

Abstract
The habenula (Hb) is a small midbrain structure that is important for brain signaling and learning from negative events. Thus, the habenula is strongly connected to both the reward system and motor regions. Increasing evidence suggests a role for the habenula in the etiology of psychiatric disorders, including mood and substance use disorders. However, no studies to date have investigated habenular resting-state functional connectivity (rsFC) in suicide-related behaviors (SB). The authors enrolled 123 individuals with major depressive disorder (MDD) or bipolar disorder (BD) and a history of suicide-related behaviors (SB+), 74 individuals with MDD or BD and a history of suicide ideation but no history of SB (SB−), and 75 healthy controls (HC). A seed-based approach was used to identify regions showing different rsFC with the Hb followed by ROI-to-ROI post-hoc comparisons. Compared to both SB− and HC, SB+ showed higher connectivity between the left Hb and the left parahippocampal gyrus, the right amygdala, and the right precentral and postcentral gyri. Patients with mood disorders displayed higher rsFC between the left Hb and left middle temporal gyrus, the left angular gyrus and the left posterior cingulate cortex as well as lower rsFC between the right Hb and the left thalamus, when compared to HC. These findings suggest that the Hb is involved in the neural circuitry of suicide. The higher habenular rsFC found in the SB+ may mediate a dysfunction in the mechanism that links the Hb with motor activity and contextual associative processing.
Keywords: habenula, suicide, bipolar disorder, major depressive disorder, resting-state fMRI
Introduction
Death by suicide is a tragic event that occurs at higher rates among individuals with a psychiatric disorder, and particularly mood disorders {1}. However, it remains largely unclear why not all depressed patients who have suicidal ideation attempt suicide. Theoretical frameworks of suicidal behavior, such as the stress-vulnerability model, indicate that suicidal behavior may result from an interaction between exposure to stressors and a vulnerability in individuals suffering from psychiatric disorders {2}. Suicide attempts have been associated with anhedonia and deficits in reward processing both in elderly {3} and adolescent {4} attempters when compared to non-attempters.
Studies on the neurobiology of suicide suggest that interrelated neurobiological systems linked to dysfunction in the stress response systems are involved. The corticotropin-releasing hormone/ hypothalamic-pituitary-adrenal axis (CRH-HPA) and the locus coeruleus-based norepinephrine (LC-NE) system, which interact with the serotonin and opioid system, are likely to be involved {5}. Despite usually small and heterogeneous, neuroimaging findings report associations between suicidal ideation as well as suicide-related behaviors with abnormalities involving the frontal neural system and the serotonergic system, including the orbitofrontal cortex, the anterior cingulate cortex (ACC) and the amygdala {6–9}. Taken together, these findings suggest that brain regions regulating the stress response, as well as the reward-dopaminergic and serotonergic systems may be involved in the circuitry underlying suicidal ideation and suicide-related behaviors.
One likely locus of concerted control over multiple neurobiological systems is the habenula (Hb), a region that links the forebrain to the midbrain. Indeed, because of its unique anatomical and functional position, the Hb modulates downstream limbic midbrain dopamine and serotonin circuits by controlling activity in the monoaminergic nuclei, the substantia nigra compacta/ventral tegmental area, and the raphe nucleus {10}. Thus, as a relay interface between the basal ganglia and the limbic system, the Hb is involved in both motivational and emotional control of behavior {11}, playing a critical role in both the stress response and the behavioral responses induced by expected reward {12; 13}. Not surprisingly, increasing evidence suggest the involvement of the Hb in the pathophysiology of a variety of psychiatric disorders, including mood disorders, schizophrenia, and substance use disorder {14; 15}. Specifically, increased connectivity and activation of the Hb has been associated with depression {16; 17} as well as sleep disturbances {18}. It is not known, however, whether the Hb is involved in suicidal ideation and suicide-related behaviors.
The aim of the current study was to compare Hb resting state functional connectivity (rsFC) between a sample of patients with mood disorder with and without a history of suicide-related behavior and healthy control subjects. We hypothesized that higher connectivity of the Hb with limbic regions would be associated with suicidal ideation and suicide-related behaviors.
Methods and Materials
Participants
The Baylor College of Medicine Internal Review Board approved the study protocol and procedures. All participants provided informed consent.
Inpatients who met criteria for current major depressive episode and with a primary DSM-IV TR diagnosis of either Major Depressive Disorder, recurrent (MDD; n=165) or Bipolar Disorder type I or type II (BD; n=32) were recruited for the study. They were culled froma sample of 316 inpatients who gave their consent to take part in the study after being admitted at The Menninger Clinic, in Houston, TX USA. Participants with comorbid substance use disorder diagnoses were included, given the pervasive comorbidity between mood and substance use disorders {19} and the need to investigate ecologically valid samples in neuroimaging studies {20}. Psychiatric diagnoses were made using the Structured Clinical Interview for DSM-IV Disorders (SCID-I/II) {21; 22}. Tobacco, alcohol and cannabis were the most prevalent substances of abuse in the sample according to the World Health Organization Alcohol, Smoking, and Substance Involvement Screening Test (WHO-ASSIST). All patients were medicated at the time of the scanning.
Healthy control (HC) subjects were recruited from the general population to serve as comparators for the rsFC analyses. HC were excluded if they had a self-reported history of psychiatric diagnosis or substance abuse. Additional exclusion criteria for both patients and controls were a previous history of traumatic brain injury with loss of consciousness of more than ten minutes or any other MRI contraindication.
Clinical measures
Participants completed a comprehensive diagnostic assessment. Suicidal ideation and suicide-related behaviors were assessed using the Columbia Scale for Rating of Suicide Severity (C-SSRS) {23; 24}. The C-SSRS is a semi structured interview that assesses severity of Suicidal Ideation, Intensity of Ideation, Suicidal Behavior and associated Lethality of Suicide Attempts. The Suicidal Ideation (SI) subscale consists of five items: (1) a passive wish to be dead, (2) nonspecific active thoughts of suicide, (3) active suicidal ideation including methods, (4) suicidal intent without a specific plan, and (5) active suicidal ideation with intent and a plan to act. The Intensity of Ideation subscale rates the frequency, duration, controllability, deterrents, and reasons for thoughts of suicide. The Suicidal Behavior (SB) assesses past (1) Actual Attempts, (2) Nonsuicidal Self-injurious Behavior, (3) Interrupted Attempts, and (4) Aborted Attempts. The lifetime SB subscale was used to stratify groups. Patients were categorized in the suicide-related behaviors group (SB+) if they responded affirmatively to any one of the following three items: Actual Attempt, Interrupted Attempt, or Aborted Attempt. They did not meet criteria for inclusion in the SB+ group if they responded affirmatively solely to the following items: Self-injurious Behavior and Preparatory Acts or Behavior. This approach is consistent with previous studies {25; 26}. Patients were included in the non suicide-related behaviors group (SB−) if they had positive response to any item of the SI but negative response to all items of the SB. The final sample groups included 123 SB+ and 74 SB−.
Depression severity was assessed using the self-rated Patient Health Questionnaire depressive scale {27}. The PHQ-9 is a reliable measure of depression severity that scores each of the 9 diagnostic criteria of Major Depressive Episode over the past 2 weeks as “0” (not at all) to “3” (nearly every day) {28}.
Neuroimaging data acquisition and preprocessing
Participants underwent magnetic resonance imaging of the brain on a 3T Siemens Trio Magnetom system. A structural T1- MPRAGE (TR= 1200ms, TE= 2.66 ms, flip angle= 12°, voxel size= 1mm isotropic, field of view (FOV)= 245 mm, and total sequence time= 4.5 min) was first acquired. A 5-minute resting-state echo-planar imaging (EPI) scan (echo time (TE)= 30 ms, repetition time (TR)= 2000 ms, flip angle= 90°, voxel size= 3.4 × 3.4 × 4 mm, field of view (FOV)= 220 mm) was subsequently acquired for each subject. During the resting-state sequences, a large “X” was displayed on the screen and the participants were asked to keep their eyes open or closed and not fall asleep.
Preprocessing of the functional data including functional realignment and unwarp, functional slice-timing correction, structural segmentation and normalization, functional normalization, ART-based functional outlier detection and scrubbing, and functional smoothing (8 mm Gaussian kernel) were done in MNI-space using CONN- fMRI Functional Connectivity toolbox v15.b {29} with SPM8 (www.fil.ion.ucl.ac.uk/spm/). The Artifact Detection Toolbox (ART; http://www.nitrc.org/projects/artifact_detect/) was used for outlier detection and scrubbing as implemented in CONN. Motion outliers were defined as any frame where the motion exceeded 2 mm. Realignment parameters were entered as first-level covariates in the toolbox. Realignment, scrubbing, white matter, and cerebrospinal fluid were entered as potential confounders in the subject-level General Linear Model (GLM). Using the anatomical component correction (aCompCor) method of flexibly removing physiological noise and movement confounds on a voxel-by-voxel level {30; 31}, each of these effects was regressed out of the BOLD signal before connectivity measures were computed by the toolbox. Functional images were then temporally band-pass filtered (0.01 < f < 0.1 Hz) to investigate low frequency correlations. Since we were interested in the Hb which is a very small region, we downsampled the images to 3×3×3 mm. In addition, the habenular seed in rsFC analysis was not smoothed, to avoid likely contamination with adjacent areas.
Functional connectivity analyses:
We first examined the rsFC between each Hb region of interest (ROI) and each voxel of the brain. The left and right Hb seed ROIs were manually created for every subject, by using the T1 image to visually identify the right and left Hb of each subject landmarks using SPM8 (www.fil.ion.ucl.ac.uk/spm/). In T1 images the Hb is clearly visible as two small triangular structures pointing into the third ventricle. Each functional ROI was a 3 × 3 × 3 mm cube placed around a central MNI coordinate in the Hb.
The mean signal time course from the seeds were extracted and the Pearson’s correlations coefficients with the time course of all other voxels of the brain were calculated. Correlations maps were calculated for each subject and correlation coefficients were then converted to normally distributed z-scores using the Fisher transformation in order to perform GLM analyses. In order to compare rsFC across the three groups (SB+, SB−, HC) assuming as null hypothesis that the three groups would show equal rsFC with the left and right habenula seeds, ANCOVA with age and gender included as covariates was performed. A voxel statistical height threshold of p < 0.001 with a cluster threshold of p < 0.05 family-wise error (FWE) corrected was used to identify connectivity differences between the three groups. Significant clusters identified in this analysis were then imported as target ROIs in the post-hoc ROI-to-ROI analysis. The connectivity values between the significant clusters and left and right Hb were extracted from each participant in order to examine the nature of the connectivity in each group and the directionality of each post-hoc contrast. The results were considered significant if p< 0.05 False Discovery Rate (FDR) corrected. To further explore whether additional factors such as MDD/BD diagnosis, substance use/abuse disorder, or medications might have affected the significant results in the between patient group comparison (SB+ vs SB−) we subsequently included these covariates in the model in a post-hoc manner to avoid reducing statistical power of the original ANCOVA. Thus, mood disorder diagnosis, alcohol and substance use/abuse disorder, and the effects of antidepressants, lithium, antiepileptics, anxiolytics and antipsychotics were entered as dichotomous covariates (absent vs. present).
Additionally, to address whether the significant findings (clusters displayed in Table 2) would be specific to the habenula signal we conducted an exploratory SB+ vs SB− ROI-to-ROI comparison including thalamic regions as seeds. Specifically, the whole bilateral thalamus based on the automated anatomical labeling atlas {32} as well as two small bilateral thalamic control ROIs were chosen as seeds. The two small bilateral thalamic seeds were created by shifting the averaged Hb ROIs coordinates by 9 mm in the anterior direction (“thalamus medial”, MNI: right 4, −15, 2; left −3, −16, 2), and in the lateral direction (“thalamus lateral”, MNI: right 13, −24, 2; left −12, −25, 2). The results were considered significant if p< 0.05 False Discovery Rate (FDR) corrected.
TABLE 2.
Seed-to-voxel analysis: any between-group differences significant results across the 3 groups.
| Region | Side | BA | Cluster size | Cluster p-value | Peak Z Value | Peak Voxel MNI Coordinates | ||
|---|---|---|---|---|---|---|---|---|
| L habenula seed | x | y | z | |||||
| middle temporal gyrus | L | 21 | 1139 | <0.001 | 4.71 | −52 | −10 | −24 |
| angular gyrus | L | 39 | 817 | < 0.001 | 4.23 | −36 | −62 | 50 |
| posterior cingulate | L | 31 | 711 | < 0.001 | 4.9 | −2 | −54 | 32 |
| parahippocampal gyrus | L | - | 313 | 0.002 | 4.54 | −30 | −22 | −22 |
| postcentral gyrus | R | 1 | 207 | 0.005 | 4.34 | 30 | −32 | 46 |
| amygdala | R | - | 173 | 0.031 | 3.86 | 16 | 0 | −24 |
| precentral gyrus | R | 4 | 156 | 0.046 | 3.74 | 44 | −22 | 70 |
| R habenula seed | ||||||||
| thalamus | L | - | 179 | 0.026 | 6.68 | −4 | −24 | 2 |
(SB−, SB+, HC), where SB− = negative history for suicide-related behaviors, SB+= positive history for suicide related behaviors, and HC= healthy controls.
BA= Brodmann area; MNI= Montreal Neurological Institute.
Results
Demographic and clinical data are reported in Table 1. The three groups did not significantly differ on age, gender, and ethnicity. The SB+ and SB− differed on the presence of psychotic features (p= 0.026), lifetime suicidal ideation (p< 0.001), and lithium (p= 0.010). No additional differences were found between patient groups.
Table 1.
Demographic and clinical variables
| SB− | SB+ | HC | F or χ2 or Fisher | df | p | ||||
|---|---|---|---|---|---|---|---|---|---|
| n=74 | n=123 | n=75 | |||||||
| Value | SD or % | Value | SD or % | Value | SD or % | ||||
| Age (years), mean (SD) | 30.3 | (11.5) | 29.7 | (11.8) | 33.1 | (9.1) | 2.26 | 2 | 0.106 |
| Gender (n), Male/Female | 38/37 | 56/67 | 38/37 | 2.34 | 2 | 0.309 | |||
| Ethnicity (n) (Caucasian/Other) | 68/6 | 110/13 | 63/12 | 2.45 | 2 | 0.294 | |||
| MDD diagnosis (n) (%) | 62 | (83.8) | 103 | (83.8) | - | - | 0.000 | 1 | 0.994 |
| Psychosis (n) (%) | 6 | (8) | 2 | (1.6) | - | - | 4.95 | 1 | 0.026 |
| Personality disorder (n) (%) | 35 | (47.3) | 64 | (52) | - | - | 0.414 | 1 | 0.520 |
| Substance use/abuse disorder (n) (%) | 39 | (53) | 52 | (42) | - | - | 2.02 | 1 | 0.155 |
| PHQ9-Depression (mean) (SD) | 16.1 | (6.8) | 17.5 | (6.1) | - | - | 2.06 | 1 | 0.153 |
| C-SSRS Ideation lifetime | 12.2 | (3.5) | 14.6 | (3.5) | - | - | 21.9 | 1 | <0.001 |
| C-SSRS total number suicide-related behaviors* lifetime | - | 4.6 | - | - | |||||
| Psychotropic medications (n) (%) | |||||||||
| Antidepressants (n) (%) | 62 | (83.8) | 103 | (83.8) | - | - | 0.000 | 1 | 0.994 |
| Lithium (n) (%) | 5 | (6.7) | 25 | (20.3) | - | - | 6.58 | 1 | 0.010 |
| Antiepileptics (n) (%) | 28 | (37.8) | 53 | (43.1) | - | - | 0.526 | 1 | 0.468 |
| Anxiolytics (n) (%) | 24 | (32.4) | 38 | (30.9) | - | - | 0.51 | 1 | 0.822 |
| Antipsychotics (n) (%) | 28 | (37.8) | 50 | (40.6) | - | - | 0.153 | 1 | 0.696 |
SB− = negative history for suicide-related behaviors; SB+ = positive history for suicide-related behaviors; HC= healthy controls; MDD= Major Depressive Disorder; PHQ-9= Patient Health Questionnaire depressive scale; C-SSRS= Columbia Scale for Rating of Suicide Severity.
Any affirmative response to items: Actual Attempt, Interrupted Attempt, or Aborted Attempt.
Risk level from “moderate” to “high”.
Whole-brain between group analysis
Results are presented in Table 2 and Figure 1.
Figure 1. Seed-to-voxel analysis: any between-group differences results across the 3 groups.

(SB−, SB+, HC), where SB− = negative history for suicide-related behaviors, SB+= positive history for suicide related behaviors, and HC= healthy controls. Significant clusters (p< 0.05 FWE-corrected) are overlaid on canonical images (ch2-bet template). Color schemes: green – significant clusters with the left habenula seed; violet – significant cluster with the right habenula seed. Talairach x,y,z- coordinates above each slice (left hemisphere corresponds to the left side of the image).
Age- and gender- adjusted ANCOVAs demonstrated that the three groups significantly differed in rsFC between the left Hb and the left middle temporal gyrus, the left angular gyrus, the left posterior cingulate; the left parahippocampal gyrus, the right amygdala, and the right precentral and postcentral gyri. The three groups significantly differed also in rsFC between the right Hb and the left thalamus.
ROI-to-ROI analysis
Results are presented in Table 3.
TABLE 3.
ROI-to-ROI post-hoc analysis results.
| RSFC pairings | SB+ > SB− | SB+ > HC | SB− > HC |
|---|---|---|---|
| L Hb - L parahippocampal g. |
t=4.94 p <0.001** |
t=4.71 p <0.001** |
t= −0.34 p= 0.734 |
| L Hb – R amygdala |
t=4.48 p <0.001** |
t=3.77 p =0.002** |
t= −0.44 p= 0.658 |
| L Hb – R precentral g. |
t=2.60 0.023* |
t=4.66 p <0.001** |
t= 2.03 p= 0.044* |
| L Hb – R postcentral g. |
t=2.50 p= 0.023* |
t=5.59 p< 0.001** |
t= 2.69 p< 0.01* |
| L Hb – L middle temporal g. |
t= 0.57 p= 0.064 |
t= 5.97 p< 0.001** |
t= 5.04 p< 0.001** |
| L Hb – L angular g. |
t= −3.72 p= 0.054 |
t=3.72 p =0.003* |
t= 5.03 p< 0.001** |
| L Hb – L posterior cingulate |
t= 1.25 p= 0.310 |
t= 4.80 p< 0.001** |
t= 3.40 p< 0.001** |
| R Hb – L thalamus |
t= − 1.07 p=0.286 |
t= −7.95 p< 0.001** |
t= −6.08 p< 0.001** |
SB− = negative history for suicide-related behaviors, SB+= positive history for suicide related behaviors, and HC= healthy controls; RSFC= resting-state functional connectivity; L= left; R= right;
p< 0.001
p< 0.05; t= t-value; p= p-value
Compared to SB−, SB+ showed higher rsFC between the left Hb and the left parahippocampal gyrus, the right amygdala, the right precentral, and right postcentral gyri (see Figure 2). After including mood disorder diagnosis, substance use and medications in the ANCOVA model, results remain significant except for the rsFC between the left Hb and the right postcentral (p=0.124) and precentral (p=0.059) gyri. No differences between the two groups were found with the right Hb seed. Compared to HC, both SB+ and SB− showed higher connectivity between the left Hb and the left middle temporal gyrus, left angular gyrus, left posterior cingulate and right postcentral gyrus. Additionally, both SB+ and SB− presented lower connectivity compared to HC between the right Hb and the left thalamus. SB+ presented also higher connectivity compared to HC between the left Hb and the left parahippocampal gyrus, and right precentral gyrus.
Figure 2.
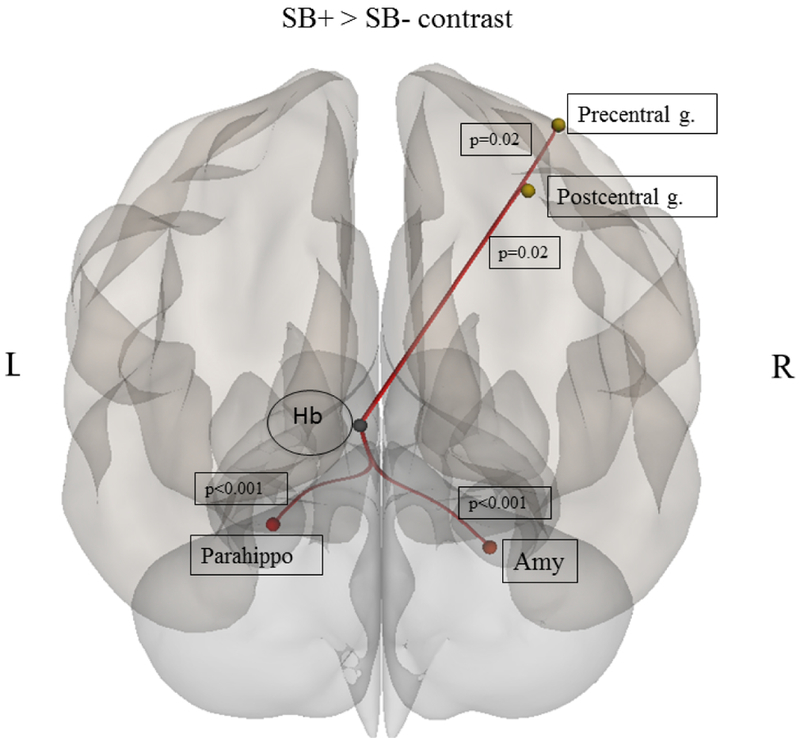
Results of between group (SB−, SB+) ROI-to-ROI post-hoc comparison, where SB− = negative history for suicide-related behaviors and SB+= positive history for suicide-related behaviors. Abbreviations: Hb = habenula; Precentral g.= precentral gyrus; Postcentral g.= postcentral gyrus; Parahippo= parahippocampal gyrus; Amy= amygdala.
As for the exploratory comparison SB− vs. SB+ with the thalamic seeds, no significant results were found between any of the target regions and either the whole AAL thalamus or thalamus lateral seeds. However, compared to SB−, SB+ showed higher rsFC between the left and right thalamus medial seeds and the left parahippocampal gyrus [Left thalamus: t=3.03, p=0.003; Right: t= 2.70, p=0.007], the right amygdala [Left thalamus t=2.93, p= 0.004; Right thalamus: t= 3.90, p< 0.001], and the right postcentral gyrus [Left thalamus: t= 2.90, p=0.004; Right thalamus: t= 2.85, p= 0.005]. SB+ also showed higher connectivity between the left thalamus medial and the posterior cingulate (t= 3.21, p=0.001).
Discussion
The present study investigated habenular rsFC differences between patients with a diagnosis of mood disorder with and without a history of suicide related-behaviors, and healthy controls. To the best of our knowledge, it represents the largest study investigating neuroimaging correlates of suicidal-related behaviors in a population of mood disorder patients. As hypothesized, we found that SB+ had higher connectivity between left Hb and several regions: left Hb and left parahippocampal gyrus, left and right amygdala, and right precentral and postcentral gyri. Moreover, patients with a diagnosis of mood disorder, independently of suicide-related status, displayed higher connectivity between left Hb and middle temporal gyrus, angular gyrus and PCC as well as decreased connectivity between right Hb and left thalamus. The present results suggest that altered connectivity of the Hb with regions of the limbic and motor systems might be involved in the neural basis of suicide-related behaviors.
Typically described as composed by medial and lateral subregions, the Hb controls behavior, cognition and emotion through extensive inputs from the basal ganglia and the limbic regions {33}. Based on these inputs, the fasciculus retroflexus carries outputs that target dopaminergic and serotonergic neurons {12}. By inhibiting the firing activity of the dopamine and serotonin systems, the Hb facilitates the evolutionary mechanism of action avoidance when the outcome is less rewarding than predicted {11}.
Our data support the evidence that altered activity of the Hb contributes to the etiology of psychiatric disorder, including depression {34}. Evidence that the Hb is hyperactive during depression comes from animal models of depression {33} and pharmacological studies {35}. Neuroimaging studies of Hb activity in MDD patients reported contrasting results. While increased activity of the Hb was detected in remitted MDD patients {36}, reduced activity was observed in depressed MDD during the prediction of negative outcome {37} and primary aversive conditioning {38}. The investigation of an association between Hb connectivity and suicide is new to the literature. However, the higher connectivity of the left Hb we found in SB+ is somewhat in line with the disrupted Hb function observed in depression. Thus, it is tempting to speculate that in the SB+ the higher connectivity with regions of the limbic and motor systems might be related, at least partially, to a possible increased sensitivity of Hb cells induced by chronic stress {12; 39; 40}. Although the exact mechanism that links suicide and chronic stress is not fully understood, stress-related alterations in cell structure have been documented in suicide attempters {41}. Chronic dysregulation of the habenula circuit may lead to long-term alterations of dopamine, serotonin, and norepinephrine release that impact behavioral functioning, particularly the learning of maladaptive coping strategies related to suicide-related behaviors {42}. Interestingly, the parahippocampal gyrus has been linked to suicidality: in an fMRI experiment using a delay discount task, suicidal patients showed longer reward delays were associated with diminished parahippocampal response {43}. The parahippocampal cortex has been hypothesized to be a critical part of a network of brain regions that processes contextual associations {44}. This makes functional connectivity with the habenula a possibly important feature of contextual associations in general, and especially those that include reward as an important feature.
Interestingly, the left Hb was strongly associated with suicidality while the right Hb was much less involved. Habenular asymmetry has been shown to be critically important for behavior in fish {45}, and more recently, rsFC of the habenula in healthy humans was shown to be asymmetric {46}. Our data suggest that habenular asymmetry may be also important for psychiatric disorder etiology, which may have implications in terms of future therapeutic intervention development, especially for neuromodulatory approaches. For example, deep brain stimulation of the Hb has been shown to possibly be therapeutic in treatment-resistant major depressive disorder {47}. This might imply that unilateral Hb stimulation, which arguably could have lesser side effects, should be explored.
Not surprisingly, significant results were also found when the medial thalamic seed was considered. While we cannot exclude this might be partially attributable to the spatial distortion of the MRI images, it could also be attributable to shared connectivity patterns between the Hb and the medial nuclei of the thalamus. Overlapping and mutually interacting circuits – e.g., similar inputs from the ventral/limbic striatum - between the two regions are already described {17; 48}.
It is important to address several limitations in the present study. First, the reliability of in vivo study of the Hb through fMRI has been debated {17; 49}, considering the small size of this structure. We tried to temper this caveat using individual-specific Hb ROIs, without attempting to isolate the medial and lateral subdivisions. However, we cannot exclude that head movement as well as spatial resolution and smoothing might have contributed to a signal contamination from the surrounding regions. Second, the current study examined suicide behavior in a mixed sample of depressed subjects diagnosed with MDD or BD. Although this study includes patients with diagnostic heterogeneity, the two disorders tend to be more similar than not when it comes to suicide-related behaviors {50}. All individuals in the SB+ and SB− groups were prescribed multiple medications at the time of MRI. Although medication status was secondarily included as covariates in the analyses performed here, the potential effects of medication status on rsFC cannot be disentangled fully from those effects attributable to suicidal behavior. Of note, a recent review of the literature {51} concluded that medication effects do not seem to provide an alternative explanation for observed differences in BOLD signal.
In summary, the present findings suggest that the Hb might be involved in the neural circuitry of suicide. As such, the higher habenular rsFC we found in the suicide-related behaviors group may mediate a dysfunction in the critical survival mechanism by which the habenula works as a suppressor of motor activity. Nonetheless, additional research focused on the identification of neuroimaging differences between patients with only suicidal ideation from those with suicide-related behaviors is needed in order to identify patients at higher risk to attempt suicide.
Acknowledgments
This work was supported by the McNair Medical Institute; American Foundation for Suicide Prevention (SRG-2–125-14); NARSAD (19295); the Veteran Health Administration (VHA5I01CX000994); and the National Institute of health (NIDA DA026539, DA09167). This research was partially supported by the Menninger Clinic Foundation; The Brown Foundation, Inc. of Houston, Texas; the George and Mary Josephine Hamman Foundation; and the Toomim Family Fund. Dr. Madan is a McNair Scholar. This material is partly the result of work supported with resources and the use of facilities at the Michael E. DeBakey VA Medical Center, Houston, TX. The authors thank the Core for Advanced MRI at Baylor College of Medicine, and Dr. Charles Neblett. The study follows the guidelines on good publication practices (61). The study sponsors were not involved in any aspect of research activities and did not approve the specific protocol or manuscript. Thus, the authors were independent from study sponsors in the context of research.
Footnotes
Financial Disclosures
None of the authors have any potential conflicts of interests to declare.
References
- 1.Oquendo MA, Galfalvy H, Russo S, et al. : Prospective study of clinical predictors of suicidal acts after a major depressive episode in patients with major depressive disorder or bipolar disorder. [Internet]. Am. J. Psychiatry 2004; 161:1433–41 [DOI] [PubMed] [Google Scholar]
- 2.Hawton K, van Heeringen K: Suicide. Lancet 2009; 373:1372–1381 [DOI] [PubMed] [Google Scholar]
- 3.Dombrovski AY, Szanto K, Clark L, et al. : Reward Signals, Attempted Suicide, and Impulsivity in Late-Life Depression. JAMA Psychiatry 2013; 70:1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Auerbach RP, Millner AJ, Stewart JG, et al. : Identifying differences between depressed adolescent suicide ideators and attempters. J. Affect. Disord 2015; 186:127–133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Oquendo MA, Sullivan GM, Sudol K, et al. : Toward a Biosignature for Suicide. Am. J. Psychiatry 2014; 171:1259–1277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cox Lippard ET, Johnston J a. Y, Blumberg HP: Neurobiological Risk Factors for Suicide. Am. J. Prev. Med 2014; 47:S152–S162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Serafini G, Pardini M, Pompili M, et al. : Understanding suicidal behavior: The contribution of recent resting-state fMRI techniques. Front. Psychiatry 2016; 7:1–4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Oquendo MA, Galfalvy H, Sullivan GM, et al. : Positron Emission Tomographic Imaging of the Serotonergic System and Prediction of Risk and Lethality of Future Suicidal Behavior. JAMA Psychiatry 2016; 10032:1–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Spoletini I, Piras F, Fagioli S, et al. : Suicidal attempts and increased right amygdala volume in schizophrenia. Schizophr. Res 2011; 125:30–40 [DOI] [PubMed] [Google Scholar]
- 10.Klemm WR: Habenular and interpeduncularis nuclei: shared components in multiple-function networks. Med. Sci. Monit 2004; 10:RA261–A273 [PubMed] [Google Scholar]
- 11.Hikosaka O, Sesack SR, Lecourtier L, et al. : Habenula: crossroad between the basal ganglia and the limbic system. J. Neurosci 2008; 28:11825–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hikosaka O: The habenula: from stress evasion to value-based decision-making. Nat. Rev. Neurosci 2010; 11:503–513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Salas R, Baldwin P, de Biasi M, et al. : BOLD Responses to Negative Reward Prediction Errors in Human Habenula. Front. Hum. Neurosci 2010; 4:36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fakhoury M: The habenula in psychiatric disorders: More than three decades of translational investigation. Neurosci. Biobehav. Rev 2017; 10.1016/j.neubiorev.2017.02.010 [DOI] [PubMed]
- 15.Velasquez KM, Molfese DL, Salas R: The role of the habenula in drug addiction [Internet]. Front. Hum. Neurosci 2014; 8:174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Proulx CD, Hikosaka O, Malinow R: Reward processing by the lateral habenula in normal and depressive behaviors. Nat. Neurosci 2014; 17:1146–1152 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ely BA, Xu J, Goodman WK, et al. : Resting-state functional connectivity of the human habenula in healthy individuals: Associations with subclinical depression. Hum. Brain Mapp 2016; 37:2369–2384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Aizawa H, Cui W, Tanaka K, et al. : Hyperactivation of the habenula as a link between depression and sleep disturbance. Front. Hum. Neurosci 2013; 7:826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Conway KP, Compton W, Stinson FS, et al. : Lifetime comorbidity of DSM-IV mood and anxiety disorders and specific drug use disorders: results from the National Epidemiologic Survey on Alcohol and Related Conditions. J. Clin. Psychiatry 2006; 67:247–57 [DOI] [PubMed] [Google Scholar]
- 20.Greene DJ, Black KJ, Schlaggar BL: Considerations for MRI study design and implementation in pediatric and clinical populations. Dev. Cogn. Neurosci 2016; 18:101–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.First M, Gibbon M, Spitzer R: Structured Clinical Interview for DSM-IV Axis II Personality Disorders (SCID-II). Am. Psychiatr Press Inc, 1997
- 22.First MB, Spitzer RL, Gibbon M, and Williams JB : Structured Clinical Interview for DSM-IV-TR Axis I Disorders. New York State Psychiatr Inst, 2002
- 23.Posner K, Brown GK, Stanley B, et al. : The Columbia-suicide severity rating scale: Initial validity and internal consistency findings from three multisite studies with adolescents and adults. Am. J. Psychiatry 2011; 168:1266–1277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Madan A, Frueh BC, Allen JG, et al. : Psychometric reevaluation of the columbia-suicide severity rating scale: Findings from a prospective, inpatient cohort of severely mentally ill adults. J. Clin. Psychiatry 2016; 77:e867–e873 [DOI] [PubMed] [Google Scholar]
- 25.Minzenberg MJ, Lesh T, Niendam T, et al. : Conflict-related anterior cingulate functional connectivity is associated with past suicidal ideation and behavior in recent-onset schizophrenia. J. Psychiatr. Res 2015; 65:95–101 [DOI] [PubMed] [Google Scholar]
- 26.Minzenberg MJ, Lesh TA, Niendam TA, et al. : Conflict-Related Anterior Cingulate Functional Connectivity Is Associated With Past Suicidal Ideation and Behavior in Recent-Onset Psychotic Major Mood Disorders. J. Neuropsychiatry Clin. Neurosci 2016; 28:299– 305 [DOI] [PubMed] [Google Scholar]
- 27.Löwe B, Kroenke K, Herzog W, et al. : Measuring depression outcome with a brief self-report instrument: Sensitivity to change of the Patient Health Questionnaire (PHQ-9). J. Affect. Disord 2004; 81:61–66 [DOI] [PubMed] [Google Scholar]
- 28.Kroenke K, Spitzer RL, Williams JBW: The PHQ-9: Validity of a brief depression severity measure. J. Gen. Intern. Med 2001; 16:606–613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Whitfield-Gabrieli S, Nieto-Castanon A: Conn: A Functional Connectivity Toolbox for Correlated and Anticorrelated Brain Networks. Brain Connect 2012; 2:125–141 [DOI] [PubMed] [Google Scholar]
- 30.Behzadi Y, Restom K, Liau J, et al. : A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. Neuroimage 2007; 37:90–101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chai XJ, Castañán AN, Öngür D, et al. : Anticorrelations in resting state networks without global signal regression. Neuroimage 2012; 59:1420–1428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tzourio-Mazoyer N, Landeau B, Papathanassiou D, et al. : Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002; 15:273–289 [DOI] [PubMed] [Google Scholar]
- 33.Benarroch EE: Habenula Recently recognized functions and potential clinical relevance. Neurology 2015; 85:992–1000 [DOI] [PubMed] [Google Scholar]
- 34.Loonen AJM, Ivanova SA: Circuits Regulating Pleasure and Happiness—Mechanisms of Depression. Front. Hum. Neurosci 2016; 10:571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Freo U, Ori C, Dam M, et al. : Effects of acute and chronic treatment with fluoxetine on regional glucose cerebral metabolism in rats: implications for clinical therapies. Brain Res 2000; 854:35–41 [DOI] [PubMed] [Google Scholar]
- 36.Roiser JP, Levy J, Fromm SJ, et al. : The Effects of Tryptophan Depletion on Neural Responses to Emotional Words in Remitted Depression. Biol. Psychiatry 2009; 66:441–450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Furman D, Gotlib IH: Habenula Responses to Potential and Actual Loss in Major Depression: Preliminary Evidence for Lateralized Dysfunction. Soc. Cogn. Affect. Neurosci 2016; 843–851 [DOI] [PMC free article] [PubMed]
- 38.Lawson RP, Drevets WC, Roiser JP: Defining the habenula in human neuroimaging studies. Neuroimage 2013; 64:722–727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kaye A, Ross DA: The Habenula: Darkness, Disappointment, and Depression. Biol. Psychiatry 2017; 81:e27–e28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Li B, Piriz J, Mirrione M, et al. : Synaptic potentiation onto habenula neurons in the learned helplessness model of depression. Nature 2011; 470:535–539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lindqvist D, Fernström J, Grudet C, et al. : Increased plasma levels of circulating cell-free mitochondrial DNA in suicide attempters: associations with HPA-axis hyperactivity. Transl. Psychiatry 2016; 6:e971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Brüdern J, Berger T, Michel K, et al. : Are Suicide Attempters Wired Differently? J. Nerv. Ment. Dis 2015; 203:514–521 [DOI] [PubMed] [Google Scholar]
- 43.Vanyukov PM, Szanto K, Hallquist MN, et al. : Paralimbic and lateral prefrontal encoding of reward value during intertemporal choice in attempted suicide. Psychol. Med 2016; 46:381– 391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Aminoff EM, Kveraga K, Bar M: The role of the parahippocampal cortex in cognition. Trends Cogn. Sci 2013; 17:379–390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Aizawa H: Habenula and the asymmetric development of the vertebrate brain. Anat Sci Int 2013; 88:1–9 [DOI] [PubMed] [Google Scholar]
- 46.Hétu S, Luo Y, Saez I, et al. : Asymmetry in functional connectivity of the human habenula revealed by high-resolution cardiac-gated resting state imaging. Hum. Brain Mapp 2016; 37:2602–2615 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sartorius A, Kiening KL, Kirsch P, et al. : Remission of Major Depression Under Deep Brain Stimulation of the Lateral Habenula in a Therapy-Refractory Patient. Biol. Psychiatry 2010; 67:e9–e11 [DOI] [PubMed] [Google Scholar]
- 48.Ikemoto S, Yang C, Tan A: Basal ganglia circuit loops, dopamine and motivation: A review and enquiry. Behav. Brain Res 2015; 290:17–31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Loonen AJM, Ivanova SA: Commentary on “A non-reward attractor theory of depression”: a Proposal to include the Habenula Connection. Neurosci Biobehav Rev 2017; http://linkinghub.elsevier.com/retrieve/pii/S0149763416305577 [DOI] [PubMed]
- 50.Nock MK, Hwang I, Sampson NA, et al. : Mental disorders, comorbidity and suicidal behavior: results from the National Comorbidity Survey Replication. Mol. Psychiatry 2010; 15:868–76 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hafeman DM, Chang KD, Garrett AS, et al. : Effects of medication on neuroimaging findings in bipolar disorder: an updated review. Bipolar Disord 2012; 14:375–410 [DOI] [PubMed] [Google Scholar]