

Abstract Abstract
Sabah, Malaysia, is well known for its extensive and diverse coral reefs. It is located on the northwestern edge of the Coral Triangle, the region with the highest marine biodiversity. Much of the marine fauna here is still unknown, especially inconspicuous animals, such as small stoloniferous octocorals, which are common on coral reefs. Here, we describe two new monospecific genera of the family Arulidae found off the coast of Kota Kinabalu, Sabah, East Malaysia; Bunga payunggen. nov. et sp. nov. and Laeta waheedaegen. nov. et sp. nov. As well, the stoloniferan genus Phenganax Alderslade & McFadden, 2011 belonging to the family Clavulariidae is expanded with three new species, P. marumisp. nov., P. subtilissp. nov., and P. stokvisisp. nov., which are all sclerite-free. Additionally, we report a possibly undescribed species, closely related to the clavulariid genera Azoriella Lopez-Gonzalez & Gili, 2001 and Cervera Lopez-Gonzalez et al., 1995. As this and other recent studies have shown, discoveries of small stoloniferous octocorals are helping to fill gaps in our knowledge of the overall systematics of Octocorallia.
Keywords: 28S rDNA, Arulidae , biodiversity, Clavulariidae , COI, Coral Triangle, mtMutS, ND6, Stolonifera , systematics, TARP, taxonomy
Introduction
Coral reefs fringe one-sixth of the world’s coastlines and constitute the most biologically diverse shallow-water marine ecosystems, supporting thousands of species (Birkeland 1997; Reaka-Kudla 1997; Roberts et al. 2002). The estimated global biodiversity of coral reefs is approximately 950,000 (±40%) species, of which 90% are not yet described (Fisher et al. 2015). One reason marine biodiversity is globally underestimated is that there are many small and cryptic species that are usually overlooked in diversity assessments (Wolf et al. 1983; Hoeksema 2017; Lau et al. 2018, 2019; Sabroux et al. 2019). An estimate from 2002 indicated that 25% of the world’s coral reefs have already been severely damaged due to global warming (Goreau et al. 2000; Roberts et al. 2002), pollution, and destructive fisheries methods (dynamite and poison) (Polunin and Roberts 1996; Roberts et al. 2002; Hughes et al. 2017; Heery et al. 2018). Damage continues to accumulate (Hughes et al. 2018), thus threatening coral reef biodiversity and possibly causing the extinction of organisms that have not been scientifically described yet (Carpenter et al. 2008; Hoeksema 2017).
The highest concentrations of coral reef species can be found in the Coral Triangle in the Central Indo-Pacific (Hoeksema 2007, 2017). Continuing analyses have resulted in the boundaries of the Coral Triangle biodiversity hotspot being refined over time (Hoeksema 2007; Huang et al. 2015; Veron et al. 2015; Lane and Hoeksema 2016). The coastline of Sabah, on the northern tip of East Malaysia, is located within the area that makes up the outer western edge of the Coral Triangle (Hoeksema 2007; Waheed and Hoeksema 2013, 2014; Waheed et al. 2015a). Malaysia is known for its extensive coral reefs that cover an area of approximately 4000 km2 with the majority (>75%) situated in Sabah (Burke et al. 2002; Waheed et al. 2007). Sabah (with its offshore islands) has an extensive history of marine research, focusing mainly on scleractinian diversity and coral cover (Lulofs 1973; Lulofs et al. 1974; Wood 1977, 1979; Mathias and Langham 1978; Nyanti and Johnston 1992; Pilcher and Cabanban 2000; Waheed and Hoeksema 2013, 2014; Reef Check Malaysia 2014, Waheed et al. 2015a, 2015b), and reef fish (Kassem et al. 2012; Reef Check Malaysia 2014; Townsend 2015).
Just off the coast of Kota Kinabalu, the capital of Sabah, there is an assemblage of five islands that make up Tunku Abdul Rahman Park (TARP): Gaya, Manukan, Sapi, Sulug, and Mamutik islands cover an area of approximately 50 km2, including their surrounding reefs and sea (Spait 2001; Reef Check Malaysia 2014; Waheed and Hoeksema 2014). TARP has been protected since 1977 and is reserved and managed by Sabah Parks. Despite the protection of the coral reefs in the park against destructive fishing and land-based developments (Spait 2001; Waheed et al. 2007), the coral cover in the park has seen a severe decline since 1994 owing to continued anthropogenic influences and a major crown-of-thorns starfish outbreak (Praveena et al. 2012; Waheed and Hoeksema 2014; Heery et al. 2018), emphasizing the urgent need for diversity research and taxonomic documentation.
For the shallow waters of the Indo-Pacific, over 100 genera in 23 families of Alcyonacea (e.g., Alderslade 2001, 2002; Fabricius and Alderslade 2001; Benayahu et al. 2004; van Ofwegen 2005), nine genera in five families of Pennatulacea (Williams 1996), and two genera in two families of Helioporacea have been described (Fabricius et al. 2007; Miyazaki and Reimer 2015). However, compared to scleractinian corals, octocorals have received little research attention and are less well documented (Fabricius and Alderslade 2001; Breedy and Cortés 2008; McFadden et al. 2010; Samimi-Namin and Van Ofwegen 2012). Despite Sabah’s location within the area that makes up the outer western edge of the Coral Triangle (Hoeksema 2007; Waheed and Hoeksema 2014; Waheed et al. 2015a), and the fact that octocorals are an abundant and species-rich group on Indo-Pacific coral reefs, relatively little research has been done on octocorals in Sabah (Kassem et al. 2012).
Zooxanthellate octocorals are, however, similarly affected by global climate change and other anthropogenic and natural threats (Prada et al. 2010; Dias et al. 2016; Van de Water et al. 2018). Additionally, octocorals are one of the most widely distributed and common benthic groups, occurring from shallow tropics to the Antarctic deep sea, and are important members of the benthic community, providing refuge and habitat for numerous organisms (Sánchez et al. 2003; Sánchez 2016). Due to their abundance, diverse structural complexity, and symbiotic relationships, octocorals play an important role in the energy transfer between plankton and other benthos (Van de Water et al. 2018) and should receive more research attention. However, research focused on octocorals is still uncommon, and the importance of their taxonomy is underestimated (McFadden and Ofwegen 2012; Benayahu et al. 2017; Conti-Jerpe and Freshwater 2017). This is particularly true for one group of octocorals, namely species of the subordinal group Stolonifera, many of which are inconspicuous, with small polyps (often ~2–3 mm in diameter) and colonies, and are hard to find, one reason for the scant amount of information available for this group (Wolf et al. 1983; McFadden and Ofwegen 2012). In the TARP area, next to sponges, algae, and hard corals, soft corals are the most dominant benthic fauna (Reef Check Malaysia 2014), however, research focusing on octocorals is completely lacking here. Therefore, there may be undescribed species, especially given the geographical location of TARP on the edge of the Coral Triangle.
The present study is the first investigation into the stoloniferous octocorals in and around the TARP area, off the coast of Kota Kinabalu, Sabah, Malaysia, and aims to improve on our understanding of the phylogenetic relationships of stoloniferous octocorals within the Octocorallia radiation. Based on our findings, we formally describe two new monotypic genera within Arulidae and three new species within the genus Phenganax Alderslade & McFadden, 2011 within Clavulariidae.
Materials and methods
Specimen collection
A total of 25 stoloniferous octocoral specimens (Table 1) was collected from eight locations (Figure 1) in and around TARP, off the coast of Kota Kinabalu, Sabah, Malaysia, between 20 and 27 March 2018. Specimens were collected at depths of 4–18 m by means of SCUBA and all voucher material was preserved in 70–80% ethanol and sub-samples in 95% ethanol. High-resolution in situ images were taken with an Olympus Tough TG-4 in an Olympus PT-056 underwater housing. Vouchers and type material have been deposited at the National Museum of Nature and Science (NSMT), Tokyo, Japan (all holotypes) and at the Borneo Marine Research Institute (IPMB), Universiti Malaysia Sabah (UMS), Sabah, Malaysia (all paratypes).
Table 1.
Overview of stoloniferous octocoral specimens collected from off NW Sabah, used in this study; including GenBank accession numbers and locality. Key: catalogue number: NSMT = National Museum of Nature and Science, Tokyo, Japan; IPMB = Borneo Marine Research Institute, Sabah, Malaysia; n.a. = not available.
| Family | Species | Catalogue/voucher number | Location | GPS (DMS) | GenBank accession numbers | |||
|---|---|---|---|---|---|---|---|---|
| 28S rDNA | COI | mtMutS | ND6 | |||||
| Arulidae | Bunga payung gen. nov. et sp. nov. | IPMB-C 01.00017 | Sepangar, E Sepangar Is. | 06°03'38.66"N, 116°04'0.65"E | MN164539 | n.a. | n.a. | MN164587 |
| B. payung gen. nov. et sp. nov. (holotype) | NSMT-Co 1679 | Sepangar, E Sepangar Is. | 06°03'38.66"N, 116°04'0.65"E | MN164540 | MN164559 | n.a. | MN164588 | |
| Laeta waheedae gen. nov. et sp. nov. | IPMB-C 01.00018 | Mid Reef, E Manukan Is. | 05°58'35.8"N, 116°00'52.2"E | MN164542 | MN164561 | MN164583 | MN164590 | |
| IPMB-C 01.00019 | Gaya Clement Reef, E Gaya Is. | 06°01'24.26"N, 116°00'13.55"E | MN164562 | MN164584 | MN164591 | |||
| L. waheedae gen. nov. et sp. nov. (holotype) | NSMT-Co 1680 | Udar, W Udar is. | 06°4'49.81"N, 116°5'13.16"E | MN164541 | MN164560 | MN164582 | MN164589 | |
| Clavulariidae | Clavulariidae sp. | NSMT-Co 1686 | Edgell Patches, W Sapi Is. | 06°00'38.7"N, 115°59'22.2"E | MN164543 | MN164563 | MN164580 | n.a. |
| IPMB-C 01.00016 | Gaya Clement Reef, W Gaya Is. | 06°01'24.26"N, 116°00'13.55"E | MN164544 | MN164564 | MN164581 | n.a. | ||
| Phenganax marumi sp. nov. | IPMB-C 01.00001 | Edgell Patches, W Sapi Is. | 06°00'38.7"N, 115°59'22.2"E | MN164545 | MN164570 | n.a. | MN164595 | |
| IPMB-C 01.00002 | Mid Reef Slope, E Manukan Is. | 05°58'38.08"N, 116°00'52.82"E | MN164546 | MN164571 | n.a. | MN164596 | ||
| IPMB-C 01.00003 | Mid Reef Slope, E Manukan Is. | 05°58'38.08"N, 116°00'52.82"E | n.a. | MN164572 | n.a. | MN164597 | ||
| IPMB-C 01.00004 | Edgell Patches, W Sapi Is. | 06°00'38.7"N, 115°59'22.2"E | MN164547 | MN164573 | n.a. | MN164598 | ||
| IPMB-C 01.00005 | Edgell Patches, W Sapi Is. | 06°00'38.7"N, 115°59'22.2"E | MN164548 | MN164574 | n.a. | MN164599 | ||
| IPMB-C 01.00006 | Gaya Clement Reef, W Gaya Is. | 06°01'24.26"N, 116°00'13.55"E | MN164549 | MN164575 | n.a. | MN164600 | ||
| IPMB-C 01.00007 | Gaya Clement Reef, W Gaya Is. | 06°01'24.26"N, 116°00'13.55"E | n.a. | MN164576 | n.a. | MN164601 | ||
| IPMB-C 01.00008 | Manukan, N Manukan Is. | 05°58'46.1"N, 116°00'10.6"E | MN164550 | MN164577 | n.a. | MN164602 | ||
| P. marumi sp. nov. (holotype) | NSMT-Co 1683 | Manukan, N Manukan Is. | 05°58'46.1"N, 116°00'10.6"E | MN164551 | n.a. | n.a. | MN164603 | |
| P. marumi sp. nov. | IPMB-C 01.00009 | Manukan, N Manukan Is. | 05°58'46.1"N, 116°00'10.6"E | MN164552 | MN164578 | n.a. | MN164604 | |
| IPMB-C 01.00010 | Manukan, N Manukan Is. | 05°58'46.1"N, 116°00'10.6"E | MN164553 | MN164579 | n.a. | MN164605 | ||
| Phenganax subtilis sp. nov. (holotype) | NSMT-Co 1684 | Sepangar, W Sepangar Is. | 06°03'38.66"N, 116°04'0.65"E | MN164554 | MN164566 | MN164586 | n.a. | |
| P. subtilis sp. nov. | IPMB-C 01.00011 | Sepangar II, W Sepangar Is. | 06°04'7.38"N, 116°04'6.76"E | MN164555 | MN164567 | n.a. | n.a. | |
| IPMB-C 01.00013 | Manukan, N Manukan Is. | 05°58'46.1"N, 116°00'10.6"E | n.a. | MN164568 | n.a. | n.a. | ||
| IPMB-C 01.00012 | Manukan, N Manukan Is. | 05°58'46.1"N, 116°00'10.6"E | n.a. | MN164569 | n.a. | n.a. | ||
| Phenganax stokvisi sp. nov. (holotype) | NSMT-Co 1685 | Mid Reef, E Manukan Is. | 05°58'35.8"N, 116°00'52.2"E | MN164556 | n.a. | n.a. | MN164592 | |
| P. stokvisi sp. nov. | IPMB-C 01.00015 | Mid Reef, E Manukan Is. | 05°58'35.8"N, 116°00'52.2"E | MN164557 | n.a. | n.a. | MN164593 | |
| IPMB-C 01.00014 | Mid Reef, E Manukan Is. | 05°58'35.8"N, 116°00'52.2"E | MN164558 | MN164565 | MN164585 | MN164594 | ||
Figure 1.

Map of eight sampling locations where stoloniferous octocorals were collected in this study, around Udar and Sepangar Islands and within Tunku Abdul Rahman Park (including Gaya, Sapi, Manukan, Mamutik, and Sulug Islands), Kota Kinabalu, Sabah, Malaysia.
Morphological examinations
Sclerites were isolated by dissolving different parts of the specimens (polyp tentacle, calyx, entire polyp and stolon) in 4% hypochlorite (household bleach). Rinsing and visualisation of sclerites followed the same protocol as described in Lau et al. (2018, 2019). Additionally, sclerites from specimens that needed more detailed examination were mounted on scanning electron microscope stubs and coated with Pd/Au for imaging on a JEOL JSM6490LV scanning electron microscope (SEM) operated at high vacuum at 15 kV.
DNA extraction, amplification, and sequencing
DNA was extracted from polyps using a DNeasy Blood and Tissue kit (Qiagen, Tokyo). PCR amplification and sequencing of three mitochondrial markers, cytochrome c oxidase subunit I (COI), mitochondrial mutS-like protein (mtMutS), and the ND6 subunit (ND6), and one nuclear ribosomal marker, 28S ribosomal DNA (28S rDNA) followed the protocols in Lau et al. (2019). The amplified products were visualised by 1% agarose gel electrophoresis. PCR products were treated with Exonuclease I and Alkaline Phosphate (Shrimp) and sent for bidirectional sequencing on an ABI 3730XL (Fasmac, Kanagawa, Japan). Sequences were assembled and edited using Geneious R11 (Kearse et al. 2012) and BioEdit (Hall 1999). COI, mtMutS, and ND6 were checked for introns, exons, and stop-codons in AliView (Larsson 2014).
Molecular phylogenetic analyses
Multiple sequence alignments were performed using MAFFT 7 (Katoh and Standley 2013) and coding markers were aligned using MACSE (Ranwez et al. 2011) under default parameters. To determine the phylogenetic position of the collected specimens, consensus sequences for markers 28S rDNA, COI and mtMutS were aligned to a reference dataset of 121 octocoral genera (n = 127 sequences), including sequences of Cornularia pabloi McFadden & van Ofwegen, 2012 and Cornularia cornucopiae (Pallas, 1766) as outgroup (total n = 134 sequences). Alignments of 893 bp for 28S rDNA, 717 bp for COI and 923 bp for mtMutS were separately run in Maximum Likelihood (ML) analyses (Suppl. material 1: Figures S1–S3), which were highly congruent with the phylogenetic construction generated with the concatenated three-marker dataset (2533 bp).
A separate phylogenetic analysis was made using a concatenated four-marker dataset to further investigate the phylogenetic position of collected clavulariid and arulid specimens. The four separate markers (28S rDNA, 804 bp; COI, 717 bp; mtMutS, 734 bp; ND6, 441 bp) were also run in ML analyses to check for congruency (Suppl. material 1: Figures S4–S7). The concatenated markers resulted in a 2696 bp dataset with a total of nine reference taxa, including Hanabira yukibana Lau, Stokvis, Imahara & Reimer, 2019 as outgroup (total n = 33).
Alignments of the separate markers were all concatenated using SequenceMatrix 1.8 (Gaurav et al. 2011). ML analyses were run with RAX-ML 8 (Stamatakis 2014) using the GTRCAT model. The best ML tree was calculated using the –D parameter. A multi-parametric bootstrap search was performed, which automatically stopped based on the extended majority rule criterion. The Bayesian inference was performed with ExaBayes 1.5 (Aberer et al. 2014) using the GTR substitution model. Four independent runs were run for 10,000,000 generations during which convergence (with a standard deviation of split frequencies <2%) had been reached. Bootstrap supports and posterior probabilities were depicted on the branches of the best ML tree using P4 (Foster 2004). The resulting tree was visualized in FigTree 1.4.2 (Rambout 2014). Additionally, average distance estimations within species and within genera were computed using MEGA X (Kumar et al. 2018) by analysing pairwise measures of genetic distances (uncorrected P) among sequences (Suppl. material 1: Tables S1–S6).
Systematic account
Class Anthozoa
Subclass Octocorallia Ehrenberg, 1831
Order Alcyonacea Lamouroux, 1812
Family Arulidae McFadden & Ofwegen, 2012
Genus. Bunga gen. nov.
7B9D9009A0D150828F13299B08B1C31F
http://zoobank.org/BFF20AFD-A854-48EB-9F6A-EB3AC2066A95
Type species.
Bunga payung sp. nov., by original designation and monotypy.
Diagnosis.
Colony with polyps connected through thin stolons, which are cylindrical in cross-section and loosely attached to hard substrate. Anthocodiae retract into clavate calyces, which do not retract into the stolon. Oral disk expanded to circular membrane, as is characteristic of arulids. Oral disk with eight shallow furrows running from intertentacular margin to mouth of polyp, dividing membrane into eight lobes. Distal two-thirds of tentacles extend from fused margins of oral membrane. Sclerites of anthocodiae are rods. Sclerites of calyx are table-radiates. Sclerites of stolon are fused table-radiates forming a sheet. Sclerites colourless. Zooxanthellate.
Remarks.
The main difference between type species Bunga payung gen. nov. et sp. nov., with Arula McFadden & Ofwegen, 2012 and HanaLau et al., 2018 is found in both polyp morphology and sclerites; the outer margins of the oral membrane are much less pronounced than seen in Arula and Hana, as well as the lobes, as the furrows appear to be shallower. The 6-radiate sclerite type could not be found in any of the Bunga gen. nov. specimens, which are present in the polyp calyx of both Arula and Hana. Bunga, similar to Hana, has fused table-radiates that form a sheet in the stolon.
Etymology.
From the Malaysian and Indonesian word bunga, meaning flower; denoting the shape of the polyps, which resemble flowers. Gender: feminine.
Bunga payung sp. nov.
A7BFFA7C748755D79DECEC7906461195
http://zoobank.org/5DDA8DCE-2333-4C45-8FA4-77021E54A211
Figure 2.

Bunga payung gen. nov. et sp. nov.: a In situ photograph, holotype (NSMT-Co 1679) b In situ photograph, paratype (IPMB-C 01.00017) c holotype in ethanol. Laeta waheedae gen. nov. et sp. nov. d holotype (NSMT-Co 1680) in ethanol e In situ photograph, holotype f In situ photograph, paratype (IPMB-C 01.00019). Scale bars: 1 mm.
Figure 4.

Sclerite types observed in Bunga payung gen. nov. et sp. nov., NSMT-Co 1679, holotype: a Anthocodial rods b table-radiates of calyx, top view c table-radiates of calyx, bottom view d table-radiates of calyx, lateral view e fragment of fused table-radiates of the stolon. Scale bars: 0.05 mm (a–d); 0.1 mm (e).
Material examined.
All specimens are from Sepangar, Sepangar Island, Kota Kinabalu, Sabah, Malaysia (06°03'38.66"N, 116°04'0.65"E), 20 March 2018 and collected by YW Lau. Holotype: NSMT-Co 1679, 9 m depth. Paratype: IPMB-C 01.00017, 10 m depth.
Description.
Colony with numerous polyps (total ~70). Polyps connected through stolons attached to rock. Stolons are thin and rounded (circular in cross-section, ~0.3 mm in diameter) and polyps are spaced apart irregularly, either adjacent to one another or spaced apart up to ~5 mm. Expanded polyps are ~2.2–3.0 mm in width and retract fully into calyces of ~1 mm wide and up to ~2 mm in height. Calyces do not retract into the stolon. The oral disk of the polyps is expanded into a circular membrane by fusion of proximal regions of adjacent tentacles (Figure 2a), as is characteristic of arulids. The oral disk has eight shallow furrows that run from intertentacular margin to mouth of polyp, dividing the membrane into eight lobes. The distal two-thirds of the tentacles extend from fused margins of the oral membrane. Tentacles with 6–10 pairs of widely spaced pinnules are arranged in a single row on either side of rachis.
Anthocodial sclerites are smooth rods, with simple tubercles at distal margin ends, 0.1–0.15 mm long (Figure 4a). Calyces contain table-radiates that range 0.06–0.19 mm in length (Figure 4b–d). Sclerites of the stolon are fused table-radiates forming a flat sheet (Figure 4e).
Polyps are brown coloured in life with a whitish oral disk, but yellowish white when preserved in ethanol. Zooxanthellate.
Morphological variation.
The paratype is a colony consisting of ~10 polyps. Polyps of the paratype colony show variation in colouration; the whitish colour is not restricted to the oral disk but is also seen in the tentacles. This could be due to differences in sclerite density (Figure 2a), as described in Alderslade and McFadden (2007) and Lau et al. (2019), or it could be due to the position of zooxanthellae in the tissue.
Distribution.
Sepangar Island, Kota Kinabalu, Sabah, Malaysia.
Remarks.
Anthocodial rods are scarce in Bunga payung gen. nov. et sp. nov. The 6-radiate type of sclerite was not observed in this genus but is present in Arula and Hana, the two other genera in the family Arulidae.
Etymology.
From the Malaysian and Indonesian word payung, which means umbrella; denoting the shape of the oral disc of the polyps, which resemble the shape of an umbrella.
Genus. Laeta gen. nov.
34C1E76EFA0854E1ADF3CA66CF0B00A9
http://zoobank.org/02B40F3A-AA78-49A5-ADA8-12F3374CDF21
Type species.
Laeta waheedae sp. nov., by original designation and monotypy.
Diagnosis.
Colony with polyps connected through stolons, which are ribbon-like. Oral disk with eight furrows, running from intertentacular margin to the mouth of the polyp, dividing the membrane into eight shallow lobes. Distal two-thirds of tentacles extend from fused margins of the oral membrane. Sclerites of anthocodiae are rods. Sclerites of calyx are table-radiates and three types of rods; (1) branched rods with high tuberculate processes, (2) club-like rods and (3) table-radiate-like rods. Sclerites of stolon are fused table-radiates, which form a sheet. Sclerites colourless. Zooxanthellate.
Remarks.
The main morphological difference between the type species, Laeta waheedae gen. nov. et sp. nov., and species of the genera Arula, Hana, and Bunga gen. nov. is found in the presence of all three types of rods in its calyces. The club-like rod with tubercles at one distal end of the rod is also seen in Hana hanatabaLau et al., 2018, however this type of sclerite was only found in the anthocodiae in Laeta waheedae and not in the calyx as in Hana. Similar to Bunga, Laeta does not have 6-radiates in the calyx. Similar to Hana and Bunga, Laeta has fused table-radiates in the stolon. Outer margins of the oral membrane are much less pronounced than in Arula and Hana, and are more similar to margins of the oral disk in Bunga.
Etymology.
From Latin laeta, meaning bright, charming, cheerful. Gender: feminine.
Laeta waheedae sp. nov.
289F7404F31D5F01A1C9F9599E3EF929
http://zoobank.org/761E7919-D311-4F0B-8B67-17E8ECDEC370
Figure 5.

Sclerite types observed in Laeta waheedae gen. nov. et sp. nov., NSMT-Co 1680, holotype: a rods of calyx b rods of calyx c anthocodial rods d table-radiates of calyx e table-radiates of calyx, top view f rods of calyx g table-radiates of calyx, lateral view h fragment of fused table-radiates of the stolon. Scale bars 0.05 mm (a–c, f); 0.02 mm (d–e); 0.01 mm (g); 0.02 mm (h).
Material examined.
All specimens are from Kota Kinabalu, Sabah, Malaysia and collected by YW Lau. Holotype: NSMT-Co 1680, Udar, east of Udar Island (06°4'49.81"N, 116°5'13.16"E), 10 m depth. Paratypes: IPMB-C 01.00018, Mid Reef, east of Manukan Island, TARP (05°58'46.1"N, 116°00'10.6"E), 8 m depth. IPMB-C 01.00019, Gaya, Gaya Island, TARP (06°01'24.26"N, 116°00'13.55"E), 11 m depth.
Description.
The colony consists of ~50 polyps, which are connected through ribbon-like stolons (0.5–0.6 mm width). Polyps are spaced apart quite regularly (~3 mm). Anthocodiae retract fully into low oval to cylindrical calyces (~1 mm in width, ~0.5 mm in height), which do not retract into the stolon. Expanded polyps 3.5–4.0 mm in diameter. Oral membrane with eight shallow furrows running from intertentacular margin to mouth of polyp, dividing the membrane into eight shallow lobes. Distal two-thirds of tentacles extend from fused margins of the oral membrane. Tentacles with 6–8 pairs of widely spaced pinnules are arranged in a single row on either side of rachis.
Sclerites of anthocodiae are small smooth rods, 0.035–0.1 mm in length, with little ornamentation (Figure 5c). Sclerites of calyx are table-radiates, 0.06–0.10 mm (Figure 5d, e, g) and three types of rods: (1) rods branched with high tuberculate processes, 0.08–0.09 mm long (Figure 5f), (2) rods with tubercles at one distal end of the rod, giving it a club-like appearance, 0.1–0.14 mm long (Figure 5a), and (3) rods with table-shaped tubercles, 0.08–0.12 mm long (Figure 5b).
Oral membrane has a blue colour in life (yellowish white when preserved in ethanol). The tentacles are brown in colour, partially with a greenish shimmer. Zooxanthellate.
Morphological variation.
The paratype colony consists of ~30 polyps. Tentacles of paratypes show variation in pinnule pairs, partly having nine pairs of widely spaced pinnules arranged in a single row on either side of rachis.
Distribution.
Udar Island, Kota Kinabalu, Sabah, Malaysia. Gaya Island, TARP, Kota Kinabalu, Sabah, Malaysia.
Remarks.
Polyps of Laeta waheedae gen. nov. et sp. nov. have the same blue-purple colour in life as those of Arula petunia McFadden & Ofwegen, 2012, but there is a difference in the outer margins and lobes of the oral membrane; both are more pronounced in Arula petunia. The holotype colony was attached to sponge tissue, but this epibiotic relation is not obligate.
Etymology.
Named after Dr. Zarinah Waheed, for her dedication to coral reef research and her guidance during fieldwork in Sabah.
Family Clavulariidae Hickson, 1894
Genus. Phenganax
Alderslade & McFadden, 2011
5B94F4202707519BA68EA0EAA5435705
Diagnosis.
(after Alderslade & McFadden 2011). Alcyonacea with erect polyps and stolons with encrusting, stoloniferous habit. Sclerites are absent. Zooxanthellate. Distribution tropical, Indo-Pacific. Type species: Phenganax parrini Alderslade & McFadden, 2011.
Phenganax marumi sp. nov.
27333CB5803E5D2685480AE7926D5905
http://zoobank.org/29410835-EB6E-4E9D-9361-CF60FE36ECFD
Figure 3.

In situ photographs of clavulariids found in and around TARP, Kota Kinabalu, Sabah, Malaysia. aClavulariidae sp., specimen NSMT-Co 1686 bPhenganax marumi sp. nov., holotype (NSMT-Co 1683) cPhenganax subtilis sp. nov., holotype (NSMT-Co 1684) dPhenganax stokvisi sp. nov., holotype (NSMT-Co 1685).
Material examined.
All specimens are from Kota Kinabalu, Sabah, Malaysia and collected by YW Lau. Holotype: NSMT-Co 1683, Manukan, Manukan Island, TARP (05°58'46.1"N, 116°00'10.6"E), 13 m depth. Paratypes: IPMB-C 01.00001: Edgell Patches, west of Sapi Island (06°00'38.7"N, 115°59'22.2"E), 16 m depth. IPMB-C 01.00002: Mid Reef Slope, east of Manukan Island, TARP (05°58'38.08"N, 116°00'52.82"E), 13 m depth. IPMB-C 01.00003: Mid Reef Slope, east of Manukan Island, TARP (05°58'38.08"N, 116°00'52.82"E), 11 m depth. IPMB-C 01.00004: Edgell Patches, west of Sapi Island (06°00'38.7"N, 115°59'22.2"E), 19 m depth. IPMB-C 01.00005: Edgell Patches, west of Sapi Island (06°00'38.7"N, 115°59'22.2"E), 16 m depth. IPMB-C 01.00006: Gaya Clement Reef, west of Gaya Island, TARP (06°01'24.26"N, 116°00'13.55"E), 12 m depth. IPMB-C 01.00007: Gaya Clement Reef, west of Gaya Island, TARP (06°01'24.26"N, 116°00'13.55"E), 11 m depth. IPMB-C 01.00008: Manukan, north of Manukan Island, TARP (05°58'46.1"N, 116°00'10.6"E), 12 m depth. IPMB-C 01.00009: Manukan, Manukan Island, TARP (05°58'46.1"N, 116°00'10.6"E), 12 m depth. IPMB-C 01.00010: Manukan, Manukan Island, TARP (05°58'46.1"N, 116°00'10.6"E), 12 m depth.
Description.
Colony is attached to rock and sponge and consists of ~15 polyps. Stolons are consistent in width throughout the colony (~0.35 mm). Polyps are spaced apart irregularly (from 0.5 mm up to 6 mm) and retract fully into calyces; calyces do not retract into stolon and are approximately 1 mm in width and 2 mm in height. Expanded polyps 4.0–5.5 mm in width. The pinnules are arranged on either side of the rachis in pairs of 12–15, sometimes spaced apart irregularly. The pinnules give a swollen appearance and are mostly conical shaped, otherwise diamond-shaped. The colony is sclerite-free. Polyps are brownish yellow in colour, with a bright yellowish white oral disk in life (whitish yellow when preserved in ethanol). Tentacles have a greenish colour interrupted with brown specks. Zooxanthellate.
Morphological variation.
Paratypes IPMB-C 01.00003 and IPMB-C 01.00007 show variation in the colouration of the polyps, having a paler and pinkish appearance. There is also variation in the number of pinnule pairs (~8 pairs) and the space between the pinnules; pinnules are sometimes so densely packed there is no space between them. Like the holotype, all paratype colonies are small colonies consisting of ~20 polyps.
Distribution.
Gaya, Sapi and Manukan Islands, TARP, Kota Kinabalu, Sabah, Malaysia.
Remarks.
Phenganax marumi sp. nov. resembles Phenganax parrini Alderslade & McFadden, 2011 in polyp morphology, but differs in colour. The numerous and densely packed polyps described for a P. parrini colony (Alderslade and McFadden 2011) have not been observed in any of the 11 collected P. marumi colonies; instead the maximum number of polyps observed in one colony was approximately 35. However, the density of P. parrini could be caused by unnatural conditions (aquarium environment) in which it was kept for scientific research purposes (Alderslade and McFadden 2011). Phenganax marumi was found on exposed spots on the reef, not sheltered from strong current or light, as opposed to P. parrini, which was found in dimly lit, sheltered locations.
Etymology.
From the Japanese word marumi, which means round or rounded; denoting the plump shape of the polyp tentacles, which give the polyp an overall plump appearance.
Phenganax subtilis sp. nov.
7A41C7E03E8059E7B416FEFF6536434F
http://zoobank.org/D01BA048-BF0F-4285-B4D3-3D071B9D1653
Material examined.
All specimens are from Kota Kinabalu, Sabah, Malaysia and collected by YW Lau. Holotype: NSMT-Co 1684, Sepangar, west of Sepangar Island (06°03'38.66"N, 116°04'0.65"E), 7 m depth. Paratypes: IPMB-C 01.00011: Sepangar II, west of Sepangar Island (06°04'7.38"N, 116°04'6.76"E), 7 m depth. IPMB-C 01.00012: Manukan, north of Manukan Island, TARP (05°58'46.1"N, 116°00'10.6"E), 12 m depth. IPMB-C 01.00013: Manukan, north of Manukan Island, TARP (05°58'46.1"N, 116°00'10.6"E), 12 m depth.
Description.
The colony consists of ~50 polyps, which are connected through thin rounded stolons with a width of approximately 0.13–0.2 mm, growing over coral rubble. Polyps are partly clustered and spaced apart irregularly (1.0–3.0 mm). Polyps retract into calyces (0.65–0.77 mm width) that are barrel shaped. Expanded polyps are 3.0–3.5 mm in width and have tentacles with pinnules that are widely spaced and arranged in pairs of 12 on either side of the rachis. No sclerites were found in any of the specimens. Polyps are brown coloured in life and whitish yellow when preserved in ethanol. Zooxanthellate.
Morphological variation.
Paratypes show variation in the number of pinnules on either side of the rachis (10–13 pairs).
Distribution.
Sepangar Island, Kota Kinabalu, Sabah, Malaysia. Manukan Island, TARP, Kota Kinabalu, Sabah, Malaysia.
Remarks.
Phenganax subtilis sp. nov. differs from P. parrini and P. marumi sp. nov. mainly in the shape of the pinnules. Phenganax subtilis does not have the plump, diamond-shaped pinnules as seen in P. marumi and P. parrini. Additionally, the colour of P. subtilis is different from P. parrini and P. marumi; brown instead of the more characteristic white-grey.
Etymology.
From Latin subtilis, meaning simple, subtle, plain; denoting the subtleness of the polyps blending into the reef background. Gender: masculine.
Phenganax stokvisi sp. nov.
DD32586E0B2E5DD7BE5BF6DB1821F2E1
http://zoobank.org/5871284F-05F8-469D-AED1-0BE797C0FEB2
Material examined.
All specimens are from dive location Mid Reef, east of Manukan Island (05°58'35.8"N, 116°00'52.2"E) TARP, Kota Kinabalu, Sabah, Malaysia and collected by YW Lau. Holotype: NSMT-Co 1685, 5 m depth. Paratypes: IPMB-C 01.00015: 4 m depth. IPMB-C 01.00014 4 m depth.
Description.
The colony consists of ~100 polyps which are densely packed and are connected through flattened ribbon-like stolons with irregular width (~0.21–0.55 mm). The colony is fragmented into two pieces with the rock it was attached to. Expanded polyps are approximately 3.0 mm in diameter and have tentacles with pinnules arranged in pairs of ten on either side of the rachis. Polyps are clustered closely together (adjacent to one another) and partly spaced apart (~1.0 mm in between clustered polyps). Polyps retract into calyces (~1.0 mm width) that are barrel shaped. No sclerites were found. Polyps are whitish grey coloured in life (whitish yellow when preserved in ethanol). Zooxanthellate.
Distribution.
Manukan Island, TARP, Kota Kinabalu, Sabah, Malaysia.
Remarks.
Phenganax stokvisi sp. nov. shows similarity to the densely packed P. parrini, polyps being in the same size range (2.5–3.0 mm width in P. parrini), only showing difference in the number of pinnule pairs and colour of the polyps (possibly due to zooxanthellae).
Etymology.
Named after Frank Robert Stokvis, whose passion for octocoral research has never changed since his first study on this topic.
Clavulariidae
sp.
4F253F95E71D5A93952393F39A2E08B4
Material examined.
All specimens are from Kota Kinabalu, Sabah, Malaysia. NSMT-Co 1686, Edgell Patches, west of Sapi Island (06°00'38.7"N, 115°59'22.2"E), 18 m depth, coll. YW Lau. IPMB-C 01.00016, Gaya Clement Reef, west of Gaya Island, TARP (06°01'24.26"N, 116°00'13.55"E), 11 m depth, coll. YW Lau.
Description.
Colonies with 20–30 polyps are connected through flattened stolons, which have a varying width of 0.5–1 mm. Colonies can be loosely attached to sponge or rocky substrates, such as coral rubble. Polyps are transparent and clustered in groups, connected by stolons with lengths up to 4–5 mm. Expanded polyps were ~6.0–7.0 mm in width when alive, with the pharynx visible in all polyps. Polyps retract fully into the calyx, which is cylinder-shaped (~1.3 mm width and up to 1.5 mm tall) and do not retract fully into the stolon. The tentacles have approximately 11 pairs of pinnules, which are widely spaced apart. No sclerites were found in the specimens. Polyps are whitish translucent when alive (yellowish white when preserved in ethanol). Azooxanthellate.
Distribution.
West of Sapi and Gaya Islands, TARP, Kota Kinabalu, Sabah, Malaysia.
Remarks.
This material, henceforth Clavulariidae sp., can be identified to the family level Clavulariidae Hickson, 1894 by its initial morphological resemblance to the type species Azoriella bayeri Lopez-Gonzalez & Gili, 2001 and Cervera atlantica Lopez-Gonzalez et al., 1995 in having similar whitish translucent polyps, although, the polyps of C. atlantica are translucently orange. However, more diagnostic morphological features and more specimens are necessary before a genus- and species-level distinction can be made.
The main difference between Clavulariidae sp. and A. bayeri can be found in the absence of sclerites in Clavulariidae sp. Additionally, both type species C. atlantica and A. bayeri have polyps that are smaller than in Clavulariidae sp.; C. atlantica, ~5.1 mm width, A. bayeri, ~3.6 mm width, and Clavulariidae sp., ~6.0–7.0 mm width. As well, C. atlantica and A. bayeri have more pinnules on either side of the tentacles than seen in Clavularidae sp.; both C. atlantica and A. bayeri have 12–14 pinnules and Clavulariidae sp. has tentacles with 11 pairs.
Molecular phylogenetic analyses
This study has produced 67 sequences, which were added to the public database GenBank; the 51 clavulariid, and 16 arulid sequences had no previous barcodes. The phylogenies resulting from the ML analyses of the separate markers (Suppl. material 1: Figures S1–S7) were highly congruent with those from the concatenated alignments for both the three- and four-marker datasets (Figures 6–7). ML and Bayesian analyses of the concatenated datasets yielded almost identical tree topologies.
Figure 6.
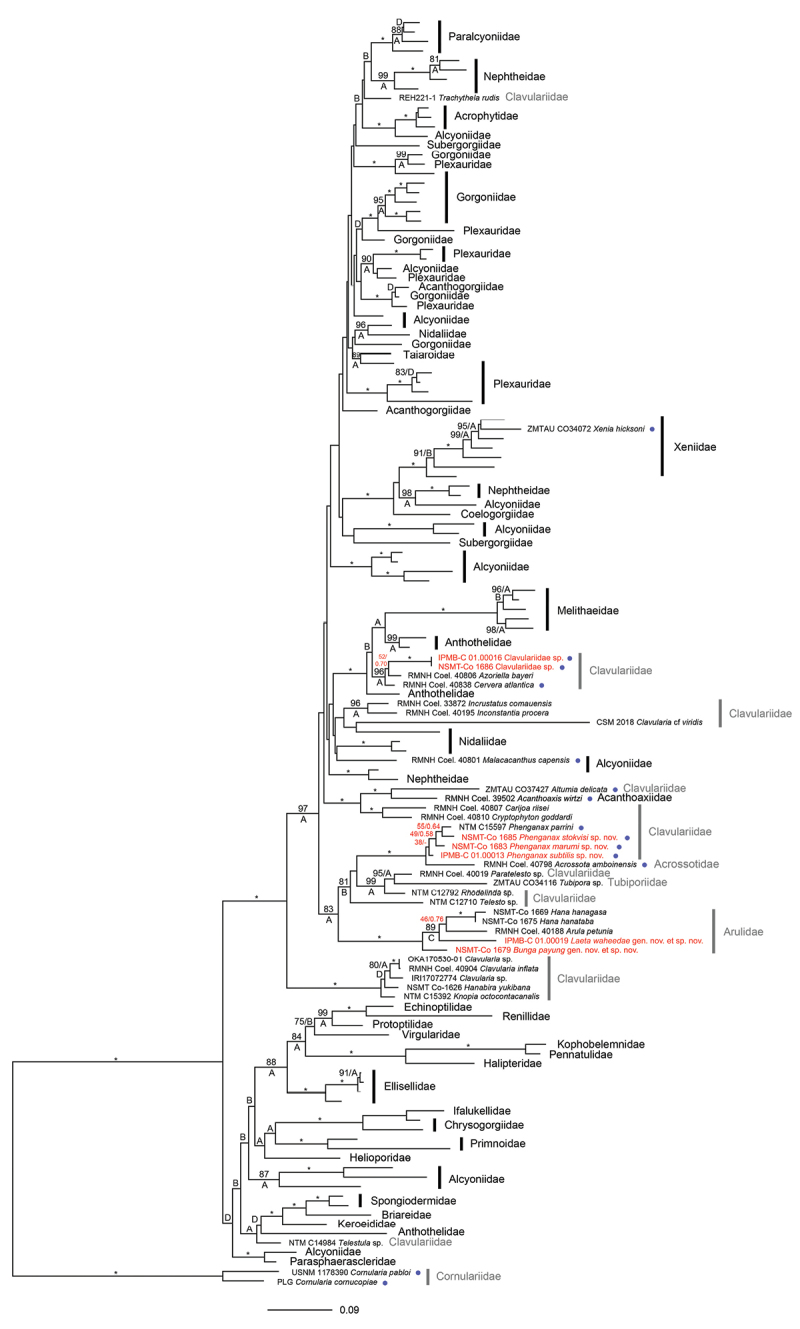
Phylogenetic relationships among 121 octocoral genera, including seven specimens (highlighted red) collected in and around TARP, Kota Kinabalu, Sabah, Malaysia, using the combined 28S rDNA+COI+mtMutS dataset (total n = 127). The best Maximum Likelihood tree is shown, with values at branches representing bootstrap probabilities (shown when > 70%; top/left) and Bayesian posterior probabilities (shown when > 0.80; bottom/right; A = 1.00, B = 0.95–0.99, C = 0.90–0.94, D = 0.80–0.89). Key: * represents 100%/1.00 for both analyses. Important values concerning target specimens are red. Non-stoloniferous families are shown with family classification only and stoloniferous families are highlighted in grey. Sclerite-free species are indicated with a blue dot. Cornularia spp. were used as outgroup.
Figure 7.

Combined 28S rDNA+COI+mtMutS+ND6 phylogenetic reconstruction for four Phenganax species and four arulid genera (Hana, Arula, Bunga gen. nov., and Laeta gen. nov.) collected in and around TARP, Kota Kinabalu, Sabah, Malaysia; Hanabira yukibana was used as outgroup (total n = 33). The best Maximum Likelihood tree is shown, with values at branches representing bootstrap probabilities (shown when > 50%; top/left) and Bayesian posterior probabilities (shown when > 0.50; bottom/right). Photograph credit: in situ images NTM C15597 Phenganax parrini, by Daniel Knop (modified from Alderslade and McFadden 2011, reproduced with permission from copyright holder). Sclerite images RMNH Coel. 40188 Arula petunia (modified from McFadden and Van Ofwegen 2012, reproduced with permission from CSIRO Publishing).
Arulidae from Sabah, Malaysia
Sequences of arulids Bunga payung gen. nov. et sp. nov. and Laeta waheedae gen. nov. et sp. nov., collected from the north of TARP (Udar, Gaya and Sepangar Islands) formed a completely-supported clade with the arulid genera Arula and Hana; 100%/1.00 for both the three- and four-marker datasets (Figures 6, 7, respectively). In both phylogenies, Bunga was sister to the remaining clade. However, it remains unresolved how Laeta waheedae, Arula, and Hana are related to one another in the four-marker phylogeny reconstruction (21%/0.56). Nonetheless, all genera in the clade have high support values.
Genetic distances (uncorrected P, expressed as percentage) between specimens of Bunga gen. nov., Laeta gen. nov., Arula, and Hana (between 2.6–4.6% for COI, 5.4–6.1% for mtMutS) indicated that the levels of pairwise differences were well above those observed among congeneric species (McFadden et al. 2011), Suppl. material 1: Table S1.
Morphological data supported the molecular data and justified the description of the two new genera Bunga and Laeta. There were distinct differences in sclerite features between Bunga, Laeta, Arula, and Hana, involving the presence and absence of sclerite types in the calyx; three types of rods (branched with high tuberculate processes, club-like rods and table-radiate-like rods), which were only observed in Laeta but not in the other sister genera, and the absence of 6-radiates in Bunga, which were present in all sister genera. There was also a difference in sclerites of the stolon; Arula is the only genus without fused table-radiates, but instead has table-radiates in the calyx. The polyp morphology did not show many obvious differences, although two of them could be observed in the outer margins of the characteristic fused oral membrane; both Bunga and Laeta had polyps with oral discs that were less pronounced at their outer margins than those of other genera, having a smooth transition from oral disc to the connecting tentacles, while the furrows dividing the oral disc into eight lobes were shallower in both of these genera (Figure 2). The combination of molecular and morphological information supports the erection of two new genera within the Arulidae.
Clavulariidae from Sabah, Malaysia
Sequences of 18 clavulariid specimens grouped together with Acrossota amboinensis (Burchardt, 1902) and Phenganax parrini Alderslade & McFadden, 2011 in a well-supported clade in both the three-marker and four-marker analyses (Figures 6, 7, respectively), in which Acrossota was sister to the remaining clade (100%/1.00). However, there was disparity in the relationships within the remaining clade in the two phylogeny reconstructions and the relationships within the remaining clade were not clearly resolved. The four-marker phylogeny reconstruction, with more specimens, gave a higher resolution (Figure 7) and here, each species had high support values, although it remained unclear how P. parrini is related to the other three species (50%/-).
On the other hand, within-genus genetic distances for Phenganax were 0.57% for COI and 3.10% for mtMutS, percentages within the range of congeneric species commonly observed in octocorals (Suppl. material 1: Table S3). When comparing pairwise differences between the four species, the values were well above averages for conspecifics; the observed values were in between 0.98–4.09% for COI and between 1.60–4.07% for mtMutS (Suppl. material 1: Table S5), supporting the division into four species (McFadden et al. 2011).
Despite the absence of sclerites, which was characteristic for the entire clade, there were visible differences in morphological polyp features. The main differences were found in tentacle pinnules and colony growth (Figure 3b–d). Phenganax marumi sp. nov. was similar to P. parrini Alderslade & McFadden, 2011 in polyp morphology, although differing in colour and polyp size; expanded polyp sizes ranged 4.0–5.5 mm in P. marumi and 2.5–3.0 mm in P. parrini. Additionally, the numerous amounts and densely packed polyps described for a P. parrini colony (Alderslade and McFadden 2011) had not been observed in any of the 11 collected P. marumi colonies; instead the maximum number of polyps observed for one colony was approximately 30–40. However, the density of P. parrini could be due to unnatural conditions (aquarium environment) in which it was kept for scientific research (Alderslade and McFadden 2011).
Phenganax marumi sp. nov. was found on exposed spots on the reef, not sheltered from strong current or light, opposed to P. parrini, which was found in dimly lit, sheltered locations. Phenganax subtilis sp. nov. differs from P. parrini and P. marumi mainly in the shape of the pinnules; P. subtilis does not have the plump, diamond-shaped pinnules as seen in P. marumi and P. parrini. As well, the colour of P. subtilis is different from P. parrini and P. marumi, being brown. Phenganax stokvisi sp. nov. shows similarity to the densely packed P. parrini, with polyps being in the same size range (2.5–3.0 mm width), but showing differences in the number of pinnule pairs and colour of the polyps (possibly due to the presence of zooxanthellae).
Differences in morphology combined with genetic distances and high support values in the four-marker phylogeny reconstruction supported the erection of the three new sister species of Phenganax parrini; P. marumi sp. nov., P. subtilis sp. nov., and P. stokvisi sp. nov.
Clavulariidae sp. had a similar DNA profile to two genera within the Clavulariidae; sequences from the two specimens clustered in a well-supported clade (96%/1.00), together with the genera Azoriella (Lopez Gonzalez & Gili, 2001) and Cervera Lopez-Gonzalez et al., 1995, with Cervera as sister to Azoriella and Clavulariidae sp. (Figure 6). Genetic distances between the two specimens and Azoriella were 1.41% for COI and 5.09% for mtMutS, values within ranges amongst intergeneric octocoral species (Suppl. material 1: Table S2). However, with the current available molecular data it remains unresolved how Clavulariidae sp. and Azoriella are related to one another, as branching was weakly supported (52%/0/70).
Discussion
From surveys at only eight different dive locations, approximately 13 hours of field work, all within approximately 60 km2 (TARP + area around Udar and Sepangar Islands), five species and two stoloniferan genera new to science were discovered. These results are not completely unexpected, as TARP is located on the outer edge of the Coral Triangle, where studies on stoloniferous octocorals are in their infancy. Of the seven families that are considered to belong to the subordinal group Stolonifera (Cordeiro et al. 2019), three have members that are confirmed to occur in the Indo-Pacific (Alderslade and McFadden 2007, 2011; Fabricius and Alderslade 2011): Acrossotidae Bourne, 1914, Tubiporidae Ehrenberg, 1828 and Clavulariidae Hickson, 1894. Of these, Clavulariidae is the only family that is not monogeneric or even monospecific, and is obviously the most speciose and common. The majority of recent studies on Indo-Pacific Clavulariidae have mainly focused on Tubipora musica Linnaeus, 1758, a reef-building alcyonarian species and Tubipora musica-sponge associations (Calcinai et al. 2013; Agustiadi and Luthfi 2017). Therefore, it is reasonable to assume that continued surveys in this area and within the Coral Triangle will result in further discoveries of undescribed stoloniferous octocorals. Such new descriptions could fill knowledge gaps in the overall systematics of Octocorallia, similar as recent discoveries of new small inconspicuous stoloniferan species have done (Alderslade and McFadden 2007, 2011; Benayahu et al. 2017; Lau et al. 2018, 2019).
Arulids of Sabah
There are minor differences in polyp morphology when comparing Bunga payung gen. nov. et sp. nov., Laeta waheedae gen. nov. et sp. nov., Arula petunia, and Hana spp.; Arula and Laeta are similar in colour, having blue colorations in the oral disc and/or tentacles. Hana spp. are similar to Arula petunia in having a very pronounced oral disc with deep furrows dividing the oral disc into eight distinct lobes, as opposed to Laeta and Bunga, which both have an oral disc that fuses into the tentacle base and shallow furrows that divides the eight lobes.
It could therefore be proposed that instead of four different genera within Arulidae, Arula, Hana, Bunga gen. nov., and Laeta gen. nov. could be five distinct species of Arula. However, the erection of a new genus for each new species is justified by the combination of sclerite morphology and molecular information. The four genera of Arulidae are distinguished morphologically by differences in sclerite form and the presence or absence of certain sclerite types in different parts of the colony or being absent completely. The only two sclerite types that all four genera have in common are the rods present in the anthocodiae and the table-radiates in the calyx, which are characteristic to the family Arulidae. Bunga is the only genus that has only these sclerites types in the polyps; anthocodial rods and table-radiates in the calyx. Arula, Hana, and Laeta have an additional sclerite type in the calyx; Arula and Hana have 6-radiates. Laeta has three different ornamented rods in the calyx, which are absent in all other genera.
Hana and Arula are the two most similar genera in terms of morphology. However, Arula is the only genus which has separate table-radiate sclerites in the stolon, and it can also be distinguished from the other genera in having table-radiates that are smaller in size (up to 0.09 mm length) and by not having table-radiates which are elongated in shape; the other three genera all have stolons with fused table-radiates which form a sheet and all have larger table-radiates up to 0.10–0.19 mm length.
Molecular analyses of the concatenated four marker dataset (28s rDNA, COI, mtMutS, and ND6 sequences) resulted in similar phylogenetic positions for Bunga payung gen. nov. et sp. nov. and Laeta waheedae gen. nov. et sp. nov. when compared to our analyses of the concatenated three marker dataset (28s rDNA, COI, and mtMutS). It should be noted that the two analyses including more specimens for Bunga and Laeta placed the four genera in slightly different phylogenetic positions. In the three-marker dataset (Suppl. material 1: Figure S8), Bunga and Laeta formed a clade, although it was not very well supported (74/-). This clade was sister to Hana and Arula, which also formed a clade. In the four-marker dataset (Figure 7), Bunga was sister to all remaining clades. Nonetheless, branches of the four-marker dataset had better support values. Therefore, we again recommend the use of the extra gene region (ND6) next to the conventional DNA markers (COI, mtMutS, 28S rDNA) commonly used in octocoral phylogenetic analyses (Lau et al. 2018, 2019).
New discoveries and descriptions of stoloniferous octocorals outside of the Coral Triangle have shown surprising octocoral novelty, at the species, genus, and family levels (Alderslade and McFadden 2011; McFadden and Ofwegen 2012; Benayahu et al. 2017; Lau et al. 2018, 2019). Arulidae, the most recently described family of stoloniferous octocorals (McFadden and Ofwegen 2012), is a good example. Arulids have unique morphological features that had not been seen before in octocorals, including an expanded fused oral disc, and a new type of sclerite, table-radiates. The species Bunga payung gen. nov. et sp. nov. and Laeta waheedae gen. nov. et sp. nov. increase the total number of species within the family Arulidae to five. Moreover, they represent the first confirmed records of arulids in and around the shallow waters of the TARP area in Sabah, Malaysia. The discovery of these two monospecific taxa demonstrates that the recently erected family Arulidae is considerably much more diverse than was originally known. For example, there is photographic evidence of an arulid from the reefs of Bali, Indonesia (McFadden and Ofwegen 2012), and it is possible that other regions in or close to the Coral Triangle harbour unknown stoloniferan diversity.
The distinctive feature of the fused oral membrane seen in arulid polyps and its function is not yet fully understood. It has been proposed by Porter (1976) and Sebens (1979) that an increase in size of the capture surface area of polyps could be positively related to the amount of prey captured (Lasker 1981; Lewis 1982), as has also been suggested for scleractinian corals (Todd et al. 2004; Todd 2008). On the other hand, a large polyp size in scleractinians has been observed to be beneficial for the capture and consumption of large prey items (Alamura et al. 2009; Hoeksema and Waheed 2012; Mehrotra et al. 2014, 2016, 2019; Musco et al. 2018). A study on octocorals by Lasker (1981) showed that there were differences in the number of nematocysts, bigger polyps having relatively less nematocysts, which could influence prey capture. The expanded oral disc seen in arulids could perhaps be an adaptation for feeding and compensation for their relatively small polyp size and lower polyp number per colony, and it would be worthwhile to conduct a study of their nematocysts.
Another compelling hypothesis involves the possibility of these zooxanthellate taxa being heterotrophic, in which an increased light-gathering surface for photosynthesis of symbionts could explain the development of the expanded oral disc. Based on currently available information, the expanded oral disc does not appear to have evolved across the Octocorallia radiation, and is unique to the family Arulidae.
Clavulariids of Sabah
Multiple factors made it initially difficult to establish with certainty if the currently described species were either congeners or possibly already described. The most important factor in this matter is the small number of morphological features that represent the new Phenganax species. The absence of sclerites makes it especially challenging to make confident judgements about evolutionary relationships between species within this genus. Additionally, the available historical literature on described stoloniferous species in the Indo- and northwestern Pacific is in a poor state. There are a number of Clavularia species listed by Alderslade and McFadden (2011) that could possibly belong to Phenganax, mainly due to the fact that these species also lack sclerites and the described distinguishing characters are alike: namely Clavularia reptans Hickson, 1894, Clavularia reptans sensu Thomson & Henderson (1906: 402), Clavularia celebensis Hickson, 1894, Clavularia pregnans Thomson & Henderson, 1906.
The main characteristic morphological difference between the Phenganax spp. described in this study and the above mentioned Clavularia spp. are the “numerous densely packed pinnules”, which are sometimes as many as 30 pinnules per row; all Phenganax spp. in the current study have six to 15 pinnules on either side of the tentacle rachis. Not all measurements (for example expanded polyp width) were included in the Clavularia spp. (Alderslade and McFadden 2011). Clavularia reptans was described as having polyps 7.0–10.0 mm width when expanded, whereas expanded polyp widths of Phenganax spp. are within the size range of 2.5–5.5 mm. Further information regarding the colouration of the tentacles and polyp body in combination with colony drawings make it doubtful that these Clavularia spp. belong to Phenganax. Additional arguments supporting the description of the Phenganax species in this study involve ecological aspects. The preferred habitat of P. parrini is dimly lit and in sheltered locations, below 10 m depth. The described Phenganax spp. in the current study, however, were found at exposed locations above 10 m depth.
Despite the fact that the relationships between the Phenganax spp., with emphasis on the position of P. parrini, seem to be unresolved, and all species lack sclerites to aid in their identification, the described species all had high support values and could be distinguished based on some features in colony growth form and polyp morphology, namely tentacle pinnules, the density of polyps, polyp size and spacing between polyps in a colony.
Similar to the new genera within Arulidae, the results for the Phenganax spp. also demonstrate that more specimens and the extra gene region (ND6) resulted in obtaining higher phylogenetic resolution and increased branch support values, and therefore should represent a more accurate reconstruction of evolutionary relationships. Future octocoral taxonomy will unquestionably build upon a combination of molecular and more traditional morphological techniques (Conti-Jerpe and Freshwater 2017); however, the lack of sclerites has important implications for future studies, as these skeletal parts are one of the major diagnostic features of the group. This emphasizes the need for improved molecular techniques, such as finding an optimum between concatenated marker datasets towards whole-genome mapping, combined with next-generation sequencing to allow more reliance on molecular information when morphological information is scarce.
The current and other recent works show that octocoral species without sclerites are not rarities; the absence of sclerites or other calciferous skeletal parts are known from within seven octocoral families (Acanthoaxiidae, Acrossotidae, Cornulariidae, Dendrobrachiidae, Alcyoniidae, Clavulariidae, Xeniidae) (Alderslade and McFadden 2007, 2011; López-González et al. 1995; Benayahu et al. 2017). Sclerites provide protection against predation, provide support and rigidity to soft colony parts, and are critical in octocoral classification (West 1997; Aharonovich and Benayahu 2012). Sclerites in different species differ in density as well as spatial organisation in the connective tissue, and thus they also differ in the degree to which they offer support and protection (Benayahu et al. 2017). It could be hypothesized that small size and inhabiting small cracks on reefs protects small stoloniferous species from exposure and has eliminated the need for sclerites in some of these species. Alternately, perhaps the absence of predation (e.g., by ovuliids or sea slugs) has made the possession of sclerites unnecessary. However, there is no obvious pattern to be found for this character in the octocoral phylogenetic radiation amongst its smallest members (Figure 6), and more work is needed to confirm this hypothesis. The absence of sclerites is an example of the gaps in our present knowledge about octocorals and indicate how much basic work in alpha taxonomy and documentation remains for this group.
Our results demonstrate that the TARP area, off the coast of Kota Kinabalu, in Sabah, Malaysia, harbours stoloniferous octocoral diversity that was previously unknown to science, which is obvious since it represents one of the first studies focusing on Stolonifera in this region. However, the newly described sclerite-free Phenganax spp. as well as Clavulariidae sp. are only small pieces of the puzzle in the needed thorough investigation of the polyphyletic family Clavulariidae (McFadden and Ofwegen 2012; Benayahu et al. 2017; Conti-Jerpe and Freshwater 2017; Lau et al. 2019), and an even smaller step towards a full understanding of the morphological and molecular distinctions amongst clades of Stolonifera and ultimately Octocorallia. Molecular data indicate that the entire Octocorallia classification needs re-structuring, but for now researchers still rely on the traditional morphological classification system (Fabricius and Alderslade 2001; Conti-Jerpe and Freshwater 2017), as evolutionary relationships are still far from being fully resolved (McFadden and Ofwegen 2012; Conti-Jerpe and Freshwater 2017). Integrative taxonomy combining not only morphology and molecular biology but also including ecology and biogeography data appears to be the best way to try to better understand the systematics of such a diverse and important group in marine benthic communities (Perez et al. 2016). To this end, marine parks should include in their goal not only the preservation of commercially important faunal diversity (primarily reef fish), but also of other reef organisms, and in particular, small and inconspicuous cryptic species, as it is these small reef inhabitants that make up the vast majority of the diversity of coral reef communities (Hoeksema 2017).
Supplementary Material
Acknowledgements
We thank Dr. Z. Waheed (Universiti Malaysia Sabah) for her support regarding research permissions and transport license. Access license to Tunku Abdul Rahman Park (TARP) was granted on authority of Sabah Biodiversity Centre and Sabah Parks (Ref: TTS/IP/100-6/2 Jld.8). The boat crew of Universiti Malaysia Sabah (UMS) is thanked for their logistic support in the field. Dr. Z. Waheed, M.F.A. Nooramli and S.Q. Pang (UMS) are thanked for help in the field. We thank Prof. E. Hirose (University of the Ryukyus) for providing access to the scanning electron microscope (SEM) used in this work. F.R. Stokvis (Naturalis Biodiversity Center) is thanked for his valuable help and advice concerning the phylogenetic analyses of the octocoral data. Dr. L.P. van Ofwegen is thanked for sharing his expert advice. We thank the reviewers, Prof. C.S. McFadden (Harvey Mudd College) and Dr. O. Breedy (University of Costa Rica), for their thorough and helpful comments, which undoubtedly have contributed to a better manuscript. Finally, we would like to thank Prof. B.W. Hoeksema for helping us with his editorial skills. Fieldwork by the first author in TARP, Kota Kinabalu, Sabah, Malaysia was funded by the Okinawa Research Core for Highly Innovative Discipline Science Projects (ORCHIDS). The first author was supported by a MEXT, Japan Student Services Organization (JASSO) scholarship. All material included in this manuscript was collected under approved collection permits and ethics guidelines.
Citation
Lau YW, Reimer JD (2019) A first phylogenetic study on stoloniferous octocorals off the coast of Kota Kinabalu, Sabah, Malaysia, with the description of two new genera and five new species. ZooKeys 872: 127–158. https://doi.org/10.3897/zookeys.872.36288
Funding Statement
Okinawa Research Core for Highly Innovative Discipline Science Projects (ORCHIDS) research grant
Supplementary materials
This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.
Yee Wah Lau, James D. Reimer
Figures S1–S8, Tables S1–S6
Data type: Adobe Acrobat Document (.pdf)
References
- Aberer AJ, Kobert K, Stamatakis A. (2014) ExaBayes: massively parallel Bayesian tree inference for the whole-genome era. Molecular Biology and Evolution 31: 2553–2556. 10.1093/molbev/msu236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agustiadi T, Luthfi OM. (2017) Diversity of stoloniferan coral (Stolonifera) at Lirang Island, Southwest Maluku (Moluccas), Indonesia. International Journal of Oceans and Oceanography 11: 21–30. [Google Scholar]
- Aharonovich D, Benayahu Y. (2012) Microstructure of octocoral sclerites for diagnosis of taxonomic features. Marine Biodiversity 42: 173–177. 10.1007/s12526-011-0102-3 [DOI] [Google Scholar]
- Alamaru A, Bronstein O, Loya Y, Dishon G. (2009) Opportunistic feeding by the fungiid coral Fungia scruposa on the moon jellyfish Aurelia aurita Coral Reefs 28: 865. 10.1007/s00338-009-0507-7 [DOI]
- Alderslade P, McFadden CS. (2007) Pinnule-less polyps: a new genus and new species of Indo-Pacific Clavulariidae and validation of the soft coral genus Acrossota and the family Acrossotidae (Coelenterata: Octocorallia). Zootaxa 1400: 27–44. 10.11646/zootaxa.1400.1.2 [DOI] [Google Scholar]
- Alderslade P, McFadden CS. (2011) A new sclerite-free genus and species of Clavulariidae (Coelenterata: Octocorallia). Zootaxa 3104: 64–68. 10.11646/zootaxa.3104.1.6 [DOI] [Google Scholar]
- Benayahu Y, Jeng MS, Perkol-Finkel S, Dai CF. (2004) Soft corals (Octocorallia: Alcyonacea) from southern Taiwan. II. Species diversity and distributional patterns. Zoological Studies 43: 548–560. [Google Scholar]
- Benayahu Y, McFadden CF, Shoham E. (2017) Search for mesophotic octocorals (Cnidaria, Anthozoa) and their phylogeny: I. A new sclerite-free genus from Eilat, northern Red Sea. ZooKeys 680: 1–11. 10.3897/zookeys.680.12727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birkeland C. (1997) Life and death of coral reefs, Chapman and Hall, New York, 536 pp. 10.1007/978-1-4615-5995-5 [DOI]
- Breedy O, Cortés J. (2008) Octocorals (Coelenterata: Anthozoa: Octocorallia) of Isla del Coco, Costa Rica. Revista de Biologia Tropical 56 (Supplement 2): 71–77.
- Burchardt E. (1902) Alcyonaceen von Thursday Island (Torres-Strasse) und von Amboina, II. Jenaische Denkschriften fur Medizinsch-naturwessen-schaftliche Gessellschaft zu Jena, 8: 655–682. [pls 54–57] [Google Scholar]
- Burke L, Selig E, Spalding M. (2002) Reefs at risk in Southeast Asia. World Resources Institute, USA, 72 pp https://wriorg.s3.amazonaws.com/s3fs-public/pdf/rrseasia_full.pdf [Google Scholar]
- Calcinai B, Bavestrello G, Bertolino M, Pica D, Wagner D, Cerrano C. (2013) Sponges associated with octocorals in the Indo-Pacific, with the description of four new species. Zootaxa 3617: 1–61. 10.11646/zootaxa.3617.1.1 [DOI] [PubMed] [Google Scholar]
- Carpenter KE, Abrar M, Aeby GS, Aronson RB, Banks S, Bruckner A, Chiriboga A, Cortés J, Charles Delbeek JC, DeVantier L, Edgar GJ, Edwards AJ, Fenner D, Guzmán HM, Hoeksema BW, Hodgson G, Johan O, Licuanan WY, Livingstone SR, Lovell ER, Moore JA, Obura DO, Ochavillo D, Polidoro BA, Precht WF, Quibilan MC, Reboton C, Richards Z, Rogers AD, Sanciangco J, Sheppard A, Sheppard C, Smith J, Stuart S, Turak E, Veron JEN, Wallace C, Weil E, Wood E. (2008) One-third of reef-building corals face elevated extinction risk from climate change and local impacts. Science 321: 560–563. 10.1126/science.1159196 [DOI] [PubMed] [Google Scholar]
- Conti-Jerpe IE, Freshwater DW. (2017) Hedera caerulescens (Alcyonacea: Alcyoniidae), a new genus and species of soft coral from the temperate North Atlantic: invasive in its known range? Invertebrate Systematics 31: 723–733. 10.1071/IS16069 [DOI]
- Dias TLP, Gondim AI. (2016) Bleaching in scleractinians, hydrocorals, and octocorals during thermal stress in a northeastern Brazilian reef. Marine Biodiversity 46: 303–307. 10.1007/s12526-015-0342-8 [DOI] [Google Scholar]
- Fabricius KE, Alderslade P. (2001) Soft corals and sea fans: a comprehensive guide to the tropical shallow water genera of the central-west Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science, Townsville.
- Fabricius KE, Alderslade P, Williams GC, Colin PL, Golbuu Y. (2007) Octocorallia in Palau, Micronesia: Effects of biogeography and coastal influences on local and regional biodiversity. In: Kayanne H, Omori M, Fabricius K, Verheij E, Colin P, Golbuu Y, Yurihira H (Eds) Coral Reefs of Palau. Palau International Coral Reef Centre, Palau, 231 pp. [Google Scholar]
- Fisher R, O’Leary RA, Low-Choy S, Mengersen K, Knowlton N, Brainard RE, Caley MJ. (2015) Species richness on coral reefs and the pursuit of convergent global estimates. Current Biology 25: 500–505. 10.1016/j.cub.2014.12.022 [DOI] [PubMed] [Google Scholar]
- Foster PG. (2004) Modeling compositional heterogeneity. Systematic Biology 53: 485–495. 10.1080/10635150490445779 [DOI] [PubMed] [Google Scholar]
- Gaurav V, Lohman DJ, Meier R. (2011) SequenceMatrix: concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 27: 171–180. 10.1111/j.1096-0031.2010.00329.x [DOI] [PubMed] [Google Scholar]
- Goreau T, McClanahan T, Hayes R, Strong A. (2000) Conservation of coral reefs after the 1998 global bleaching event. Conservation Biology 14: 5–15. 10.1046/j.1523-1739.2000.00011.x [DOI] [Google Scholar]
- Hall TA. (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series 41: 95–98. jwbrown.mbio.ncsu.edu/JWB/papers/1999Hall1.pdf
- Heery EC, Hoeksema BW, Browne NK, Reimer JD, Ang PO, Huang D, Friess DA, Chou LM, Loke LHL, Saksena-Taylor P, Alsagoff N, Yeemin T, Sutthacheep M, Vo ST, Bos AR, Gumanao GS, Syed Hussein MA, Waheed Z, Lane DJW, Johan O, Kunzmann A, Jompa J, Suharsono, Taira D, Bauman AG, Todd PA. (2018) Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Marine Pollution Bulletin 135: 654–681. 10.1016/j.marpolbul.2018.07.041 [DOI] [PubMed] [Google Scholar]
- Hoeksema BW. (2007) Delineation of the Indo-Malayan centre of maximum marine biodiversity: The Coral Triangle. In: Renema W. (Ed.) Biogeography, Time, and Place: Distributions, Barriers, and Islands.Springer, Dordrecht, 117–178. 10.1007/978-1-4020-6374-9_5 [DOI]
- Hoeksema BW. (2017) The hidden biodiversity of tropical coral reefs. Biodiversity 18: 8–12. 10.1080/14888386.2017.1307787 [DOI] [Google Scholar]
- Hoeksema BW, Waheed Z. (2012) It pays to have a big mouth: mushroom corals ingesting salps at Sabah, Malaysia. Marine Biodiversity 42: 297–302. 10.1007/s12526-012-0110-y [DOI] [Google Scholar]
- Huang D, Licuanan WY, Hoeksema BW, Chen CA, Ang PO, Huang H, Lane DJW, Vo ST, Waheed Z, Amri AY, Yeemin T, Chou LM. (2015) Extraordinary diversity of reef corals in the South China Sea. Marine Biodiversity 45: 157–168. 10.1007/s12526-014-0236-1 [DOI] [Google Scholar]
- Hughes TP, Bellwood DR, Connolly SR. (2002) Biodiversity hotspots, centres of endemicity, and the conservation of coral reefs. Ecology Letters 5: 775–784. 10.1046/j.1461-0248.2002.00383.x [DOI] [Google Scholar]
- Hughes TP, Kerry JT, Álvarez-Noriegal M, Álvarez-Romero JG, Anderson KD, Baird AH, Babcock RC, Beger M, Bellwood DR, Berkelmans R, Bridge TC, Butler IR, Byrne M, Cantin NE, Comeau S, Connolly SR, Cumming GS, Dalton SJ, Diaz-Pulido G, Eakin CM, Figueira WF, Gilmour JP, Harrison HB, Heron SF, Hoey AS, Hobbs JPA, Hoogenboom MO, Kennedy EV, Kuo CY, Lough JM, Lowe RJ, Liu G, McCulloch MT, Malcolm HA, McWilliam MJ, Pandolfi JM, Pears RJ, Pratchett MS, Schoepf V, Simpson T, Skirving WJ, Sommer B, Torda G, Wachenfeld DR, Willis BL, Wilson SK. (2017) Global warming and recurrent mass bleaching of corals. Nature 543: 373–377. 10.1038/nature21707 [DOI] [PubMed] [Google Scholar]
- Hughes TP, Kerry JT, Simpson T. (2018) Large‐scale bleaching of corals on the Great Barrier Reef. Ecology 99: 501. 10.1002/ecy.2092 [DOI] [PubMed]
- Johnson JY. (1861) Notes on the sea-anemones of Madeira, with descriptions of new species. Proceedings of the Zoological Society of London 1861: 298–306. [Google Scholar]
- Kassem K, Hoeksema BW, Affendi YA. (2012) Semporna Marine Ecological Expedition. WWF-Malaysia, NCB Naturalis, Universiti Malaysia Sabah. Kota Kinabalu, Malaysia, 267 pp. [Google Scholar]
- Katoh K, Standley DM. (2013) MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Molecular Biology and Evolution 30: 772–780. 10.1093/molbev/mst010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kearse M, Moir M, Wilson A, Stones-havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Meintjes P, Drummond A. (2012) Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28: 1647–1649. 10.1093/bioinformatics/bts199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S, Stecher G, Li M, Knyaz C, Tamura K. (2018) MEGA X: Molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution 35: 1547–1549. 10.1093/molbev/msy096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lane DJW, Hoeksema BW. (2016) Mesophotic mushroom coral records at Brunei Darussalam support westward extension of the Coral Triangle to the South China Sea waters of Northwest Borneo. Raffles Bulletin of Zoology 64: 204–212. [Google Scholar]
- Larsson A. (2014) AliView: a fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 30: 3276–3278. 10.1093/bioinformatics/btu531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lasker HR. (1981) A comparison of the particulate feeding abilities of three species of gorgonian soft coral, Marine Ecology Progress Series 5: 61–67. https://www.int-res.com/articles/meps/5/m005p061.pdf
- Lau YW, Stokvis FR, Ofwegen LP van, Reimer JD. (2018) Stolonifera from shallow waters in the north-western Pacific: a description of a new genus and two new species within the Arulidae (Anthozoa, Octocorallia). ZooKeys 790: 1–19. 10.3897/zookeys.790.28875 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau YW, Stokvis FR, Imahara Y, Reimer JD. (2019) The stoloniferous octocoral, Hanabira yukibana, gen. nov., sp. nov., of the southern Ryukyus has morphological and symbiont variation. Contributions to Zoology 88: 54–77. 10.1163/18759866-20191355 [DOI] [Google Scholar]
- Lewis JB. (1982) Feeding behaviour and feeding ecology of the Octocorallia (Coelenterata: Anthozoa) Journal of Zoology 196: 371–384. 10.1111/j.1469-7998.1982.tb03509.x [DOI]
- López-González PJ, Gili JM. (2001) A new genus and species of stoloniferous octocoral from the Azores Archipelago (Cnidaria: Anthozoa). Bulletin of the Biological Society of Washington 10: 130–139. [Google Scholar]
- López-González PJ, Ocaña O, García-Gómez JC, Núñez J. (1995) North-eastern Atlantic and Mediterranean species of Cornulariidae Dana, 1846 (Anthozoa: Stolonifera) with the description of a new genus. Zoologische Mededelingen Leiden 69: 261–272. [Google Scholar]
- Lulofs RB. (1973) A reef survey of Pulau Gaya and associated islands Part I. World Wide Fund for Nature (WWF) Malaysia, 16 pp.
- Lulofs RB, Langham NPE, Mathias JA. (1974) A reef survey of Pulau Gaya and associated islands Part II. World Wide Fund for Nature (WWF) Malaysia, 7 pp.
- Mathias JA, Langham NPE. (1978) Chapter 5: Coral reefs. In: Chua TE, Mathias JA. (Eds) Coastal resources of West Sabah.Universiti Sains Malaysia, Penang, 117–151.
- McFadden CS, Ofwegen LP van. (2012) Stoloniferous octocorals (Anthozoa, Octocorallia) from South Africa, with descriptions of a new family of Alcyonacea, a new genus of Clavulariidae, and a new species of Cornularia (Cornulariidae). Invertebrate Systematics 26: 331–356. 10.10071/IS12035 [DOI] [Google Scholar]
- McFadden CS, Sanchez JA, France SC. (2010) Molecular phylogenetic insights into the evolution of Octocorallia: a review. Integrative and Comparative Biology 50: 389–410. 10.1093/icb/icq056 [DOI] [PubMed] [Google Scholar]
- McFadden CS, Benayahu Y, Pante E, Thoma JN, Nevarez PA, France SC. (2011) Limitations of mitochondrial gene barcoding in Octocorallia. Molecular Ecology Resources 11: 19–31. 10.1111/j.1755-0998.2010.02875.x [DOI] [PubMed] [Google Scholar]
- Mehrotra R, Scott CM, Rohrer JM, Hoeksema BW. (2015) Predation on a sacoglossan gastropod by a mushroom coral. Coral Reefs 34: 517. 10.1007/s00338-015-1285-z [DOI]
- Mehrotra R, Scott CM, Hoeksema BW. (2016) A large gape facilitates predation on salps by Heteropsammia corals. Marine Biodiversity 46: 323–324. 10.1007/s12526-015-0379-8 [DOI] [Google Scholar]
- Mehrotra R, Scott CM, Monchanin C, Phongsuwan N, Caballer M, Chavanich S, Hoeksema BW. (2019) Selective consumption of sacoglossan sea slugs (Mollusca: Gastropoda) by scleractinian corals (Cnidaria: Anthozoa). PLoS ONE 14: e0215063. 10.1371/journal.pone.0215063 [DOI] [PMC free article] [PubMed]
- Miyazaki Y, Reimer JD. (2015) A new genus and species of octocoral with aragonite calcium-carbonate skeleton (Octocorallia, Helioporacea) from Okinawa, Japan. ZooKeys 511: 1–23. 10.3897/zookeys.511.9432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Musco L, Vega Fernandez T, Caroselli E, Roberts JM, Badalamenti F. (2018) Protocooperation among small polyps allows the coral Astroides calycularis to prey on large jellyfish. Ecology 99: 2400–2401. 10.1002/ecy.2413 [DOI] [PubMed] [Google Scholar]
- Nyanti L, Johnston NA. (1992) The coral reefs of the Tunku Abdul Rahman Park, Sabah. Sabah Society Journal 9: 323–348. [Google Scholar]
- Pérez CD, de Moura Neves B, Cordeiro RT, Williams GC, Cairns SD. (2016) Diversity and Distribution of Octocorallia. In: Goffredo S, Dubinsky Z. (Eds) The Cnidaria, Past, Present and Future.Springer, Cham, 109–123. 10.1007/978-3-319-31305-4_8 [DOI]
- Pilcher N, Cabanban AS. (2000) The status of coral reefs in eastern Malaysia. In: Global Coral Reef Monitoring Network (GCRMN) Report. Australia Institute of Marine Science, Townsville, 81 pp. [Google Scholar]
- Polunin NVC, Roberts CM. (1996) Reef Fisheries, Chapman and Hall, London, 477 pp. 10.1007/978-94-015-8779-2 [DOI]
- Porter JW. (1976) Autotrophy, heterotrophy and resource partitioning in Caribbean reef building corals. The American Naturalist 110: 731–742. 10.1086/283100 [DOI] [Google Scholar]
- Prada C, Weil E, Yoshioka PM. (2010) Octocoral bleaching during unusual thermal stress. Coral Reefs 29: 41–45. 10.1007/s00338-009-0547-z [DOI] [Google Scholar]
- Praveena SM, Siraj SS, Aris AZ. (2012) Coral reefs studies and threats in Malaysia: a mini review, Reviews in Environmental Science and Biotechnology 11: 27–39. 10.1007/s11157-011-9261-8 [DOI]
- Rambout A. (2014) FigTree.
- Ranwez V, Harispe S, Delsuc F, Douzery EJP. (2011) MACSE: Multiple Alignment of Coding SEquences accounting for frameshifts and stop codons. PLoS ONE 6: e22594. 10.1371/journal.pone.0022594 [DOI] [PMC free article] [PubMed]
- Reaka-Kudla ML. (1997) The global biodiversity of coral reefs: A comparison with rain forests, 83–108. In: Reaka-Kudla ML, Wilson DE, Wilson EO (Eds) Biodiversity II: Understanding and protecting our biological resources. Joseph Henry Press, Washington, DC, 549 pp citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.466.2268&rep=rep1&type=pdf [Google Scholar]
- Reef Check Malaysia (2014) Status of coral reefs in Malaysia, 2014. Saving our reefs, Research, Education, Conservation, 72 pp https://data.nodc.noaa.gov/coris/library/NOAA/Non-CRCP/Corals/20150320_RCM_Survey_Report_2014.pdf [Google Scholar]
- Roberts CM, Mittermeier CG, Schueler FW. (2002) Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 295: 1280–1285. 10.1126/science.1067728 [DOI] [PubMed] [Google Scholar]
- Sabroux R, Hassanin A, Corbari L. (2019) Four times more species of sea spiders (Arthropoda: Pycnogonida) in Martinique Island (Lesser Antilles). Marine Biodiversity 49: 1519–1535. 10.1007/s12526-019-00957-9 [DOI] [Google Scholar]
- Samimi-Namin K, van Ofwegen LP. (2012) The octocoral fauna of the Gulf. In: Riegl BM, Purkis S. (Eds) Coral Reefs of the Gulf: Adaptation to Climatic Extremes.Springer; Netherlands, 225–252. 10.1007/978-94-007-3008-3_12 [DOI]
- Sánchez JA. (2016) Diversity and evolution of octocoral animal forests at both sides of Tropical America. 20–21. In: Rossi S, Bramanti L, Gori A, Orejas Saco del Valle C (Eds) Marine Animal Forests. Springer, Cham. 10.1007/978-3-319-17001-5_39-1 [DOI]
- Sánchez JA, McFadden CS, France SC, Lasker HR. (2003) Molecular phylogenetic analyses of shallow-water Caribbean octocorals. Marine Biology 142: 975–9872. 10.1007/s00227-003-1018-7 [DOI] [Google Scholar]
- Sebens KP. (1979) The energetics of asexual reproduction and colony formation in benthic marine invertebrates. American Zoologist 19: 683–697. 10.1093/icb/19.3.683 [DOI] [Google Scholar]
- Spait M. (2001) Paper 16. Marine park management: Issues and challenges, 6th SITE Research Seminar, 13–14 September 2001: 1–11. 10.1016/S1353-4858(01)01102-3 [DOI]
- Stamatakis A. (2014) RAxML version 8: a tool for phylogenetic analyses and post-analysis of large phylogenies. Bioinformatics 30: 1312–1313. 10.1093/bioinformatics/btu033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd PA. (2008) Morphological plasticity in scleractinian corals. Biological Reviews 83: 315–337. 10.1111/j.1469-185X.2008.00045.x [DOI] [PubMed] [Google Scholar]
- Todd PA, Sidle RC, Lewin-Koh NJI. (2004) An aquarium experiment for identifying the physical factors inducing morphological change in two massive scleractinian corals. Journal of Experimental Marine Biology and Ecology 299: 97–113. 10.1016/j.jembe.2003.09.005 [DOI] [Google Scholar]
- Townsend D. (2015) First comprehensive list of the coral reef fishes of Tunku Abdul Rahman Park, Sabah, Malaysia (Borneo). Check List 11: 1762. 10.15560/11.7.1762 [DOI]
- Van de Water JAJM, Allemand D, Ferrier-Pagés C. (2018) Host-microbe interactions in octocoral holobionts – recent advances and perspectives. Microbiome 6: 64. 10.1186/s40168-018-0431-6 [DOI] [PMC free article] [PubMed]
- Van Soest RWM, Verseveldt J. (1987) Unique symbiotic octocoral-sponge association from Komodo. Indo-Malayan Zoology 4: 27–32. [Google Scholar]
- Veron J, Stafford-Smith M, DeVantier L, Turak E. (2015) Overview of distribution patterns of zooxanthellate Scleractinia Frontiers in Marine Science 1: 81. 10.3389/fmars.2014.00081 [DOI]
- Waheed Z, Hoeksema BW. (2013) A tale of two winds: species richness patterns of reef corals around the Semporna Peninsula, Malaysia. Marine Biodiversity 43: 37–51. 10.1007/s12526-012-0130-7 [DOI] [Google Scholar]
- Waheed Z, Hoeksema BW. (2014) Diversity patterns of scleractinian corals at Kota Kinabalu, Malaysia, in relation to exposure and depth. Raffles Bulletin of Zoology 62: 66–82. https://lkcnhm.nus.edu.sg/app/uploads/2017/06/62rbz066-082.pdf [Google Scholar]
- Waheed Z, Adnan FAF, Hwa LCH, Hashim SRM. (2007) Status of coral reefs and sedimentation at Kota Kinabalu: a preliminary study at Gaya Bay and Sepangar Bay. Borneo Journal of Marine Science and Aquaculture: 27–43. http://wwwsst.ums.edu.my/data/file/BMzNMit3WmB3.pdf
- Waheed Z, van Mil HGJ, Syed Hussein MA, Jumin R, Golam Ahad B, Hoeksema BW. (2015a) Coral reefs at the northernmost tip of Borneo: an assessment of scleractinian species richness patterns and benthic reef assemblages. PLoS ONE 10(12): e0146006. 10.1371/journal.pone.0146006 [DOI] [PMC free article] [PubMed]
- Waheed Z, Benzoni F, van der Meij SET, Terraneo TI, Hoeksema BW. (2015b) Scleractinian corals (Fungiidae, Agariciidae and Euphylliidae) of Pulau Layang-Layang, Spratly Islands, with a note on Pavona maldivensis (Gardiner, 1905). ZooKeys 517: 1–37. 10.3897/zookeys.517.9308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- West JM. (1997) Plasticity in the sclerites of a gorgonian coral: tests of water motion, light level, and damage cues. The Biological Bulletin 192: 279–289. 10.2307/1542721 [DOI] [PubMed] [Google Scholar]
- Williams RB. (1996) The rediscovery of Cervera atlantica (Johnson, 1861) (Cnidaria: Octocorallia): notes on its identification, ecology and geographical distribution. Bulletin Zoölogisch Museum Amsterdam 15: 65–74. [Google Scholar]
- Wolf NG, Bermingham EB, Reaka ML. (1983) Relationships between fishes and mobile benthic invertebrates on coral reefs, 89–96. In: ML Reaka (Ed.) The ecology of deep and shallow coral reefs, Vol. 1. Washington, DC: National Oceanic and Atmospheric National Undersea Research Program. aoml.noaa.gov/general/lib/CREWS/SaltRiver/salt_river26.pdf
- Wood EM. (1977) Coral reefs in Sabah: present damage and potential dangers. Malayan Nature Journal 31: 49–57. [Google Scholar]
- Wood EM. (1979) Ecological study of coral reefs in Sabah. Technical report, WWF Project Number MYS 15, Petaling Jaya, Malaysia, 163 pp. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.
Yee Wah Lau, James D. Reimer
Figures S1–S8, Tables S1–S6
Data type: Adobe Acrobat Document (.pdf)