

Abstract
The endocannabinoid (eCB) signaling system plays a key role in short-term and long-term synaptic plasticity in brain regions involved in various neural functions ranging from action selection to appetite control. This review will explore the role of eCBs in shaping neural circuit function to regulate behaviors. In particular, we will discuss the behavioral consequences of eCB mediated long-term synaptic plasticity in different brain regions. This review brings together evidence from in vitro and ex vivo studies and points out the need for more in vivo studies.
Keywords: Arachidonoylethanolamine, 2-arachidonoylglycerol, cannabinoid receptors, depolarization-induced suppression of inhibition, depolarization-induced suppression of excitation, long-term depression, long-term potentiation
Graphical Abstract
INTRODUCTION
Endogenous cannabinoids, also referred to as endocannabinoids (eCBs), are small lipid-derived signaling molecules found throughout the body, including the central and peripheral nervous systems.1–4 In the brain, these signaling molecules differ from canonical vesicle-packaged neurotransmitters, such as glutamate and monoamine neuromodulators, such as dopamine. eCBs are directly synthesized from membrane phospholipids in response to neuronal activity.5–7 These fatty acid derivatives cannot be packaged into vesicles, and there is no obvious mechanism for intracellular storage. Thus, their production is mainly thought to be “on-demand” and “use-dependent”.5–7 In addition to the traditional “on-demand” synthesis of eCBs, there is evidence for presynthesized pools of eCBs that may be released in a stimulus specific manner.8,9 It is not known how these eCB “pools” are maintained within a cell or released. In the nervous system, eCBs mainly modulate synaptic transmission via “retrograde signaling” in which postsynaptic release of the molecule leads to actions on presynaptic receptors.10–12 There is evidence that eCBs can participate in direct receptor-mediated (nonretrograde) signaling, sometimes even in a receptor-independent manner.13–16 Due to their transient and tightly regulated production, eCBs have the potential to quickly adjust the gain of incoming signals to modify circuit function and behavior.
There are two types of eCBs, arachidonoylethanolamine (anandamide or AEA) and 2-arachidonoylglycerol (2-AG), that differ in their biosynthesis/degradation pathways and affinity for binding to the cannabinoid type 1 and 2 receptors (CB1 and CB2).2,17–21 2-AG is the most abundant and efficacious eCB ligand, while AEA is a lower efficacy ligand. Differences in desensitization between the two eCBs lead to differential time course of responses to agonist application, but desensitization appears to be too slow to contribute to the time course of effects on synaptic transmission.4,22,23 The duration of modulation by eCBs at synapses is most likely determined by enzymatic degradation and possibly by a poorly characterized reuptake mechanism.24–29 AEA can also directly act on multiple ionic channels at low concentrations, presumably in its tonic signaling state.13,30–33
Both AEA and 2AG appear to be synthesized on the postsynaptic side of the synapse. Indeed, the enzyme that catalyzes 2-AG synthesis, diacylglycerol lipase (DAGL), is found in the postsynaptic structure of many neurons.34–36 The main pathway for AEA synthesis in brain is not fully certain, but physiological studies support a postsynaptic role for synthesis and release.37,38 The eCB degradation pathways are segregated across the synapse, with the AEA-catabolic enzyme fatty acid amide hydrolase (FAAH) localized mainly in postsynaptic structures and the 2-AG-degrading enzyme monoacylglycerol lipase (MGL) located mainly on the presynaptic side.39,40 The differences in efficacy and the spatial segregation of the degradation pathways for the two eCBs support the hypothesis that AEA and 2-AG have differential functional roles in eCB mediated signaling in different brain regions or at different synapses within these brain regions (Figure 1).
Figure 1.

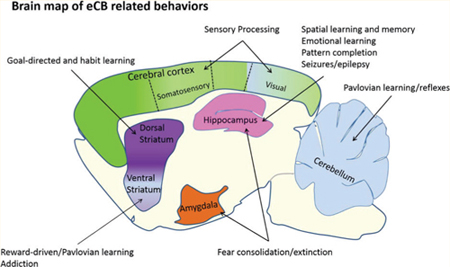
Proposed eCB mediated synaptic plasticity functional map in rodent brain. Schematic illustration of brain structures, specialized functions, and microcircuitry involved in eCB-LTD. eCBs have the potential to modulate goal-directed, habitual, and reward-driven behaviors in the striatum by DA regulation of eCB mediated LTD at excitatory synapses and eCB mediated LLD and LTD at inhibitory synapses. eCBs work in a layer specific manner at excitatory and inhibitory synapses in the somatosensory and visual cortices to process incoming sensory information. Hippocampal eCB synaptic depression has been reported at CCK-basket cell interneuron-pyramidal cell synapses and at excitatory terminals of Schaffer collaterals arising from CA3. There, eCB-mediated plasticity is postulated to play an important role in learning and memory formation, and can contribute to seizures and epileptic disorders. In the cerebellar cortex, eCBs mediate synaptic depression of Purkinje cell inputs to control motor learning and conditioning of reflexes. eCB mediated changes in synaptic transmission in the hippocampus and amygdala modulate fear-associated memory formation. DA, dopamine; FC, frontal cotex; OFC, obritofrontal cortex, MSN, medium spiny neuron; CIN, cholinergic interneuron; FSI, fast-spiking interneurons; glut, glutamatergic; PC, purkinje cell; PF, parallel fibers; CF, climbing fibers; GC, granule cell; CCK, cholecytokinin; SC, Schaffer collaterals; PV, parvalbumin positive neurons.
There are two modes of eCB signaling, tonic vs phasic, by which eCBs can regulate neurotransmitter release to affect network activity. In the hippocampus, AEA is thought to be the constitutive tonic eCB messenger, while 2-AG is considered the neuromodulatory phasic eCB messenger. However, it is yet to be determined if this pattern is observed in other brain regions. Indeed, it is likely that eCB signaling may not strictly adhere to one eCB type being tonic and the other phasic, but both AEA and 2-AG may switch between the different modes of signaling. Both eCBs can be produced in an activity dependent manner and may be differentially produced in different brain regions under different physiological conditions or pathophysiological states.23,38,41–43 Further evidence to support this hypothesis comes from experiments in which neuronal stimulation can selectively increase tissue levels of 2-AG but not AEA, even though both are present.41,44,45
As mentioned above, both eCBs bind to and activate CB1 and CB2 receptors. These are G-protein-coupled receptors (GPCRs) that upon activation signal through the Gαi/o family of heterotrimeric G-proteins.1,46 CB1 receptors are abundantly expressed in the CNS in brain regions associated with learning and memory, such as hippocampus, striatum, amygdala, and prefrontal cortex47,48 In these areas, CB1 receptors are highly expressed on glutamatergic and GABAergic (γ-aminobutyric acid) presynaptic terminals, made by both projection neurons and interneurons, making them ideally placed to fine-tune the regulation of synaptic efficacy and neuronal activity in response to incoming signals.47,49,50
The GABAergic interneurons that express CB1 in basolateral amygdala (BLA), hippocampus and neocortex are predominantly basket cells, but Schaffer collateral-associated interneurons in the CA1 hippocampal subfield also express CB1.49,51,52 One common feature of these CB1-expressing interneurons in hippocampus and other cortical regions is that they express and presumably release the cholecystokinin (CCK) neuropeptide (Figure 1). It is interesting to note that excitation-secretion coupling in the presynaptic terminals of CCK-positive interneurons in BLA and hippocampus requires the N-type voltage-gated calcium channel (Cav2.2), and inhibition of this channel appears to be the predominant mechanism through which CB1 inhibits GABA release at these synapses.53,54 The specificity of inhibition of release from this interneuron subtype appears to influence certain frequencies of activity within the hippocampus.55 The observation that this inhibition is relieved when CCK-positive neurons fire at higher frequencies indicates that CB1 may have a high-pass filtering function within this microcircuit.56 The functional implications of the specificity for synapses using N-type channels is not yet clear, but the channel subtype may contribute to the frequency-dependent effects of CB1 inhibition.
CB2 receptors are expressed mostly in the periphery,3,47,57,58 with very little expression in the CNS.59–61 The eCB system has been shown to be involved in both short and long-term decreases in synaptic transmission at glutamatergic and GABAergic synapses in many different brain regions.12,62 This review will discuss the functional implications of eCB-mediated synaptic modifications in behaviors involving different brain regions (Figure 1), by giving an overview of in vitro and limited in vivo studies in an attempt to link physiology and behavior.
FUNCTIONAL ROLE OF ENDOCANNABINOID MEDIATED DSE AND DSI
Activation of presynaptic CB1 receptors by the release of postsynaptic eCBs often begins with increases in postsynaptic intracellular calcium and/or activation of Gαq-coupled GPCRs. The eCB release can suppress synaptic transmission transiently for a minute or less. One such form of short-term plasticity is called depolarization-induced suppression of inhibition (DSI) or excitation (DSE) and can be induced by a brief (seconds) postsynaptic depolarization.5,63–70 This depolarization leads to increases in postsynaptic intracellular calcium via the activation of voltage-gated calcium channels to trigger eCB mobilization.5,24,43,65,67,68 eCB mobilization can also be triggered by activation of Gαq-coupled GPCRs such as the muscarinic acetylcholine receptors and group 1 metabotropic glutamate receptors to induce short-term depression (STD) and can further enhance the magnitude and expression of DSI and DSE.6,7,71 It is important to state that STD and DSI/DSE cooperatively interact and do not directly oppose each other. It is not clear which type of eCB is involved in short-term plasticity and whether the type of eCB varies with brain region. However, 2-AG appears to be the most likely candidate for involvement in DSI/DSE based on experiments showing that increased 2-AG signaling enhances these forms of plasticity.72–74 Cell-type specific deletion of 2-AG signaling reduces the magnitude of DSI and DSE in striatum, supporting a role for this eCB in this brain region.75 The transient suppression of synaptic transmission caused by STD or DSI/DSE may help to modulate neuronal activity by altering neuronal excitability and/or resetting neuronal responsiveness to incoming afferent information to influence behavior. To date, eCB mediated DSI/DSE has been observed in many brain regions, but it is best characterized in the hippocampus and cerebellum.76–79 It is likely that DSI/DSE has distinct physiological and behavioral functions depending on the synapses and brain region in which it is expressed.
Despite the implication of DSI/DSE and STD in neuronal modulation that affects circuit function, little is known about the occurrence of these forms of short-term plasticity in vivo. In fact, there are questions concerning the physiological relevance, if any, of these forms of transient synaptic plasticity.80 A computational model of presynaptic input firing predicts that the temporal summation of convergent inputs can produce a calcium dependent mobilization of eCB to result in DSI in vivo.81 Melis and colleagues, demonstrated that a brief afferent stimulus train, mimicking firing rates in vivo, can induce an eCB-mediated STD in vitro and more importantly in vivo in the ventral tegmental area (VTA), a brain region involved in reward learning.82 This dampening of neuronal activity could signal changes in reward information, thereby locally modulating neuronal activity to shape circuit function. Additionally, eCB mediated STD is augmented after a behavioral conditioning task, indicating that this can itself be modulated by changes in vivo as a result of environmental experience.83,84 Although there is no definitive evidence that DSI/DSE suppresses synaptic transmission in vivo, the aforementioned results demonstrate that it is possible for STD, lasting a few seconds, to sculpt neuronal firing rates to affect neural function. It is likely that these transient changes can encode alterations in reward value in real time during a behavioral task. Additionally, the fast dynamic switch between activation and inhibition at different synapses or the interruption of ongoing activity may help in action selection during a behavioral task. It is well-known that these forms of eCB mediated synaptic plasticity constitute a widespread phenomenon in the CNS.12,68 However, little research has been done to examine the contribution of eCB mediated shortterm plasticity to behavior. More research on identifying conditions and molecular mechanisms that promote STD in vivo is needed to fully understand the physiological and functional significance of eCB mediated transient synaptic plasticity.
FUNCTIONAL SIGNIFICANCE OF ENDOCANNABINOID MEDIATED LTD
In addition to eCBs mediating short-term synaptic plasticity, the eCB system can also produce longer-term changes in synaptic efficacy lasting hours to weeks that are referred to as long-term potentiation (LTP) and long-term depression (LTD). eCB-LTD is notably the best characterized and widespread form of eCB mediated long-term synaptic plasticity and can occur at both glutamatergic and GABAergic synapses throughout the brain.85–95 Endocannabinoid-dependent LTD can be induced by a transient increase in neuronal activity that leads to increases in calcium that result in the synthesis/release of eCBs from the postsynaptic cell to bind to presynaptic CB1 receptors.85,96,97 Depending on the brain region, eCB-LTD can be altered by neuromodulators, such as dopamine, which can influence cAMP signaling to facilitate LTD.42,90,98–101
eCB LTD can also occur in response to activation of metabotropic receptors, such as metabotropic glutamate type 5 (mGluR5) and serotonin type 2 (5-HT2) GPCRs that stimulate diacylglycerol formation leading to increased 2-AG production, and produce increased calcium release from intracellular stores to participate in mobilizing eCBs.6,71,90,102 LTD initiated by eCB activation of CB1 receptors persists long after CB1 receptor activation has ended.86,88,103–105 It is still unclear whether the same type of eCB (2-AG vs AEA) is involved in mediating the long-term changes in synaptic plasticity resulting from transient increases of afferent neuronal activity and those induced by postsynaptic activation of metabotropic receptors. AEA has been implicated as the eCB signaling molecule mediating eCB-LTD in the dorsal striatum (DS).38,106 Ade and Lovinger showed that during development, there is a switch from LTP to eCB-LTD.38 More interestingly, this switch in synaptic plasticity is accompanied by a developmental increase in AEA production levels.38 Also, it is not known how CB1 receptor activation can differentially mediate both short-term and long-term changes in synaptic plasticity. Chevaleyre and colleagues demonstrated that cAMP/PKA signaling and Rim 1α function downstream of CB1 receptor activation are needed for eCB-LTD and not STD.103 However, more research identifying the intracellular signaling components downstream of CB1 receptor activation is needed to fully understand the mechanistic difference between CB1 receptor-mediated STD and LTD. Long-term changes in synaptic plasticity at corticostriatal synapses are postulated to contribute to mechanisms of action control, such as skill and instrumental learning (Figure 1). In fact, aberrant corticostriatal synaptic plasticity has been implicated in numerous movement disorders, including Parkinson’s and Huntington’s diseases.98,107–109
GOAL-DIRECTED/HABITUAL BEHAVIORS
Prefrontal Cortex-Striatal eCB-LTD.
The prefrontal cortex (PFC) refers to cortical subregions that sit at the anterior of the frontal cortex. These cortices send glutamatergic afferents to a variety of subcortical regions including BLA, other allocortical and neocortical areas, nucleus accumbens (NAc) and dorsomedial striatum (DMS).110,111 The prefrontal cortices are generally thought to have roles in such advanced cognitive functions as cognitive and behavioral flexibility, decision making, executive function, and perhaps even consciousness.112–115 However, more specific functions of these cortices and their projections include roles in reversal learning and extinction of fear conditioning.116–118 There are ongoing debates as to whether rodents possess true prefrontal cortical areas, but it is clear that many regions of mouse and rat frontal cortex have connectivity similar to that of primate PFC, and there is some evidence that the functions of these areas within cortical-subcortical circuits are similar across species. For the purposes of this review, we will focus on eCB actions on neurons that are part of 3 PFC regions where these neuromodulatory actions are well characterized, the infralimbic and prelimbic medial prefrontal areas (ILC, PLC respectively, often referred to as mPFC when not explicitly differentiated) and the orbitofrontal cortex (OFC). The CB1 receptor has been localized to neurons in the PFC, with especially prominent mRNA expression in ILC, PLC, and OFC (Allen Brain Atlas; http://www.brain-map.org/).49,119,120 Receptors have been localized to presynaptic terminals of both GABAergic and glutamatergic neurons within these PFC subregions,121,122 and the receptor is also present on terminals of glutamatergic neurons that project from PFC to subcortical regions (Figure 1).123–125 Thus, the eCB signaling system is well positioned to influence processing within PFC as well as PFC communication and control of other brain regions.
Some of the initial evidence for eCB and CB1 receptormediated synaptic plasticity came from electrophysiological experiments in medial PFC slices. The work of Crepel and colleagues showed that a CB1 receptor agonist inhibited glutamatergic transmission at synapses onto layer V PLC projection neurons via an apparent presynaptic mechanism.126 A CB1 receptor inverse agonist increased glutamatergic transmission, revealing some of the first evidence for tonic receptor activity that depresses transmitter release. These investigators also found that activation of CB1 receptors increased the proportion of neurons showing HFS-induced LTD, while receptor antagonism favored LTP. In hindsight, these data clearly presage the discovery of eCB-LTD a few years later, although at the time these investigators did not distinguish between a modulatory role for eCBs and CB1 and a central role in the LTD induction mechanism. Subsequent studies have examined eCB-LTD at other synapses in the PFC, as well as the roles of eCBs, CB1 and LTD in a variety of PFC functions. At inhibitory synapses onto layer V PFC pyramidal neurons, eCBs contribute to LTD of inhibitory synapses (iLTD) that requires activation of a group I mGluR and is modulated by dopamine via dopamine D2 receptors (D2Rs).127 This dopaminergic modulation is intriguing as the CB1 and D2Rs are situated in close proximity to one another on the GABAergic terminals that show eCB-mediated plasticity. The GABAergic neuronal subtypes that express these receptors are not yet clear, but it is probable that these neurons include those that make perisomatic synapses and are similar to CB1-receptor expressing basket cells in other cortical regions.
Endocannabinoids and CB1 receptors expressed by OFC projection neurons appear to have important roles in stimulus-response (S-R) or “habit learning” (Figure 1).125 Experiments in the Costa laboratory first found a role for eCBs/CB1 receptors in this type of learning.128 Further investigation of this role revealed that knocking out CB1 receptors in mouse OFC neurons prevented S-R learning, and this effect was also observed when the knockout was restricted to glutamatergic projection neurons.125 Using a retrograde viral-based strategy in the latter experiments, these investigators also showed that CB1 receptors inhibit glutamatergic synapses from OFC projection neurons onto medium spiny neurons (MSNs) in the DMS, and that this modulation was lost in the projection neuron-specific CB1 receptor KO condition. These findings provide some of the most direct evidence of an effect of CB1 on a specific corticostriatal synapse. Mimicking the effect of presynaptic CB1 with Gi/o-DREADD expression/activation at OFC-DMS synapses fostered S-R learning, including experiments in which the DREADD agonist was applied locally within DMS. It is postulated that presynaptic suppression of OFC inputs fosters S-R learning, most likely by inhibition of OFC-derived information about outcome value status that normally supports competing action-outcome or “goal-directed” instrumental performance.
It must be noted that CB1 receptors on OFC-DMS presynaptic terminals may not be the only such receptors involved in S-R learning. Examination of the mechanisms underlying promotion of S-R learning after chronic Δ9-tetrahydrocannabinol (THC) treatment indicated that receptors and eCB-LTD in the dorsolateral striatum also have important roles in this learning process.129 This study puts forth the hypothesis that bidirectional synaptic plasticity, allowing for synaptic flexibility, is important for selecting the appropriate goal-directed or habitual behaviors. Thus, there may be several cellular and brain regional loci at which the eCB/CB1 system contributes to instrumental learning and environmental and drug effects on this learning.
There is a growing literature describing effects of drugs of abuse and other environmental manipulations of eCB synaptic actions in the PFC, as well as on goal-directed behaviors (Figure 1). Exposing rats to repeated in vivo THC administration in adult mouse leads to a loss of eCB-LTD at glutamatergic synapses between PFC and NAc.105 The loss of eCB-LTD is also observed following exposure to THC during adolescence in female rats, and this is accompanied by altered maturation of several molecules involved in eCB signaling and other forms of synaptic plasticity.130 Many of these changes persist into adulthood. Adolescent exposure to CB1 receptor agonist also produces lasting impairment in other aspects of PFC function, and the performance of cognitive tasks that involve the PFC.131–134 In vivo exposure to cocaine impairs eCB-LTD at glutamatergic synapse in PLC,135 and even a single in vivo injection of this drug can impair induction of eCB-LTD at PFC synapses in the NAc.136 Endocannabinoid actions in the PFC may also have roles in psychiatric disorders.137–139 Drugs of abuse interfere with bidirectional plasticity at certain synapses, resulting in the inability to properly update normal patterns of plasticity (i.e., LTD and LTP) in key circuits in response to incoming sensory stimuli (Figure 1). More detailed reviews on the role of eCBs in addiction and addiction-related behaviors can be found elsewhere.140–142
Prefrontal Cortex-Striatal eCB Modulated DA Release.
Recently it has been shown that CB1 receptors on terminals of mPFC-NAc projections modulate cholinergically driven DA release in vitro and in vivo.143 This eCB-mPFC modulation of DA signaling can be mediated by either 2-AG or AEA This study also provided evidence that “on-demand” activation of 2-AG-CB1 receptor signaling suppresses DA release under conditions where this release is driven by brief bursts of stimulation of NAc cholinergic interneurons. The cellular source of the 2-AG produced by this stimulation is not yet clear. The NAc is perfectly positioned to integrate excitatory signals from PFC, hippocampus, and amygdala, as well as dopaminergic signals from VTA to affect downstream output circuitry to control motivated goal-directed behavior.144–146 Indeed, eCBs and CB1 receptors in mPFC and NAc can modulate goal directed behavior in a drug reinstatement paradigm.147 It is believed that maladaptive long-term changes at these inputs contribute to addictive behaviors.148,149 Previously it has been shown that mPFC-NAc terminal activation can promote self-stimulation,150 indicating a role for this projection in reward-driven behavior. Recently published data have extended these findings by demonstrating that CB1 receptor modulation of mPFC-NAc afferents can alter this self-stimulation.143 Mateo and colleagues showed that 2AG signaling via CB1 receptor activation reduces mPFC-NAc self-stimulation in an instrumental task.143 One possible mechanism is that activation of CB1 receptors located on mPFC-NAc terminals decreases glutamate release either through STD or LTD, thereby decreasing cholinergically driven DA release in the NAc to modulate reward-driven instrumental responding. Given that mPFC-NAc self-stimulation responding is elevated on the first day of training, eCB-LTD may not be needed for the initial goal-directed learning process, but may be needed for the maintenance of mPFC-NAc responding. The role of habit processes in this self-stimulation paradigm is not yet known, but 2-AG and CB1 receptors could also play a role in the development of habitual responding.
Parkinson’s Disease: Corticostriatal eCB-LTD.
Parkinson’s disease (PD) is a progressive neurodegenerative disorder that is characterized by profound motor dysfunction151–153arising from the gradual loss of DA neurons in substantia nigra pars compacta (SNc) that project to the DS, an area important for movement control, goal directed and habitual behaviors.154,155 The loss of DA receptor activation leads to changes in corticostriatal plasticity that affect basal ganglia network function.108,156 Specifically, corticostriatal LTD is lost after in vivo DA depletion when measured in vitro98,157,158 or in vivo.159 This form of LTD is dependent on eCB signaling85,99,108 and DA-D2R activation98,99,157,158,160 The site of the DA-D2Rs involved in this LTD is still under debate. Some pharmacological studies have implicated striatal cholinergic interneurons,99,160 while others have implicated DA-D2R-containing MSNs.101,156,161 Nonetheless, DA-D2R activation enhances AEA eCB release in striatum,42 and AEA has been postulated as the eCB signaling molecule involved in corticostriatal LTD.38,106,108 Flowever, a role for 2-AG in striatal LTD has not been ruled out by any means. Other neurotransmitters and receptors, including the A2A adenosine receptor,160,161 have also been implicated in the modulation of striatal eCB-LTD, but the precise mechanism of action of these receptors remains unclear.
Following DA depletion, there is an enhancement of eCB levels162,163 that is associated with the inhibition of movement.163 Both corticostriatal LTD and the effects of DA depletion on movement can be restored by DA receptor activation and the enhancement of eCB signaling.42,108,156,158,164,90 This tightly orchestrated activation of DA receptors and the appropriate levels of eCB signaling are necessary for the expression of corticostriatal LTD160 and proper voluntary movement.108 Although the loss eCB-LTD has been the central focus of corticostriatal dysfunction following DA-depletion, there are also alterations in the expression and induction of LTP. Depending on the degree of DA depletion, corticostriatal LTP is lost158,165 or aberrant LTP can be induced using stimulation paradigms that normally induce eCB-LTD.98,108,156 More interestingly, bidirectional plasticity at corticostriatal synapses is lost, leaving only unidirectional changes in corticostriatal efficacy following DA depletion.156,166 These findings suggest that the DA depletioninduced loss of synaptic flexibility contributes to the motor deficits in PD and bidirectional synaptic modification is critical for proper movement control.
GABAergic-STRIATAL eCB MEDIATED LONG-LASTING DISINHIBITION
Disinhibition is a mechanism through which eCB activation of CB1 receptors can induce a long-lasting increase in glutamatergic activation of striatal neurons. Low-moderate frequency intrastriatal electrical stimulation induces a longlasting decrease in GABAergic transmission at synapses onto MSNs.95,167,168 When the synaptically driven massed firing of striatal neurons is measured in field potential recordings (i.e., the population spike, PS, driven by synaptic activation), this low-moderate-frequency stimulation induces a long-lasting increase in PS amplitude that persists after the stimulus protocol. This increase is not observed when GABAa receptors are blocked, indicating a role for fast inhibitory synaptic transmission in this form of plasticity. Thus, this synaptic change has been termed long-lasting disinhibition (DLL; Figure 1).167 Antagonists of CB1 receptors block this DLL, and the underlying LTD of GABAergic synaptic transmission is prevented by knocking out CB1 receptors in striatal MSNs and fast-spiking GABAergic interneurons.168 Two forms of eCB-LTD have been observed at GABAergic synapses onto striatal MSNs.168 Differences in postsynaptic membrane potential, eCB subtype, and presynaptic neuronal subtype determine which type of iLTD is expressed.168 Thus, there may be circumstances where the underlying eCB-mediated processes contributing to DLL differ, and it remains to be determined when these different forms of plasticity might occur in vivo.
Exposure to ethanol (EtOH) alters DLL induction and expression. In rats that consumed EtOH intermittently for 7 weeks DLL can no longer be induced by moderate-frequency electrical stimulation.167 The ability of a CB1 receptor agonist to produce disinhibition was also impaired in striatum from these EtOH-consuming rats. The role in EtOH-related behaviors of CB1 receptors on different striatal GABAergic neurons and the iLTD and DLL processes needs to be explored.
ASSOCIATIVE MEMORY FORMATION
Medial Prefrontal Cortex-Amygdala eCB-LTD.
The role of eCBs/CB1 on mPFC-amygdala projections in synaptic plasticity and learning and memory has also been examined (Figure 1). It is well-known that glutamatergic projections from mPFC to the BLA influence the extinction of aversive memories.118,169–171 The hippocampus also plays a role in the associative processing in fear conditioning paradigms.172 The CB1 receptors in mPFC itself also appear to have roles in fear consolidation and extinction, and activation of these receptors can impair retrieval of these memories in the fearpotentiated startle paradigm.173 Several studies have implicated AEA signaling in the extinction of aversive memories.174–177 Disruption of eCB/CB1 receptor signaling is selective for the extinction of aversive memories because extinction of non-aversive memories in an appetitive behavioral paradigm is not impaired by CB1 antagonism.178 The extinction of fear memories or relearning of “new” memories that overrides the previous “old” memories may be mediated by eCB-LTD signaling. In the amygdala, eCB-LTD has been reported at both the inhibitory and excitatory terminals.123,179–181 Emerging evidence suggests that DSI is not the mechanism through which eCBs and CB1 receptor participate in fear memory extinction. Réintroduction of CB1 receptors in forebrain GABAergic neurons in mice that lack CB1 receptors restored DSI in the hippocampus and amygdala, but did not restore fear extinction.182 The authors report only “marginally” improved extinction of fear conditioning after the genetic restoration of CB1 receptors. One possible mechanism is that CB1 receptor activation results in eCB-LTD at the inhibitory synapses that may result in the disinhibition of LTP at excitatory synapses to facilitate associative learning.88,89,183 eCB-LTD can be spatially restricted to reduce synaptic transmission at selected presynaptic terminals, such that disinhibition can occur if effects are restricted to GABAergic synapses.88,183 Although this has not been shown to occur in the amygdala, this metaplasticity has been shown to occur in the hippocampus, as discussed later in this review. With the advancement of genetic techniques, researcher can begin to probe synapsespecific eCB-LTD and its role in behavior. These new avenues of research will undoubtedly deepen our understanding of the physiological and functional significance of this form of synaptic plasticity and others.
SENSORY PROCESSING
Somatosensory Cortex eCB-LTD.
The roles of eCBs and CB1 receptors in somatosensory cortex have been explored mainly in relation to the development of the whisker barrel cortex (Figure 1). The whisker barrel area is a rather large part of the rodent somatosensory cortex devoted to the receipt and processing of tactile information from the facial whiskers.184 This system is used for perception of environmental information of great value to animals that often navigate tight spaces in low-light conditions. This whisker barrel system is also well-known for its ability to adapt to changing sensory information, and several types of synaptic plasticity occur at synapses in different whisker barrel cortical layers.185–190
The postnatal development of whisker barrels and their physiological responses has been characterized in detail, and plasticity in these responses can be seen following trimming of whiskers during a certain developmental time period.187,191,192 This synaptic plasticity appears to have “organizational” effects that determine subsequent synaptic interactions and physiological responses within the circuit. Among the forms of synaptic plasticity observed at glutamatergic whisker barrel synapses is a form of LTD mediated by eCBs and CB1 receptors. At synapses made by layer 4 glutamatergic neurons onto neurons in layer 2/3 of barrel cortex, evidence of LTD after whisker trimming has been observed in brain slices from rats examined at postnatal day 12.193 This LTD has an apparent presynaptic locus of expression, and depends on retrograde eCB signaling.194 One physiological role for this layer 2/3 eCB-LTD would be to reduce responsiveness to unimportant sensory stimuli during an important period of cortical development.195 This idea is supported by the observation that blockade of CB1 receptor signaling leads to inappropriate whisker tuning in L2/3 neurons and blurring of the whisker map.196 Thus, eCB-mediated plasticity is important in the early period of somatosensory cortical physiological shaping and later responsiveness to the environment.
Both GABAergic and glutamatergic synapses onto somatosensory cortex neurons are modulated by eCBs and CB1 receptors in young and young adult rodents.197,198 This modulation occurs at synapses onto pyramidal neurons in layers 2/3 and 5, and not at synapses onto interneurons.199 Short-term depression has been the main type of plasticity investigated at these synapses. Interactions between eCBs and brain-derived neurotrophic factor (BDNF) have also been described in somatosensory cortex.200,201 One such interaction involves BDNF effects on layer 2/3 pyramidal neurons leading to endocannabinoid production and release.200,202 This stimulation involves activation of the TrkB neurotrophin receptor. This mechanism contributes to LTD induction at GABAergic synapses onto layer 2/3 pyramidal neurons, triggered by theta burst stimulation,202 and 2-AG appears to be the eCB that mediates this LTD. At present there is very little information about how eCB-dependent plasticity in the different layers alters sensory processing mediated by this cortical area.
Endocannabinoids and CB1 receptors also participate in long-lasting “self-modulation” of layer V low-threshold spiking inhibitory interneurons and layer 2/3 pyramidal neurons in somatosensory cortex.203–205 While this physiological change is not strictly speaking LTD, it will have a long-lasting influence on cortical function. This modulation is seen as a long-lasting hyperpolarization following induction of a brief burst of action potentials during recordings from these neurons. Pharmacological experiments indicate that the hyperpolarization involves tonic CB1 receptor activation leading to increased potassium current in the somatodendritic compartment of these neurons. The effect can be mimicked by CB1 receptor agonist application. It appears that 2-AG mediates this selfinhibition.204 The net effect of the long-lasting self-inhibition of low-threshold spiking neurons is to provide a window during which their strong inhibition of pyramidal neurons is relieved, permitting enhanced glutamatergic cortical output,203 while the effect in a subset of pyramidal neurons may inhibit corticocortical transmission.205
Visual Cortex eCB-LTD.
The eCBs and CB1 receptors also have important organizational roles in development of visual cortex (VC), similar to their actions in the whisker barrel system (Figure 1).206 As in that somatosensory system, several types of synaptic plasticity in VC are involved in system development, and altering sensory input early in development alters subsequent visually driven physiology.207 The maturation and function of VC is often assessed by examining the response of neurons in different cortical layers to light presented to one or the other eye, the well-known ocular dominance columns.207,208 The column pattern is altered by visual deprivation to a single eye, as synapses responding to input from the two eyes appear to compete for influence on the VC neurons. This pattern of deprivation-induced plasticity in layer 2/3 VC is altered by CB1 receptor antagonist application in young animals, but this treatment does not appear to alter plasticity in layer 4.208 Endocannabinoid-dependent LTD occurs at glutamatergic and GABAergic synapses in VC, especially in layer 2/3.209–211 Prior monocular deprivation occludes LTD induction at glutamatergic layer 2/3 synapses.209 The iLTD is especially prominent during the “critical period” when effects of visual deprivation are maximal. This developmental stage-dependent iLTD underlies a visual experience-dependent decrease in release probability at GABAergic synapses that helps set the final efficacy for mature synapses.210 Keeping mice in the dark between eye opening and the onset of puberty also alters aspects of VC development, including GABA release at synapses in layer 2/3.212 This effect is mimicked by knocking out CB1 receptors, and alterations in plasticity within layer 5 are also observed.212 These findings indicate that eCB-LTD has prominent roles in the physiology of developing visual cortex, but the consequences for adult visual function are not so clear. The status of eCB-LTD in adult visual cortex and roles in visual function are also still unclear.
Cerebellar eCB-LTD.
The LTD observed at granule cell/ parallel fiber (PF) synapses onto Purkinje neurons in cerebellar cortex was arguably the first form of LTD to be described in the CNS. Consequently, much is known about the mechanisms of this “cerebellar LTD” and its role in cerebellar-based forms of learning and memory including pavlovian conditioning and adaptation of reflexes.213 Only recently was any role for eCBs in this LTD suggested. Work in the Regehr laboratory indicated that cerebellar LTD is prevented by a CB1 receptor antagonist and by inhibition of DAG lipase,214 and cannot be induced in mice lacking CB1 receptors specifically in cerebellar granule cells.215 This eCB/CB1 role was initially quite surprising given that the bulk of evidence indicates that cerebellar LTD involves a postsynaptic expression mechanism with no indication of a presynaptic role.213 While this apparent conundrum has not been fully resolved, the eCB/CB1 system may help to regulate the timing of LTD induction by allowing for optimal suppression of parallel fiber inputs by subsequent activation of climbing fiber inputs.216 Antagonism of CB1 receptors also prevents LTD at PF synapses induced by moderate frequency PF stimulation in brain slices and in vivo,217,218 and there is evidence that presynaptic mechanisms are involved in expression of this type of LTD.217 A presynaptically expressed form of eCB-LTD is also observed at PF synapses onto stellate interneurons.91 Interestingly, there is evidence for an eCB role in cerebellum-dependent eyeblink conditioning,219 but it will be interesting to examine the roles of this signaling system in other forms of cerebellum-dependent learning and memory (Figure 1).
HIPPOCAMPAL ENDOCANNABINOIDS, LEARNING AND MEMORY, SEIZURES, AND EPILEPSY
The hippocampus has well-known roles in different types of explicit learning, including spatial learning.220,221 In addition, eCB/CB1-mediated synaptic plasticity and metaplasticity are well characterized in the hippocampus, especially at GABAergic synapses (Figure 1)88,183 There is also extensive literature describing cannabinoid effects on learning and memory involving the hippocampus.222–224 There is a growing body of information about how hippocampal eCBs and CB1 contribute to learning and memory. Intrahippocampal infusion of AM251 impairs consolidation of fear memory.225 Interestingly, CB1 antagonist treatment can enhance acquisition of trace fear conditioning (a learning process known to involve the hippocampus),226 and antagonists can enhance performance on spatial learning tasks.227–229 Endocannabinoids also appear to contribute to spatial memory retrieval, depending on the level of emotional arousal.230
The explicit relationship between eCB-mediated plasticity and hippocampal-based learning and memory has been the subject of several recent studies. In mice lacking MGL, LTP induced by theta-burst stimulation at glutamatergic synapses in the CA1 subregion is enhanced.29 These mice also show prolonged DSI that may underlie the facilitation of LTP. In spatial and object recognition tasks, these MGL knockout mice show improved performance, consistent with the idea that disinhibitory eCB-dependent synaptic plasticity can facilitate both LTP and learning. Using a trace-conditioning task, Zhu and co-workers showed that mice with deficiency in an eCB-mediated form of metaplasticity involving iLTD and enhanced LTP perform poorly.231 Enhancing eCB signaling with an MGL inhibitor enhanced acquisition and performance on this temporally based associative learning task. This enhancement was associated with changes in iLTD indicative of in vivo metaplastitity. In contrast, strong inhibition of MGL impairs LTP in the hippocampal CA1 region, and forms of learning involving the hippocampus.232 The different effects of the MGL inhibitors in these two studies may reflect the task used or the efficacy of the inhibitor in enhancing hippocampal 2-AG levels. Impairment of LTP and hippocampal-based learning and memory was also observed with a FAAH inhibitor.233 The effects of experimental manipulations that alter eCBs and CB1 may also depend on the extent to which a given manipulation alters eCB suppression of GABAergic transmission,88 which would favor LTP and learning, versus inhibiting glutamatergic transmission, which might impair LTP.41
Endocannabinoid and CB receptors roles in brain hyperexcitability, seizures, and epilepsy have been investigated since the discovery of these biomolecules. Effects of eCBs on GABAergic and glutamatergic synapses in brain regions with key roles in limbic and tonic-clonic seizures can alter the balance of synaptic excitation and inhibition in ways that may promote or reduce seizure susceptibility. This topic was recently reviewed in some depth,55,234,235 and thus, only a brief summary is presented here. Alterations in eCB signaling in relation to epilepsy are best characterized in the hippocampal formation, as this brain region has important roles in limbic seizures (Figure 1). A temporal pattern of changes in tissue eCB levels and CB1 expression has been observed in epilepsy models. Downregulation of CB1 expression is generally observed throughout the hippocampal formation in the acute seizure phase, followed by upregulation in the chronic epileptic phase,55,234,236 but changes in receptor levels and function also vary across hippocampal subregions in some seizure models.237 Both 2-AG and AEA levels are increased in the acute phases of pilocarpine and kainic acid-induced seizures, but there is not as much information on eCB levels in the chronic phase.55,238 Retrograde signaling by eCBs at GABAergic hippocampal synapses is also increased in animal models of seizure disorders.239 This enhanced modulation likely results in enhanced excitability in this brain region.
Increased CB1 expression on GABAergic terminals is also observed in post-mortem tissue from human epilepsy patients, although downregulation was also observed on glutamatergic terminals.240,241 Examination of the living human brain using positron emission tomography imaging with a CB1 ligand has revealed increased CB1 binding in the temporal lobe ipsilateral to a seizure focus in patients with temporal lobe epilepsy and hippocampal sclerosis, while binding was decreased bilaterally in the insular cortex.242 However, it is not clear if these changes are due to alterations in receptor expression or differences in eCB occupancy of the receptors during the imaging period. It is interesting in this context that AEA levels are reduced in cerebrospinal fluid of temporal lobe epilepsy patients.243 Overall, these findings indicate that the eCB/CB1 signaling system is altered by seizures and in epileptic disorders, but more information is needed to determine the role of eCBmediated synaptic plasticity in the development of these disorders.
There is also ample evidence that altering eCB/CB1 signaling affects seizures and epilepsy in animal models, but the effects of experimental manipulations of this system have been mixed. Peripheral administration of efficacious CB1 agonists generally has an anticonvulsant effect in a variety of animal seizure models, although seizure-enhancing effects have also been observed.244–246 When CB1 antagonists are administered peripherally, they produce mainly pro-convulsant effects in these models.238,247–250 However, antagonists can prevent development of experimental febrile or head-injuryinduced seizures especially when given early in the development of epilepsy.251,252 Deleting CB1 receptors from glutamatergic forebrain synapses enhances kainic acid induced convulsions, consistent with a role for excess synaptic excitation due to loss of eCB modulation.253 While compounds targeted at the eCB/CB1 signaling system clearly affect seizures and epileptic disorders, therapeutic use of any such compounds will require a more precise understanding of the interactions with different circuits involved in different seizure types and forms of epilepsy.
The finding that cannabidiol, a phytocannabinoid that does not interact with CB1 receptors, has some efficacy for reduction of seizures in animal models and in certain childhood epilepsies needs to be mentioned in the context of this discussion.235,254,255 However, it must also be noted that there is no strong evidence that this action involves eCBs or CB receptors.
FUNCTIONAL RELEVANCE OF ENDOCANNABINOID LTP
In addition to well characterized eCB and CB1 receptor roles in LTD, participation of these molecules in LTP has also been postulated.95,256,257 Using a spike-timing-dependent plasticity (STDP) procedure with a low number of pre- and postsynaptic spike pairings, Cui and co-workers observed a form of LTP at glutamatergic synapses onto dorsal striatal MSNs that is blocked by inhibition of eCB production and a CB1 receptor inverse agonist, and is lost in CB1 receptor KO mice.256 A role for TRPV1 in this STDP-LTP has also been proposed based on antagonist actions, but it would be helpful to determine if this form of plasticity is lost in animals lacking TRPV1. A role for dopamine in this STDP-LTP has also been postulated based on blockage by a D2 receptor antagonist or a combination of D1 and D2 receptor antagonists. The locus of the mechanisms responsible for this increased synaptic transmission appears to be presynaptic. Interestingly, both eCB-LTD and LTP can be induced in the same neuron by altering the number of pre- and postsynaptic spike pairs. A form of LTP involving eCBs and CB1 receptors has also been reported in the hippocampal CA1 region.258 This eCB-LTP requires group I mGluR activation, and appears to involve retrograde 2-AG signaling resulting in presynaptic facilitation of glutamate release specific to Schaffer collateral synapses.
A form of LTP that requires CB1 receptors has also been observed in whisker barrel cortex layer V pyramidal neurons of young rats.259 Activation of blood flow in the whisker barrel cortex of adult rat by whisker stimulation is potentiated by CB1 receptor agonist administration, but there are mixed effects of a CB1 receptor inverse agonist depending on the route of peripheral administration that remain unexplained.260,261 This type of plasticity may provide a mechanism for strengthening synapses during relatively short windows of intense combined afferent and postsynaptic coactivation, such as might take place in associative learning tasks. However, it remains to be determined if eCB-LTP occurs in vivo.
CONCLUSIONS
In the last two decades, the eCB system has emerged as a key regulator of synaptic function. Short-term and long-term forms of eCB mediated synaptic plasticity have been reported in many different brain regions at both inhibitory and excitatory synapses. While there is ample evidence that altering eCB-CB1 signaling affects many behaviors, to date, there is no direct evidence that eCB mediated LTD or LTP in vivo contributes to these behaviors per se. Three general circuit mechanisms emerge again and again for eCB mediated LTD and LTP that may influence behaviors. Endocannabinoid LTD can participate in two of these circuit effects. One mechanism is the net disinhibition of local circuits, in which inhibition of GABAergic synapses allows for enhanced synaptic output at a subset of synapses. On the other hand, eCB-LTD can exert its inhibitory actions on neural circuits on a micro and macro level to suppress unwanted neuronal activity. Lastly, eCB-LTP can strengthen synapses to reinforce or maintain a certain behavior. These complex changes in potentiation, inhibition, and disinhibition in synaptic output are delicately modulated by eCBs to help shape synaptic and behavioral function. Understanding how these mechanisms contribute to behavior will require determining if and when these types of plasticity take place in vivo during behavioral performance or learning and memory. Preventing this plasticity in a temporally and spatially specific manner can then be used to determine if eCB-mediated long-lasting synaptic plasticity has a crucial role in these in vivo processes. With the powerful new approaches available to the modern neurobiologist it should now be possible to perform such experiments. Ultimately, this line of research will produce a better understanding of mechanisms involved in learning, memory, effects of drugs of abuse, and addiction.
Funding
This work was supported by The Division of Intramural Clinical and Biological Research of the National Institute on Alcohol Abuse and Alcoholism, project # ZIA AA000416 12.
ABBREVIATIONS
- 2-AG
2-arachidonoylglycerol
- AEA
arachidonoylethanolamine
- DSE
depolarization-induced suppression of excitation
- LTD
long-term depression
- LTP
long-term potentiation
Footnotes
Notes
The authors declare no competing financial interest.
REFERENCES
- (1).Matsuda LA, Lolait SJ, Brownstein M. j., Young AC, and Bonner TI (1990) Structure of a cannabinoid receptor and functional expression of the cloned cDNA Nature 346 (6284), 561–4. [DOI] [PubMed] [Google Scholar]
- (2).Devane WA, Hanus L, Breuer A, Pertwee RG., Stevenson LA, Griffin G, Gibson D, Mandelbaum A, Etinger A, and Mechoulam R (1992) Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 258 (5090), 1946–9. [DOI] [PubMed] [Google Scholar]
- (3).Munro S, Thomas KL., and Abu-Shaar M (1993) Molecular characterization of a peripheral receptor for cannabinoids. Nature 365 (6441), 61–5. [DOI] [PubMed] [Google Scholar]
- (4).Sugiura T, Kondo S, Sukagawa A, Nakane S, Shinoda A, Itoh K, Yamashita A, and Waku K (1995) 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun 215 (1), 89–97. [DOI] [PubMed] [Google Scholar]
- (5).Kreitzer AC, and Regehr WG (2001) Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto Purkinje cells. Neuron 29 (3), 717–27. [DOI] [PubMed] [Google Scholar]
- (6).Varma N, Carlson GC, Ledent C, and Alger BE (2001) Metabotropic glutamate receptors drive the endocannabinoid system in hippocampus. J. Neurosci. 21 (24), RC188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (7).Kim J, Isokawa M, Ledent C, and Alger BE (2002) Activation of muscarinic acetylcholine receptors enhances the release of endogenous cannabinoids in the hippocampus. J. Neurosci 22 (23), 10182–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (8).Edwards DA, Kim J, and Alger BE (2006) Multiple mechanisms of endocannabinoid response initiation in hippocampus. J. Neurophysiol 95 (1), 67–75. [DOI] [PubMed] [Google Scholar]
- (9).Min R, Di Marzo V, and Mansvelder HD (2010) DAG lipase involvement in depolarization-induced suppression of inhibition: does endocannabinoid biosynthesis always meet the demand? Neuroscientist 16 (6), 608–13. [DOI] [PubMed] [Google Scholar]
- (10).Alger BE (2002) Retrograde signaling in the regulation of synaptic transmission: focus on endocannabinoids. Prog. Neurobiol 68 (4), 247–86. [DOI] [PubMed] [Google Scholar]
- (11).Katona I, and Freund TF (2012) Multiple functions of endocannabinoid signaling in the brain. Annu. Rev. Neurosci 35, 529–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (12).Kano M, Ohno-Shosaku T, Hashimotodani Y, Uchigashima M, and Watanabe M (2009) Endocannabinoid-mediated control of synaptic transmission. Physiol. Rev 89 (1), 309–80. [DOI] [PubMed] [Google Scholar]
- (13).Venance L, Piomelli D, Glowinski J, and Glaume C (1995) Inhibition by anandamide of gap junctions and intercellular calcium signalling in striatal astrocytes. Nature 376 (6541), 590–4. [DOI] [PubMed] [Google Scholar]
- (14).Grueter BA, Brasnjo G, and Malenka RC (2010) Postsynaptic TRPV1 triggers cell type-specific long-term depression in the nucleus accumbens. Nat. Neurosci 13 (12), 1519–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (15).Chavez AE, Chiu CQ, and Castillo PE (2010) TRPV1 activation by endogenous anandamide triggers postsynaptic long-term depression in dentate gyrus. Nat. Neurosci 13 (12), 1511–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (16).Puente N, Cui Y, Lassalle O, Lafourcade M, Georges F, Venance L, Grandes P, and Manzoni OJ (2011) Polymodal activation of the endocannabinoid system in the extended amygdala. Nat. Neurosci 14 (12), 1542–7. [DOI] [PubMed] [Google Scholar]
- (17).Cravatt BF, Giang DK, Mayfield SP, Boger DL, Lerner RA, and Gilula NB (1996) Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 384 (6604), 83–7. [DOI] [PubMed] [Google Scholar]
- (18).Dinh TP, Carpenter D, Leslie FM, Freund TF, Katona I, Sensi SL, Kathuria S, and Piomelli D (2002) Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc. Natl. Acad. Sci. U. S. A 99 (16), 10819–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (19).Sugiura T, Kobayashi Y, Oka S, and Waku K (2002) Biosynthesis and degradation of anandamide and 2-arachidonoylglycerol and their possible physiological significance. Prostaglandins, Leukotrienes Essent. Fatty Acids 66 (2–3), 173–92. [DOI] [PubMed] [Google Scholar]
- (20).Grotenhermen F (2005) Cannabinoids. Curr. Drug Targets: CNS Neurol. Disord 4 (5), 507–30. [DOI] [PubMed] [Google Scholar]
- (21).Wang J, and Ueda N (2009) Biology of endocannabinoid synthesis system. Prostaglandins Other Lipid Mediators 89 (3–4), 112–9. [DOI] [PubMed] [Google Scholar]
- (22).Luk T, Jin W, Zvonok A, Lu D, Lin XZ, Chavkin C, Makriyannis A, and Mackie K (2004) Identification of a potent and highly efficacious, yet slowly desensitizing CB1 cannabinoid receptor agonist. Br. J. Pharmacol 142 (3), 495–500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (23).Kim J, and Alger BE (2010) Reduction in endocannabinoid tone is a homeostatic mechanism for specific inhibitory synapses. Nat. Neurosci 13 (5), 592–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (24).Kim J, and Alger BE (2004) Inhibition of cyclooxygenase-2 potentiates retrograde endocannabinoid effects in hippocampus. Nat. Neurosci 7 (7), 697–8. [DOI] [PubMed] [Google Scholar]
- (25).Fowler CJ (2007) The contribution of cyclooxygenase-2 to endocannabinoid metabolism and action. British journal of pharmacology 152 (5), 594–601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (26).Straiker A, Hu SS, Long JZ, Arnold A, Wager-Miller J, Cravatt BF, and Mackie K (2009) Monoacylglycerol lipase limits the duration of endocannabinoid-mediated depolarization-induced suppression of excitation in autaptic hippocampal neurons. Molecular pharmacology 76 (6), 1220–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (27).Straiker A, Wager-Miller J, Hu SS, Blankman JL, Cravatt BF, and Mackie K (2011) COX-2 and fatty acid amide hydrolase can regulate the time course of depolarization-induced suppression of excitation. British journal of pharmacology 164 (6), 1672—83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (28).Schlosburg JE, Blankman JL, Long JZ, Nomura DK, Pan B, Kinsey SG, Nguyen PT, Ramesh D, Booker L, Burston JJ, Thomas EA, Selley DE, Sim-Selley LJ, Liu QS, Lichtman AH, and Cravatt BF (2010) Chronic monoacylglycerol lipase blockade causes functional antagonism of the endocannabinoid system. Nat. Neurosci 13 (9), 1113–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (29).Pan B, Wang W, Zhong P, Blankman JL, Cravatt BF, and Liu QS (2011) Alterations of endocannabinoid signaling, synaptic plasticity, learning, and memory in monoacylglycerol lipase knock-out mice. J. Neurosci 31 (38), 13420–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (30).Poling JS, Rogawski MA, Salem N Jr., and Vicini S (1996) Anandamide, an endogenous cannabinoid, inhibits Shakerrelated voltage-gated K+ channels. Neuropharmacology 35 (7), 983–91. [DOI] [PubMed] [Google Scholar]
- (31).Maingret F, Patel AJ, Lazdunski M, and Honore E (2001) The endocannabinoid anandamide is a direct and selective blocker of the background K(+) channel TASK-1. EMBO J. 20 (1–2), 47–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (32).Chemin J, Monteil A, Perez-Reyes E, Nargeot J, and Lory P (2001) Direct inhibition of T-type calcium channels by the endogenous cannabinoid anandamide. EMBO J. 20 (24), 7033–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (33).Gantz SC, and Bean BP (2017) Cell-Autonomous Excitation of Midbrain Dopamine Neurons by Endocannabinoid-Dependent Lipid Signaling. Neuron 93 (6), 1375–1387.e2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (34).Katona I, Urban GM, Wallace M, Ledent C, Jung KM, Piomelli D, Mackie K, and Freund TF (2006) Molecular composition of the endocannabinoid system at glutamatergic synapses. J. Neurosd. 26 (2l), 5628–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (35).Yoshida T, Fukaya M, Uchigashima M, Miura E, Kamiya H, Kano M, and Watanabe M (2006) Localization of diacylglycerol lipase-alpha around postsynaptic spine suggests close proximity between production site of an endocannabinoid, 2-arachidonoyl-glycerol, and presynaptic cannabinoid CB1 receptor. J.Neurosci 26 (18), 4740–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (36).Yoshino H, Miyamae T, Hansen G, Zambrowicz B, Flynn M, Pedicord D, Blat Y, Westphal RS, Zaczek R, Lewis DA, and Gonzalez-Burgos G (2011) Postsynaptic diacylglycerol lipase mediates retrograde endocannabinoid suppression of inhibition in mouse prefrontal cortex. J. Physiol 589 (Pt 20), 4857–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (37).Di Marzo V (2011) Endocannabinoid signaling in the brain: biosynthetic mechanisms in the limelight. Nat. Neurosci 14 (1), 9–15. [DOI] [PubMed] [Google Scholar]
- (38).Ade KK, and Lovinger DM (2007) Anandamide regulates postnatal development of long-term synaptic plasticity in the rat dorsolateral striatum. J. Neurosci 21 (9), 2403–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (39).Blankman JL, Simon GM, and Cravatt BF (2007) A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem. Biol 14 (12), 1347–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (40).Gulyas AL, Cravatt BF, Bracey MH, Dinh TP, Piomelli D, Boscia F, and Freund TF (2004) Segregation of two endocannabinoid-hydrolyzing enzymes into pre- and postsynaptic compartments in the rat hippocampus, cerebellum and amygdala. European journal of neuroscience 20 (2), 441—58. [DOI] [PubMed] [Google Scholar]
- (41).Stella N, Schweitzer P, and Piomelli D (1997) A second endogenous cannabinoid that modulates long-term potentiation. Nature 388 (6644), 773–8. [DOI] [PubMed] [Google Scholar]
- (42).Giuffrida A, Parsons LH, Kerr TM, Rodriguez de Fonseca F, Navarro M, and Piomelli D (1999) Dopamine activation of endogenous cannabinoid signaling in dorsal striatum. Nat. Neurosci 2 (4), 358–63. [DOI] [PubMed] [Google Scholar]
- (43).Hashimotodani Y, Ohno-Shosaku T, and Kano M (2007) Presynaptic monoacylglycerol lipase activity determines basal endocannabinoid tone and terminates retrograde endocannabinoid signaling in the hippocampus. J. Neurosci 21(5), 1211–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (44).Maejima T, Oka S, Hashimotodani Y, Ohno-Shosaku T, Aiba A, Wu D, Waku K, Sugiura T, and Kano M (2005) Synaptically driven endocannabinoid release requires Ca2+−assisted metabotropic glutamate receptor subtype 1 to phospholipase Cbeta4 signaling cascade in the cerebellum. J. Neurosci 25 (29), 6826–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (45).Caille S, Alvarez-Jaimes L, Polis L, Stouffer DG, and Parsons LH (2007) Specific alterations of extracellular endocannabinoid levels in the nucleus accumbens by ethanol, heroin, and cocaine self-administration. J. Neurosci 21(14), 3695–702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (46).Devane WA, Dysarz FA 3rd, Johnson MR, Melvin LS, and Howlett AC (1988) Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol 34 (5), 605—13. [PubMed] [Google Scholar]
- (47).Herkenham M, Lynn AB, Little MD, Johnson MR, Melvin LS, de Costa BR, and Rice KC (1990) Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. U. S. A 87 (5), 1932–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (48).Tsou K, Brown S, Sanudo-Pena MC, Mackie K, and Walker JM (1998) Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience 83 (2), 393–411. [DOI] [PubMed] [Google Scholar]
- (49).Marsicano G, and Lutz B (1999) Expression of the cannabinoid receptor CB1 in distinct neuronal subpopulations in the adult mouse forebrain. European journal of neuroscience 11 (12), 4213–25. [DOI] [PubMed] [Google Scholar]
- (50).Lovinger DM (2010) Neurotransmitter roles in synaptic modulation, plasticity and learning in the dorsal striatum. Neuropharmacology 58 (7), 951–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (51).Bodor AL, Katona I, Nyiri G, Mackie K, Ledent C, Hajos N, and Freund TF (2005) Endocannabinoid signaling in rat somatosensory cortex: laminar differences and involvement of specific interneuron types. J. Neurosci 25 (29), 6845–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (52).Eggan SM, Melchitzky DS, Sesack SR, Fish KN, and Lewis DA (2010) Relationship of cannabinoid CB1 receptor and cholecystokinin immunoreactivity in monkey dorsolateral prefrontal cortex. Neuroscience 169 (4), 1651–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (53).Szabo GG, Lenkey N, Holderith N, Andrasi T, Nusser Z, and Hajos N (2014) Presynaptic calcium channel inhibition underlies CB(1) cannabinoid receptor-mediated suppression of GABA release. J. Neurosci 34 (23), 7958–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (54).Azad SC, Kurz J, Marsicano G, Lutz B, Zieglgansberger W, and Rammes G (2008) Activation of CB1 specifically located on GABAergic interneurons inhibits LTD in the lateral amygdala. Learn. Mem 15 (3), 143–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (55).Soltesz I, Alger BE, Kano M, Lee SH, Lovinger DM, Ohno-Shosaku T, and Watanabe M (2015) Weeding out bad waves: towards selective cannabinoid circuit control in epilepsy. Nat. Rev. Neurosci 16 (5), 264–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (56).Foldy C, Neu A, Jones MV, and Soltesz I (2006) Presynaptic, activity-dependent modulation of cannabinoid type 1 receptor-mediated inhibition of GABA release. J. Neurosci 26 (5), 1465–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (57).Herkenham M, Lynn AB, Johnson MR, Melvin LS, de Costa BR, and Rice KC (1991) Characterization and localization of cannabinoid receptors in rat brain: a quantitative in vitro autoradiographic study. J. Neurosci 11 (2), 563–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (58).Galiegue S, Mary S, Marchand J, Dussossoy D, Carriere D, Carayon P, Bouaboula M, Shire D, Fur G, and Casellas P (1995) Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Ear. J. Biochem 232 (l), 54–61. [DOI] [PubMed] [Google Scholar]
- (59).Van Sickle MD, Duncan M, Kingsley PJ, Mouihate A, Urbani P, Mackie K, Stella N, Makriyannis A, Piomelli D, Davison JS, Marnett LJ, Di Marzo V, Pittman QJ, Patel KD, and Sharkey KA (2005) Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 310 (5746), 329–32. [DOI] [PubMed] [Google Scholar]
- (60).Morgan NH, Stanford IM, and Woodhall GL (2009) Functional CB2 type cannabinoid receptors at CNS synapses. Neuropharmacology 51 (4), 356–68. [DOI] [PubMed] [Google Scholar]
- (61).Atwood BK, and Mackie K (2010) CB2: a cannabinoid receptor with an identity crisis. Br. J. Pharmacol 160 (3), 467–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (62).Chevaleyre V, Takahashi KA, and Castillo PE (2006) Endocannabinoid-mediated synaptic plasticity in the CNS. Annu. Rev. Neurosci 29, 37–76. [DOI] [PubMed] [Google Scholar]
- (63).Pitler TA, and Alger BE (1992) Postsynaptic spike firing reduces synaptic GABAA responses in hippocampal pyramidal cells. J. Neurosci 12 (10), 4122–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (64).Pitier TA, and Alger BE (1994) Depolarization-induced suppression of GABAergic inhibition in rat hippocampal pyramidal cells: G protein involvement in a presynaptic mechanism. Neuron 13 (6), 1447–55. [DOI] [PubMed] [Google Scholar]
- (65).Llano I, Leresche N, and Marty A (1991) Calcium entry increases the sensitivity of cerebellar Purkinje cells to applied GABA and decreases inhibitory synaptic currents. Neuron 6 (4), 565–74. [DOI] [PubMed] [Google Scholar]
- (66).Vincent P, Armstrong CM, and Marty A (1992) Inhibitory synaptic currents in rat cerebellar Purkinje cells: modulation by postsynaptic depolarization. J. Physiol 456, 453–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (67).Wilson RI, and Nicoll RA (2001) Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 410 (6828), 588–92. [DOI] [PubMed] [Google Scholar]
- (68).Wilson RI, and Nicoll RA (2002) Endocannabinoid signaling in the brain. Science 296 (5568), 678–82. [DOI] [PubMed] [Google Scholar]
- (69).Ohno-Shosaku T, Tsubokawa H, Mizushima I, Yoneda N, Zimmer A, and Kano M (2002) Presynaptic cannabinoid sensitivity is a major determinant of depolarization-induced retrograde suppression at hippocampal synapses. J. Neurosci 22 (10), 3864–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (70).Diana MA, and Marty A (2004) Endocannabinoid-mediated short-term synaptic plasticity: depolarization-induced suppression of inhibition (DSI) and depolarization-induced suppression of excitation (DSE). Br. J. Pharmacol 142(1), 9–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (71).Maejima T, Hashimoto K, Yoshida T, Aiba A, and Kano M (2001) Presynaptic inhibition caused by retrograde signal from metabotropic glutamate to cannabinoid receptors. Neuron 31 (3), 463–75. [DOI] [PubMed] [Google Scholar]
- (72).Makara JK, Mor M, Fegley D, Szabo SI, Kathuria S, Astarita G, Duranti A, Tontini A, Tarzia G, Rivara S, Freund TF, and Piomelli D (2005) Selective inhibition of 2-AG hydrolysis enhances endocannabinoid signaling in hippocampus. Nat. Neurosci 8 (9), 1139–41. [DOI] [PubMed] [Google Scholar]
- (73).Pan B, Wang W, Long JZ, Sun D, Hillard CJ, Cravatt BF, and Liu QS (2009) Blockade of 2-arachidonoylglycerol hydrolysis by selective monoacylglycerol lipase inhibitor 4-nitrophenyl 4-(dibenzo[d] [1,3]dioxol-5-yl(hydroxy)methyl)piperidine-1-carboxy-late (JZL184) Enhances retrograde endocannabinoid signaling. J. Pharmacol. Exp. Ther 331 (2), 591–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (74).Szabo B, Urbanski MJ, Bisogno T, Di Marzo V, Mendiguren A, Baer WU, and Freiman I (2006) Depolarizationinduced retrograde synaptic inhibition in the mouse cerebellar cortex is mediated by 2-arachidonoylglycerol. J. Physiol 577 (Pt 1), 263–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (75).Shonesy BC, Parrish WP, Haddad JK, Stephenson JR, Baldi R, Bluett RJ, Marks CR, Centanni SW, Folkes OM, Spiess K, Augustin SM, Mackie K, Lovinger DM, Winder DG, Patel S, and Colbran RJ (2017) Role of striatal direct pathway 2-arachidonoylglycerol signaling in sociability and repetitive behavior. Biol. Psychiatry, DOI: 10.1016/j.biopsych.2017.11.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (76).Melis M, Pistis M, Perra S, Muntoni AL, Pillolla G, and Gessa GL (2004) Endocannabinoids mediate presynaptic inhibition of glutamatergic transmission in rat ventral tegmental area dopamine neurons through activation of CB1 receptors. J. Neurosci 24 (1), 53–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (77).Jo YH, Chen YJ, Chua SC Jr., Talmage DA, and Role LW (2005) Integration of endocannabinoid and leptin signaling in an appetite-related neural circuit. Neuron 48 (6), 1055–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (78).Zhu PJ, and Lovinger DM (2005) Retrograde endocannabinoid signaling in a postsynaptic neuron/synaptic bouton preparation from basolateral amygdala. J. Neurosci 25 (26), 6199–207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (79).Narashima M, Uchigashima M, Hashimoto K, Watanabe M, and Kano M (2006) Depolarization-induced suppression of inhibition mediated by endocannabinoids at synapses from fast-spiking interneurons to medium spiny neurons in the striatum. European journal of neuroscience 24 (8), 2246–52. [DOI] [PubMed] [Google Scholar]
- (80).Hampson RE, Zhuang SY, Weiner JL, and Deadwyler SA (2003) Functional significance of cannabinoid-mediated, depolarization-induced suppression of inhibition (DSI) in the hippocampus. J. Neurophysiol 90 (1), 55–64. [DOI] [PubMed] [Google Scholar]
- (81).Zhuang S, Hampson RE, and Deadwyler SA (2005) Behaviorally relevant endocannabinoid action in hippocampus: dependence on temporal summation of multiple inputs. Behav. Pharmacol 16 (5–6), 463–71. [DOI] [PubMed] [Google Scholar]
- (82).Melis M, Perra S, Muntoni AL, Pillolla G, Lutz B, Marsicano G, Di Marzo V, Gessa GL, and Pistis M (2004) Prefrontal cortex stimulation induces 2-arachidonoyl-glycerol-mediated suppression of excitation in dopamine neurons. J. Neurosci 24 (47), 10707–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (83).Patel S, Kingsley PJ, Mackie K, Marnett LJ, and Winder DG (2009) Repeated homotypic stress elevates 2-arachidonoylglycerol levels and enhances short-term endocannabinoid signaling at inhibitory synapses in basolateral amygdala. Neuropsychopharmacology 34 (13), 2699–709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (84).Kamprath K, Romo-Parra H, Haring M, Gaburro S, Doengi M, Lutz B, and Pape HC (2011) Short-term adaptation of conditioned fear responses through endocannabinoid signaling in the central amygdala. Neuropsychopharmacology 36 (3), 652–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (85).Gerdeman GL, Ronesi J, and Lovinger DM (2002) Postsynaptic endocannabinoid release is critical to long-term depression in the striatum. Nat. Neurosci 5 (5), 446–51. [DOI] [PubMed] [Google Scholar]
- (86).Robbe D, Kopf M, Remaury A, Bockaert J, and Manzoni OJ (2002) Endogenous cannabinoids mediate long-term synaptic depression in the nucleus accumbens. Proc. Natl. Acad. Sci. U. S. A 99 (12), 8384–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (87).Sjostrom PJ, Turrigiano GG, and Nelson SB (2004) Endocannabinoid-dependent neocortical layer-5 LTD in the absence of postsynaptic spiking. J. Neurophysiol 92 (6), 3338–43. [DOI] [PubMed] [Google Scholar]
- (88).Chevaleyre V, and Castillo PE (2003) Heterosynaptic LTD of hippocampal GABAergic synapses: a novel role of endocannabinoids in regulating excitability. Neuron 38 (3), 461–72. [DOI] [PubMed] [Google Scholar]
- (89).Azad SC, Monory K, Marsicano G, Cravatt BF, Lutz B, Zieglgansberger W, and Rammes G (2004) Circuitry for associative plasticity in the amygdala involves endocannabinoid signaling. J. Neurosci 24 (44), 9953–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (90).Kreitzer AC, and Malenka RC (2005) Dopamine modulation of state-dependent endocannabinoid release and longterm depression in the striatum. J. Neurosci 25 (45), 10537–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (91).Soler-Llavina GJ, and Sabatini BL (2006) Synapse-specific plasticity and compartmentalized signaling in cerebellar stellate cells. Nat. Neurosci 9 (6), 798–806. [DOI] [PubMed] [Google Scholar]
- (92).Yasuda H, Huang Y, and Tsumoto T (2008) Regulation of excitability and plasticity by endocannabinoids and PKA in developing hippocampus. Proc. Natl. Acad. Sci. U. S. A 105 (8), 3106–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (93).Pan B, Hillard CJ, and Liu QS (2008) Endocannabinoid signaling mediates cocaine-induced inhibitory synaptic plasticity in midbrain dopamine neurons. J. Neurosci 28 (6), 1385–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (94).Adermark L, and Lovinger DM (2007) Combined activation of L-type Ca2+ channels and synaptic transmission is sufficient to induce striatal long-term depression. J. Neurosci 27 (25), 6781–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (95).Adermark L, and Lovinger DM (2009) Frequencydependent inversion of net striatal output by endocannabinoiddependent plasticity at different synaptic inputs. J. Neurosci 29 (5), 1375–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (96).Calabresi P, Pisani A, Mercuri NB, and Bernardi G (1994) Post-receptor mechanisms underlying striatal long-term depression. J. Neurosci 14 (8), 4871–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (97).Choi S, and Lovinger DM (1997) Decreased probability of neurotransmitter release underlies striatal long-term depression and postnatal development of corticostriatal synapses. Proc. Natl. Acad. Sci. U. S. A 94 (6), 2665–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (98).Calabresi P, Saiardi A, Pisani A, Baik JH, Centonze D, Mercuri NB, Bernardi G, and Borrelli E (1997) Abnormal synaptic plasticity in the striatum of mice lacking dopamine D2 receptors. J. Neurosci 17 (12), 4536–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (99).Wang Z, Kai L, Day M, Ronesi J, Yin HH, Ding J, Tkatch T, Lovinger DM, and Surmeier DJ (2006) Dopaminergic control of corticostriatal long-term synaptic depression in medium spiny neurons is mediated by cholinergic interneurons. Neuron 50 (3), 443–52. [DOI] [PubMed] [Google Scholar]
- (100).Kheirbek MA, Britt JP, Beeler JA, Ishikawa Y, McGehee DS, and Zhuang X (2009) Adenylyl cyclase type 5 contributes to corticostriatal plasticity and striatum-dependent learning. J. Neurosci 29 (39), 12115–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (101).Augustin SM, Beeler JA, McGehee DS, and Zhuang X (2014) Cyclic AMP and afferent activity govern bidirectional synaptic plasticity in striatopallidal neurons. J. Neurosci 34 (19), 6692–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (102).Best AR, and Regehr WG (2008) Serotonin evokes endocannabinoid release and retrogradely suppresses excitatory synapses. J. Neurosci 28 (25), 6508–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (103).Chevaleyre V, Heifets BD, Kaeser PS, Sudhof TC, and Castillo PE (2007) Endocannabinoid-mediated long-term plasticity requires cAMP/PKA signaling and RIM1alpha. Neuron 54 (5), 801–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (104).Ronesi J, and Lovinger DM (2005) Induction of striatal long-term synaptic depression by moderate frequency activation of cortical afferents in rat. J. Physiol 562 (Pt 1), 245–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (105).Mato S, Robbe D, Puente N, Grandes P, and Manzoni OJ (2005) Presynaptic homeostatic plasticity rescues long-term depression after chronic Delta 9-tetrahydrocannabinol exposure. J. Neurosci 25 (50), 11619–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (106).Ronesi J, Gerdeman GL, and Lovinger DM (2004) Disruption of endocannabinoid release and striatal long-term depression by postsynaptic blockade of endocannabinoid membrane transport. J. Neurosci 24 (7), 1673–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (107).Klapstein GJ, Fisher RS, Zanjani H, Cepeda C, Jokel ES, Chesselet MF, and Levine MS (2001) Electrophysiological and morphological changes in striatal spiny neurons in R6/2 Huntington’s disease transgenic mice. J. Neurophysiol 86 (6), 2667–77. [DOI] [PubMed] [Google Scholar]
- (108).Kreitzer AC, and Malenka RC (2007) Endocannabinoidmediated rescue of striatal LTD and motor deficits in Parkinson’s disease models. Nature 445 (7128), 643–7. [DOI] [PubMed] [Google Scholar]
- (109).Zhuang X, Mazzoni P, and Kang UJ (2013) The role of neuroplasticity in dopaminergic therapy for Parkinson disease. Nat. Rev. Neurol 9 (5), 248–56. [DOI] [PubMed] [Google Scholar]
- (110).Gabbott PL, Warner TA, Jays PR, Salway P, and Busby SJ (2005) Prefrontal cortex in the rat: projections to subcortical autonomic, motor, and limbic centers. J. Comp. Neurol 492 (2), 145–77. [DOI] [PubMed] [Google Scholar]
- (111).Knapska E, Radwanska K, Werka T, and Kaczmarek L (2007) Functional internal complexity of amygdala: focus on gene activity mapping after behavioral training and drugs of abuse. Physiol. Rev 87 (4), 1113–73. [DOI] [PubMed] [Google Scholar]
- (112).Courtney SM, Petit L, Haxby JV, and Ungerleider LG (1998) The role of prefrontal cortex in working memory: examining the contents of consciousness. Philos. Trans. R Soc., B 353 (1377), 1819–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (113).Miller EK, and Cohen JD (2001) An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci 24, 167–202. [DOI] [PubMed] [Google Scholar]
- (114).Dalley JW, Cardinal RN, and Robbins TW (2004) Prefrontal executive and cognitive functions in rodents: neural and neurochemical substrates. Neurosci. Biobehav. Rev 28 (7), 771—84. [DOI] [PubMed] [Google Scholar]
- (115).Bodovitz S (2008) The neural correlate of consciousness. J. Theor. Biol 254 (3), 594–8. [DOI] [PubMed] [Google Scholar]
- (116).Orsini CA, and Maren S (2012) Neural and cellular mechanisms of fear and extinction memory formation. Neurosci. Biobehav. Rev 36 (7), 1773–802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (117).Milad MR, and Quirk GJ (2012) Fear extinction as a model for translational neuroscience: ten years of progress. Annu. Rev. Psychol 63, 129–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (118).Quirk GJ, and Mueller D (2008) Neural mechanisms of extinction learning and retrieval. Neuropsychopharmacology 33 (1), 56–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (119).Heng L, Beverley JA, Steiner H, and Tseng KY (2011) Differential developmental trajectories for CB1 cannabinoid receptor expression in limbic/assodative and sensorimotor cortical areas. Synapse 65 (4), 278–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (120).Matsuda LA, Bonner TI, and Lolait SJ (1993) Localization of cannabinoid receptor mRNA in rat brain. J. Comp. Neurol 327 (4), 535–50. [DOI] [PubMed] [Google Scholar]
- (121).Trettel J, and Levine ES (2002) Cannabinoids depress inhibitory synaptic inputs received by layer 2/3 pyramidal neurons of the neocortex. J. Neurophysiol 88 (1), 534–9. [DOI] [PubMed] [Google Scholar]
- (122).Hill EL, Gallopin T, Ferezou I, Cauli B, Rossier J, Schweitzer P, and Lambolez B (2007) Functional CB1 receptors are broadly expressed in neocortical GABAergic and glutamatergic neurons. J. Neurophysiol 97 (4), 2580–9. [DOI] [PubMed] [Google Scholar]
- (123).Azad SC, Eder M, Marsicano G, Lutz B, Zieglgansberger W, and Rammes G (2003) Activation of the cannabinoid receptor type 1 decreases glutamatergic and GABAergic synaptic transmission in the lateral amygdala of the mouse. Learn. Mem 10 (2), 116–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (124).Domenici MR, Azad SC, Marsicano G, Schierloh A, Wotjak CT, Dodt HU, Zieglgansberger W, Lutz B, and Rammes G (2006) Cannabinoid receptor type 1 located on presynaptic terminals of principal neurons in the forebrain controls glutamatergic synaptic transmission. J. Neurosci 26 (2l), 5794–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (125).Gremel CM, Chancey JH, Atwood BK, Luo G, Neve R, Ramakrishnan C, Deisseroth K, Lovinger DM, and Costa RM (2016) Endocannabinoid Modulation of Orbitostriatal Circuits Gates Habit Formation. Neuron 90 (6), 1312–1324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (126).Auclair N, Otani S, Soubrie P, and Crepel F (2000) Cannabinoids modulate synaptic strength and plasticity at glutamatergic synapses of rat prefrontal cortex pyramidal neurons. J. Neurophysiol 83 (6), 3287–93. [DOI] [PubMed] [Google Scholar]
- (127).Chiu CQ, Puente N, Grandes P, and Castillo PE (2010) Dopaminergic modulation of endocannabinoid-mediated plasticity at GABAergic synapses in the prefrontal cortex. J Neurosci. 30 (21), 7236–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (128).Hilario MR, Clouse E, Yin HH, and Costa RM (2007) Endocannabinoid signaling is critical for habit formation. Front. Integr. Neurosci 1, 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (129).Nazzaro C, Greco B, Cerovic M, Baxter P, Rubino T, Trusel M, Parolaro D, Tkatch T, Benfenati F, Pedarzani P, and Tonini R (2012) SK channel modulation rescues striatal plasticity and control over habit in cannabinoid tolerance. Nat. Neurosci 15 (2), 284–93. [DOI] [PubMed] [Google Scholar]
- (130).Rubino T, Prini P, Piscitelli F, Zamberletti E, Trusel M, Melis M, Sagheddu C, Ligresti A, Tonini R, Di Marzo V, and Parolaro D (2015) Adolescent exposure to THC in female rats disrupts developmental changes in the prefrontal cortex. Neurobiol. Dis 73, 60–9. [DOI] [PubMed] [Google Scholar]
- (131).Cass DK, Flores-Barrera E, Thomases DR, Vital WF, Caballero A, and Tseng KY (2014) CB1 cannabinoid receptor stimulation during adolescence impairs the maturation of GABA function in the adult rat prefrontal cortex. Mol. Psychiatry 19 (5), 536–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (132).O’Shea M, Singh ME, McGregor IS, and Mallet PE (2004) Chronic cannabinoid exposure produces lasting memory impairment and increased anxiety in adolescent but not adult rats. J. Psychopharmacol 18 (4), 502–8. [DOI] [PubMed] [Google Scholar]
- (133).Renard J, Krebs MO, Jay TM, and Le Pen G (2013) Long-term cognitive impairments induced by chronic cannabinoid exposure during adolescence in rats: a strain comparison. Psychopharmacology 225 (4), 781–90. [DOI] [PubMed] [Google Scholar]
- (134).Schneider M, and Koch M (2003) Chronic pubertal, but not adult chronic cannabinoid treatment impairs sensorimotor gating, recognition memory, and the performance in a progressive ratio task in adult rats. Neuropsychopharmacology 28 (10), 1760–9. [DOI] [PubMed] [Google Scholar]
- (135).Kasanetz F, Lafourcade M, Deroche-Gamonet V, Revest JM, Berson N, Balado E, Fiancette JF, Renault P, Piazza PV, and Manzoni OJ (2013) Prefrontal synaptic markers of cocaine addiction-like behavior in rats. Mol. Psychiatry 18 (6), 129–31. [DOI] [PubMed] [Google Scholar]
- (136).Fourgeaud L, Mato S, Bouchet D, Hemar A, Worley PF, and Manzoni OJ (2004) A. single in vivo exposure to cocaine abolishes endocannabinoid-mediated long-term depression in the nucleus accumbens. J. Neurosci 24 (31), 6939–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (137).Tan H, Ahmad T, Loureiro M, Zunder J, and Laviolette SR (2014) The role of cannabinoid transmission in emotional memory formation: implications for addiction and schizophrenia. Frontiers in psychiatry 5, 73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (138).Wyrofsky R, McGonigle P, and Van Bockstaele EJ (2015) Drug discovery strategies that focus on the endocannabinoid signaling system in psychiatric disease. Expert Opin. Drug Discovery 10 (1), 17–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (139).Bukalo O, Pinard CR, and Holmes A (2014) Mechanisms to medicines: elucidating neural and molecular substrates of fear extinction to identify novel treatments for anxiety disorders. British journal of pharmacology 171 (20), 4690–718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (140).Sidhpura N, and Parsons LH (2011) Endocannabinoidmediated synaptic plasticity and addiction-related behavior. Neuropharmacology 61 (7), 1070–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (141).Parsons LH, and Hurd YL (2015) Endocannabinoid signalling in reward and addiction. Nat. Rev. Neurosci 16 (10), 579–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (142).Zlebnik NE, and Cheer JF (2016) Drug-Induced Alterations of Endocannabinoid-Mediated Plasticity in Brain Reward Regions. J. Neurosci 36 (40), 10230–10238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (143).Mateo Y, Johnson KA, Covey DP, Atwood BK, Wang HL, Zhang S, Gildish I, Cachope R, Bellocchio L, Guzman M, Morales M, Cheer JF, and Lovinger DM (2017) Endocannabinoid Actions on Cortical Terminals Orchestrate Local Modulation of Dopamine Release in the Nucleus Accumbens. Neuron 96 (5), 1112–1126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (144).Sesack SR, and Grace AA (2010) Cortico-Basal Ganglia reward network: microcircuitry. Neuropsychopharmacology 35 (1), 27–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (145).Kalivas PW (2009) The glutamate homeostasis hypothesis of addiction. Nat. Rev. Neurosci 10 (8), 561–72. [DOI] [PubMed] [Google Scholar]
- (146).Kauer JA, and Malenka RC (2007) Synaptic plasticity and addiction. Nat. Rev. Neurosci 8 (11), 844–58. [DOI] [PubMed] [Google Scholar]
- (147).Alvarez-Jaimes L, Polis I, and Parsons LH (2008) Attenuation of cue-induced heroin-seeking behavior by cannabinoid CB1 antagonist infusions into the nucleus accumbens core and prefrontal cortex, but not basolateral amygdala. Neuropsychopharmacology 33 (10), 2483–93. [DOI] [PubMed] [Google Scholar]
- (148).Berke JD, and Hyman SE (2000) Addiction, dopamine, and the molecular mechanisms of memory. Neuron 25 (3), 515–32. [DOI] [PubMed] [Google Scholar]
- (149).Vanderschuren LJ, and Kalivas PW (2000) Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: a critical review of preclinical studies. Psychopharmacology 151(2–3), 99–120. [DOI] [PubMed] [Google Scholar]
- (150).Britt JP, Benaliouad F, McDevitt RA, Stuber GD, Wise RA, and Bonci A (2012) Synaptic and behavioral profile of multiple glutamatergic inputs to the nucleus accumbens. Neuron 76 (4), 790–803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (151).Lemstra AW, Verhagen Metman L, Lee JI, Dougherty PM, and Lenz FA (1999) Tremor-frequency (3–6 Hz) activity in the sensorimotor arm representation of the internal segment of the globus pallidus in patients with Parkinson’s disease. Neurosci. Lett 267 (2), 129–32. [DOI] [PubMed] [Google Scholar]
- (152).Jankovic J (2008) Parkinson’s disease: clinical features and diagnosis. J. Neurol., Neurosurg. Psychiatry 79 (4), 368–76. [DOI] [PubMed] [Google Scholar]
- (153).Gelb DJ, Oliver E, and Gilman S (1999) Diagnostic criteria for Parkinson disease. Arch. Neurol 56 (1), 33–9. [DOI] [PubMed] [Google Scholar]
- (154).Graybiel AM, Aosaki T, Flaherty AW, and Kimura M (1994) The basal ganglia and adaptive motor control. Science 265 (5180), 1826–31. [DOI] [PubMed] [Google Scholar]
- (155).Yin HH, and Knowlton BJ (2006) The role of the basal ganglia in habit formation. Nat. Rev. Neurosci. 7 (6), 464–76. [DOI] [PubMed] [Google Scholar]
- (156).Shen W, Flajolet M, Greengard P, and Surmeier DJ (2008) Dichotomous dopaminergic control of striatal synaptic plasticity. Science 321 (5890), 848–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (157).Bagetta V, Picconi B, Marinucci S, Sgobio C, Pendolino V, Ghiglieri V, Fusco FR, Giampa C, and Calabresi P (2011) Dopamine-dependent long-term depression is expressed in striatal spiny neurons of both direct and indirect pathways: implications for Parkinson’s disease. J. Neurosci 31 (35), 12513–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (158).Calabresi P, Galletti F, Saggese E, Ghiglieri V, and Picconi B (2007) Neuronal networks and synaptic plasticity in Parkinson’s disease: beyond motor deficits. Parkinsonism Relat. Disord 13 (Suppl 3), S259–62. [DOI] [PubMed] [Google Scholar]
- (159).Belujon P, Lodge DJ, and Grace AA (2010) Aberrant striatal plasticity is specifically associated with dyskinesia following levodopa treatment. Mov. Disord 25 (11), 1568–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (160).Tozzi A, de Iure A, Di Filippo M, Tantucci M, Costa C, Borsini F, Ghiglieri V, Giampa C, Fusco FR, Picconi B, and Calabresi P (2011) The distinct role of medium spiny neurons and cholinergic interneurons in the D(2)/A(2)A receptor interaction in the striatum: implications for Parkinson’s disease. J. Neurosci 31 (5), 1850–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (161).Lerner TN, and Kreitzer AC (2012) RGS4 is required for dopaminergic control of striatal LTD and susceptibility to parkinsonian motor deficits. Neuron 73 (2), 347–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (162).Gubellini P, Picconi B, Bari M, Battista N, Calabresi P, Centonze D, Bernardi G, Finazzi-Agro A, and Maccarrone M (2002) Experimental parkinsonism alters endocannabinoid degradation: implications for striatal glutamatergic transmission. J. Neurosci 22 (16), 6900–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (163).Di Marzo V, Hill MP, Bisogno T, Crossman AR, and Brotchie JM (2000) Enhanced levels of endogenous cannabinoids in the globus pallidus are associated with a reduction in movement in an animal model of Parkinson’s disease. FASEB J. 14 (10), 1432–8. [DOI] [PubMed] [Google Scholar]
- (164).Calabresi P, Maj R, Pisani A, Mercuri NB, and Bernardi G (1992) Long-term synaptic depression in the striatum: physiological and pharmacological characterization. J. Neurosci 12 (11), 4224–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (165).Picconi B, Centonze D, Hakansson K, Bernardi G, Greengard P, Fisone G, Cenci MA, and Calabresi P (2003) Loss of bidirectional striatal synaptic plasticity in L-DOPA-induced dyskinesia. Nat. Neurosci 6 (5), 501–6. [DOI] [PubMed] [Google Scholar]
- (166).Thiele SL, Chen B, Lo C, Gertler TS, Warre R, Surmeier JD, Brotchie JM, and Nash JE (2014) Selective loss of bi-directional synaptic plasticity in the direct and indirect striatal output pathways accompanies generation of parkinsonism and 1-DOPA induced dyskinesia in mouse models. Neurobiol. Dis 71, 334–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (167).Adermark L (2011) Modulation of endocannabinoidmediated long-lasting disinhibition of striatal output by cholinergic interneurons. Neuropharmacology 61 (8), 1314–20. [DOI] [PubMed] [Google Scholar]
- (168).Mathur BN, Tanahira C, Tamamaki N, and Lovinger DM (2013) Voltage drives diverse endocannabinoid signals to mediate striatal microcircuit-specific plasticity. Nat. Neurosci 16 (9), 1275–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (169).Milad MR, and Quirk GJ (2002) Neurons in medial prefrontal cortex signal memory for fear extinction. Nature 420 (6911), 70–4. [DOI] [PubMed] [Google Scholar]
- (170).Santini E, Ge H, Ren K, Pena de Ortiz S, and Quirk GJ (2004) Consolidation of fear extinction requires protein synthesis in the medial prefrontal cortex. J. Neurosci 24 (25), 5704–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (171).Maren S, and Quirk GJ (2004) Neuronal signalling of fear memory. Nat. Rev. Neurosci 5 (11), 844–52. [DOI] [PubMed] [Google Scholar]
- (172).Shin LM, and Liberzon I (2010) The neurocircuitry of fear, stress, and anxiety disorders. Neuropsychopharmacology 35 (1), 169–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (173).Kuhnert S, Meyer C, and Koch M (2013) Involvement of cannabinoid receptors in the amygdala and prefrontal cortex of rats in fear learning consolidation, retrieval and extinction. Behav. Brain Res. 250, 274–84. [DOI] [PubMed] [Google Scholar]
- (174).Chhatwal JP, Davis M, Maguschak KA, and Ressler KJ (2005) Enhancing cannabinoid neurotransmission augments the extinction of conditioned fear. Neuropsychopharmacology 30 (3), 516–24. [DOI] [PubMed] [Google Scholar]
- (175).Marsicano G, Wotjak CT, Azad SC, Bisogno T, Rammes G, Cascio MG, Hermann H, Tang J, Hofmann C, Zieglgansberger W, Di Marzo V, and Lutz B (2002) The endogenous cannabinoid system controls extinction of aversive memories. Nature 418 (6897), 530–4. [DOI] [PubMed] [Google Scholar]
- (176).Varvel SA, Wise LE, Niyuhire F, Cravatt BF, and Lichtman AH (2007) Inhibition of fatty-acid amide hydrolase accelerates acquisition and extinction rates in a spatial memory task Neuropsychopharmacology 32 (5), 1032–41. [DOI] [PubMed] [Google Scholar]
- (177).Gunduz-Cinar O, MacPherson KP, Cinar R, GambleGeorge J, Sugden K, Williams B, Godlewski G, Ramikie TS, Gorka AX, Alapafuja SO, Nikas SP, Makriyannis A, Poulton R, Patel S, Hariri AR, Caspi A, Moffitt TE, Kunos G, and Holmes A (2013) Convergent translational evidence of a role for anandamide in amygdala-mediated fear extinction, threat processing and stressreactivity. Mol. Psychiatry 18 (7), 813–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (178).Holter SM, Kallnik M, Wurst W, Marsicano G, Lutz B, and Wotjak CT (2005) Cannabinoid CB1 receptor is dispensable for memory extinction in an appetitively-motivated learning task Eur. J. Pharmacol 510 (1–2), 69–74. [DOI] [PubMed] [Google Scholar]
- (179).Katona I, Sperlagh B, Sik A, Kafalvi A, Vizi ES, Mackie K, and Freund TF (1999) Presynaptically located CB1 cannabinoid receptors regulate GABA release from axon terminals of specific hippocampal interneurons. J. Neurosci 19 (11), 4544–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (180).Tsou K, Mackie K, Sanudo-Pena MC, and Walker JM (1999) Cannabinoid CB1 receptors are localized primarily on cholecystokinin-containing GABAergic interneurons in the rat hippocampal formation. Neuroscience 93 (3), 969–75. [DOI] [PubMed] [Google Scholar]
- (181).Katona I, Rancz EA, Acsady L, Ledent C, Mackie K, Hajos N, and Freund TF (2001) Distribution of CB1 cannabinoid receptors in the amygdala and their role in the control of GABAergic transmission. J. Neurosci 21 (23), 9506–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (182).Remmers F, Lange MD, Hamann M, Ruehle S, Pape HC, and Lutz B (2017) Addressing sufficiency of the CB1 receptor for endocannabinoid-mediated functions through conditional genetic rescue in forebrain GABAergic neurons. Brain Struct. Fund 222 (8), 3431–3452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (183).Chevaleyre V, and Castillo PE (2004) Endocannabinoidmediated metaplasticity in the hippocampus. Neuron 43 (6), 871–81. [DOI] [PubMed] [Google Scholar]
- (184).Woolsey TA, and Van der Loos H (1970) The structural organization of layer IV in the somatosensory region (SI) of mouse cerebral cortex. The description of a cortical field composed of discrete cytoarchitectonic units. Brain Res. 17 (2), 205–42. [DOI] [PubMed] [Google Scholar]
- (185).Broser P, Grinevich V, Osten P, Sakmann B, and Wallace DJ (2008) Critical period plasticity of axonal arbors of layer 2/3 pyramidal neurons in rat somatosensory cortex: layer-specific reduction of projections into deprived cortical columns. Cerebral cortex 18 (7), 1588–603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (186).Buonomano DV, and Merzenich MM (1998) Cortical plasticity: from synapses to maps. Annu. Rev. Neurosci 21, 149–86. [DOI] [PubMed] [Google Scholar]
- (187).Celikel T, Szostak VA, and Feldman DE (2004) Modulation of spike timing by sensory deprivation during induction of cortical map plasticity. Nat. Neurosci 7 (5), 534–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (188).Shepherd GM, Pologruto TA, and Svoboda K (2003) Circuit analysis of experience-dependent plasticity in the developing rat barrel cortex. Neuron 38 (2), 277–89. [DOI] [PubMed] [Google Scholar]
- (189).Feldman DE (2000) Timing-based LTP and LTD at vertical inputs to layer II/III pyramidal cells in rat barrel cortex. Neuron 27 (1), 45–56. [DOI] [PubMed] [Google Scholar]
- (190).Feldman DE (2009) Synaptic mechanisms for plasticity in neocortex. Annu. Rev. Neurosci 32, 33–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (191).Fox K (1992) A critical period for experience-dependent synaptic plasticity in rat barrel cortex. J. Neurosci 12 (5), 1826–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (192).Stern EA, Maravall M, and Svoboda K (2001) Rapid development and plasticity of layer 2/3 maps in rat barrel cortex in vivo. Neuron 31 (2), 305–15. [DOI] [PubMed] [Google Scholar]
- (193).Bender KJ, Allen CB, Bender VA, and Feldman DE (2006) Synaptic basis for whisker deprivation-induced synaptic depression in rat somatosensory cortex. J. Neurosci 26 (l6), 4155–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (194).Bender VA, Bender KJ, Brasier DJ, and Feldman DE (2006) Two coincidence detectors for spike timing-dependent plasticity in somatosensory cortex. J. Neurosci 26 (16), 4166–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (195).Allen CB, Celikel T, and Feldman DE (2003) Long-term depression induced by sensory deprivation during cortical map plasticity in vivo. Nat. Neurosci 6 (3), 291–9. [DOI] [PubMed] [Google Scholar]
- (196).Li L, Bender KJ, Drew PJ, Jadhav SP, Sylwestrak E, and Feldman DE (2009) Endocannabinoid signaling is required for development and critical period plasticity of the whisker map in somatosensory cortex Neuron 64 (4), 537–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (197).Trettel J, and Levine ES (2003) Endocannabinoids mediate rapid retrograde signaling at interneuron right-arrow pyramidal neuron synapses of the neocortex. J. Neurophysiol 89 (4), 2334–8. [DOI] [PubMed] [Google Scholar]
- (198).Fortin DA, and Levine ES (2007) Differential effects of endocannabinoids on glutamatergic and GABAergic inputs to layer 5 pyramidal neurons. Cerebral Cortex 17 (1), 163–74. [DOI] [PubMed] [Google Scholar]
- (199).Lemtiri-Chlieh F, and Levine ES (2007) Lack of depolarization-induced suppression of inhibition (DSI) in layer 2/3 interneurons that receive cannabinoid-sensitive inhibitory inputs. J. Neurophysiol 98 (5), 2517–24. [DOI] [PubMed] [Google Scholar]
- (200).Lemtiri-Chlieh F, and Levine ES (2010) BDNF evokes release of endogenous cannabinoids at layer 2/3 inhibitory synapses in the neocortex J. Neurophysiol 104 (4), 1923–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (201).Zhao L, and Levine ES (2014) BDNF-endocannabinoid interactions at neocortical inhibitory synapses require phospholipase C signaling. J. Neurophysiol 111 (5), 1008–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (202).Yeh ML, Selvam R, and Levine ES (2017) BDNFinduced endocannabinoid release modulates neocortical glutamatergic neurotransmission. Synapse 71 (5), e21962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (203).Bacci A, Huguenard JR, and Prince DA (2004) Longlasting self-inhibition of neocortical interneurons mediated by endocannabinoids. Nature 431 (7006), 312–6. [DOI] [PubMed] [Google Scholar]
- (204).Marinelli S, Pacioni S, Bisogno T, Di Marzo V, Prince DA, Huguenard JR, and Bacci A (2008) The endocannabinoid 2-arachidonoylglycerol is responsible for the slow self-inhibition in neocortical interneurons. J. Neurosci 28 (50), 13532–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (205).Marinelli S, Pacioni S, Cannich A, Marsicano G, and Bacci A (2009) Self-modulation of neocortical pyramidal neurons by endocannabinoids. Nat. Neurosci 12 (12), 1488–90. [DOI] [PubMed] [Google Scholar]
- (206).Bouchard JF, Casanova C, Cecyre B, and Redmond WJ (2016) Expression and Function of the Endocannabinoid System in the Retina and the Visual Brain. Neural Plast. 2016, 9247057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (207).Wiesel TN, and Hubel DH (1963) Single-Cell Responses in Striate Cortex of Kittens Deprived of Vision in One Eye. J. Neurophysiol 26, 1003–17. [DOI] [PubMed] [Google Scholar]
- (208).Liu CH, Heynen AJ, Shuler MG, and Bear MF (2008) Cannabinoid receptor blockade reveals parallel plasticity mechanisms in different layers of mouse visual cortex. Neuron 58 (3), 340–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (209).Crozier RA, Wang Y, Liu CH, and Bear MF (2007) Deprivation-induced synaptic depression by distinct mechanisms in different layers of mouse visual cortex. Proc. Natl. Acad. Sci. U. S. A 104 (4), 1383–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (210).Jiang B, Huang S, de Pasquale R, Millman D, Song L, Lee HK, Tsumoto T, and Kirkwood A (2010) The maturation of GABAergic transmission in visual cortex requires endocannabinoidmediated LTD of inhibitory inputs during a critical period. Neuron 66 (2), 248–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (211).Sun W, Wang L, Li S, Tie X, and Jiang B (2015) Layer-specific endocannabinoid-mediated long-term depression of GABAergic neurotransmission onto principal neurons in mouse visual cortex. European journal of neuroscience 42(3), 1952–65. [DOI] [PubMed] [Google Scholar]
- (212).Jiang B, Sohya K, Sarihi A, Yanagawa Y, and Tsumoto T (2010) Laminar-specific maturation of GABAergic transmission and susceptibility to visual deprivation are related to endocannabinoid sensitivity in mouse visual cortex. J. Neurosci 30 (42), 14261–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (213).Ito M, Yamaguchi K, Nagao S, and Yamazaki T (2014) Long-term depression as a model of cerebellar plasticity. Prog. Brain Res. 210, 1–30. [DOI] [PubMed] [Google Scholar]
- (214).Safo PK, and Regehr WG (2005) Endocannabinoids control the induction of cerebellar LTD. Neuron 48 (4), 647–59. [DOI] [PubMed] [Google Scholar]
- (215).Carey MR, Myoga MH, McDaniels KR, Marsicano G, Lutz B, Mackie K, and Regehr WG (2011) Presynaptic CB1 receptors regulate synaptic plasticity at cerebellar parallel fiber synapses. J. Neurophysiol 105 (2), 958–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (216).Safo P, and Regehr WG (2008) Timing dependence of the induction of cerebellar LTD. Neuropharmacology 54 (1), 213–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (217).Qiu DL, and Knopfel T (2009) Presynaptically expressed long-term depression at cerebellar parallel fiber synapses. Pfluegers Arch. 457 (4), 865–75. [DOI] [PubMed] [Google Scholar]
- (218).Chu CP, Zhao GY, Jin R, Zhao SN, Sun L, and Qiu DL (2014) Properties of 4 Hz stimulation-induced parallel fiber-Purkinje cell presynaptic long-term plasticity in mouse cerebellar cortex in vivo. European journal of neuroscience 39 (10), 1624–31. [DOI] [PubMed] [Google Scholar]
- (219).Kishimoto Y, and Kano M (2006) Endogenous cannabinoid signaling through the CB1 receptor is essential for cerebellumdependent discrete motor learning. J. Neurosci 26 (34), 8829–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (220).Moser EI, Moser MB, and McNaughton BL (2017) Spatial representation in the hippocampal formation: a history. Nat. Neurosci 20 (11), 1448–1464. [DOI] [PubMed] [Google Scholar]
- (221).Yonelinas AP (2013) The hippocampus supports highresolution binding in the service of perception, working memory and long-term memory. Behav. Brain Res. 254, 34–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (222).Heyser C. j., Hampson RE, and Deadwyler SA (1993) Effects of delta-9-tetrahydrocannabinol on delayed match to sample performance in rats: alterations in short-term memory associated with changes in task specific firing of hippocampal cells. J. Pharmacol. Exp. Ther 264 (1), 294–307. [PubMed] [Google Scholar]
- (223).Lichtman AH, Dimen KR, and Martin BR (1995) Systemic or intrahippocampal cannabinoid administration impairs spatial memory in rats. Psychopharmacology 119 (3), 282–90. [DOI] [PubMed] [Google Scholar]
- (224).Davies SN, Pertwee RG, and Riedel G (2002) Functions of cannabinoid receptors in the hippocampus. Neuropharmacology 42 (8), 993–1007. [DOI] [PubMed] [Google Scholar]
- (225).de Oliveira Alvares L, Engelke DS, Diehl F, Scheffer-Teixeira R, Haubrich J, de Freitas Cassini L, Molina VA, and Quillfeldt JA (2010) Stress response recruits the hippocampal endocannabinoid system for the modulation of fear memory. Learn. Mem 17 (4), 202–9. [DOI] [PubMed] [Google Scholar]
- (226).Reich CG, Mohammadi MH, and Alger BE (2008) Endocannabinoid modulation of fear responses: learning and statedependent performance effects. J. Psychopharmacol 22 (7), 769–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (227).Lichtman AH (2000) SR 141716A enhances spatial memory as assessed in a radial-arm maze task in rats. Eur.J. Pharmacol 404 (l–2), 175–9. [DOI] [PubMed] [Google Scholar]
- (228).Robinson L, McKillop-Smith S, Ross NL, Pertwee RG, Hampson RE, Platt B, and Riedel G (2008) Hippocampal endocannabinoids inhibit spatial learning and limit spatial memory in rats. Psychopharmacology 198 (4), 551–63. [DOI] [PubMed] [Google Scholar]
- (229).Wolff MC, and Leander JD (2003) SR141716A, a cannabinoid CB1 receptor antagonist, improves memory in a delayed radial maze task Eur. J. Pharmacol 477 (3), 213–7. [DOI] [PubMed] [Google Scholar]
- (230).Morena M, De Castro V, Gray JM, Palmery M, Trezza V, Roozendaal B, Hill MN, and Campolongo P (2015) Training-Associated Emotional Arousal Shapes Endocannabinoid Modulation of Spatial Memory Retrieval in Rats. J. Neurosci 35 (41), 13962–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (231).Xu J, Antion MD, Nomura T, Kraniotis S, Zhu Y, and Contractor A (2014) Hippocampal metaplasticity is required for the formation of temporal associative memories. J. Neurosci 34 (50), 16762–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (232).Griebel G, Pichat P, Beeske S, Leroy T, Redon N, Jacquet A, Francon D, Bert L, Even L, Lopez-Grancha M, Tolstykh T, Sun F, Yu Q, Brittain S, Arlt H, He T, Zhang B, Wiederschain D, Bertrand T, Houtmann J, Rak A, Vallee F, Michot N, Auge F, Menet V, Bergis OE, George P, Avenet P, Mikol V, Didier M, and Escoubet J (2015) Selective blockade of the hydrolysis of the endocannabinoid 2-arachidonoylglycerol impairs learning and memory performance while producing antinociceptive activity in rodents. Sci. Rep 5, 7642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (233).Basavarajappa BS, Nagre NN, Xie S, and Subbanna S (2014) Elevation of endogenous anandamide impairs LTP, learning, and memory through CB1 receptor signaling in mice. Hippocampus 24 (7), 808–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (234).Blair RE, Deshpande laxmikant S, and DeLorenzo RJ (2015) Endocannabinoids and epilepsy In Cannabinoids in neurologic and mental disease (Fattore L, Ed.), 1st ed., pp 125–172, Chapter 6, Academic Press, Waltham, MA. [Google Scholar]
- (235).Reddy DS, and Golub VM (2016) The Pharmacological Basis of Cannabis Therapy for Epilepsy. J. Pharmacol. Exp. Ther 357 (1), 45–55. [DOI] [PubMed] [Google Scholar]
- (236).Karlocai MR, Toth K, Watanabe M, Ledent C, Juhasz G, Freund TF, and Magloczky Z (2011) Redistribution of CB1 cannabinoid receptors in the acute and chronic phases of pilocarpineinduced epilepsy. PLoS One 6 (11), e27196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (237).Falenski KW, Blair RE, Sim-Selley LJ, Martin BR, and DeLorenzo RJ (2007) Status epilepticus causes a long-lasting redistribution of hippocampal cannabinoid type 1 receptor expression and function in the rat pilocarpine model of acquired epilepsy. Neuroscience 146 (3), 1232–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (238).Wallace MJ, Blair RE, Falenski KW, Martin BR, and DeLorenzo RJ (2003) The endogenous cannabinoid system regulates seizure frequency and duration in a model of temporal lobe epilepsy. J. Pharmacol. Exp. Ther 307 (1), 129–37. [DOI] [PubMed] [Google Scholar]
- (239).Chen K, Ratzliff A, Hilgenberg L, Gulyas A, Freund TF, Smith M, Dinh TP, Piomelli D, Mackie K, and Soltesz I (2003) Long-term plasticity of endocannabinoid signaling induced by developmental febrile seizures. Neuron 39 (4), 599–611. [DOI] [PubMed] [Google Scholar]
- (240).Ludanyi A, Eross L, Czirjak S, Vajda J, Halasz P, Watanabe M, Palkovits M, Magloczky Z, Freund TF, and Katona I (2008) Downregulation of the CB1 cannabinoid receptor and related molecular elements of the endocannabinoid system in epileptic human hippocampus. J. Neurosci 28 (12), 2976–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (241).Magloczky Z, Toth K, Karlocai R, Nagy S, Eross L, Czirjak S, Vajda J, Rasonyi G, Kelemen A, Juhos V, Halasz P, Mackie K, and Freund TF (2010) Dynamic changes of CB1-receptor expression in hippocampi of epileptic mice and humans. Epilepsia 51 (Suppl 3), 115–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (242).Goffin K, Van Paesschen W, and Van Laere K (2011) In vivo activation of endocannabinoid system in temporal lobe epilepsy with hippocampal sclerosis. Brain 134 (Pt 4), 1033–40. [DOI] [PubMed] [Google Scholar]
- (243).Romigi A, Bari M, Placidi F, Marciani MG, Malaponti M, Torelli F, Izzi F, Prosperetti C, Zannino S, Corte F, Chiaramonte C, and Maccarrone M (2010) Cerebrospinal fluid levels of the endocannabinoid anandamide are reduced in patients with untreated newly diagnosed temporal lobe epilepsy. Epilepsia 51 (5), 768–72. [DOI] [PubMed] [Google Scholar]
- (244).Stadnicki SW, Schaeppi U, Rosenkrantz H, and Braude MC (1974) Delta9-tetrahydrocannabinol: subcortical spike bursts and motor manifestations in a Fischer rat treated orally for 109 days. Life Sci. 14 (3), 463–72. [DOI] [PubMed] [Google Scholar]
- (245).Martin P, and Consroe P (1976) Cannabinoid induced behavioral convulsions in rabbits. Science 194 (4268), 965–7. [DOI] [PubMed] [Google Scholar]
- (246).Gordon E, and Devinsky O (2001) Alcohol and marijuana: effects on epilepsy and use by patients with epilepsy. Epilepsia 42 (10), 1266–72. [DOI] [PubMed] [Google Scholar]
- (247).Deshpande LS, Sombati S, Blair RE, Carter DS, Martin BR, and DeLorenzo RJ (2007) Cannabinoid CB1 receptor antagonists cause status epilepticus-like activity in the hippocampal neuronal culture model of acquired epilepsy. Neurosci. Lett 411 (1), 11–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (248).Marsicano G, Goodenough S, Monory K, Hermann H, Eder M, Cannich A, Azad SC, Cascio MG, Gutierrez SO, van der Stelt M, Lopez-Rodriguez ML, Casanova E, Schutz G, Zieglgansberger W, Di Marzo V, Behl C, and Lutz B (2003) CB1 cannabinoid receptors and on-demand defense against excitotoxidty. Science 302 (5642), 84–8. [DOI] [PubMed] [Google Scholar]
- (249).Shafaroodi H, Samini M, Moezi L, Homayoun H, Sadeghipour H, Tavakoli S, Hajrasouliha AR, and Dehpour AR (2004) The interaction of cannabinoids and opioids on pentylenetetrazole-induced seizure threshold in mice. Neuropharmacology 47 (3), 390–400. [DOI] [PubMed] [Google Scholar]
- (250).Wallace MJ, Martin BR, and DeLorenzo RJ (2002) Evidence for a physiological role of endocannabinoids in the modulation of seizure threshold and severity. Eur. J. Pharmacol 452 (3), 295–301. [DOI] [PubMed] [Google Scholar]
- (251).Chen K, Neu A, Howard AL, Foldy C, Echegoyen J, Hilgenberg L, Smith M, Mackie K, and Soltesz I (2007) Prevention of plasticity of endocannabinoid signaling inhibits persistent limbic hyperexcitability caused by developmental seizures. J. Neurosci 27 (1), 46–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (252).Echegoyen J, Armstrong C, Morgan RJ, and Soltesz I (2009) Single application of a CB1 receptor antagonist rapidly following head injury prevents long-term hyperexcitability in a rat model. Epilepsy Res. 85 (1), 123–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (253).Monory K, Massa F, Egertova M, Eder M, Blaudzun H, Westenbroek R, Relsch W, Jacob W, Marsch R, Ekker M, Long J, Rubenstein JL, Goebbels S, Nave KA, During M, Klugmann M, Wolfel B, Dodt HU, Zieglgansberger W, Wotjak CT, Mackie K, Elphick MR, Marsicano G, and Lutz B (2006) The endocannabinoid system controls key epileptogenic circuits in the hippocampus. Neuron 51 (4), 455–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (254).Devinsky O, Cilio MR, Cross H, Fernandez-Ruiz J, French J, Hill C, Katz R, Di Marzo V, Jutras-Aswad D, Notcutt WG, Martinez-Orgado J, Robson PJ, Rohrback BG, Thiele E, Whalley B, and Friedman D (2014) Cannabidiol: pharmacology and potential therapeutic role in epilepsy and other neuropsychiatric disorders. Epilepsia 55 (6), 791–802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (255).Maa E, and Figi P (2014) The case for medical marijuana in epilepsy. Epilepsia 55 (6), 783–6. [DOI] [PubMed] [Google Scholar]
- (256).Cui Y, Paille V, Xu H, Genet S, Delord B, Fino E, Berry H, and Venance L (2015) Endocannabinoids mediate bidirectional striatal spike-timing-dependent plasticity. J. Physiol 593 (13), 2833–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (257).Cui Y, Prokin I, Xu H, Delord B, Genet S, Venance L, and Berry H (2016) Endocannabinoid dynamics gate spike-timing dependent depression and potentiation. eLife 5, e13185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (258).Xu JY, Zhang J, and Chen C (2012) Long-lasting potentiation of hippocampal synaptic transmission by direct cortical input is mediated via endocannabinoids. J. Physiol 590 (10), 2305–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (259).Maglio LE, Noriega-Prieto JA, Maraver MJ, and Fernandez de Sevilla D (2017) Endocannabinoid-Dependent Long-Term Potentiation of Synaptic Transmission at Rat Barrel Cortex. Cerebral cortex, 1–14. [DOI] [PubMed] [Google Scholar]
- (260).Patel S, Gerrits R, Muthian S, Greene AS, and Hillard CJ (2002) The CB1 receptor antagonist SR141716 enhances stimulusinduced activation of the primary somatosensory cortex of the rat. Neurosci. Lett 335 (2), 95–8. [DOI] [PubMed] [Google Scholar]
- (261).Ho WS, Patel S, Thompson JR, Roberts CJ, Stuhr KL, and Hillard CJ (2010) Endocannabinoid modulation of hyperaemia evoked by physiologically relevant stimuli in the rat primary somatosensory cortex. Br.J. Pharmacol 160 (3), 736–46. [DOI] [PMC free article] [PubMed] [Google Scholar]