

ABSTRACT
Parabiosis experiments in mice demonstrated that a young environment could partially rejuvenate multiple tissues of old organisms. However, the circulating mediators responsible of such effect have been elusive so far. Novel results suggest that extracellular vesicles isolated from plasma of young mice increase lifespan in old mice. Here we integrate these findings in a larger framework, advancing the hypothesis that circulating vesicles may mediate the beneficial effect of a young milieu on ageing.
KEYWORDS: Extracellular vesicles, ageing, senescence, SASP, eNAMPT, NAD+, miRNAs, SIRT1, inflammaging, lifespan, physical activity
The desire of eternal youth and immortality pervaded human culture from its inception. Historical, mythological, and religious tales are full of reports on increased fitness or even rejuvenation following specific practices. In particular, blood transfusion, drinking, or replacement have long been suggested as health-promoting behaviours [1]. In the late 50s, parabiosis experiments provided some scientific consistency to these beliefs. Indeed, a shared circulatory system was sufficient to increase bone weight and density of old mice when joined to younger ones [2]. The same experimental design was applied to demonstrate a lifespan-enhancing effect of young blood [3]. Many years later, elegant reports mainly from Rando’s and Wagers’ groups demonstrated a rejuvenation-promoting effect of young blood in a wide variety of cells and tissues, e.g. stem cells, muscle, brain, and the heart [4,5]. However, the pursuit of the circulating factors responsible for such effects did not achieve the same success. In fact, the suggested pro-regeneration role of growth differentiation factor 11, a member of the TGFβ superfamily, has been questioned [6].
Extracellular vesicles (EVs) are membrane-coated nanoparticles actively released by almost all cell types. EVs are usually categorized according to their size. Specifically, small vesicles of a diameter <100 nM derive either from multivesicular bodies (= exosomes), or from the plasma membrane, while larger vesicles derive mainly from the plasma membrane. Increasing evidence indicates that both are able to shuttle and deliver functional proteins and nucleic acids in a paracrine and systemic manner. Blood contains a heterogeneous mixture of EVs of different origins, which are currently being characterized for therapeutic and diagnostic purposes [7,8].
The effects of EVs are now attracting intense interest also in the context of ageing and age-related diseases (ARDs) [9]. In particular, senescent cells (SCs) are emerging as major drivers of ageing and key contributors to inflammaging, the age-associated pro-inflammatory drift that promotes the development of ARDs. Indeed, the genetic or pharmacological removal of SCs is sufficient to prolong lifespan and ameliorate the development of a plethora of ARDs in mice. Notably, these effects are accompanied by a decrease in circulating and tissue levels of pro-inflammatory cytokines and other soluble mediators, known to be the main components of SCs secretome [10]. Recent evidence suggests that EVs are also central constituents of the SCs secretome [11]. In particular, SCs secrete an increased amount of EVs [9], excreting pro-inflammatory DNA and possibly spreading pro-ageing signals [12,13]. In addition, qualitative alterations in EVs cargos underly the development of vascular calcification and osteoporosis, two prototypical ARDs [9]. Conversely, a seminal paper suggests that a 4-month injection of small EVs derived from hypothalamic neural stem cells and rich in specific miRNAs into the hypothalamic third ventricle is sufficient to ameliorate some age-associated detrimental outcomes in C57BL/6 mice, including hypothalamic inflammation and the drop in physical activity [14]. These and other observations prompted the hypothesis that EVs are central mediators of the circulating communicosome fostering inflammaging. In that framework, we hypothesized that the chronic administration of EVs purified from a young healthy mouse to an old one should ameliorate some age-associated phenotypes [15]. This experimental approach appeared to be enough feasible and robust to demonstrate a tangible role of EVs in the ageing process.
Yoshida and colleagues have now shown a clear pro-longevity role for EVs isolated from young mouse plasma [16]. Indeed, they injected EVs isolated from 4-to-12-month-old mice into 26-month-old female mice once a week until sacrifice and observed an increase of 10.2% and of 15.8% in median and maximal lifespan, respectively, in mice receiving the treatment vs. vehicle-treated mice of the same age. Of note, one of the most studied, non-genetic, lifespan-promoting intervention, i.e. pharmacological mTOR inhibition, produced comparable increases in rodent’s lifespan [17]. Also, mice receiving EVs appeared to be healthier and more active. The authors attributed these beneficial effects to the high amount of extracellular nicotinamide phosphoribosyltransferase (eNAMPT) shuttled by EVs from young mice. Indeed, treatment of old mice with EVs isolated from WT adipocytes media phenocopied the physical activity-promoting effect, while EVs derived from Nampt-KO adipocytes did not. Thus, the authors proposed a framework where EVs-associated eNAMPT is internalized into target cells and enhances the intracellular NMN/NAD+ biosynthesis, as elegantly demonstrated in the hypothalamus, hippocampus, pancreas, and retina of adipose tissue-specific Nampt knock-in (ANKI) mice [16]. In turn, the rise in intracellular NAD+ levels would counteract the age-induced decline in metabolic, cognitive, and physical functions, as previously demonstrated for other NAD+-modulating interventions [18].
The observation that also ANKI female mice exhibit a longer median, but not maximal, lifespan does not exclude other putative, beneficial molecules loaded in EVs from young plasma. Among the 181 proteins detectable in EVs by proteomic analysis, 3% showed age-related trends. However, EVs shuttle almost all existing biological molecules, including miRNAs [8]. We recently characterized the miRNAs repertoire shuttled in EVs secreted by non-senescent and senescent cells identifying a senescence–associated miRNAs signature targeting SIRT1 [15, unpublished data]. Since SIRT1 is one of the major downstream targets of altered NAD+ concentration [18], overall data hint at the existence of an EVs-mediated, synergistic protein/miRNAs-based modulation of SIRT1 function and expression (Figure 1). Of note, the age-related decline in the EVs content of eNAMPT observed in mice was mirrored in plasma of subjects of different ages, suggesting the relevance of this phenomenon for human ageing [16]. These results reinforce the central role of NAD+ metabolism and SIRT1 in the ageing process [18]. On the other side, the authors highlight complex alterations in circulating EVs payload during ageing.
Figure 1.
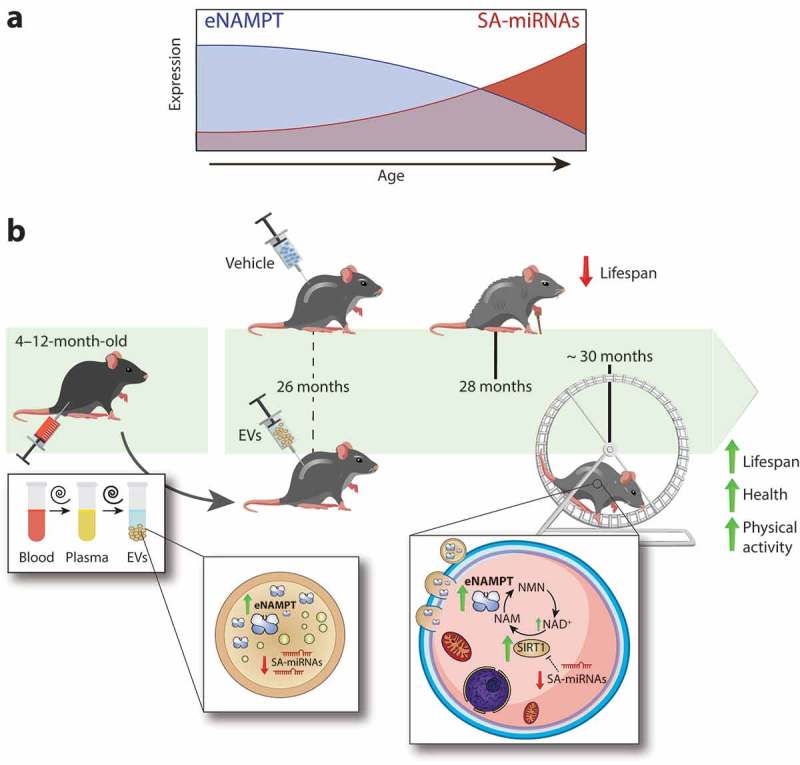
Chronic treatment with extracellular vesicles of young mice prolongs lifespan in old mice. (a) The amount of extracellular nicotinamide phosphoribosyltransferase (eNAMPT) within extracellular vesicles (EVs) decreases during ageing, while the content of senescence-associated (SA)-miRNAs increases progressively. (b) EVs isolated from plasma of young mice contain high levels of eNAMPT and low levels of SA-miRNAs. EVs from young mice injected intraperitoneally into old mice once-a-week produce a tangible increase in lifespan coupled by an enhanced physical activity and a healthier aspect [16]. Shuttled eNAMPT is functionally active in receiving tissues, thus increasing NAD+ levels and pushing SIRT1 activation. Low-levels of SA-miRNAs promote a higher SIRT1 expression [15], implying that young EVs may concertedly sustain SIRT1-related pathways, ultimately promoting healthy longevity.
The field of EVs is still in its infant phase and major questions are still open regarding the relative contribution of various tissues to the plasmatic EVs pool and the effective disease-modifying potential of EVs alterations [19]. For instance, a seminal paper suggested that adipose tissue is a major contributor to the circulating pool of miRNAs shuttled by EVs [20]. However, other studies suggest that platelets, leukocytes, and the endothelium are major contributor to the plasmatic EVs pool [21]. Also, acquired knowledge regarding EVs content has been challenged by the observation that EVs have a substantially more restricted repertoire of molecules than what previously thought [22]. In the lifespan experiments, the authors used a commercially available isolation kit to achieve the highest yields of EVs [16], which may have influenced the composition of the preparation [7]. Replication studies with other separation methods would boost the validity of these findings. Broadly, these new results should prompt more research to extensively characterize EVs alterations during ageing, in order to disentangle the effective impact of these mediators on the ageing milieu.
Transfusion of plasma from young donors to old patients with Alzheimer’s disease has reached the clinical stage, with already available promising results [23]. Considering the low immunogenic potential of EVs [8], it is tempting to speculate that the suggested beneficial effects of young blood may be recapitulated by EVs transfusion, a hypothesis deserving exploration in the future. In the meanwhile, these new findings suggest that the untraceable Fountain of Youth could be “hidden” within EVs circulating in young organisms.
Funding Statement
This work was supported in part by a grant from La Maratò de TV3 to AC and by the Italian Ministry of Health [Ricerca Corrente];Fundació la Marató de TV3;Ministero della Salute [Ricerca Corrente].
Author contributions
FP, FO and AC conceived the idea and wrote the manuscript. AG, JS, EM, VdN, PdC, and LLS collected relevant literature, prepared the figure, and revised the manuscript for intellectual content. The final version of the manuscript was approved by all authors.
Declaration of interest
None of the authors have competing interests.
References
- [1].Learoyd P. The history of blood transfusion prior to the 20th century–part 1. Transfus Med. 2012;22:308–4. [DOI] [PubMed] [Google Scholar]
- [2].Horrington EM, Pope F, Lunsford W, et al. Age changes in the bones, blood pressure, and diseases of rats in parabiosis. Gerontologia. 1960;4:21–31. [DOI] [PubMed] [Google Scholar]
- [3].Ludwig FC, Elashoff RM. Mortality in syngeneic rat parabionts of different chronological age. Trans N Y Acad Sci. 1972;34:582–587. [DOI] [PubMed] [Google Scholar]
- [4].Conboy IM, Conboy MJ, Wagers AJ, et al. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature. 2005. February 17;433(7027):760–764. [DOI] [PubMed] [Google Scholar]
- [5].Hofmann B. Young blood rejuvenates old bodies: a call for reflection when moving from mice to men. Transfus Med Hemother. 2018. January;45(1):67–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Brun CE, Rudnicki MA. GDF11 and the mythical fountain of youth. Cell Metab. 2015;22:54–56. [DOI] [PubMed] [Google Scholar]
- [7].Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018. November 23;7(1):1535750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19:213–228. [DOI] [PubMed] [Google Scholar]
- [9].Takasugi M. Emerging roles of extracellular vesicles in cellular senescence and aging. Aging Cell. 2018;17:e12734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci. 2014. June;69(Suppl 1):S4–S9. [DOI] [PubMed] [Google Scholar]
- [11].Terlecki-Zaniewicz L, Lämmermann I, Latreille J, et al. Small extracellular vesicles and their miRNA cargo are anti-apoptotic members of the senescence-associated secretory phenotype. Aging (Albany NY). 2018. May 19;10(5):1103–1132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Takasugi M, Okada R, Takahashi A, et al. Small extracellular vesicles secreted from senescent cells promote cancer cell proliferation through EphA2. Nat Commun. 2017. June 6;8: 15729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Takahashi A, Okada R, Nagao K, et al. Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat Commun. 2017;8:15287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Zhang Y, Kim MS, Jia B, et al. Hypothalamic stem cells control ageing speed partly through exosomal miRNAs. Nature. 2017;548:52–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Prattichizzo F, Micolucci L, Cricca M, et al. Exosome-based immunomodulation during aging: A nano-perspective on inflamm-aging. Mech Ageing Dev. 2017;168:44–53. [DOI] [PubMed] [Google Scholar]
- [16].Yoshida M, Satoh A, Lin JB, et al. Extracellular vesicle-contained eNAMPT delays aging and extends lifespan in mice. Cell Metab. 2019;30:329–342.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Harrison DE, Strong R, Sharp ZD, et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009;460:392–395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Rajman L, Chwalek K, Sinclair DA. Therapeutic potential of NAD-boosting molecules: the in vivo evidence. Cell Metab. 2018;27:529–547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Shah R, Patel T, Freedman JE. Circulating extracellular vesicles in human disease. N Engl J Med. 2018. September 6;379(10):958–966. [DOI] [PubMed] [Google Scholar]
- [20].Thomou T, Mori MA, Dreyfuss JM, et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature. 2017. February 23;542(7642):450–455. Epub 2017 Feb 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Brahmer A, Neuberger E, Esch-Heisser L, et al. Platelets, endothelial cells and leukocytes contribute to the exercise-triggered release of extracellular vesicles into the circulation. J Extracell Vesicles. 2019. May 28;8(1):1615820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Jeppesen DK, Fenix AM, Franklin JL, et al. Reassessment of exosome composition. Cell. 2019. April 4;177(2):428–445.e18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Sha SJ, Deutsch GK, Tian L, et al. Safety, tolerability, and feasibility of young plasma infusion in the plasma for alzheimer symptom amelioration study: a randomized clinical trial. JAMA Neurol. 2019;76:35–40. [DOI] [PMC free article] [PubMed] [Google Scholar]