, 39(12), 2438–2446. 10.1111/acer.12894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pradier, B. , Erxlebe, E. , Markert, A. , & Racz, I. (2015). Interaction of cannabinoid receptor 2 and social environment modulates chronic alcohol consumption. Behavioural Brain Research, 287, 163–171. 10.1016/j.bbr.2015.03.051 [DOI] [PubMed] [Google Scholar]
- Reibaud, M. , Obinu, M. C. , Ledent, C. , Parmentier, M. , Bohme, G. A. , & Imperato, A. (1999). Enhancement of memory in cannabinoid CB1 receptor knock‐out mice. European Journal of Pharmacology, 379, R1–R2. 10.1016/S0014-2999(99)00496-3 [DOI] [PubMed] [Google Scholar]
- Rimondini, R. , Arlinde, C. , Sommer, W. , & Heilig, M. (2002). Long‐lasting increase in voluntary ethanol consumption and transcriptional regulation in the rat brain after intermittent exposure to alcohol. The FASEB Journal, 16, 27–35. [DOI] [PubMed] [Google Scholar]
- Rivera, P. , Blanco, E. , Bindila, L. , Alen, F. , Vargas, A. , Rubio, L. , … Suárez, J. (2015). Pharmacological activation of CB2 receptors counteracts the deleterious effect of ethanol on cell proliferation in the main neurogenic zones of the adult rat brain. Frontiers in Cellular Neuroscience, 9, 379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberto, M. , Cruz, M. , Bajo, M. , Siggins, G. R. , Parsons, L. H. , & Schweitzer, P. (2010). The endocannabinoid system tonically regulates inhibitory transmission and depresses the effect of ethanol in central amygdala. Neuropsychopharmacology, 35(9), 1962–1972. 10.1038/npp.2010.70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson, S. L. , Alexander, N. J. , Bluett, R. J. , Patel, S. , & McCool, B. A. (2016). Acute and chronic ethanol exposure differentially regulate CB1 receptor function at glutamatergic synapses in the rat basolateral amygdala. Neuropharmacology, 108, 474–484. 10.1016/j.neuropharm.2015.12.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez de Fonseca, F. , Roberts, A. J. , Bilbao, A. , Koob, G. F. , & Navarro, M. (1999). Cannabinoid receptor antagonist SR141716A decreases operant ethanol self administration in rats exposed to ethanol‐vapor chambers. Acta Pharmacologica Sinica, 20(12), 1109–1114. [PubMed] [Google Scholar]
- Rubio, M. , de Miguel, R. , Fernandez‐Ruiz, J. , Gutierrez‐Lopez, D. , Carai, M. A. , & Ramos, J. A. (2009). Effects of a short‐term exposure to alcohol in rats on FAAH enzyme and CB1 receptor in different brain areas. Drug and Alcohol Dependence, 99(1–3), 354–358. [DOI] [PubMed] [Google Scholar]
- Rubio, M. , Fernandez‐Ruiz, J. , de Miguel, R. , Maestro, B. , Michael Walker, J. , & Ramos, J. A. (2008). CB1 receptor blockade reduces the anxiogenic‐like response and ameliorates the neurochemical imbalances associated with alcohol withdrawal in rats. Neuropharmacology, 54(6), 976–988. [DOI] [PubMed] [Google Scholar]
- Rubio, M. , McHugh, D. , Fernandez‐Ruiz, J. , Bradshaw, H. , & Walker, J. M. (2007). Short‐term exposure to alcohol in rats affects brain levels of anandamide, other N‐acylethanolamines and 2‐arachidonoyl‐glycerol. Neuroscience Letters, 421(3), 270–274. 10.1016/j.neulet.2007.05.052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt, L. G. , Samochowiec, J. , Finckh, U. , Fiszer‐Piosik, E. , Horodnicki, J. , Wendel, B. , … Hoehe, M. R. (2002). Association of a CB1 cannabinoid receptor gene (CNR1) polymorphism with severe alcohol dependence. Drug and Alcohol Dependence, 65(3), 221–224. [DOI] [PubMed] [Google Scholar]
- Schneider, M. (2009). Cannabis use in pregnancy and early life and its consequences: Animal models. European Archives of Psychiatry and Clinical Neuroscience, 259(7), 383–393. 10.1007/s00406-009-0026-0 [DOI] [PubMed] [Google Scholar]
- Serrano, A. , & Parsons, L. H. (2011). Endocannabinoid influence in drug reinforcement, dependence and addiction‐related behaviors. Pharmacology & Therapeutics, 132(3), 215–241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serrano, A. , Pavon, F. J. , Buczynski, M. W. , Schlosburg, J. , Natividad, L. A. , Polis, I. Y. , … Parsons, L. H. (2018). Deficient endocannabinoid signaling in the central amygdala contributes to alcohol dependence‐related anxiety‐like behavior and excessive alcohol intake. Neuropsychopharmacology, 43, 1840–1850. 10.1038/s41386-018-0055-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sloan, M. E. , Gowin, J. L. , Yan, J. , Schwandt, M. L. , Spagnolo, P. A. , Sun, H. , … Ramchandani, V. A. (2018). Severity of alcohol dependence is associated with the fatty acid amide hydrolase Pro129Thr missense variant. Addiction Biology, 23(1), 474–484. 10.1111/adb.12491 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soyka, M. , Koller, G. , Schmidt, P. , Lesch, O. M. , Leweke, M. , Fehr, C. , … Mann, K. F. (2008). Cannabinoid receptor 1 blocker rimonabant (SR 141716) for treatment of alcohol dependence: Results from a placebo‐controlled, double‐blind trial. Journal of Clinical Psychopharmacology, 28(3), 317–324. 10.1097/JCP.0b013e318172b8bc [DOI] [PubMed] [Google Scholar]
- Spagnolo, P. A. , Ramchandani, V. A. , Schwandt, M. L. , Kwako, L. E. , George, D. T. , Mayo, L. M. , … Heilig, M. (2016). FAAH gene variation moderates stress response and symptom severity in patients with posttraumatic stress disorder and comorbid alcohol dependence. Alcoholism, Clinical and Experimental Research, 40(11), 2426–2434. 10.1111/acer.13210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stefanis, N. C. , Delespaul, P. , Henquet, C. , Bakoula, C. , Stefanis, C. N. , & Van Os, J. (2004). Early adolescent cannabis exposure and positive and negative dimensions of psychosis. Addiction, 99(10), 1333–1341. 10.1111/j.1360-0443.2004.00806.x [DOI] [PubMed] [Google Scholar]
- Stopponi, S. , Fotio, Y. , Domi, A. , Borruto, A. M. , Natividad, L. , Roberto, M. , … Cannella, N. (2017). Inhibition of fatty acid amide hydrolase in the central amygdala alleviates co‐morbid expression of innate anxiety and excessive alcohol intake. Addiction Biology, 23(6), 1223–1232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbanna, S. , & Basavarajappa, B. S. (2014). Pre‐administration of G9a/GLP inhibitor during synaptogenesis prevents postnatal ethanol‐induced LTP deficits and neurobehavioral abnormalities in adult mice. Experimental Neurology, 261, 34–43. 10.1016/j.expneurol.2014.07.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbanna, S. , Joshi, V. , & Basavarajappa, B. S. (2018). Activity‐dependent signaling and epigenetic abnormalities in mice exposed to postnatal ethanol. Neuroscience, 392, 230–240. 10.1016/j.neuroscience.2018.07.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbanna, S. , Nagre, N. N. , Shivakumar, M. , Joshi, V. , Psychoyos, D. , Kutlar, A. , … Basavarajappa, B. S. (2018). CB1R‐mediated activation of caspase‐3 causes epigenetic and neurobehavioral abnormalities in postnatal ethanol‐exposed mice. Frontiers in Molecular Neuroscience, 11, 45 10.3389/fnmol.2018.00045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbanna, S. , Nagre, N. N. , Shivakumar, M. , Umapathy, N. S. , Psychoyos, D. , & Basavarajappa, B. S. (2014). Ethanol induced acetylation of histone at G9a exon1 and G9a‐mediated histone H3 dimethylation leads to neurodegeneration in neonatal mice. Neuroscience, 258, 422–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbanna, S. , Nagre, N. N. , Umapathy, N. S. , Pace, B. S. , & Basavarajappa, B. S. (2015). Ethanol exposure induces neonatal neurodegeneration by enhancing CB1R Exon1 histone H4K8 acetylation and up‐regulating CB1R function causing neurobehavioral abnormalities in adult mice. The International Journal of Neuropsychopharmacology, 18(5), 1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbanna, S. , Shivakumar, M. , Psychoyos, D. , Xie, S. , & Basavarajappa, B. S. (2013). Anandamide‐CB1 receptor signaling contributes to postnatal ethanol‐induced neonatal neurodegeneration, adult synaptic and memory deficits. Journal of Neuroscience, 33(15), 6350–6366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talani, G. , & Lovinger, D. M. (2015). Interactions between ethanol and the endocannabinoid system at GABAergic synapses on basolateral amygdala principal neurons. Alcohol, 49(8), 781–794. 10.1016/j.alcohol.2015.08.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Twitchell, W. , Brown, S. , & Mackie, K. (1997). Cannabinoids inhibit N‐ and P/Q‐type calcium channels in cultured rat hippocampal neurons. Journal of Neurophysiology, 78(1), 43–50. 10.1152/jn.1997.78.1.43 [DOI] [PubMed] [Google Scholar]
- van den Wildenberg, E. , Janssen, R. G. , Hutchison, K. E. , van Breukelen, G. J. , & Wiers, R. W. (2007). Polymorphisms of the dopamine D4 receptor gene (DRD4 VNTR) and cannabinoid CB1 receptor gene (CNR1) are not strongly related to cue‐reactivity after alcohol exposure. Addiction Biology, 12(2), 210–220. [DOI] [PubMed] [Google Scholar]
- Varodayan, F. P. , Bajo, M. , Soni, N. , Luu, G. , Madamba, S. G. , Schweitzer, P. , & Roberto, M. (2017). Chronic alcohol exposure disrupts CB1 regulation of GABAergic transmission in the rat basolateral amygdala. Addiction Biology, 22(3), 766–778. 10.1111/adb.12369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varodayan, F. P. , Soni, N. , Bajo, M. , Luu, G. , Madamba, S. G. , Schweitzer, P. , … Roberto, M. (2016). Chronic ethanol exposure decreases CB1 receptor function at GABAergic synapses in the rat central amygdala. Addiction Biology, 21(4), 788–801. 10.1111/adb.12256 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinod, K. Y. , Kassir, S. A. , Hungund, B. L. , Cooper, T. B. , Mann, J. J. , & Arango, V. (2010). Selective alterations of the CB1 receptors and the fatty acid amide hydrolase in the ventral striatum of alcoholics and suicides. Journal of Psychiatric Research, 44(9), 591–597. 10.1016/j.jpsychires.2009.11.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinod, K. Y. , Yalamanchili, R. , Thanos, P. K. , Vadasz, C. , Cooper, T. B. , Volkow, N. D. , & Hungund, B. L. (2008). Genetic and pharmacological manipulations of the CB1 receptor alter ethanol preference and dependence in ethanol preferring and nonpreferring mice. Synapse, 62(8), 574–581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinod, K. Y. , Yalamanchili, R. , Xie, S. , Cooper, T. B. , & Hungund, B. L. (2006). Effect of chronic ethanol exposure and its withdrawal on the endocannabinoid system. Neurochemistry International, 49(6), 619–625. 10.1016/j.neuint.2006.05.002 [DOI] [PubMed] [Google Scholar]
- Viscomi, M. T. , Oddi, S. , Latini, L. , Pasquariello, N. , Florenzano, F. , Bernardi, G. , … Maccarrone, M. (2009). Selective CB2 receptor agonism protects central neurons from remote axotomy‐induced apoptosis through the PI3K/Akt pathway. The Journal of Neuroscience, 29(14), 4564–4570. 10.1523/JNEUROSCI.0786-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Viudez‐Martinez, A. , Garcia‐Gutierrez, M. S. , Navarron, C. M. , Morales‐Calero, M. I. , Navarrete, F. , Torres‐Suarez, A. I. , & Manzanares, J. (2018). Cannabidiol reduces ethanol consumption, motivation and relapse in mice. Addiction Biology, 23(1), 154–164. 10.1111/adb.12495 [DOI] [PubMed] [Google Scholar]
- Wang, L. , Liu, J. , Harvey‐white, J. , Zimmer, A. , & Kunos, G. (2003). Endocannabinoid signaling via CB1 receptors is involved in ethanol preference and its age‐dependent decline in mice. Proceedings of the National Academy of Sciences of the United States of America, 100, 1393–1398. 10.1073/pnas.0336351100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warnault, V. , Houchi, H. , Barbier, E. , Pierrefiche, O. , Vilpoux, C. , Ledent, C. , … Naassila, M. (2007). The lack of CB1 receptors prevents neuroadapatations of both NMDA and GABAA receptors after chronic ethanol exposure. Journal of Neurochemistry, 102(3), 741–752. 10.1111/j.1471-4159.2007.04577.x [DOI] [PubMed] [Google Scholar]
- Wilson, D. A. , Peterson, J. , Basavaraj, B. S. , & Saito, M. (2011). Local and regional network function in behaviorally relevant cortical circuits of adult mice following postnatal alcohol exposure. Alcoholism: Clinical and Experimental Research, 35, 1974–1984. 10.1111/j.1530-0277.2011.01549.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu, C. S. , Jew, C. P. , & Lu, H. C. (2011). Lasting impacts of prenatal cannabis exposure and the role of endogenous cannabinoids in the developing brain. Future Neurology, 6(4), 459–480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, H. Y. , Gao, M. , Shen, H. , Bi, G. H. , Yang, H. J. , Liu, Q. R. , … Xi, Z. X. (2016). Expression of functional cannabinoid CB receptor in VTA dopamine neurons in rats. Addiction Biology, 22(3), 752–765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng, L. , Wu, X. , Dong, X. , Ding, X. , & Song, C. (2015). Effects of chronic alcohol exposure on the modulation of ischemia‐induced glutamate release via cannabinoid receptors in the dorsal hippocampus. Alcoholism, Clinical and Experimental Research, 39(10), 1908–1916. 10.1111/acer.12845 [DOI] [PubMed] [Google Scholar]
- Zhou, Y. , Schwartz, B. I. , Giza, J. , Gross, S. S. , Lee, F. S. , & Kreek, M. J. (2017). Blockade of alcohol escalation and “relapse” drinking by pharmacological FAAH inhibition in male and female C57BL/6J mice. Psychopharmacology, 234(19), 2955–2970. 10.1007/s00213-017-4691-9 [DOI] [PMC free article] [PubMed] [Google Scholar]