

Abstract
Background
Mangrove forests provide many ecosystem services, including the provision of habitat that supports avian biodiversity. However, hurricanes can knock down trees, alter hydrologic connectivity, and affect avian habitat. In 1995, Hurricanes Opal and Roxanne destroyed approximately 1,700 ha of mangrove forest in Laguna de Términos, Mexico. Since then, hydrological restoration has been implemented to protect the mangrove forest and its biodiversity.
Methods
Since avian communities are often considered biological indicators of ecosystem quality, avian diversity and species relative abundance were evaluated as indicators of mangrove restoration success by comparing undisturbed mangrove patches with those affected by the hurricanes. Using bird surveys, similarity analyses, and generalized linear models, we evaluated the effects of water quality variables and forest structure on the relative abundance and diversity of the avian community in disturbed, restored, and undisturbed mangrove patches.
Results
Higher bird species richness and relative abundances were found in disturbed and restored sites compared to the undisturbed site. After restoration, values of frequency of flooding, water temperature, tree density, and the number of tree species were more similar to that of the undisturbed site than to the values of the disturbed one. Such variables influenced the relative abundance of bird guilds in the different habitat conditions. Furthermore, some insectivorous bird species, such as the Yellow Warbler and Tropical Kingbird, were found to be similarly abundant in both undisturbed and restored sites, but absent or very low in occurrence at the disturbed site.
Conclusions
Collectively, our results strongly suggest that hydrologic restoration helps to enhance niche availability for different bird guilds, including water and canopy bird species. Our work can help inform management strategies that benefit avian communities in mangrove forests and wetland systems.
Keywords: Hydroperiod, Biodiversity, Rehabilitation, Coastal wetland, Hydrology, Laguna de terminos, Birds, Ecological indicator
Introduction
Mangrove forests provide valuable ecosystem services. These highly productive ecosystems can prevent erosion, trap sediments, and provide wind protection for coastal communities (Warren-Rhodes et al., 2011). Mangrove forests also support important nursery habitats for a variety of organisms including pelagic and marine vertebrates and invertebrates, mammals, and birds (Nagelkerken et al., 2008; Lee et al., 2014; Serafy et al., 2015). In Mexico, mangroves provide important habitats for specialist species such as the Mangrove Cuckoo (Coccizus minor), the Mangrove Finch (Camarhynchus heliobates), the Mangrove Hummingbird (Amazilia buocardi), and the Mangrove Warbler (Setophaga petechia bryanti) (Polidoro et al., 2010; Andrade et al., 2012; Gardner et al., 2012; Buelow & Sheaves, 2015). Mangrove forests can also improve water quality (Wang et al., 2010), and provide economically important products. Nevertheless, it is estimated that about 35% of the global mangrove cover has been lost, mainly due to forest clearance for fish farming, urbanization, habitat fragmentation, and fuel and timber production (Valiela, Bowen & York, 2001; Alongi, 2008; Donato et al., 2011).
In Mexico, four mangrove species are distributed along the Gulf of Mexico coast: red (Rhizophora mangle), black (Avicennia germinans), white (Laguncularia racemosa), and button (Conocarpus erectus) mangroves, covering approximately 742,000 hectares (ha) (Giri et al., 2011). The estimated value of mangroves due to fisheries, carbon sequestration, forestry coastal protection, erosion control, water treatment and other environmental, recreational and traditional uses is about $80,000 to $194,000 USD per ha−1yr−1 (Salem & Mercer, 2012; Costanza et al., 2014). Such a value represents up to a total of $59–143 billion USD for the Mexican mangroves. Globally, Mexico ranks fourth in terms of total mangrove area (Giri et al., 2011); however, as in other parts of the world, the rate of degradation of mangrove forests in Mexico has been high in the past century. Mangrove cover has been reduced by approximately 10% of the original area in the last 40 years, and about 2% (15,000 ha) of the area that remains has been classified as disturbed (Valderrama et al., 2014). In most mangrove forests, there has been a loss of connectivity and a decrease in heterogeneity, which has reduced faunal diversity, including avian populations (Mohd-Azlan, Noske & Lawes, 2015; Hauser et al., 2017; Amir, 2018).
Because restoration can help mitigate climate change effects and the consequential biodiversity loss (Nilsson & Aradottir, 2013), interest in the ecological restoration of coastal wetlands is currently growing (Palmer, Hondula & Koch, 2014; Suding et al., 2015). The disturbance of coastal wetlands is often accompanied by changes in hydrological patterns, including the hydroperiod, which is defined as the amount of time, the frequency, and the level with which a wetland is covered by water, a key factor determining success in wetland restoration (Turner & Lewis, 1996; Zaldívar-Jiménez et al., 2010; Wortley, Hero & Howes, 2013). For instance, sediment deposition in the tidal channels affects the amount of time and the frequency at which a mangrove is flooded (Woodroffe et al., 2016). These kinds of changes affect the water quality and the composition of plant and animal communities (Schaffelke, Mellors & Duke, 2005; Krauss et al., 2006; Crase et al., 2013). Hence, the success of ecological restoration can be assessed through a system of indicators that generate information about the recovery of wetland functions. Such a system might include landscape features, biogeochemical processes (Cvetkovic & Chow-Fraser, 2011; Zhang et al., 2012), ecosystem services, and the composition of biological assemblages (e.g., vegetation, crustaceans, mollusk, and vertebrates) (Thornton & Johnstone, 2015; Salmo, Tibbetts & Duke, 2016). Therefore, species richness, abundance, and community structure can be used to evaluate the biological outcomes of restoration efforts (Zhao et al., 2016).
Because of their diverse roles within the trophic webs, bird communities are key elements for describing the energetics of ecosystems (Wenny et al., 2011; Adame et al., 2015; McFadden, Kauffman & Bhomia, 2016), and are considered useful indicators of ecosystem health (Canterbury et al., 2000; Bryce, Hughes & Kaufmann, 2002; Catterall et al., 2012). For instance, monitoring of pollutants such as DDT and organochlorines, and the effects of variations in hydroperiod in the Everglades were carried out using water and wading bird species (Frederick et al., 2009; Lavoie et al., 2010; Lantz, Gawlik & Cook, 2011; Boyle, Dorn & Cook, 2012). Moreover, bird diversity has been used as an indicator of temporal changes in mangrove health (Behrouzi-Rad, 2014), and to assess the impacts of climate change and coastal development (Ogden et al., 2014). While avian community monitoring can be a useful tool for evaluating the health of wetland ecosystems, its use for assessment of ecological restoration has been rarely employed (Weller, 1995; Cui et al., 2009; Gyurácz, Bánhidi & Csuka, 2011; Li et al., 2011; Catterall et al., 2012; Zou et al., 2014).
Laguna de Términos is a coastal lagoon located in Mexico along the southwestern coast of the Gulf of Mexico. It is the second largest coastal lagoon in Mexico, supporting approximately 259,000 ha of mangrove forest (33% of all mangrove forest in Mexico) and 262 bird species, among other vertebrate taxa (Villalobos-Zapata & Mendoza-Vega, 2010). As a result, Laguna de Términos has been recognized as a Ramsar wetland of international importance (Chape, Spalding & Jenkins, 2008). However, due to fisheries, oil-extraction activities, illegal timber exploitation, and urbanization, nearly 26% of the mangrove habitat in this lagoon is considered degraded (Zaldívar-Jiménez et al., 2017). Additionally, in 1995, the lagoon was affected by Hurricanes Opal (category 4) and Roxana (category 3), which destroyed 1,700 ha of mangrove (Pérez-Ceballos et al., 2013). Fallen trees blocked some creeks (hereafter tidal channels), mainly the secondary ones. The silting of channels altered the hydroperiod patterns and biogeochemical conditions, leading to the mortality of adult trees and inhibiting the natural regeneration of the mangrove. Moreover, this condition has led to a constantly increasing area of dead trees around the points affected by the hurricanes. In order to increase the resilience of these mangrove forests, protect their biological diversity, educate others, and contribute to the sustainable development of the adjacent local communities, from October 2010 to November 2012, restoration activities were implemented. In brief, restoration activities included an environmental and social diagnosis, as well as the formulation of a management plan before the restoration implementation. The primary restoration activity was the desilting and unblocking of the main and secondary tidal channels. Where needed, new secondary channels were dug based on the microtopography analysis of each site selected for restoration. Desilting, unblocking, and channel digging were carried out by local women and men only by hand.
The objective of this study was to use the changes on the avian abundance and diversity as indicators of the restoration success of a mangrove site. We specifically asked: (1) whether the avian community structure differs between not obviously affected mangrove patches and those affected by the hurricanes; (2) whether hydroperiod and water quality influence the diversity or the abundance of birds; and (3) which species could be used as indicator species of post-restoration recovery. The sampling was carried out on a landscape mosaic with different strata: undisturbed patches, patches without restoration, and patches three years after restoration. The water quality variation among the study sites, along with the avian community diversity and abundance of species, were evaluated as indicators of mangrove restoration success.
Materials & Methods
Study area
Laguna de Términos is a coastal wetland located in Campeche, Mexico. It covers about 150 km2 and is connected to the Gulf of Mexico by two inlets at the east and west sides of Isla del Carmen, a calcareous sandbar that supports 5,900 ha of mangrove, of which around 26% are disturbed. The annual average rainfall and temperature are approximately 1,420 mm and 27 ° C respectively (David & Kjerfve, 1998). The study area is an estuary known as Bahamitas, located on the east of Isla del Carmen (637787.52E, 2066226.35N, and 633872.60E, 2064181.96N UTM Q15, Fig. 1). The hydrological restoration was implemented from October 2010 to November 2012 through desilting and unblocking of natural tidal channels. Where needed, new secondary channels were created to induce natural regeneration of the vegetation, and to enhance the water quality (oxygenation and salinity) of 1,300 ha of disturbed mangrove through water exchange (Zaldívar-Jiménez et al., 2017). The creation of new tidal channels relied on the modeling of hydrological flow-paths after analysis of a digital elevation model created from the obtained microtopography data. All the work was undertaken by hand using shovels, and involved local inhabitants (10 people ha−1), who were trained through workshops on habitat restoration and environmental education, as well as being advised on social and community organization for sustainable development through bird watching and catch-and-release fishing (Zaldívar-Jiménez et al., 2017).
Figure 1. Location of Bahamitas estuaries with different disturbance levels in Laguna de Términos, Campeche Mexico.

Sampling sites and forest structure
Based on the digital analysis of a Worldview 2013 image and subsequent field surveys, three different patches for sampling were identified and selected according to their condition of degradation: (i) First were undisturbed sites with well-established adult trees and no significant human activities. Here, at least 80% of the trees were alive and there was no evident alteration of the hydrological connectivity due to the effects of hurricanes. (ii) Second were disturbed sites with all the trees dead, except for some few individuals at the edges, and no seedling establishment or with no more than 10% of scrub mangroves alive. (iii) Third were restored sites that showed similar conditions to the degraded site prior to restoration activities implemented during 2010 and 2012 to allow water to flow in and out through the topography of the wetland. After restoration (three years), this site already showed the establishment of seedlings and some saplings, and no more than 50% of the scrub mangroves were dead.
For each sampling site, two 10 m × 10 m random sampling plots were surveyed to determine the forest structure and the number of live trees. On the same plots, measures of diameter at breast height (DBH), canopy height, and basal area were collected to determine the forest cover. Tree density was estimated by counts of all trees with DBH >2.5 cm (Schaeffer-Novelli, Vale & Cintrón, 2015). These data were used to investigate the effects of the vegetation structure on bird abundance in each sampling site (McElhinny et al., 2005; Azhar et al., 2013).
Hydrologic and water quality parameters
To assess the relationship of bird abundance with the water quality parameters, 11 sampling points were established every 100 m within each survey site to measure the water depth, as well as the temperature and salinity of surficial water of the tidal channels. Water depth was measured using a ruler at the center of the tidal channels. Temperature and salinity were recorded using a parametric probe, YSI-30 (YSI Incorporated, Yellow Springs, OH, USA). The pH and oxidation reduction potential (ORP) were recorded using a portable tester, HI916 (Hanna Instruments, Inc., Woonsocket, RI, USA). All these data were recorded twice monthly. The hydroperiod patterns among the sampling sites were also compared by contrasting the tidal range (the level of the flooding in cm), the flooding duration (the time in hours that a site stand flooded), and the flooding frequency (the number of times per month that a place floods). These measures were recorded during the entire sampling period every 60 min using a HOBO U20-001-01-Ti logger placed at each sampling site.
Birds survey
To estimate species richness and abundance, monthly bird surveys (December 2014 to June 2015) were conducted by boat. This period of time was selected because it allows finding both migrant birds in winter and residents species in spring and early summer. Counting points (n = 11 for each sampling site) were used to record bird species and count individuals per species within each sampling site. We carried out a systematic sampling with a randomly selected start point at each sampling area. Then, the next ten counting points were separated 100 m from each other. Because a count at a particular point can be affected by whether the neighboring points are above or below their averages (Pendleton, 1995), we tested for independence of our counting points through a Pearson’s test of conditional independence. Additionally, we used the function Moran included in the R package ape (Paradis, Claude & Strimmer, 2004) to treat the abundance data across counting points with a Moran’s index test to ensure that they were not spatially biased. As recommended for field methods in bird surveys (Gregory, Gibbons & Donald, 2004), we used a minimum of four visits to each counting point (four in winter and four in spring). To avoid double count, a single observer counted individual birds at standard intervals of time (10 min). To help deal with varying detectability of different species, observations were made for two consecutive days (Bibby et al., 1992; Gregory, Gibbons & Donald, 2004; Schmidt, McIntyre & MacCluskie, 2013). To deal with species mimicry, besides song and call recognition, birds were identified by a trained observer that sought field marks using 10 × 42 binoculars. We recorded all species and individuals seen or heard within the first 20 m radius at each counting point. All species and individuals observed on the vegetation, water, or flying within the observation radius (up to 15 m above) were counted. We did not account for passing or transient birds flying on the sampling areas that did not stop there to feed or rest. The sampling effort was equivalent to approximately 77 h of surveying over the three sampling sites. All bird species observed outside the sampling radius were also recorded but not included in further analyses of species abundance. The reliability of our sampling design concerning species detectability, temporal, and size representativeness was assessed by the implementation of an evaluation framework for ecological research (Battisti, Dodaro & Franco, 2014).
Statistical analyses
The data on hydroperiod and environmental variables were explored for normality through Shapiro–Wilk tests, which are ratios of two estimates of the variance in a normal distribution calculated from a set of observations (Royston, 1995). Data that met the assumptions of normality and independence were analyzed using a one-way analysis of variance (ANOVA) to compare the means between the data from all sampling sites. When significant differences were found, ANOVA results were further examined through an honestly-significant-difference Tukey’s test, a multiple comparisons procedure to find the differences between all levels of a factor once the hypothesis of equality from the ANOVA test is rejected. When the data did not meet the assumption of normality, we used Kruskal-Wallis one-way analysis of variance by ranks, a non-parametric alternative to ANOVA for multiple comparisons. To contrast the levels of a factor from this analysis, we used the function posthoc.kruskal.nemenyi.test from the R package PMCMR, a post-hoc alternative to performing multiple comparisons for non-parametrics (Pohlert, 2014).
Species richness and abundance for each sampling site were calculated based on the number of recorded individuals per species. Diversity was estimated using the inverse of Simpson’s Index (1 Σ(n∕N)2) which indicates greater diversity as the resulting value approaches 1, while the dominance (the extent to which a taxon is more numerous than others) was assessed through the Berger-Parker Index (d = Nmax∕NT), in which the lower the value of d results in a more even dominance in the sample (for details on the formulae see Ingram (2008)). To reduce biases caused by non-detected species, the expected species richness was calculated using the Jack1 estimator, which is adequate to estimate actual species richness when the number of sampling units is small (<20 samples or individuals) or when the samples are not the same size. It uses the total number of observed species in a set of samples, the total number of unique species in each sample, and the number of samples for the calculations (Smith & Van Belle, 1984; Gotelli & Colwell, 2011). Abundance and evenness (how equal the bird community is numerically) among sampling sites were compared through rank-abundance curves. The curves were constructed as implemented in the R package BiodiversityR (Kindt & Coe, 2005). From the pooled data, the total number of individuals was calculated to obtain their abundance (y-axis) and then ranked from the most to the least abundant species (x-axis). Then, the same procedure was implemented for each sampling location (Kindt & Coe, 2005). Key bird species to particular sampling sites were identified on the basis of their abundance through a one-way Simper test carried out in PRIMER 7 (Anderson, Gorley & Clarke, 2008). The Simper test estimated the contribution of each species abundance to the total dissimilarity among the sampling sites using Bray-Curtis distances, which helps find discriminating features within habitats that explain differences in community composition (Clarke, 1993). Additionally, we estimated the indicator species index value (IndVal) to find the value of particular species to each sampling site as indicators of their condition. The IndVal uses both the relative abundance (instead of the absolute abundance), and the relative frequency of each species to estimate its value as a percentage (Roberts, 2016). We also investigated the relationship between species abundance and environmental variables by means of redundancy (RDA) and multivariate analyses through generalized linear models (glms). The RDA was selected because there was a linear response of the abundance of birds to the measured environmental variables. For both the RDA and the glms, the birds’ abundances were used as the dependent variable, and habitat levels, environmental characteristics, and forest structure were explanatory variables. The RDA test was carried out as implemented in BiodiversityR while the glms were fitted through the R package mvabund (Wang et al., 2012). The mvabund functions help test for interactions through multiple testing to predict abundance between sites or treatments. The primary function of mvabund (manyglm) fits a glm to each species in the dataset using a common group of environmental variables. This approach uses a resampling-based hypothesis testing to infer which environmental variables relate to multivariate abundances at community or taxon-specific levels (Wang et al., 2012). The independence of the abundance estimates across sampling sites, as well as the quadratic mean–variance and log linearity in the dataset, were checked through the functions plot and meanvar.plot as implemented in the same package before fitting the models. Several glms were fitted to tests for variable effects. The data set included the mangrove degradation condition as a categorical variable within the environmental matrix. We performed the same analysis but with the data grouped by functional groups according to the birds primary source of food: insectivores, nectarivores, scavengers, macroinvertivores (polychaetes, mollusks, crustaceans), frugivores, omnivores, and meat eaters (fishes, small reptilians, amphibians, and mammals). Because of the difficultly of measuring this directly in the field, this group classification was sourced from literature on local species (MacKinnon, 2013; Fagan & Komar, 2016), and from specialized web sources (https://www.allaboutbirds.org). Models were ranked according to the Akaike’s information criterion (AIC) (Burnham & Anderson, 2002; Burnham, Anderson & Huyvaert, 2011). All tests were set to be significant at 0.05 level, if not indicated otherwise.
Results
Pearson’s test of conditional independence showed that although the sampling points in each site were closely located, they were independent (χ2 = 26.166, df = 20, p = 0.1604). Similarly, no evidence of spatial autocorrelation of the abundance was detected across sampling points (Moran’s Index value = 0.0074, p = 0.3335).
Water quality parameters and forest structure
The frequency of flooding was approximately two and three times greater at the restored and undisturbed sites, respectively, than at the disturbed one (ANOVA test F2,18 = 12.47, p = 0.003, Fig. S1). However, there were no significant differences among treatments in tidal range or flooding duration (ANOVA test F2,18 = 2.259, p = 0.133; F2,18 = 2.416, p = 0.118, respectively).
Environmental variables such as salinity concentration, water depth, pH, and temperature were found to be statistically different among sites (Table 1). The mangrove structure also showed significant differences between sites in terms of the number of tree species, basal area, and tree density (Table 1). However, due to the size of the remaining live trees in the disturbed and restored sites, no statistical differences in the heights of trees between sampled sites were observed (Fig. S2). Finally, the ORP values were not statistically different between treatments. However, the ongoing water exchange due to the greater frequency of flooding in the undisturbed and restored areas might indicate higher oxygen concentration in the water (all ORP values were positive).
Table 1. Comparison of the mean values of hydroperiod, water-quality parameters and forest structure between studied areas following restoration at the restored site.
Only statistically significant results are shown.
| Variable | Disturbed | Restored | Undisturbed | Test | df | p | Post hoc |
|---|---|---|---|---|---|---|---|
| Flooding frequency (times/month) | 2.71 | 9.00 | 14.42 | F = 12.47 | 2.00 | 0. 003 |
R-D = 0.038 U-D < 0.001 |
| Flooding duration (h) | 588.14 | 404.42 | 314.71 | F = 2.41 | 2.00 | 0.118 | – |
| Tidal range (m) | 0.09 | 0.09 | 0.05 | F = 2.25 | 2.00 | 0.133 | – |
| Salinity (PSU) | 34.93 | 34.12 | 33.28 | χ2 = 10.25 | 2.00 | 0. 005 | U-D = 0.004 |
| pH | 8.15 | 8.11 | 8.02 | F = 12.32 | 2.30 | 0. 001 | U-D < 0.001 U-R = 0.023 |
| Redox potential | 82.09 | 91.18 | 85.18 | χ2 = 3.42 | 2.00 | 0.180 | – |
| Temperature (°C) | 28.02 | 29.94 | 28.87 | χ2 = 11.81 | 2.00 | 0. 002 | R-D = 0.001 |
| Depth (cm) | 54.62 | 74.96 | 117.54 | F = 31.87 | 2.00 | <0. 001 | U-D < 0.001 U-R < 0.001 R-D = 0.043 |
| No. of tree species | 1.09 | 1.36 | 1.90 | χ2 = 12.16 | 2.00 | 0. 002 | U-D = 0.008 |
| Tree density (trees/ha) | 345 | 763 | 709 | χ2 = 6.45 | 2.00 | 0. 039 | R-D = 0.035 |
| Basal area (m2) | 1.29 | 4.31 | 7.02 | χ2 = 15.39 | 2.00 | <0. 001 | U-D < 0.001 |
Notes.
In the Test column, F is for ANOVA and χ2 is for Kruskal–Wallis rank sum test. In the Post hoc column, comparisons between pairs of sites (R = restored, D = degraded, U = undisturbed), were tested for significance using either HDS Tukey’s or Nemenyi’s tests. A dash means that the test was not carried out for that variable.
Species diversity and abundance
Fifty-three avian species were recorded in the surveys across the sampling sites (Table S1). One of them, the Reddish Egret (Egretta rufescens) is near threatened while the rest are of least concern according to the IUCN (2016). Regionally, five species are under special protection, and one, the Yellow-crowned Night-heron (Nyctanassa violacea), is a threatened species (SEMARNAT, 2010). For almost half of the recorded species (49%), the populations are trending to increase while 24.5% show a decreasing trend (IUCN, 2016).
The total bird species richness was higher in the disturbed and restored than in the undisturbed site, and, for all sampling sites, the observed species richness was lower than expected (Table 2). This result is consistent with our empirical knowledge of the bird community found in the vicinity of our sampling sites. For instance, species such as the migrant Lesser Scaup (Aythya affinis), the Northern Pintail (Anas acuta), and the American Wigeon (Anas americana), as well as the resident mangrove inhabitants, the Muscovy Duck (Cairina moschata) and the White-fronted Parrot (Amazona albifrons) (both with important population declines in Mexico), were expected but not observed. Diversity indexes showed that there was a clear dominance of few species belonging to the families Ardeidae and Threskiornithidae, especially in the restored site. Even though our sampling effort captured most of the species richness in the study area, the species saturation curve did not reach a flat shape at the end of the surveys (Fig. S3). Accordingly, an increased sampling effort might be necessary to show the true richness of the study area. When pooled data were analyzed, the most abundant species were the Least Sandpiper (Calidris minutilla), the Mangrove Warbler, the Black-bellied Whistling Duck (Dendrocigna autumnalis), the Green Heron (Butorides virescens), the White Ibis (Eudocimus albus), and the Great-tailed Grackle (Quiscalus mexicanus). Because birds detected in flight were only about 10% of all counts and because there were no statistical differences in the abundance between the whole set of data vs. the data that excluded birds detected in flight (Wilcoxon’s test with continuity correction, W = 1505.5, p = 0.7716), for further analyses, we used the full dataset. Species abundance and evenness were similar for undisturbed and disturbed sites. Although the restored site showed higher bird abundance, this was also the location with the least evenness in species distribution. For instance, higher numbers of species such as the Least Sandpiper and the White Ibis, which depend on mangrove habitats with open or semi-open forest structure, were observed in the restored mangrove patches, while the Mangrove Warbler, the Tropical Kingbird (Tyrannus melancholicus), and the American White Pelican (Pelecanus erythrorhynchos) were the least abundant at this site (Fig. 2).
Table 2. Avian community diversity indexes at the study locations.
| Habitat | n | Observed richness | Mean ± SD | Expected richness | 1-Simpson | Berger parker |
|---|---|---|---|---|---|---|
| Undisturbed | 11 | 28 | 5.27 ± 2.28 | 44.2 | 0.906 | 0.207 |
| Restored | 11 | 41 | 9.00 ± 3.38 | 46.3 | 0.893 | 0.230 |
| Disturbed | 11 | 41 | 9.36 ± 2.20 | 54.5 | 0.945 | 0.133 |
Notes.
n, number of sampling points.
Figure 2. Ranking of the abundance of bird species by sampling site.

Only the six most abundant bird species for each site are shown: Least Sandpiper (LS), Yellow Warbler (YW), White Ibis (WI), Green Heron (GH), Great Egret (GE), Tropical Kingbird (TK), Great-tailed Grackle (GG), Magnificent Frigatebird (MF), American White pelican (AWP), Black-bellied Whistling Duck (BWD), Green Kingfisher (GK), and Roseate Spoonbill (RS).
The RDA model was statistically significant (Pseudo- F8,24 = 2.57, p = 0.030), but only a small part of the variation in species abundance within sampling sites was explained by environmental variables since the proportion of unconstrained variance was larger than that of the constrained (0.53 and 0.46, respectively, after 500 permutations for all eigenvalues). When only the first eigenvalue was analyzed, no significant influence of the environmental variables on birds abundance was observed (Fig. S4); however, a trend in some variables such as water temperature, salinity, and tree density to influence the number of birds across sampling sites was detected (Pseudo- F1,24 = 17.97, p = 0.055). To explore the effects of environmental variables on the abundance of birds further, five glms were fitted: one with only environmental variables and not accounting for habitat condition and others that included habitat condition and multiple interactions between environmental variables; however, none of these reached convergence. Thus, the analysis was restricted to additive models between habitat condition and environmental variables. Because pH and water depth showed high correlation with salinity (Spearman’s rank correlation Rho = 0.486, p = 0.001; Rho = − 0.595, p = 0.001, respectively), these two variables were dropped from the models. According to the AIC scores, the best models showed significant effects of the environmental variables and forest structure on the abundance of bird species (Table S2). Habitat condition, i. e., the condition of the mangrove as disturbed, restored or undisturbed, showed more substantial effects on the number of detected birds as well as on the distribution of the bird guilds across sites (Fig. 3). Additionally, vegetation cover, as well as the number of tree species, also significantly positively influenced the birds’ abundance (Table 3A). When the same models were fitted to each single species, neither environmental variables nor the habitat condition showed significant effects on the estimated abundances (Padj = 0.730–0.900 for all species and all variables). The multivariate analysis by functional group showed that only detectability significantly influenced the abundance of bird guilds, while the other variables did not show significant effects on the abundance of any of the analyzed functional groups (Table 3B). Similar to the univariate results by species, no significant result was observed when functional groups were analyzed separately (Padj = 0.172–0.932 for all groups and all variables).
Figure 3. Abundance of bird functional groups by site.
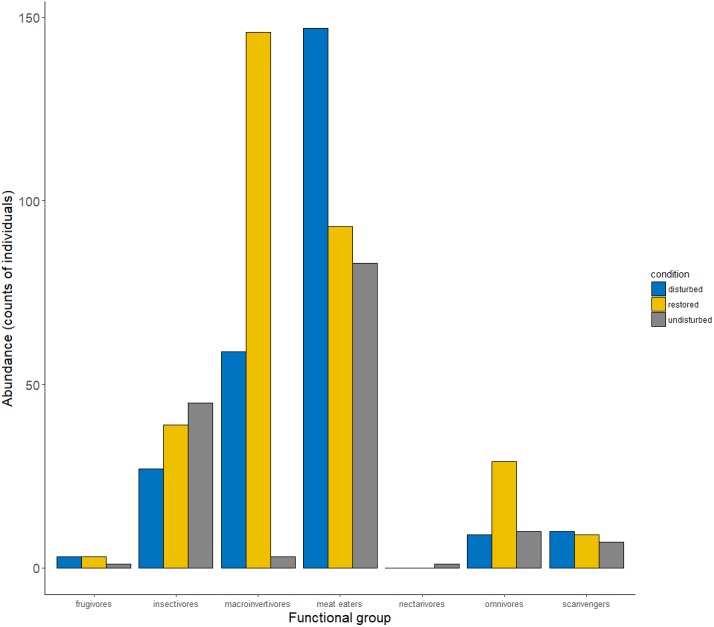
In the restored site, the abundance of guilds such as macroinvertivores and insectivores increased as compared with the disturbed site.
Table 3. Results of the multivariate analysis (best fitted glm).
The greater the deviance, the stronger the effects of the environmental variables on the abundance of bird species (A) and the abundance of functional groups (B).
| A | B | |||
|---|---|---|---|---|
| Site feature | Deviance | P | Deviance | P |
| Habitat condition | 162.50 | 0.012 | 22.598 | 0.110 |
| Water temperature | 113.29 | 0.001 | – | – |
| Redox | 82.83 | 0.029 | – | – |
| Water salinity | 71.86 | 0.040 | – | – |
| No. Tree species | 83.02 | 0.003 | 8.623 | 0.388 |
| Basal area | 82.33 | 0.023 | 6.053 | 0.605 |
| Tree density | 76.50 | 0.035 | 6.719 | 0.490 |
| Basal area: Tree density | 45.00 | 0.157 | – | – |
| Detectability | – | – | 17.780 | 0.019 |
Notes.
Either dash is for variables not included in the models, or for models not reaching convergence when including those variables.
Key species in mangrove forests
According to the similar percentage analysis, wader bird species, such as the Least Sandpiper, the White Ibis, and the Great Egret (Ardea alba) are more associated with the disturbed site. Insectivores such as the Yellow Warbler and the Tropical Kingbird, along with the Green Heron, were more associated with the undisturbed habitat condition. The species associated with the restored site were the Least Sandpiper, the insectivorous Yellow Warbler, and the Green Heron (Table 4). Even though the estimated IndVal value suggested that three bird species might be considered indicators of the different habitat conditions, these results were not significant (Table 5).
Table 4. Similarity analysis of species abundance among sites.
In (A), the comparison of Undisturbed (UN) vs Restored (RS) is shown, (B) Undisturbed vs Disturbed (DS), and (C) Restored vs Disturbed. Species are ordered by their contribution according to the dissimilarity:standard deviation ratio (D/SD).
| (A) | Av. dissimilarity = 76.83 | |||
|---|---|---|---|---|
| Av.Abund UN | Av.Abund RS | Contribution (%) | D/SD | |
| Yellow Warbler | 1.45 | 1.14 | 6.11 | 1.21 |
| Green Heron | 0.89 | 1.08 | 7.35 | 1.12 |
| White Ibis | 0.84 | 0.56 | 5.84 | 1.04 |
| Double-crested Cormorant | 0.31 | 0.70 | 4.72 | 0.96 |
| Tropical Kingbird | 0.61 | 0.09 | 3.72 | 0.89 |
| Great-tailed Grackle | 0.26 | 0.88 | 6.32 | 0.65 |
| Least Sandpiper | 0.09 | 1.14 | 5.55 | 0.58 |
Table 5. Indicator species index (IndVal) values for key species within each sampling site.
A percentage >25% and a p-value 0.05 mean that the selected species are good indicators for a given habitat condition.
| Species | Habitat condition | IndVal (%) | p |
|---|---|---|---|
| Tropical Kingbird | Undisturbed | 39 | 0.068 |
| Blue-winged Teal | Restored | 29 | 0.057 |
| Mangrove Swallow | Disturbed | 27 | 0.951 |
Discussion
The restored tidal channels allowed more frequent water exchange between the Bahamitas estuary and the Laguna de Términos, which increased the frequency of flooding. This, in turn, improved the mangrove soil quality by decreasing the salinity and enabled the natural regeneration of the forest cover. Such changes positively impacted the abundance of insectivorous birds. When comparing the undisturbed and restored sites with the disturbed areas, in the latter, larger duration of flooding favors anaerobic conditions, hindering the oxidation and enabling the rising of soil sulfides concentration (Reddy & Delaune, 2008), which affects the ecophysiological functioning of individual plants and is toxic to aquatic fauna (Lamers et al., 2013). On the other hand, the lack of connectivity with the main lagoon and the absence of mangrove vegetation, as well as the low water exchange and higher rate of evaporation are factors causing higher pH and salinity conditions (Tam et al., 2009; Molnar et al., 2014). These conditions are also likely inhibiting the establishment and growth of seedlings and plants. This effect has been observed in other areas of Laguna de Términos (Agraz-Hernández et al., 2015). By contrast, the undisturbed site showed more frequent water exchange and dense vegetation cover, leading to slower evaporation rate and lower salinity, as posited by Lee et al. (2008). Although there were not yet statistical dissimilarities between the disturbed and the restored sites, only three years after restoration, the opening and desilting of tidal channels brought down the porewater salinity values (Fig. 4) and other components of water quality in the restored site, being overall more similar to the undisturbed site than to the disturbed one, which probably helped to increase the tree canopy cover, litterfall production and tree growth (Kathiresan & Bingham, 2001; Kathiresan, 2002; Polidoro et al., 2010; Kamali & Hashim, 2011). This effect in mangroves has been demonstrated in other sites of the coast of Campeche, where higher rates of litterfall production are associated with lower water salinity (Chan-Keb et al., 2018). Besides the fishing and mud-foraging species characteristic of estuarine areas (Kobza et al., 2004), higher litterfall productivity, in turn, broadens the spectrum of resources by increasing niches for insectivorous birds’ prey species in the restored sites.
Figure 4. The observed changes in porewater salinity though time at the restored site from the beginning to the end of the hydrological restoration (2011–2012), and two years later.

The horizontal lines in the boxes are medians, the open circles are the means, and whiskers are the minimum and maximum values.
Both the disturbed and restored sites showed higher bird abundance and species richness than the undisturbed one. This is similar to the findings reported in structurally complex habitats and island setting, which are found to be more diverse if their habitats show heterogeneity (Acevedo & Aide, 2008; Jones, Marsden & Linsley, 2003), even if they are disturbed, as were two of our study sites. The results from the models highlight the fact that more heterogeneous habitats such as the restored site and, to some extent, the disturbed one result in more species-rich places, allowing for the existence of more structured bird communities than in the more homogenous undisturbed site (Báldi, 2008). This effect is present because high-quality undisturbed mangrove patches surround the disturbed and restored mangrove sites, increasing their heterogeneity. Additionally, while canopy species such as insectivorous birds might use bushes and dead trees from the restored and disturbed sites to feed or rest, bird species that prefer open and semi-open habitats (sandpipers, avocets, cormorants, etc.) are not present in the densely vegetated undisturbed mangrove. They take advantage of the open areas at the disturbed site, which are adequate foraging niches for waders because of the long-lasting floods there. Accordingly, the high abundance of these species could be indicative of sites with impacts affecting the hydroperiod or the vegetation cover. Because of mangrove forests being structurally homogenous when compared to other forest habitats (Mohd-Azlan, Noske & Lawes, 2015), the number of bird species recorded was not as high as expected when surveys include other types of tropical forests. This outcome might also be due to habitat preferences of the different bird species recorded, or could be the effect of short-term monitoring (Canterbury et al., 2000). However, the effects of the sampling duration and size might be negligible, since the heterogeneity of the forest structure traits, such as cover and composition, results in a more complex assemblage of bird communities at the landscape level (Kroll et al., 2014). Hence, the avian community structure might be more influenced by the heterogeneity, diversity and phenology of the mangrove forest than by the size of the sampled sites (Mohd-Azlan, Noske & Lawes, 2012). Moreover, Chacin et al. (2015) demonstrated that hydrological fragmentation does not always negatively affect avian abundance since the loss of hydrological connectivity might result in prey concentration, facilitating forage activities for some species of fishing birds. We agree with this hypothesis but only if tidal patterns allow a cyclic interruption-reconnection of the main and secondary tidal channels to allow flooding and drainage of the mangrove sites. The results of the multivariate analysis supported this idea. Moreover, it demonstrated that bird abundance was more influenced by habitat condition (i.e., hydroperiod and forest structure) than by the measured water parameters. Although there were no clear biological effects of the measured water parameters on the abundance of the birds in our study area, they surely determine resource availability because of their effect on the primary producers, and hence on the presence of the benthonic, fish and crustacean communities (Holguin, Vazquez & Bashan, 2001; Sandilyan & Kathiresan, 2015), on which many bird species feed.
As the characteristics of the habitat influence the distribution of wading species, the sites with less canopy cover provide better foraging areas for waders (Bancroft, Gawlik & Rutchey, 2002; Curado et al., 2013). Additionally, the selection of open and semi-open areas likely reduces the predation risk and increases the foraging efficiency of wading species (Pomeroy, 2006; Chacin et al., 2015). Notwithstanding, while tall and broad canopy vegetation might negatively affect the foraging efficiency of water birds, it might become more beneficial for the insectivores (Tavares & Siciliano, 2013). Thus, the presence of wading and diving species, together with canopy and undergrowth species using the edges of primary habitat and the emerging vegetation within the restored site, may have influenced the proportions of bird abundance and richness in the study area.
Because our sampling sites are located within a landscape matrix of disturbed and undisturbed patches, the geographical distribution of the study locations likely influenced the number of species found in each sampling site. For example, recordings of birds common to undisturbed sites, such as the Mangrove Warbler, were relatively frequent in the restored and disturbed sites because this species uses features of the disturbed areas as rest spots or merely in the movement between undisturbed patches, as other species do (Mohd-Azlan, Noske & Lawes, 2015). This behavior might prevent the identification of particular species from each sampling location just on the basis of observation. Thus, estimates of abundance might be a better indicator. For instance, the abundance of the Mangrove Warbler was higher in both undisturbed and restored areas, although we recorded this species in all studied sites. Additionally, based on the species-by-species abundance, we obtained evidence of non-random use of the restored site as foraging habitats for this and other insectivores (Table 4).
The regrowth of vegetation cover induced by the restoration activities, demonstrated by the increase in the height (0–55 cm) of the mangrove scrub and increased recruitment (0–79 individualsha−1) (see Echeverría-Ávila et al., 2019), improved the availability of resources and, hence, the presence and the abundance of insectivorous birds such as the Mangrove Warbler and the Tropical Kingbird. A very close relative of the former, the Yellow Warbler (Setophaga petechia), has been considered a key species because of its sensitivity to changes in environmental conditions and specific habitat needs (Lowther et al., 1999), and because its populations may change according to the habitat management practices and food availability (Salgado-Ortiz, Marra & Robertson, 2009). Since the higher abundance of the Mangrove Warbler was apparently related to the undisturbed areas, it demonstrates its importance as an indicator of habitat with negligible impacts, or of habitats showing signs of recovery. Also, the open and semi-open areas in the restored site contributed to the higher availability of resources for different bird guilds (Ortega-Alvarez & Lindig-Cisneros, 2012; Buelow & Sheaves, 2015). These areas were more attractive to birds which usually flock in large groups and forage on the ground, mud or inundated areas, leading to higher abundances than in the densely-vegetated undisturbed site. Even though the IndVal was non-significant, both approaches used to identify key species suggested that insectivores better represented the undisturbed site. According to the Simper analysis, the species that best represents the restored mangrove is a meat-eater (the Green Heron), whereas the IndVal suggested that an omnivore (the Blue-winged Teal [Anas discors]) could be an indicator for this site. This lack of coincidence might relate to differing information used by each approach. While Simper uses only the information regarding abundance, IndVal uses the relative abundance and also incorporates the relative frequency of occurrence.
Additionally, the presence of species such as ibises and wood storks is considered evidence of success after the implementation of restoration activities (Ortega-Alvarez & Lindig-Cisneros, 2012; Zhao et al., 2016). This is because they depend mostly on communities of vertebrates and invertebrates that are sensitive to changes in water and soil quality, induced among other factors, by the alteration of the hydroperiod (Ogden et al., 2014). Accordingly, the presence and abundance of the insectivorous and wading bird species similar to those found in the study area are important elements by which to evaluate and monitor the effectiveness of habitat restoration projects in mangrove ecosystems.
Implications for Conservation
There is no doubt that natural phenomena, such as hurricanes, contribute to habitat heterogeneity. However, severe climatic events may alter the microtopography and hydrological connectivity and make difficult post-hurricane natural regeneration. Such is especially the case in habitats that depend on recurrent flooding and drainage (hydroperiods) such as mangroves. By altering the hydroperiod, dead mangrove areas may increase in size through time. The loss of the vegetation cover and the alteration of the environmental conditions may, in turn, lead to biotic homogenization (Martínez-Ruiz & Renton, 2018). After degradation due to loss of hydrological connectivity occurs in mangroves, restoration of water flow through deblocking of main and secondary tidal channels increases the habitat heterogeneity, allowing the resettlement of ecosystem services and strengthening the ecological relationships and structure of the biotic communities living there. It may also improve the hydro-edaphic factors, such as nutrients, water level, and porewater salinity (McKee et al., 2002), from which the micro and macrobenthos, as well as fisheries and birds, are dependent.
Long flooding periods limit the abundance of wading birds, especially the small species foraging on the macrobenthos. Thus, identifying degraded areas and monitoring the hydroperiod, before and after hydrological restoration, will allow for better conservation strategies for the mangroves and their avian communities. As the hydrological connectivity improves mangrove heterogeneity by regenerating vegetation, avian communities can become more diverse, acquiring a higher number of species from different functional groups. Thus, densely vegetated mangroves, together with the restoration of patches unable to regenerate themselves, are essential to maximizing the abundance of specialized bird guilds.
To realize the positive effects of restoration activities, it is necessary to create a link between the restoration programs and the local communities through environmental and economic development education. For instance, birdwatching and sport fishing (catch and release) can be alternatives to socio-economic growth for the benefit of human coastal communities through the development of a green touristic industry in restored places (Skelton & Allaway, 1996).
We expect that the restoration activities implemented will increase and maintain the habitat’s heterogeneity in the medium and the long term and will strengthen the resilience of the mangrove forests.
Conclusions
The restoration activities in the estuaries of Laguna de Términos first helped the natural regeneration of the mangrove forest through the water movement caused by the opening and desilting of tidal channels, making tidal periodicity comparable to that of natural conditions while also reducing water salinity. In addition, water exchange likely favored fish and crustacean production and the appearance of mudflat and shallow water zones, which are attractive foraging areas for different bird guilds. Consequently, habitat heterogeneity and the availability of resources increased, and the avian community became more diverse, especially regarding the abundance of insectivorous birds in the restored areas.
Supplemental Information
Same letters indicate no differences between the study areas. On the right side the hydroperiod components (D–F). The star represents significant differences among different mangrove conditions. The boxes show the inter-quartile range, the thick black lines within the boxes are the medians, and the whiskers are the minimum and maximum values.
{kind=link}
(A) show the number of tree species. In (B) the density of mangrove tress is shown. (C) shows the measured height of the mangrove trees at each sampling site. (D) shows the cover expressed as basal area at each sampling site.
{kind=link}
The different studied sites are disturbed (triangles), restored (crosses), and undisturbed (circles). In both graphics, bars indicate ±1.5 standard deviations.
{kind=link}
{kind=link}
Area: restored (R), undisturbed (U), disturbed (D) NOM-059: Is the list of threatened species recognized by the Mexican Environmetal Authorities. The categories are: least concern (LC), subject to special protection (SP), threatened (TH). Pop.Trend are the population trends as reported by the International Union for the Conservation of Nature (IUCN).
H, habitat condition; R, measured Redox; T, water temperature; S, water salinity; Sps, number of tree species; B, Basal area; Dn, trees density; Dt, detectability.
Acknowledgments
We thank Herminia Rejón Salazar and the ‘Community of Mangrove Restorers’ from Isla Aguada for their support with the field work, Emma Guevara Carrió, Mario Alejandro Gómez Ponce, Hernán Álvarez Guillén and Andrés Reda Deara for their assistance with logistics and field data collection. We thank José Nava from CONANP-Laguna de Términos for the facilities to carry out our surveys.
Funding Statement
This work was supported by Comisión Nacional Forestal, Gulf of Mexico Large Marine Ecosystem Project, Instituto de Ciencias del Mar y Limnología UNAM, Universidad Autónoma del Carmen (grant number CA-CONAFOR-UNACAR-GOM LME 2014-2016). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests. Arturo Zaldívar-Jiménez is a mangrove restoration specialist and manager of ATEC Asesoría Técnica y Estudios Costeros SCP.
Author Contributions
Julio Cesar Canales-Delgadillo analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Rosela Perez-Ceballos conceived and designed the experiments, analyzed the data, contributed reagents/materials/analysis tools, prepared figures and/or tables.
Mario Arturo Zaldivar-Jimenez conceived and designed the experiments, analyzed the data, contributed reagents/materials/analysis tools.
Martin Merino-Ibarra authored or reviewed drafts of the paper, approved the final draft.
Gabriela Cardoza performed the experiments.
Jose-Gilberto Cardoso-Mohedano authored or reviewed drafts of the paper.
Data Availability
The following information was supplied regarding data availability:
The raw data is available in the Supplemental Files.
References
- Acevedo & Aide (2008).Acevedo M, Aide T. Bird community dynamics and habitat associations in karsts. mangrove and Pterocarpous forest fragments in an urban zone in Puerto Rico. Caribbean Journal of Science. 2008;44:402–416. [Google Scholar]
- Adame et al. (2015).Adame M, Fry B, Gamboa J, Herrera-Silveira J. Nutrient subsidies delivered by seabirds to mangrove islands. Marine Ecology Progress Series. 2015;525:15–4. doi: 10.3354/meps11197. [DOI] [Google Scholar]
- Agraz-Hernández et al. (2015).Agraz-Hernández CM, Chan-Keb CA, Iriarte-Vivar S, Posada Venegas G, Vega Serratos B, Osti Sáenz J. Phenological variation of Rhizophora mangle and ground water chemistry associated to changes of the precipitation. Hidrobiológica. 2015;25:49–61. [Google Scholar]
- Alongi (2008).Alongi DM. Mangrove forests: resilience, protection from tsunamis, and responses to global climate change. Estuarine, Coastal and Shelf Science. 2008;76:1–13. doi: 10.1016/j.ecss.2007.08.024. [DOI] [Google Scholar]
- Amir (2018).Amir AA. Mitigate risk for Malaysia’s mangroves. Science. 2018;359(6382):1342.2–1343. doi: 10.1126/science.aas9139. [DOI] [PubMed] [Google Scholar]
- Anderson, Gorley & Clarke (2008).Anderson MJ, Gorley RN, Clarke KR. PERMANOVA+ for PRIMER: guide to software and statistical methods. PRIMER-E, Ltd; Plymouth: 2008. [Google Scholar]
- Andrade et al. (2012).Andrade LL, Leite D, Ferreira E, Ferreira L, Paula GR, Maguire M, Hubert C, Peixoto R, Domingues R, Rosado A. Microbial diversity and anaerobic hydrocarbon degradation potential in an oil-contaminated mangrove sediment. BMC Microbiology. 2012;12:186. doi: 10.1186/1471-2180-12-186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azhar et al. (2013).Azhar B, Lindenmayer DB, Wood J, Fischer J, Manning A, McElhinny C, Zakaria M. The influence of agricultural system, stand structural complexity and landscape context on foraging birds in oil palm landscapes. Ibis. 2013;155:297–312. doi: 10.1111/ibi.12025. [DOI] [Google Scholar]
- Báldi (2008).Báldi A. Habitat heterogeneity overrides the species–area relationship. Journal of Biogeography. 2008;35:675–681. doi: 10.1111/j.1365-2699.2007.01825.x. [DOI] [Google Scholar]
- Bancroft, Gawlik & Rutchey (2002).Bancroft GT, Gawlik DE, Rutchey K. Distribution of wading birds relative to vegetation and water depths in the Northern Everglades of Florida, USA. Waterbirds. 2002;25:265–391. doi: 10.1675/1524-4695(2002)025[0265:DOWBRT]2.0.CO;2. [DOI] [Google Scholar]
- Battisti, Dodaro & Franco (2014).Battisti C, Dodaro G, Franco D. The data reliability in ecological research: a proposal for a quick self-assessment tool. Natural History Sciences. 2014;1:75–79. doi: 10.4081/nhs.2014.61. [DOI] [Google Scholar]
- Behrouzi-Rad (2014).Behrouzi-Rad B. Population dynamic and species diversity of wintering waterbirds in Mangroves Wetland (Persian Gulf) in 1983 and 2013. International Journal of Marine Science. 2014;4:1–7. [Google Scholar]
- Bibby et al. (1992).Bibby CJ, Burgess ND, Hill DA, Bibby CJ, Burgess ND, Hill DA. Bird census techniques. Academic Press; London: 1992. [Google Scholar]
- Boyle, Dorn & Cook (2012).Boyle RA, Dorn NJ, Cook MI. Nestling diet of three sympatrically nesting wading bird species in the Florida Everglades. Waterbirds. 2012;35:154–159. doi: 10.1675/063.035.0116. [DOI] [Google Scholar]
- Bryce, Hughes & Kaufmann (2002).Bryce SA, Hughes RM, Kaufmann PR. Development of a bird integrity index: using bird assemblages as indicators of riparian condition. Environmental Management. 2002;30:294–310. doi: 10.1007/s00267-002-2702-y. [DOI] [PubMed] [Google Scholar]
- Buelow & Sheaves (2015).Buelow C, Sheaves M. A birds-eye view of biological connectivity in mangrove systems. Estuarine, Coastal and Shelf Science. 2015;152:33–43. doi: 10.1016/j.ecss.2014.10.014. [DOI] [Google Scholar]
- Burnham & Anderson (2002).Burnham K, Anderson D. Model selection and multimodel inference: a practical information-theoretic approach. Springer Verlag; New York: 2002. [Google Scholar]
- Burnham, Anderson & Huyvaert (2011).Burnham KP, Anderson DR, Huyvaert KP. AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons. Behavioral Ecology and Sociobiology. 2011;65:23–35. doi: 10.1007/s00265-010-1029-6. [DOI] [Google Scholar]
- Canterbury et al. (2000).Canterbury GE, Martin TE, Petit DR, Petit LJ, Bradford DF. Bird communities and habitat as ecological indicators of forest condition in regional monitoring. Conservation Biology. 2000;14:544–558. doi: 10.1046/j.1523-1739.2000.98235.x. [DOI] [Google Scholar]
- Catterall et al. (2012).Catterall CP, Freeman AND, Kanowski J, Freebody K. Can active restoration of tropical rainforest rescue biodiversity? A case with bird community indicators. Biological Conservation. 2012;146:53–61. doi: 10.1016/j.biocon.2011.10.033. [DOI] [Google Scholar]
- Chacin et al. (2015).Chacin DH, Giery ST, Yeager LA, Layman CA, Brian-Langerhans R. Does hydrological fragmentation affect coastal bird communities? A study from Abaco Island, The Bahamas. Wetlands Ecology and Management. 2015;23:551–557. doi: 10.1007/s11273-014-9389-8. [DOI] [Google Scholar]
- Chan-Keb et al. (2018).Chan-Keb CA, Agraz-Hernández CM, Muñiz Salazar R, Posada-Vanegas G, Osti-Sáenz J, Reyes Castellano JE, Conde-Medina KP, Vega-Serratos BE. Ecophysiological response of rhizophora mangle to the variation in hydrochemistry during five years along the coast of Campeche, México. Diversity. 2018;10:9. doi: 10.3390/d10010009. [DOI] [Google Scholar]
- Chape, Spalding & Jenkins (2008).Chape S, Spalding M, Jenkins M. The world’s protected areas: status, values and prospects in the 21st century. University of California; Berkeley: 2008. [Google Scholar]
- Clarke (1993).Clarke KR. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology. 1993;18:117–143. doi: 10.1111/j.1442-9993.1993.tb00438.x. [DOI] [Google Scholar]
- Costanza et al. (2014).Costanza R, De Groot R, Sutton P, Van der Ploeg S, Anderson SJ, Kubiszewski I, Farber S, Turner RK. Changes in the global value of ecosystem services. Global Environmental Change. 2014;26:152–158. doi: 10.1016/j.gloenvcha.2014.04.002. [DOI] [Google Scholar]
- Crase et al. (2013).Crase B, Liedloff A, Vesk PA, Burgman MA, Wintle BA. Hydroperiod is the main driver of the spatial pattern of dominance in mangrove communities. Global Ecology and Biogeography. 2013;22:806–817. doi: 10.1111/geb.12063. [DOI] [Google Scholar]
- Cui et al. (2009).Cui B, Yang Q, Yang Z, Zhang K. Evaluating the ecological performance of wetland restoration in the Yellow River Delta, China. Ecological Engineering. 2009;35:1090–1103. doi: 10.1016/j.ecoleng.2009.03.022. [DOI] [Google Scholar]
- Curado et al. (2013).Curado G, Figueroa E, Sanchez MI, Castillo JM. Avian communities in Spartina maritima restored and non-restored salt marshes. Bird Study. 2013;60:185–194. doi: 10.1080/00063657.2013.790875. [DOI] [Google Scholar]
- Cvetkovic & Chow-Fraser (2011).Cvetkovic M, Chow-Fraser P. Use of ecological indicators to assess the quality of Great Lakes coastal wetlands. Ecological Indicators. 2011;11:1609–1622. doi: 10.1016/j.ecolind.2011.04.005. [DOI] [Google Scholar]
- David & Kjerfve (1998).David LT, Kjerfve B. Tides and currents in a two-inlet coastal lagoon: laguna de Términos, México. Continental Shelf Research. 1998;18:1057–1079. doi: 10.1016/S0278-4343(98)00033-8. [DOI] [Google Scholar]
- Donato et al. (2011).Donato DC, Kauffman JB, Murdiyarso D, Kurnianto S, Stidham M, Kanninen M. Mangroves among the most carbon-rich forests in the tropics. Nature Geoscience. 2011;4:293–297. doi: 10.1038/ngeo1123. [DOI] [Google Scholar]
- Echeverría-Ávila et al. (2019).Echeverría-Ávila S, Pérez-Ceballos R, Zaldívar-Jiménez A, Canales-Delgadillo J, Brito-Pérez R, Merino-Ibarra M, Vovides A. Regeneración natural de sitios de manglar degradado en respuesta a la restauración hidrológica. Madera y Bosques. 2019;25:e2511754. doi: 10.21829/myb.2019.2511754. [DOI] [Google Scholar]
- Fagan & Komar (2016).Fagan J, Komar O. Field guide to birds of Northern Central America. HMH Publishing Company; New York: 2016. [Google Scholar]
- Frederick et al. (2009).Frederick P, Gawlik DE, Ogden JC, Cook MI, Lusk M. The white ibis and wood stork as indicators for restoration of the everglades ecosystem. Ecological Indicators. 2009;9:S83–S95. doi: 10.1016/j.ecolind.2008.10.012. [DOI] [Google Scholar]
- Gardner et al. (2012).Gardner CJ, De Ridder C, De Ridder B, Jasper LD. Birds of Ambondrolava mangrove complex, Southwest Madagascar. Check List. 2012;8:1–7. doi: 10.15560/8.1.001. [DOI] [Google Scholar]
- Giri et al. (2011).Giri C, Ochieng E, Tieszen LL, Zhu Z, Singh A, Loveland T, Masek J, Duke N. Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecology and Biogeography. 2011;20:154–159. doi: 10.1111/j.1466-8238.2010.00584.x. [DOI] [Google Scholar]
- Gotelli & Colwell (2011).Gotelli NJ, Colwell RK. Estimating species richness. In: Levin SA, editor. Biological diversity: frontiers in measuring biodiversity. Oxford University Press; New York: 2011. pp. 39–54. [Google Scholar]
- Gregory, Gibbons & Donald (2004).Gregory RD, Gibbons DW, Donald PF. Bird census and survey techniques. In: Sutherland WJ, Newton I, Green R, editors. Bird ecology and conservation. Oxford: Oxford University Press; 2004. pp. 17–56. [Google Scholar]
- Gyurácz, Bánhidi & Csuka (2011).Gyurácz J, Bánhidi P, Csuka A. Successful restoration of water level and surface area restored migrant bird populations in a Hungarian wetland. Biologia. 2011;66:1177–1182. doi: 10.2478/s11756-011-0132-0. [DOI] [Google Scholar]
- Hauser et al. (2017).Hauser LT, Nguyen Vu G, Nguyen BA, Dade E, Nguyen HM, Nguyen TTQ, Le TQ, Vu LH, Tong ATH, Pham HV. Uncovering the spatio-temporal dynamics of land cover change and fragmentation of mangroves in the Ca Mau peninsula, Vietnam using multi-temporal SPOT satellite imagery (2004–2013) Applied Geography. 2017;86:197–207. doi: 10.1016/j.apgeog.2017.06.019. [DOI] [Google Scholar]
- Holguin, Vazquez & Bashan (2001).Holguin G, Vazquez P, Bashan Y. The role of sediment microorganisms in the productivity, conservation, and rehabilitation of mangrove ecosystems: an overview. Biology and Fertility of Soils. 2001;33:265–278. doi: 10.1007/s003740000319. [DOI] [Google Scholar]
- Ingram (2008).Ingram J. Berger–Parker index. In: Jorgensen SE, Fath BD, editors. Encyclopedia of ecology. Elsevier; Oxford: 2008. pp. 332–334. [Google Scholar]
- IUCN (2016).IUCN The IUCN red list of threatened species. Version 2016-3. 2016. https://www.iucn.org/resources/conservation-tools/iucn-red-list-threatened-species https://www.iucn.org/resources/conservation-tools/iucn-red-list-threatened-species
- Jones, Marsden & Linsley (2003).Jones MJ, Marsden SJ, Linsley MD. Effects of habitat change and geographical variation on the bird communities of two Indonesian islands. Biodiversity and Conservation. 2003;12:1013–1032. doi: 10.1023/A:1022810104690. [DOI] [Google Scholar]
- Kamali & Hashim (2011).Kamali B, Hashim R. Mangrove restoration without planting. Ecological Engineering. 2011;37:387–391. doi: 10.1016/j.ecoleng.2010.11.025. [DOI] [Google Scholar]
- Kathiresan (2002).Kathiresan K. Why are mangrove degrading? Current Sicience. 2002;83(10):1246–1249. [Google Scholar]
- Kathiresan & Bingham (2001).Kathiresan K, Bingham B. Biology of mangroves and mangrove Ecosystems. Advances in Marine Biology. 2001;40:81–251. doi: 10.1016/S0065-2881(01)40003-4. [DOI] [Google Scholar]
- Kindt & Coe (2005).Kindt R, Coe R. Tree diversity analysis: a manual and software for common statistical methods for ecological and biodiversity studies. World Agroforestry Center; Nairobi: 2005. [Google Scholar]
- Kobza et al. (2004).Kobza M, Trexler JC, Loftus WF, Perry SA. Community structure of fishes inhabiting aquatic refuges in a threatened Karst wetland and its implications for ecosystem management. Biological Conservation. 2004;116:153–165. doi: 10.1016/S0006-3207(03)00186-1. [DOI] [Google Scholar]
- Krauss et al. (2006).Krauss KW, Doyle TW, Twilley RR, Rivera-Monroy VH, Sullivan JK. Evaluating the relative contributions of hydroperiod and soil fertility on growth of south Florida mangroves. Hydrobiologia. 2006;569:311–324. doi: 10.1007/s10750-006-0139-7. [DOI] [Google Scholar]
- Kroll et al. (2014).Kroll AJ, Ren Y, Jones JE, Giovanini J, Perry RW, Thill RE, White D, Wigley TB. Avian community composition associated with interactions between local and landscape habitat attributes. Forest Ecology and Management. 2014;326:46–57. doi: 10.1016/j.foreco.2014.04.011. [DOI] [Google Scholar]
- Lamers et al. (2013).Lamers L, Govers L, Janssen I, Geurts J, Van der Welle M, Van Katwijk M, Van der Heide T, Roelofs J, Smolders A. Sulfide as a soil phytotoxin—a review. Frontiers in Plant Science. 2013;4:268. doi: 10.3389/fpls.2013.00268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lantz, Gawlik & Cook (2011).Lantz SM, Gawlik DE, Cook MI. The effects of water depth and emergent vegetation on foraging success and habitat selection of wading birds in the everglades. Waterbirds. 2011;34:439–447. doi: 10.1675/063.034.0406. [DOI] [Google Scholar]
- Lavoie et al. (2010).Lavoie RA, Champoux L, Rail JF, Lean DRS. Organochlorines, brominated flame retardants and mercury levels in six seabird species from the Gulf of St. Lawrence (Canada): relationships with feeding ecology, migration and molt. Environmental Pollution. 2010;158:2189–2199. doi: 10.1016/j.envpol.2010.02.016. [DOI] [PubMed] [Google Scholar]
- Lee et al. (2008).Lee RY, Porubsky WP, Feller IC, McKee KL, Joye SB. Porewater biogeochemistry and soil metabolism in dwarf red mangrove habitats (Twin Cays, Belize) Biogeochemistry. 2008;87:181–198. doi: 10.1007/s10533-008-9176-9. [DOI] [Google Scholar]
- Lee et al. (2014).Lee SY, Primavera JH, Dahdouh-Guebas F, McKee K, Bosire JO, Cannicci S, Diele K, Fromard F, Koedam N, Marchand C, Mendelssohn I, Mukherjee N, Record S. Ecological role and services of tropical mangrove ecosystems: a reassessment. Global Ecology and Biogeography. 2014;23:726–743. doi: 10.1111/geb.12155. [DOI] [Google Scholar]
- Li et al. (2011).Li D, Chen S, Guan L, Lloyd H, Liu Y, Lv J, Zhang Z. Patterns of waterbird community composition across a natural and restored wetland landscape mosaic, Yellow River Delta, China. Estuarine, Coastal and Shelf Science. 2011;91:325–332. doi: 10.1016/j.ecss.2010.10.028. [DOI] [Google Scholar]
- Lowther et al. (1999).Lowther PE, Celada N, Klein C, Rimner C, Spector DA. Yellow Warbler (Dendroica petechia) In: Poole A, Gill F, editors. The birds of North America, no. 454. The Birds of North America Online; Ithaca: 1999. [Google Scholar]
- MacKinnon (2013).MacKinnon B. Sal a Pajarear Yucatán (Guía de Aves) La Vaca Independiente S.A. de C.V. Ciencias; Merida: 2013. [Google Scholar]
- Martínez-Ruiz & Renton (2018).Martínez-Ruiz M, Renton K. Habitat heterogeneity facilitates resilience of diurnal raptor communities to hurricane disturbance. Forest Ecology and Management. 2018;426:134–144. doi: 10.1016/j.foreco.2017.08.004. [DOI] [Google Scholar]
- McElhinny et al. (2005).McElhinny C, Gibbons P, Brack C, Bauhus J. Forest and woodland stand structural complexity: its definition and measurement. Forest Ecology and Management. 2005;218:1–24. doi: 10.1016/j.foreco.2005.08.034. [DOI] [Google Scholar]
- McFadden, Kauffman & Bhomia (2016).McFadden TN, Kauffman JB, Bhomia RK. Effects of nesting waterbirds on nutrient levels in mangroves, Gulf of Fonseca, Honduras. Wetlands Ecology and Management. 2016;24:217–229. doi: 10.1007/s11273-016-9480-4. [DOI] [Google Scholar]
- McKee et al. (2002).McKee KL, Feller IC, Popp M, Wanek W. Mangrove isotopic (δ 15N and δ 13C) fractionation across a nitrogen vs. phosphorus limitation gradient. Ecology. 2002;83:1065–1075. [Google Scholar]
- Mexico’s Secretariat of Environment and Natural Resources (SEMARNAT) (2010).Mexico’s Secretariat of Environment and Natural Resources (SEMARNAT) Norma oficial Mexicana NOM-059-SEMARNAT-2010. 2010. http://www.dof.gob.mx/normasOficiales/4254/semarnat/semarnat.htm. [26 January 2019]. http://www.dof.gob.mx/normasOficiales/4254/semarnat/semarnat.htm
- Mohd-Azlan, Noske & Lawes (2012).Mohd-Azlan J, Noske RA, Lawes MJ. Avian species-assemblage structure and indicator bird species of mangroves in the Australian monsoon tropics. Emu—Austral Ornithology. 2012;112:287–297. doi: 10.1071/MU12018. [DOI] [Google Scholar]
- Mohd-Azlan, Noske & Lawes (2015).Mohd-Azlan J, Noske R, Lawes M. The role of habitat heterogeneity in structuring mangrove bird assemblages. Diversity. 2015;7(2):118–136. doi: 10.3390/d7020118. [DOI] [Google Scholar]
- Molnar et al. (2014).Molnar N, Marchand C, Deborde J, Della Patrona L, Meziane T. Seasonal pattern of the biogeochemical properties of mangrove sediments receiving shrimp farm effluents (New Caledonia) Journal of Aquaculture Research & Development. 2014;05:1–13. [Google Scholar]
- Nagelkerken et al. (2008).Nagelkerken I, Blaber SJM, Bouillon S, Green P, Haywood M, Kirton LG, Meynecke JO, Pawlik J, Penrose HM, Sasekumar A, Somerfield PJ. The habitat function of mangroves for terrestrial and marine fauna: a review. Aquatic Botany. 2008;89:155–185. doi: 10.1016/j.aquabot.2007.12.007. [DOI] [Google Scholar]
- Nilsson & Aradottir (2013).Nilsson C, Aradottir AL. Ecological and social aspects of ecological restoration: new challenges and opportunities for northern regions. Ecology and Society. 2013;18:35. [Google Scholar]
- Ogden et al. (2014).Ogden JC, Baldwin JD, Bass OL, Browder JA, Cook MI, Frederick PC, Frezza PE, Galvez RA, Hodgson AB, Meyer KD, Oberhofer LD, Paul AF, Fletcher PJ, Davis SM, Lorenz JJ. Waterbirds as indicators of ecosystem health in the coastal marine habitats of Southern Florida: 2. Conceptual ecological models. Ecological Indicators. 2014;44:128–147. doi: 10.1016/j.ecolind.2014.03.008. [DOI] [Google Scholar]
- Ortega-Alvarez & Lindig-Cisneros (2012).Ortega-Alvarez R, Lindig-Cisneros R. Feathering the scene: the effects of ecological restoration on birds and the role birds play in evaluating restoration outcomes. Ecological Restoration. 2012;30:116–127. doi: 10.3368/er.30.2.116. [DOI] [Google Scholar]
- Palmer, Hondula & Koch (2014).Palmer MA, Hondula KL, Koch BJ. Ecological restoration of streams and rivers: shifting strategies and shifting goals. Annual Review of Ecology, Evolution, and Systematics. 2014;45:247–269. doi: 10.1146/annurev-ecolsys-120213-091935. [DOI] [Google Scholar]
- Paradis, Claude & Strimmer (2004).Paradis E, Claude J, Strimmer K. APE: analyses of phylogenetics and evolution in R language. Bioinformatics. 2004;20:289–290. doi: 10.1093/bioinformatics/btg412. [DOI] [PubMed] [Google Scholar]
- Pendleton (1995).Pendleton GW. John RC, Sauer JR, Sam D, editors. Effects of sampling strategy, detection probability, and independence of counts on the use of point counts. Albany: U.S. Department of Agriculture, Forest Service, Pacific SouthwestResearch StationMonitoring bird populations by point counts. General technical report PSW-GTR-149. 1995:131–134.
- Pérez-Ceballos et al. (2013).Pérez-Ceballos R, Zaldívar-Jiménez A, Ladrón de Guevara P, Amador del Angel L, Endañú Huerta E, Guevara-Carrió E, Herrera-Silveira J. Los manglares de la región Atasta-Sabancuy, Campeche, México. Programa piloto de conservación y restauración de manglares dentro del Proyecto Integral para el Gran Ecosistema Marino del Golfo de México. Ciudad del Carmen, Campeche, MéxicoTechnical report. 2013
- Pohlert (2014).Pohlert T. The pairwise multiple comparison of mean ranks package (PMCMR) https://CRAN.R-project.org/package=PMCMR R package. 2014
- Polidoro et al. (2010).Polidoro BA, Carpenter KE, Collins L, Duke NC, Ellison AM, Ellison JC, Farnsworth EJ, Fernando ES, Kathiresan K, Koedam NE, Livingstone SR, Miyagi T, Moore GE, Nam VN, Ong JE, Primavera JH, Salmo SG, Sanciangco JC, Sukardjo S, Wang Y, Yong JWH. The loss of species: mangrove extinction risk and geographic areas of global concern. PLOS ONE. 2010;5(4):e10095. doi: 10.1371/journal.pone.0010095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pomeroy (2006).Pomeroy AC. Tradeoffs between food abundance and predation danger in spatial usage of a stopover site by western sandpipers, Calidris mauri. Oikos. 2006;112:629–637. doi: 10.1111/j.0030-1299.2006.14403.x. [DOI] [Google Scholar]
- Reddy & Delaune (2008).Reddy KR, Delaune RD. Oxygen. In: Reddy KR, Delaune RD, editors. Biogeochemistry of Wetlands. CRC Press; Boca Raton: 2008. pp. 185–213. [Google Scholar]
- Roberts (2016).Roberts DW. labdvs: ordination and multivariate analysis for ecology. R package version 1.8-0https://rdrr.io/cran/labdsv/ 2016
- Royston (1995).Royston P. Remark AS R94: a remark on algorithm AS 181: the W test for normality. Applied Statistics. 1995;44:547–551. doi: 10.2307/2986146. [DOI] [Google Scholar]
- Salem & Mercer (2012).Salem ME, Mercer DE. The economic value of mangroves: a meta-analysis. Sustainability. 2012;4:359–383. doi: 10.3390/su4030359. [DOI] [Google Scholar]
- Salgado-Ortiz, Marra & Robertson (2009).Salgado-Ortiz J, Marra PP, Robertson RJ. Breeding seasonality of the mangrove warbler (Dendroica petechia bryanti) from southern Mexico. Ornitologia Neotropical. 2009;20:255–263. [Google Scholar]
- Salmo, Tibbetts & Duke (2016).Salmo S, Tibbetts I, Duke N. Colonization and shift of mollusc assemblages as a restoration indicator in planted mangroves in the Philippines. Biodiversity and Conservation. 2016;26:865–881. [Google Scholar]
- Sandilyan & Kathiresan (2015).Sandilyan S, Kathiresan K. Density of waterbirds in relation to habitats of Pichavaram mangroves, Southern India. Journal of Coastal Conservation. 2015;19:131–139. doi: 10.1007/s11852-015-0376-x. [DOI] [Google Scholar]
- Schaeffer-Novelli, Vale & Cintrón (2015).Schaeffer-Novelli Y, Vale CC, Cintrón G. Guia para estudo de a’reas de manguezal. Estrutura, função e flora. In: Turra A, Denadai MR, editors. Protocolos para o monitoramento de habitats bentonicos costeiros. Instituto Oceanográfico da Universidade de Sao Paulo; Sao Paulo: 2015. pp. 62–80. [Google Scholar]
- Schaffelke, Mellors & Duke (2005).Schaffelke B, Mellors J, Duke NC. Water quality in the Great Barrier Reef region: responses of mangrove, seagrass and macroalgal communities. Marine Pollution Bulletin. 2005;51:279–296. doi: 10.1016/j.marpolbul.2004.10.025. [DOI] [PubMed] [Google Scholar]
- Schmidt, McIntyre & MacCluskie (2013).Schmidt JH, McIntyre CL, MacCluskie MC. Accounting for incomplete detection: what are we estimating and how might it affect long-term passerine monitoring programs? Biological Conservation. 2013;160:130–139. doi: 10.1016/j.biocon.2013.01.007. [DOI] [Google Scholar]
- Serafy et al. (2015).Serafy JE, Shideler GS, Araújo RJ, Nagelkerken I. Mangroves enhance reef fish abundance at the caribbean regional scale. PLOS ONE. 2015;10:e0142022. doi: 10.1371/journal.pone.0142022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skelton & Allaway (1996).Skelton NJ, Allaway WG. Oxygen and pressure changes measured in situ during flooding in roots of the Grey Mangrove Avicennia marina (Forssk.) Vierh. Aquatic Botany. 1996;54:165–175. doi: 10.1016/0304-3770(96)01043-1. [DOI] [Google Scholar]
- Smith & Van Belle (1984).Smith EP, Van Belle G. Nonparametric estimation of species richness. Biometrics. 1984;40:119–129. doi: 10.2307/2530750. [DOI] [Google Scholar]
- Suding et al. (2015).Suding K, Higgs E, Palmer M, Callicott JB, Anderson CB, Baker M, Gutrich JJ, Hondula KL, LaFevor MC, Larson BMH, Randall A, Ruhl JB, Schwartz KZS. Committing to ecological restoration. Science. 2015;348:638–640. doi: 10.1126/science.aaa4216. [DOI] [PubMed] [Google Scholar]
- Tam et al. (2009).Tam NFY, Wong AHY, Wong MH, Wong YS. Mass balance of nitrogen in constructed mangrove wetlands receiving ammonium-rich wastewater: effects of tidal regime and carbon supply. Ecological Engineering. 2009;35:453–462. doi: 10.1016/j.ecoleng.2008.05.011. [DOI] [Google Scholar]
- Tavares & Siciliano (2013).Tavares DC, Siciliano S. Variação temporal na abundância de espe’cies de aves aqua’ticas em uma lagoa costeira do Norte Fluminense do estado do Rio de Janeiro, sudeste do Brasil. Biotemas. 2013;27:121–132. [Google Scholar]
- Thornton & Johnstone (2015).Thornton SR, Johnstone RW. Mangrove rehabilitation in high erosion areas: assessment using bioindicators. Estuarine, Coastal and Shelf Science. 2015;165:176–184. doi: 10.1016/j.ecss.2015.05.013. [DOI] [Google Scholar]
- Turner & Lewis (1996).Turner RE, Lewis RR. Hydrologic restoration of coastal wetlands. Wetland Ecology and Management. 1996;4:65–72. doi: 10.1007/BF01876229. [DOI] [Google Scholar]
- Valderrama et al. (2014).Valderrama L, Troche C, Rodriguez MT, Marquez D, Vázquez B, Velázquez S, Vázquez A, Cruz MI, Ressl R. Evaluation of mangrove cover changes in mexico during the 1970–2005 period. Wetlands. 2014;34:747–758. doi: 10.1007/s13157-014-0539-9. [DOI] [Google Scholar]
- Valiela, Bowen & York (2001).Valiela I, Bowen JL, York JK. Mangrove forests: one of the world’s threatened major tropical environments. BioScience. 2001;51:807–815. doi: 10.1641/0006-3568(2001)051[0807:MFOOTW]2.0.CO;2. [DOI] [Google Scholar]
- Villalobos-Zapata & Mendoza-Vega (2010).Villalobos-Zapata GJ, Mendoza-Vega J. La Biodiversidad en Campeche: Estudio de Estado. CONABIO, U.A.C. ECOSUR; Campeche: 2010. [Google Scholar]
- Wang et al. (2010).Wang M, Zhang J, Tu Z, Gao X, Wang W. Maintenance of estuarine water quality by mangroves occurs during flood periods: a case study of a subtropical mangrove wetland. Marine Pollution Bulletin. 2010;60:2154–2160. doi: 10.1016/j.marpolbul.2010.07.025. [DOI] [PubMed] [Google Scholar]
- Wang et al. (2012).Wang Y, Naumann U, Wright ST, Warton DI. Mvabund—an R package for model-based analysis of multivariate abundance data. Methods in Ecology and Evolution. 2012;3:471–474. doi: 10.1111/j.2041-210X.2012.00190.x. [DOI] [Google Scholar]
- Warren-Rhodes et al. (2011).Warren-Rhodes K, Schwarz AM, Boyle LN, Albert J, Agalo SS, Warren R, Bana A, Paul C, Kodosiku R, Bosma W, Yee D, Rönnbäck P, Crona B, Duke NC. Mangrove ecosystem services and the potential for carbon revenue programmes in Solomon Islands. Environmental Conservation. 2011;38:485–496. doi: 10.1017/S0376892911000373. [DOI] [Google Scholar]
- Weller (1995).Weller MW. Use of two waterbird guilds as evaluation tools for the kissimmee river restoration. Restoration Ecology. 1995;3:211–224. doi: 10.1111/j.1526-100X.1995.tb00171.x. [DOI] [Google Scholar]
- Wenny et al. (2011).Wenny DG, DeVault TL, Johnson MD, Kelly D, Sekercioglu CH, Tomback DF, Whelan CJ. The need to quantify ecosystem services provided by birds. The Auk. 2011;128:1–14. doi: 10.1525/auk.2011.10248. [DOI] [Google Scholar]
- Woodroffe et al. (2016).Woodroffe CD, Rogers K, McKee KL, Lovelock CE, Mendelssohn IA, Saintilan N. Mangrove sedimentation and response to relative sea-level rise. Annual Review of Marine Science. 2016;8:243–266. doi: 10.1146/annurev-marine-122414-034025. [DOI] [PubMed] [Google Scholar]
- Wortley, Hero & Howes (2013).Wortley L, Hero JM, Howes M. Evaluating ecological restoration success: a review of the literature. Restoration Ecology. 2013;21:537–543. doi: 10.1111/rec.12028. [DOI] [Google Scholar]
- Zaldívar-Jiménez et al. (2017).Zaldívar-Jiménez A, Ladrón de Guevara-Porras P, Pérez-Ceballos R, Díaz-Mondragón S, Rosado-Solórzano R. US-Mexico joint gulf of Mexico large marine ecosystem based assessment and management: experience in community involvement and mangrove wetland restoration in Términos Lagoon, Mexico. Environmental Development. 2017;22:206–213. doi: 10.1016/j.envdev.2017.02.007. [DOI] [Google Scholar]
- Zaldívar-Jiménez et al. (2010).Zaldívar-Jiménez A, Herrera-Silveira J, Teutli-Hernandez C, Comin F, Andrade JL, Molina CC, Ceballos RP. Conceptual framework for mangrove restoration in the Yucatan Peninsula. Ecological Restoration. 2010;28:333–342. doi: 10.3368/er.28.3.333. [DOI] [Google Scholar]
- Zhang et al. (2012).Zhang JP, Shen CD, Ren H, Wang J, Han WD. Estimating change in sedimentary organic carbon content during mangrove restoration in Southern China using carbon isotopic measurements. Pedosphere. 2012;22:58–66. doi: 10.1016/S1002-0160(11)60191-4. [DOI] [Google Scholar]
- Zhao et al. (2016).Zhao Q, Bai J, Huang L, Gu B, Lu Q, Gao Z. A review of methodologies and success indicators for coastal wetland restoration. Ecological Indicators. 2016;60:442–452. doi: 10.1016/j.ecolind.2015.07.003. [DOI] [Google Scholar]
- Zou et al. (2014).Zou Y, Liu J, Yang X, Zhang M, Tang C, Wang T. Impact of coastal wetland restoration strategies in the chongming dongtan wetlands, China: waterbird community composition as an indicator. Acta Zoologica Academiae Scientiarum Hungaricae. 2014;60:185–198. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Same letters indicate no differences between the study areas. On the right side the hydroperiod components (D–F). The star represents significant differences among different mangrove conditions. The boxes show the inter-quartile range, the thick black lines within the boxes are the medians, and the whiskers are the minimum and maximum values.
(A) show the number of tree species. In (B) the density of mangrove tress is shown. (C) shows the measured height of the mangrove trees at each sampling site. (D) shows the cover expressed as basal area at each sampling site.
The different studied sites are disturbed (triangles), restored (crosses), and undisturbed (circles). In both graphics, bars indicate ±1.5 standard deviations.
Area: restored (R), undisturbed (U), disturbed (D) NOM-059: Is the list of threatened species recognized by the Mexican Environmetal Authorities. The categories are: least concern (LC), subject to special protection (SP), threatened (TH). Pop.Trend are the population trends as reported by the International Union for the Conservation of Nature (IUCN).
H, habitat condition; R, measured Redox; T, water temperature; S, water salinity; Sps, number of tree species; B, Basal area; Dn, trees density; Dt, detectability.
Data Availability Statement
The following information was supplied regarding data availability:
The raw data is available in the Supplemental Files.