

Abstract
In sports, peripheral vision is expected to play an important role in tasks that demand distributed attention and motion-change detection. By using the Multiple-Object-Tracking (MOT) task, these demands were simulated in a well-controlled laboratory environment. Participants tracked four target out of ten moving objects (6 distractors) and pressed a button when one of the ten objects stopped. Detection rates for tracked targets were compared to detection rates of non-tracked distractors at eccentricities between 5° and 25°. The study's aim was to test how the location of attention affects peripheral motion detection. The results show a large attention effect because target stops were detected in 89 % and distractor stops only in 55 % of the trials. Distractor stops were more likely detected when they occurred closer to the fovea while target stops were detected at all eccentricities. That means, orienting attention at target objects facilitates the peripheral detection of their motion changes in monitoring tasks. Having distractors closer to the fovea increases the chance to also detect motion changes of unattended objects. On a theoretical level, results support a tracking mechanism with object-based attention, serial covert attention shifts and flexible but limited attentional resources. On a practical level, sports' experts should use their extensive knowledge to locate attention on most-relevant objects and reduce the eccentricity to other objects to detect motion changes of attended and unattended objects.
Keywords: Neuroscience, Covert attention, Change detection, Sustained attention, Eye movement, Multiple object tracking
1. Introduction
In a soccer corner kick situation, the goalkeeper must process two sources of information: the ball flight and the position of players surrounding him. Thus, attention must be distributed to multiple peripheral locations (up to 21 players in the goalkeeper's visual environment). Since the attentional capacity of the goalkeeper is likely too low to attend to the movements of all players, selective attention is located on players that are likely to score a goal while other players receive less or even no attention. This selection, however, can be costly if a player receiving no attention suddenly initiates a movement to score a goal. The question arises: How does selective visual attention in such a monitoring task affect the motion detection of attended compared with unattended objects that are located in peripheral vision?
In sports situations, such as the one explained above, it was emphasized that peripheral vision has to be used to process relevant motion information of players (Vater et al., 2017c). This strategy, to make use of peripheral vision, seems most functional as peripheral vision is used for encoding a dynamically changing visual environment (Rosenholtz, 2016). Also, motion processing is, in contrast to visual acuity, less impaired in the periphery when compared with the fovea (To et al., 2011). Since visual selective attention has been shown to affect visual capabilities, this motion sensitivity might be enhanced at attended locations (Carrasco et al., 2006).
To examine how selective attention affects peripheral motion sensitivity, the Multiple Object Tracking (MOT) task (Pylyshyn and Storm, 1988) seems to mirror the demands of the soccer corner kick situation (Faubert and Sidebottom, 2012). In MOT, a set of identically looking objects is presented and the to-be-tracked targets are initially highlighted. After this highlighting phase, all objects begin to move and can collide with another and change their direction. After this motion phase, all objects stop and the task for participants is to recall the initially highlighted targets. Interestingly, during the monitoring process of the targets, eye-tracking studies have shown that gaze is often located on the “centroid” location (center of mass of the targets), such that targets are monitored using peripheral vision (Fehd and Seiffert, 2008, 2010; Vater et al., 2017a; Zelinsky and Neider, 2008) or, in attentional terms, selective covert attention (Giordano et al., 2009; Vater et al., 2016). At the same time, identically looking non-targets (distractors) are likely to be inhibited during the monitoring of the targets (Bettencourt and Somers, 2009; Huff et al., 2012; Pylyshyn, 2006; Pylyshyn et al., 2008; Sears and Pylyshyn, 2000). The analysis of ERP signals revealed, that neural activity for targets is enhanced but that there is at least no active suppression of distractors because N1 amplitudes for probes on distractors were the same as for probes on empty space but lower than on targets (Drew et al., 2009). This finding is supported by behavioral studies, for example, by Alvarez and Oliva (2008) found that participants can locate a missing target or distractors after a tracking phase and results show that even distractors are represented above chance level. Similarly, Meyerhoff et al. (2015) could show that distractors are integrated in target tracking, because displacements of distractors impaired target tracking. Thus, attention is most likely enhanced on targets compared with distractors but information of distractors are incorporated in target tracking.
Meyerhoff et al. (2017) provide a detailed overview on existing MOT theories and tracking mechanisms, including the original FINST theory (Pylyshyn, 1989, 2001), the perceptual grouping model (Yantis, 1992), the multifocal attention theory (Cavanagh and Alvarez, 2005), the FLEX model (Alvarez and Franconeri, 2007), and the spatial interference theory (Franconeri et al., 2010). When summarizing common features and empirical evidence for those theories, MOT tracking mechanisms are characterized by, among others, limited but flexible attentional resources, the use of motion information for tracking but not necessarily for extrapolation of target-motion paths, and an object-based (especially target-based) rather than space-based attention (Meyerhoff et al., 2017).
When transferring these MOT findings to the goalkeepers tracking problem, it is likely that actions of attended players are better detected than actions of unattended players because they receive more attention (as do targets in MOT). Nevertheless, since distractors in MOT are presumably not actively inhibited and attentional locations can be flexibly adjusted, the detection of movements of unattended players might still be possible, especially because peripheral vision is motion sensitive and sudden motion changes are known to capture visual attention (Cosman and Vecera, 2010; Pratt et al., 2010).
Some studies already looked at detection rates for changes of targets (with a high amount of attention) and distractors (with a low amount of attention). For example, Sears and Pylyshyn (2000) contrasted detection rates for target- and distractor-form changes and found that responses to distractor changes were delayed compared with responses to targets. In line with that, Bahrami (2003) found that color and shape changes are more often missed in distractors than in targets. A limitation of these studies is that they do not take the visual system into account because it is know that eccentricity affects peripheral form and color vision (Hansen et al., 2009; Rosenholtz, 2016; Strasburger, 2005; Strasburger et al., 2011). Since the eccentricity of perceived changes was not measured in these studies, it remains unclear if peripheral vision was used to different degrees in their experimental conditions and might explain their attention effect on target and distractor changes. Moreover, these studies did not use a motion-change detection task that is most relevant to the current research question.
An applicable method on how to measure the eccentricity of change events has been used by Vater et al. (2017a) who could show that form-change detection rates of targets decrease with eccentricity while motion-change detection rates of targets do not. They also controlled that this eccentricity effect is not due to saliency differences between the form- and motion-change conditions and compared a target slowdown with lower saliency with a target stop with higher saliency. Results showed that both, stops and slowdowns, are equally well detected at both eccentricities. On the one hand, these results are in line with the expected motion-sensitivity in peripheral vision. On the other hand it underlines that the results obtained by Sears and Pylyshyn (2000) and Bahrami (2003) could be influenced by eccentricity. Thus, a comparison of the ability to detect target- and distractor-motion changes with peripheral vision where retinal eccentricity of the change events is controlled with eye-tracking is still pending.
Together, previous findings indicate that attention is mostly bound to the targets and only to some degree located on distractors and that peripheral vision is used for the monitoring of targets’ motion and the detection of target-motion changes. So far studies comparing change-detection rates for targets and distractors did not measure visual eccentricity of perceived changes and did not apply the detection of motion changes on targets and distractors. Therefore, the current study will look at the effect of attention on motion-change detection in peripheral vision. Two competing hypotheses will be tested in the current study: 1) if peripheral motion detection is independent of the location of attention and purely bottom-up, motion-changes should be detected on targets and distractors to similar degree and 2) if peripheral motion detection is dependent on the location of attention, which is in MOT located on the targets, higher detection rates should be observed for target- than for distractor-motion changes.
2. Methods
2.1. Participants
Fourteen students (seven women and seven men; aged 20.4 ± 0.9 years) participated in the experiment and received course credits in return. The sample size was determined a priori on the basis of previous studies (Fehd and Seiffert, 2008; Vater et al., 2017a), and checked a priori with a G*Power analysis based on the effect sizes found in Vater et al. (2017a), which, due to large effect sizes (all > ηp2 = .33), revealed a minimum of 10 participants. Participants had self-reported normal or corrected to-normal vision and were unaware of the research question. The experiment was conducted in accordance with the Declaration of Helsinki and ethics were approved by ethics committee of the university.
2.2. Stimuli
MATLAB (2016) was used to calculate the linear motion paths of 10 white squares (35 mm × 35 mm corresponding to 1° x 1° of visual angle). At the beginning of each trial, all squares appeared in (quasi-)random starting positions (no object overlap) and the 4 targets were highlighted by red frames (line width: 15 mm; frame and stimulus together covering an area of 1.7° x 1.7° of visual angle). After 2 s, the target-defining cues disappeared, and all stimuli accelerated on straight-line paths for 1 s to reach a final speed of 6°/s. This final speed was sustained for 4 s, followed by a deceleration phase of 1 s, after which all squares stopped. This pattern resulted in a total object-motion duration of 6 s. In the following and final 3 s of each trial, a number appeared on each object and participants were to identify the initially highlighted targets by naming the respective numbers projected onto the now stationary squares. During the motion phase, a repulsion mechanism (Fehd and Seiffert, 2008, 2010; Vater et al., 2016, 2017a) was used to redirect a square whenever the distance from the rectangular frame or the next square fell below a certain threshold (35 mm corresponding to 1° of visual angle). Based on these constraints, 40 root trials were created with different motion paths.
To implement the change-detection task, each of these 40 trials was presented with either a target-motion change, a distractor-motion change or no change, leading to 120 trials in total. In the 40 target-change trials, one initially highlighted target stopped for 0.5 s at a random time point between 3 and 4.5 s of the motion phase at eccentricities between 5° and 25° of visual angle from the centroid (i.e., the center of mass of all targets), which has been shown to be a common location of gaze for target monitoring (Fehd and Seiffert, 2008, 2010; Vater et al., 2016, 2017a). In the distractor- change trials, all initially highlighted targets of the 40 target-change trials were changed. The stopping object, however, was the same as in the respective target-change trial (see Fig. 1). Eccentricities of the distractor changes were calculated from the new centroid and matched to the eccentricities in the target-change trials now being also between 5° and 25° of visual angle. After a practice block with 12 trials, the 120 test trials were presented in ten test blocks with 12 trials each. In each block, four target-change, four distractor-change and four no-change trials were presented in a randomized order.
Fig. 1.

Example target identification phase at the beginning of a MOT- trial. The four targets to be tracked and recalled are identified with a red frame (here grey). The circled object is the object that (eventually) stops for 0.5 s during the trial. In target-change trials (left image), this object is initially highlighted as a target, whereas in distractor-change trials (right image), this object is not highlighted. Aside from initial target identification, all object trajectories were the same in both conditions.
2.3. Apparatus
The vertical and horizontal rotations of the right eye were captured with a monocular eye-tracking system (EyeSeeCam, ESC, 220 Hz, EyeSeeTec GmbH, Fürstenfeldbruck, Germany) via infrared reflections from the pupil and the cornea (accuracy: 0.5° of the visual angle; resolution: 0.01° RMS within 25° of the field of view). Positional information of retro-reflective markers attached to the ESC were tracked by a 12-camera OptiTrack system (sample rate: 200 Hz) and streamed in real time over Ethernet to a control PC. Additionally, a Wii remote controller (Nintendo, Kyoto, Japan) was connected via Bluetooth to this control PC. A custom software application on the control PC synchronized the three data streams. After this synchronization, it was possible to calculate a three-dimensional gaze vector in the laboratory reference to analyze the gaze position on the screen in relation to the displayed stimuli and the button press in time steps of 5 ms. A large screen (height: 1.87 m; width: 3.01 m) with back projection (InFocus IN 5110 projector; InFocus, Portland, OR) was used to display video stimuli. The rectangular frame for the MOT task covered an area of 1.40 m × 1.40 m (i.e., 40° x 40° of visual angle) in the middle of the screen.
2.4. Procedure
Participants were individually tested in the institute's sensorimotor laboratory in a 1-hr session. After reading the general information about the study, signing the participation agreement and a consent form, the eye-tracking system was fitted to the participant's head. Subsequently, participants were positioned at 2.0 m distance from the screen to read the task instructions. The participants' task was to recall the four targets cued at the beginning of each trial by naming the respective numbers projected onto the objects at the end of each trial. In addition, they had to press a button on a Wii controller with the thumb of their dominant hand as fast as possible when they detected a stop of one of the ten objects. After providing the instructions, the ESC calibration routine was conducted. For the calibration, the participant consecutively fixated on 5 dots creating a two-dimensional axis with an origin, and with dots separated by the distance of 8.5° of visual angle (Kredel et al., 2015). After each of the 10 test blocks with 12 trials each, the EyeSeeCam was recalibrated if the point of gaze deviated more than 1° of visual angle from one of the points of the calibration grid. The numbers of the four recalled targets were recorded in writing by an experimenter at the end of each trial. No feedback of the responses' correctness was given after the trials.
2.5. Measures and analysis
2.5.1. Gaze distance
After the raw gaze data were downsampled to 200 Hz and smoothed with a 41-point, third-order Savitzky–Golay filter, the intersection point between the gaze vector and the screen could be calculated. First, as a manipulation check, the eccentricity (i.e., the distance from the gaze location to the stop-object) was calculated in visual degree for the 40 target- and distractor-change-trials for each individual. For successful manipulation, the average eccentricity should not differ between the target- and the distractor-change condition. Second, to check the predicted centroid-monitoring strategy, gaze distances were calculated to the centroid. Both gaze distances were computed at the time frame of the object-stop onset. Gaze distances to the stop-object will further be compared between detection trials and no-detection trials to examine critical eccentricities for target- und distractor-change detection.
2.5.2. Detection accuracy
To test the two competing hypotheses, the percentage of trials with correctly detected object stops will be compared for the target- and distractor-change condition. A correct detection was counted if the button was pressed after a target or distractor has stopped (false alarms, i.e., trials with a button press but no object stop, were detected in 1.82% of all trials and not included in the analyses).
2.5.3. Detection time
To control for a potential speed-accuracy tradeoff (i.e., higher accuracy but slower detections and vice versa), the detection time was computed as the duration between the onset of the object stop and the onset of the button press.
2.5.4. Recall accuracy
To control if the tracking difficulty in the target- and distractor-stop condition was similar, the percentage of trials in which all four initially highlighted targets could be recalled was computed.
2.5.5. Analysis
All dependent variables were analyzed with repeated-measures ANOVAs. For the eccentricity manipulation check, the ANOVA included the two change locations (target vs. distractor) and the gaze-reference object (stop-object vs. centroid). To test for differences between gaze distances between detected and not-detected motion stops, the ANOVA included the two change locations (target vs. distractor) and decision correctness (correct vs. incorrect). For the analyses of detection accuracy, detection time and recall accuracy the ANOVA only included the two change locations (target vs. distractor). Significant main or interaction effects were further analyzed with Bonferroni-corrected pairwise comparisons. A posteriori effect sizes were computed as partial eta squared, η2p and an alpha level of α = .05 was used to test for any differences. Mathworks MATLAB 2016a was used for the aggregation and analyses of all variables and IBM SPSS Statistics 25 for the statistical analyses.
3. Results
3.1. Gaze distance
3.1.1. Eccentricity manipulation check
As can be seen in Fig. 2, the centroid was closer to the participant's gaze position than the stop-object, which led to a main effect for the factor reference object, F(1,11) = 2592.70, p < .001, ηp2 = 1.00. This pattern did not differ between the target- and distractor-change condition, F(1,11) = 0.52, p = .486, ηp2 = .05, and there was also no interaction between the factors change location and reference object, F(1,11) = 1.29, p = .281, ηp2 = .11.
Fig. 2.

Gaze distance (M, 95% CI) to the stop-object (line) and the centroid (diamond) for the target-stop trials (left) and the distractor-stop trials (right).
3.1.2. Gaze distance for detected and not-detected motion stops
Two Participants could not be included for the analyses because they detected all target stops (mean gaze distance to the targets in these trials was 15.45° and 13.59°, respectively). A main effect for decision correctness was observed, F(1,9) = 21.91, p = .001, ηp2 = .71, indicating that stops were better detected when being closer to the gaze position. This main effect, however, was overruled by an interaction between change location and decision correctness, F(1,9) = 35.42, p < .001, ηp2 = .80. Fig. 3 shows, that gaze-distance differed between detected and not-detected distractor stops (p < .001, ηp2 = .95), while this was not the case for target stops (p = .548, ηp2 = .04). Also, the gaze distance for detected distractor stops was smaller than for detected target stops (p < .001, ηp2 = .85) and gaze distance for not-detected distractor stops was significantly larger than for not-detected target stops (p = .020, ηp2 = .47; see also Fig. 4 for the distribution of gaze distances for detected and not detected stops.).
Fig. 3.

Gaze distance to the stop-object (M, 95% CI) for detected and not-detected target stops (dashed line) and distractor stops (solid line). Significant differences are marked with *, indicating, p < .05 and **, indicating p < .001. Distractor only means that there is a significant gaze-distance difference between detected and not detected distractors but not targets.
Fig. 4.

Gaze distances for each change-detection trial for target changes (black dots) and distractor changes (white dots), for the trials with detection of the stop (left) and for trials where the stop was not detected (right). The x-axis in both figures represents a trial count from the first to the last trial for detected and not-detected target and distractor stops.
3.2. Detection accuracy and time
Detection rates for target stops were higher (88.8 %) than for distractor stops (54.6 %), which revealed a statistically significant difference, F(1,11) = 36.33, p < .001, ηp2 = .77 (Fig. 5, left). There was also a main effect for detection time, F(1,11) = 15.04, p = .003, ηp2 = .58, indicating that target stops were detected faster than distractor stops (Fig. 5, right).
Fig. 5.

Detection accuracy (M, 95% CI) for target and distractor stops (left) and detection time (M, 95% CI) for target and distractor stops (right). Significant differences are marked with *, indicating p < .01 and **, indicating p < .001.
3.3. Recall accuracy
There were no significant differences in recall accuracy between the target- and the distractor-change trials, F(1,11) = 0.27, p = .61, ηp2 = .02, indicating that tracking difficulty was similar (Fig. 6).
4. Discussion
Dual-task situations, such as tracking a number of targets while detecting target changes, places demands on the attentional and the visual system. In the current study, it was tested if the location of selective attention affects the ability to detect motion changes of targets and distractors. Compared to previous studies, retinal eccentricity was measured to take the capabilities of the visual system better into account. The results show that selective attention and retinal eccentricity influence detection rates of target- and distractor-motion changes with peripheral vision differently. While detection rates for target stops are about 30% higher than those for distractor stops, changes of unattended objects are more likely to be missed when the gaze distance to the change object (i.e., the eccentricity) is too large.
These results indicate that covert attention facilitates the accuracy and speed of detecting motion changes in MOT, because target stops were detected in 89 % of the trials while distractor stops were detected only in 55 % of the trials and target stops were detected faster (786 ms) than distractor stops (941 ms, see Fig. 5). Since there were no tracking-difficulty differences between the target- and distractor-change trials (Fig. 6), attentional demands in object monitoring can be ruled out as an alternative explanation. Rather, the location of selective attention might have caused impairments in performance. Since participants preferably locate their selective attention on the targets (Cavanagh and Alvarez, 2005), sustained covert attention seems to boost visual capabilities to detect motion changes in MOT. While visual capabilities such as contrast sensitivity can be enhanced with attention (Carrasco et al., 2006), it has not yet been shown that distributed attention facilitates motion-change detection in a monitoring task. Moreover, the results extend findings by Vater et al. (2017a), who showed that motion changes can be detected with peripheral vision at large eccentricities. When taking the current results into account, it seems that this is only true for the detection of target-motion changes but not for distractor-motion changes. Thus, the motion-sensitivity in peripheral vision is clearly affected by the location of attention.
Fig. 6.
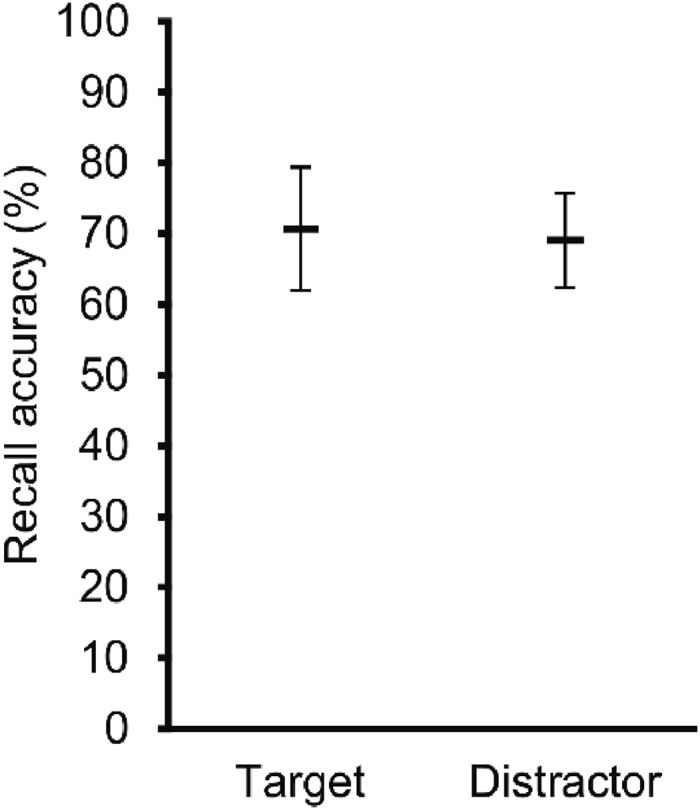
Recall accuracy (M, 95%-CI) for target- and distractor-stop trials.
The results of the current study show a similar pattern as those obtained by Sears and Pylyshyn (2000), who found that form-change detection rates are lower for distractor than for target changes. However, since Sears and Pylyshyn (2000) did not measure gaze eccentricities to the change location, and it is known that peripheral vision is limited in spatial acuity and form vision (Rosenholtz, 2016; Strasburger, 2005; Strasburger et al., 2011; To et al., 2011), their results could alternatively be explained by limitations in the visual system. This alternative explanation can be ruled out for the current results, as eccentricity was measured and there were no eccentricity differences to the change object in target- and distractor trials (Fig. 2). Based on the analyses of eccentricities in trials with and without stop detections, it was found that eccentricities higher than approximately 20° impair the ability to detect distractor-motion changes (Fig. 3). In contrast, eccentricities lower than approximately 15° increase the likelihood to detect distractor changes. Thus, covert attention maybe flexibly located to all objects including distractors when object eccentricity is low (Alvarez and Franconeri, 2007; Cavanagh and Alvarez, 2005) while attentional flexibility seems impaired when distractors are too far in the periphery. Bottom-up attentional mechanisms might be involved for the detection of sudden motion changes of unattended objects at near eccentricities (Cosman and Vecera, 2010; Pratt et al., 2010), but not at far eccentricities. That means, a functional MOT gaze behavior should reduce the eccentricity to objects to a minimum. This could not only help to reduce the previously observed effect of peripheral crowding (Franconeri et al., 2008; Meyerhoff et al., 2016; Vater et al., 2017b) but also for the now observed detection of distractor changes.
When relating results to existing theoretical tracking mechanisms, first, it can be supported that attention seems rather object based than space based as distractor changes should have been detected more often with the latter (Alvarez and Franconeri, 2007; Cavanagh and Alvarez, 2005; Huff and Papenmeier, 2013; Luu and Howe, 2015; Meyerhoff et al., 2017; Papenmeier et al., 2014). It has been suggested that this object-based tracking is characterized by a perceptual grouping of target objects (Huynh et al., 2017; Yantis, 1992). Second, the rather slow motion-change detection times seem to support a mechanism where covert attention is switched between all objects in a serial fashion (Holcombe and Chen, 2013; Howe et al., 2010). The slower detection times in the distractor change condition could indicate that systematic covert-attention searches, on objects other than the targets, are rather slow (Wolfe et al., 2000). While some other studies reported a serial tracking mechanism which was characterized by overt rather than covert tracking (Li et al., 2018, 2019), it could be the case that the additional detection task used in this and previous studies might increase the costs of an overt scanning because an object change might be missed due to the saccadic suppression (Vater et al., 2017a, 2017b). Third, results indicate that distractors are not generally inhibited as was suggested by Bettencourt and Somers (2009). Rather, attentional resources could be flexibly located on all objects, including targets and distractors (Alvarez and Oliva, 2008; Drew et al., 2009; Howe and Ferguson, 2015; Meyerhoff et al., 2015). When referring these findings back to the two predictions, a pure bottom-up mechanism of motion detection in peripheral vision can be ruled out as detection performance was considerably impaired for the detection of distractor stops. Since eccentricity seems to influence the ability to detect distractor changes, future MOT research should investigate if botton-up attentional mechanisms might only be used at small eccentricities.
When referring obtained results back to the corner-kick example from the introduction and the perceptual demands placed on goalkeepers, detecting movements of peripheral players seems to depend on the location of attention and the eccentricity of a moving player. Based on the current results, the chance to detect movements of an unattended player is 34% lower compared with the detection of movements of an attended player. Since peripheral motion detection for unattended objects also occurs later, an appropriate response by the goalkeeper might be initiated too late. Thus, it is important to locate attention on important objects in the periphery. Further, it seems that even unattended object's motion can be detected if the eccentricity is not too large. A potentially efficient gaze and attention strategy could be to have the gaze position between important players but with minimized eccentricities to other players and to then attend to players that are most likely involved in the situation to speed up the movement-detection rates and movement-detection times. While some eye-tracking studies have already shown that athletes actually position their gaze not on a specific information source but between information sources, sometimes even looking at blank space, the direction of attention has yet to be defined in these situations (Vater et al., 2019).
In summary, in tasks that demand distributed attention like MOT and potentially also goalkeeping in soccer, peripheral vision seems to play an important role for object tracking and change detection. The current results show that peripheral vision is naturally used for the detection of motion changes. While changes on targets are detected in 89%, detection rates are only 55 % for distractors although both changes occur on average on the same eccentricity. Retinal eccentricity, nevertheless, is important for change detection as distractor changes could still be detected if the change occurred closer to the fovea. On a theoretical level, results support MOT mechanisms that explain performance with object-based attention, serial covert attention shifts and flexible but limited attentional resources. On a practical level, especially for sports, it seems important to know which objects are relevant for decision making (attention) and where these objects will be located in the environment, to be able to optimally adjust the gaze position and reduce retinal eccentricity (vision). With such a gaze strategy, the use of peripheral vision for detecting motion changes seems to be most functional not only in MOT but presumably also in sports.
Declarations
Author contribution statement
Christian Vater: Conceived and designed the experiments; Performed the experiments; Analyzed and interpreted the data; Contributed reagents, materials, analysis tools or data; Wrote the paper.
Funding statement
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Competing interest statement
The authors declare no conflict of interest.
Additional information
No additional information is available for this paper.
Acknowledgements
I would like to thank Ernst-Joachim Hossner and Ralf Kredel, who helped to develop this experimental paradigm and discuss the results.
References
- Alvarez G.A., Franconeri S.L. How many objects can you track? Evidence for a resource-limited attentive tracking mechanism. J. Vis. 2007;7:14. doi: 10.1167/7.13.14. [DOI] [PubMed] [Google Scholar]
- Alvarez G.A., Oliva A. The representation of simple ensemble visual features outside the focus of attention: research article. Psychol. Sci. 2008;19:392–398. doi: 10.1111/j.1467-9280.2008.02098.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bahrami B. Object property encoding and change blindness in multiple object tracking. Vis. Cogn. 2003;10:949–963. [Google Scholar]
- Bettencourt K.C., Somers D.C. Effects of target enhancement and distractor suppression on multiple object tracking capacity. J. Vis. 2009;9:9. doi: 10.1167/9.7.9. [DOI] [PubMed] [Google Scholar]
- Carrasco M., Giordano A.M., McElree B. Attention speeds processing across eccentricity: feature and conjunction searches. Vis. Res. 2006;46:2028–2040. doi: 10.1016/j.visres.2005.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanagh P., Alvarez G.A. Tracking multiple targets with multifocal attention. Trends Cogn. Sci. 2005;9:349–354. doi: 10.1016/j.tics.2005.05.009. [DOI] [PubMed] [Google Scholar]
- Cosman J.D., Vecera S.P. Attentional capture by motion onsets is modulated by perceptual load. Atten. Percept. Psychophys. 2010;72:2096–2105. doi: 10.3758/bf03196686. [DOI] [PubMed] [Google Scholar]
- Drew T., McCollough A.W., Horowitz T.S., Vogel E.K. Attentional enhancement during multiple-object tracking. Psychon. Bull. Rev. 2009;16:411–417. doi: 10.3758/PBR.16.2.411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faubert J., Sidebottom L. Perceptual-cognitive training of athletes. J. Clin. Sport Psychol. 2012;6:85–102. [Google Scholar]
- Fehd H.M., Seiffert A.E. Eye movements during multiple object tracking: where do participants look? Cognition. 2008;108:201–209. doi: 10.1016/j.cognition.2007.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fehd H.M., Seiffert A.E. Looking at the center of the targets helps multiple object tracking. J. Vis. 2010;10:19. doi: 10.1167/10.4.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franconeri S.L., Jonathan S.V., Scimeca J.M. Tracking multiple objects is limited only by object spacing, not by speed, time, or capacity. Psychol. Sci. 2010;21:920–925. doi: 10.1177/0956797610373935. [DOI] [PubMed] [Google Scholar]
- Franconeri S.L., Lin J.Y., Pylyshyn Z.W., Fisher B., Enns J.T. Evidence against a speed limit in multiple-object tracking. Psychon. Bull. Rev. 2008;15:802–808. doi: 10.3758/pbr.15.4.802. [DOI] [PubMed] [Google Scholar]
- Giordano A.M., McElree B., Carrasco M. On the automaticity and flexibility of covert attention: a speed-accuracy trade-off analysis. J. Vis. 2009;9:30. doi: 10.1167/9.3.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen T., Pracejus L., Gegenfurtner K.R. Color perception in the intermediate periphery of the visual field. J. Vis. 2009;9:26. doi: 10.1167/9.4.26. [DOI] [PubMed] [Google Scholar]
- Holcombe A.O., Chen W.-Y. Splitting attention reduces temporal resolution from 7 Hz for tracking one object to <3 Hz when tracking three. J. Vis. 2013;13:12. doi: 10.1167/13.1.12. [DOI] [PubMed] [Google Scholar]
- Howe P.D.L., Cohen M.A., Pinto Y., Horowitz T.S. Distinguishing between parallel and serial accounts of multiple object tracking. J. Vis. 2010;10:11. doi: 10.1167/10.8.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howe P.D.L., Ferguson A. The identity-location binding problem. Cogn. Sci. 2015;39:1622–1645. doi: 10.1111/cogs.12204. [DOI] [PubMed] [Google Scholar]
- Huff M., Papenmeier F. It is time to integrate: the temporal dynamics of object motion and texture motion integration in multiple object tracking. Vis. Res. 2013;76:25–30. doi: 10.1016/j.visres.2012.10.001. [DOI] [PubMed] [Google Scholar]
- Huff M., Papenmeier F., Zacks J.M. Visual target detection is impaired at event boundaries. Vis. Cogn. 2012;20:848–864. [Google Scholar]
- Huynh D., Tripathy S.P., Bedell H.E., Öğmen H. The reference frame for encoding and retention of motion depends on stimulus set size. Atten. Percept. Psychophys. 2017;79:888–910. doi: 10.3758/s13414-016-1258-5. [DOI] [PubMed] [Google Scholar]
- Kredel R., Klostermann A., Hossner E.-J. Automated vectorbazed gaze analysis for perception-action diagnostics. In: Heinen T., editor. Advances in Visual Perception Research. Nova Science Publisher; New York, NY: 2015. pp. 45–59. [Google Scholar]
- Li J., Oksama L., Hyönä J. Close coupling between eye movements and serial attentional refreshing during multiple-identity tracking. J. Cogn. Psychol. 2018;30:609–626. [Google Scholar]
- Li J., Oksama L., Hyönä J. Model of multiple identity tracking (MOMIT) 2.0: resolving the serial vs. parallel controversy in tracking. Cognition. 2019;182:260–274. doi: 10.1016/j.cognition.2018.10.016. [DOI] [PubMed] [Google Scholar]
- Luu T., Howe P.D.L. Extrapolation occurs in multiple object tracking when eye movements are controlled. Atten. Percept. Psychophys. 2015;77:1919–1929. doi: 10.3758/s13414-015-0891-8. [DOI] [PubMed] [Google Scholar]
- Meyerhoff H.S., Papenmeier F., Huff M. Studying visual attention using the multiple object tracking paradigm: a tutorial review. Atten. Percept. Psychophys. 2017;79:1255–1274. doi: 10.3758/s13414-017-1338-1. [DOI] [PubMed] [Google Scholar]
- Meyerhoff H.S., Papenmeier F., Jahn G., Huff M. Distractor locations influence multiple object tracking beyond interobject spacing: evidence from equidistant distractor displacements. Exp. Psychol. 2015;62:170–180. doi: 10.1027/1618-3169/a000283. [DOI] [PubMed] [Google Scholar]
- Meyerhoff H.S., Papenmeier F., Jahn G., Huff M. Not FLEXible enough: exploring the temporal dynamics of attentional reallocations with the multiple object tracking paradigm. J. Exp. Psychol. Hum. Percept. Perform. 2016;42:776–787. doi: 10.1037/xhp0000187. [DOI] [PubMed] [Google Scholar]
- MATLAB . The MathWorks Inc.; Natick, Massachusetts: 2016. version 9.0.0 (R2016a) [Google Scholar]
- Papenmeier F., Meyerhoff H.S., Jahn G., Huff M. Tracking by location and features: object correspondence across spatiotemporal discontinuities during multiple object tracking. J. Exp. Psychol. Hum. Percept. Perform. 2014;40:159–171. doi: 10.1037/a0033117. [DOI] [PubMed] [Google Scholar]
- Pratt J., Radulescu P.V., Guo R.M., Abrams R.A. It's alive! Animate motion captures visual attention. Psychol. Sci. 2010;21:1724–1730. doi: 10.1177/0956797610387440. [DOI] [PubMed] [Google Scholar]
- Pylyshyn Z.W. The role of location indexes in spatial perception: a sketch of the FINST spatial-index model. Cognition. 1989;32:65–97. doi: 10.1016/0010-0277(89)90014-0. [DOI] [PubMed] [Google Scholar]
- Pylyshyn Z.W. Visual indexes, preconceptual objects, and situated vision. Cognition. 2001;80:127–158. doi: 10.1016/s0010-0277(00)00156-6. [DOI] [PubMed] [Google Scholar]
- Pylyshyn Z.W. Some puzzling findings in multiple object tracking (MOT): II. Inhibition of moving nontargets. Vis. Cogn. 2006;14:175–198. [Google Scholar]
- Pylyshyn Z.W., Haladjian H.H., King C.E., Reilly J.E. Selective nontarget inhibition in multiple object tracking. Vis. Cogn. 2008;16:1011–1021. [Google Scholar]
- Pylyshyn Z.W., Storm R.W. Tracking multiple independent targets: evidence for a parallel tracking mechanism. Spat. Vis. 1988;3:179–197. doi: 10.1163/156856888x00122. [DOI] [PubMed] [Google Scholar]
- Rosenholtz R. Capabilities and limitations of peripheral vision. Annu. Rev. Vis. Sci. 2016;2:437–457. doi: 10.1146/annurev-vision-082114-035733. [DOI] [PubMed] [Google Scholar]
- Sears C.R., Pylyshyn Z.W. Multiple object tracking and attentional processing. Can. J. Exp. Psychol. 2000;54:1–14. doi: 10.1037/h0087326. [DOI] [PubMed] [Google Scholar]
- Strasburger H. Unfocussed spatial attention underlies the crowding effect in indirect form vision. J. Vis. 2005;5:1024–1037. doi: 10.1167/5.11.8. [DOI] [PubMed] [Google Scholar]
- Strasburger H., Rentschler I., Jüttner M. Peripheral vision and pattern recognition: a review. J. Vis. 2011;11 doi: 10.1167/11.5.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- To M.P.S., Regan B.C., Wood D., Mollon J.D. Vision out of the corner of the eye. Vis. Res. 2011;51:203–214. doi: 10.1016/j.visres.2010.11.008. [DOI] [PubMed] [Google Scholar]
- Vater C., Kredel R., Hossner E.-J. Detecting single-target changes in multiple object tracking: the case of peripheral vision. Atten. Percept. Psychophys. 2016;78:1004–1019. doi: 10.3758/s13414-016-1078-7. [DOI] [PubMed] [Google Scholar]
- Vater C., Kredel R., Hossner E.-J. Detecting target changes in multiple object tracking with peripheral vision: more pronounced eccentricity effects for changes in form than in motion. J. Exp. Psychol. Hum. Percept. Perform. 2017;43:903–913. doi: 10.1037/xhp0000376. [DOI] [PubMed] [Google Scholar]
- Vater C., Kredel R., Hossner E.-J. Disentangling vision and attention in multiple-object tracking: how crowding and collisions affect gaze anchoring and dual-task performance. J. Vis. 2017;17:21. doi: 10.1167/17.5.21. [DOI] [PubMed] [Google Scholar]
- Vater C., Kredel R., Hossner E.-J. Examining the functionality of peripheral vision: from fundamental understandings to applied sport science. Curr. Iss. Sport Sci. 2017;2 010. [Google Scholar]
- Vater C., Williams A.M., Hossner E.-J. International Review of Sport and Exercise Psychology. Advance online publication; 2019. What do we see out of the corner of our eye? The role of visual pivots and gaze anchors in sport. [Google Scholar]
- Wolfe J.M., Alvarez G.A., Horowitz T.S. Perception: attention is fast but volition is slow. Nature. 2000;406:691. doi: 10.1038/35021132. [DOI] [PubMed] [Google Scholar]
- Yantis S. Multielement visual tracking: attention and perceptual organization. Cogn. Psychol. 1992;24:295–340. doi: 10.1016/0010-0285(92)90010-y. [DOI] [PubMed] [Google Scholar]
- Zelinsky G.J., Neider M.B. An eye movement analysis of multiple object tracking in a realistic environment. Vis. Cogn. 2008;16:553–566. [Google Scholar]