

Abstract
Quercetin, a natural flavonol existing in many food resources, has been reported to be an effective antimicrobial and anti-inflammatory agent for restricting the inflammation in periodontitis. In this study, we aimed to investigate the anti-inflammatory effects of quercetin on Porphyromonas gingivalis (P. gingivalis) lipopolysaccharide- (LPS-) stimulated human gingival fibroblasts (HGFs). HGFs were pretreated with quercetin prior to LPS stimulation. Cell viability was evaluated by 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay. The levels of inflammatory cytokines, including interleukin-1β (IL-1β), interleukin-6 (IL-6), and tumor necrosis factor-α (TNF-α), along with chemokine interleukin-8 (IL-8), were determined by enzyme-linked immunosorbent assay (ELISA). The mRNA levels of IL-1β, IL-6, IL-8, TNF-α, IκBα, p65 subunit of nuclear factor-kappa B (NF-κB), peroxisome proliferator-activated receptor-γ (PPAR-γ), liver X receptor α (LXRα), and Toll-like receptor 4 (TLR4) were measured by real-time quantitative PCR (RT-qPCR). The protein levels of IκBα, p-IκBα, p65, p-p65, PPAR-γ, LXRα, and TLR4 were characterized by Western blotting. Our results demonstrated that quercetin inhibited the LPS-induced production of IL-1β, IL-6, IL-8, and TNF-α in a dose-dependent manner. It also suppressed LPS-induced NF-κB activation mediated by TLR4. Moreover, the anti-inflammatory effects of quercetin were reversed by the PPAR-γ antagonist of GW9662. In conclusion, these results suggested that quercetin attenuated the production of IL-1β, IL-6, IL-8, and TNF-α in P. gingivalis LPS-treated HGFs by activating PPAR-γ which subsequently suppressed the activation of NF-κB.
1. Introduction
More than 50% of the adult worldwide are affected by periodontal disease which can lead to the destruction of tooth-supporting tissues and even teeth loss [1]. Dental plaque (suggested initiative factor for periodontitis) and host immune response contribute to the pathogenesis of periodontitis [2]. Destruction of periodontal tissues is considered mainly due to an inappropriate host response to dental plaque or the corresponding microbial products such as lipopolysaccharide (LPS). Inflammatory mediators were found elevated accordingly in subjects with severe periodontitis [3]. Porphyromonas gingivalis (P. gingivalis) has been regarded as one of the most important pathogens causing periodontal disease [4]. It leads to the destruction of teeth supporting tissues by inducing host and immune response [5]. LPS from P. gingivalis is considered to be a major virulence factor for periodontal inflammation [6].
Human gingival fibroblasts (HGFs), predominating in gingival connective tissue, are closely related to remodeling of periodontal soft tissues [7]. They can recognize LPS and mediate host immune response in periodontal lesions through interacting with bacteria directly [8]. They have been suggested a vital role in regulating the production of many inflammatory mediators such as interleukin-1β (IL-1β), interleukin-6 (IL-6), interleukin-8 (IL-8), and tumor necrosis factor-α (TNF-α) within gingival tissue [9]. These mediators are found upregulated in both gingival crevicular fluid (GCF) and periodontal tissues of periodontal patients [10]. Therefore, suppressing these inflammatory mediators or blocking the involved signaling pathway may help to restrict the initiation and progression of periodontal disease [11].
As one of the best characterized Toll-like receptors (TLRs) and principal receptor of LPS, TLR4 which can be generated in HGFs is involved in the production of inflammatory mediators and activation of downstream transcription factors such as nuclear factor-kappa B (NF-κB) induced by LPS [12]. The upregulated TLR4 and inflammatory mediators aforementioned contribute to the injury of periodontal tissues [13]. Role of both peroxisome proliferator-activated receptor-γ (PPAR-γ) and liver X receptor α (LXRα) in regulating immune response has been demonstrated previously, and LPS-induced inflammatory responses can be inhibited by the activation of either PPAR-γ or LXRα [14, 15].
There exists a strong positive correlation between periodontal disease and many systemic ones such as cardiopathy [16]. Hence, preventing periodontal disease is beneficial to the health of whole body. Although conventional procedures such as brushing and flossing are generally effective to get rid of dental plaque and manage the progression of periodontal disease, the derived damage such as gingival recession and cementum abrasion has also been reported [17]. In addition, long-term use of antibiotics such as tetracycline and amoxicillin may result in increased drug-resistant resident microbial strains [18]. Since inflammatory responses can be suppressed by reducing inflammatory mediators, bioactive products capable to inhibit these mediators may be helpful to treating periodontal disease [11]. Quercetin (Q), a natural flavonol existing in many food resources such as vegetables and fruits, exhibited long-lasting and strong anti-inflammatory capability in various types of cells in both animal and human models [19]. It suppressed the production of cyclooxygenase (COX) and lipoxygenase (LOX) induced typically by inflammation in vitro [20]. Its anti-inflammatory activity was also reported in vivo [21]. It has been reported to be an effective antimicrobial and anti-inflammatory agent for restricting the inflammation in periodontitis [22, 23]. To the best of our knowledge, the anti-inflammatory effects of quercetin on LPS-stimulated HGFs have not been reported. In the present work, we aimed to investigate the anti-inflammatory effects of quercetin on P. gingivalis LPS-stimulated HGFs and the underlying anti-inflammatory mechanism.
2. Materials and Methods
2.1. Chemicals and Reagents
Quercetin was bought from the National Institute for Pharmaceutical and Biological Products Control (Beijing, China). 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) and bicinchoninic acid (BCA) protein concentration determination kits were purchased from Solarbio Life Science Co., Ltd (Beijing, China). LPS from P. gingivalis was supplied by Invitrogen (St. Louis, CA, USA). Rabbit monoclonal antibodies against IκBα, phosphorylated IκBα (p-IκBα), p65 subunit of nuclear factor-kappa B (NF-κB) (p65), phosphorylated p65 (p-p65) and β-actin were purchased from Cell Signaling Technology Inc (Beverly, MA, USA), with rabbit monoclonal antibodies against PPAR-γ, LXRα and TLR4 from Abcam Inc. (Cambridge, UK). Horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG was provided by Biosynthesis Biotechnolgogy Co., Ltd (Beijing,China). Enhanced chemiluminescence (ECL) kit was bought from Yanxi Co., Ltd (Shanghai, China). And the enzyme-linked immunosorbent assay (ELISA) kits were provided by Westtang Co., Ltd (Shanghai, China). All the other reagents used in the present work are of analytical grade.
2.2. Cell Culture
HGFs were purchased from Bena Culture Collection (BNCC, Jiangsu, China) and cultured in Dulbecco's Modified Eagle Medium (DMEM) containing 10% fatal bovine serum (FBS), penicillin (100 U/mL) and streptomycin (100 μg/mL) at 37°C with 5% CO2. Cells between the 4th and 8th passages were used in the present work. Quercetin was dissolved in dimethyl sulfoxide (DMSO) and diluted using DMEM. This work was performed in five groups: control, LPS, LPS+5 μM Q, LPS+10 μM Q, and LPS+20 μM Q. As to the last three groups employing quercetin and LPS, cells were pretreated with quercetin for 1 h prior to P. gingivalis LPS stimulation, with control and LPS groups pretreated using the same amount of DMSO instead.
2.3. Cell Viability
The effects of LPS and quercetin on cell viability of HGFs were measured by MTT assay based on the reduction of MTT to formazan by mitochondrial succinate dehydrogenase in viable cells. In brief, HGFs were seeded in a 96-well plate at a density of 1×104 cells/well and treated with different concentrations of quercetin (final concentration at 5 μM, 10 μM and 20 μM, respectively) with and without LPS (final concentration at 1 μg/mL) for 24 h. Pre-treatment of quercetin was performed prior to the stimulation of LPS. Then 20 uL MTT (5 mg/mL) was added to each well, followed by incubation for another 4 h at 37°C with 5% CO2. After the medium was removed, 150 uL DMSO was added to each well to dissolve the insoluble formazan crystals in viable cells. Then the formazan concentrations were quantified by measuring the absorbance at 450 nm using a microplate spectraphotometer (Multiskan GO, Thermo, USA). Cell viability was expressed relative to the untreated control group which was regarded as 100%.
2.4. ELISA Assay
Protein levels of IL-1β, L-6, IL-8 and TNF-α in HGFs culture supernatants were determined using corresponding ELISA kits (Westtang Bio-tech, Shanghai, China) in accordance with the producer's instructions. Briefly, HGFs were seeded in a 24-well plate at a density of 2×105 cells/ well. The cells were pretreated with quercetin for 1 h, followed by subsequent stimulation of LPS (final concentration at 1 μg/mL) for 24 h. One hundred μL standards or samples were added to each well of reaction plate and kept at 37°C for 40 min after being mixed fully. Then the reaction plate was washed 5 times with phosphate-buffered saline (PBS) and added 50 μL biotinylated-antibodies working solution for each well and kept at 37°C for 20 min after being mixed fully. After being washed as aforementioned, 100 μL enzyme conjugate working solution was added into each well and kept at 37°C for 10 min after being mixed fully. After being washed as aforementioned, 100 μL TMB solution was added into each well and kept at 37°C for 15 min in dark prior to the addition of 100μL stopping solution into each well. The absorbance at 450 nm was determined using a microplate spectraphotometer (Multiskan GO, Thermo, USA) within 30 min. The levels of IL-1β, L-6, IL-8 and TNF-α in cell culture supernatants, expressed as pg/mL, were quantified based on each corresponding standard curve. Samples were tested in triplicate and each experiment was repeated 3 times independently.
2.5. Real-Time Quantitative PCR (RT-qPCR)
mRNA levels of IL-1β, IL-6, IL-8, TNF-α, IκBα, p65, PPAR-γ, LXRα and TLR4 in HGFs were evaluated by RT-qPCR. HGFs were seeded in a 6-well plate at a density of 1×106 cells/well and pretreated with quercetin (final concentration at 5 μM, 10 μM and 20 μM, respectively) for 1 h, followed by LPS stimulation for 3 h. Then the cells were washed three times using PBS buffer. Total RNA was extracted from HGFs using TRIzol reagent (Invitrogen, CA, USA) and the 1st strand cDNA was synthesized using a commercial kit (Takara, Otsu, Japan) according to the producer's instructions. Next, mRNA levels of the targets aforementioned were evaluated by RT-qPCR using an iQ5 RT-qPCR detection system (Bio-Rad, CA, USA) in a 20 μL reaction system containing approximately 50 ng cDNA, 10 μM of each primer and 10 μL SsoFast EvaGreen Supermix (Bio-Rad Hercules, CA, USA). The RT-qPCR reaction starts with one cycle of 95°C for 30 s, followed by 40 cycles of 95°C for 5 s, 60°C for 30 s, and 72°C for 30 s. After RT-qPCR, a melting curve assay was immediately performed from 60°C to 95°C at a transition rate of 0.5°C/s to avoid the amplification of unspecific products. The primers used in the present work were presented in Table 1. The mRNA levels of these targets were calculated using the 2−△△Ct method and normalized against β-actin which was used as an internal reference gene. The results were expressed as fold changes to control.
Table 1.
Primers used for RT-qPCR.
| Target | Primer Sequence (5'–3') |
|---|---|
| IL-1β | F: TAGGGCTGGCAGAAAGGGAACA |
| R: GTGGGAGCGAATGACAGAGGGT | |
|
| |
| IL-6 | F: CGCCTTCGGTCCAGTTGCC |
| R: GCCAGTGCCTCTTTGCTGCTTT | |
|
| |
| IL-8 | F: CTCTTGGCAGCCTTCCTGATTTC |
| R: TTTTCCTTGGGGTCCAGACAGAG | |
|
| |
| TNF-α | F: AACATCCAACCTTCCCAAACGC |
| R:TGGTCTCCAGATTCCAGATGTCAGG | |
|
| |
| IκBα | F:CACTCCATCCTGAAGGCTACCAACTAC |
| R: ATCAGCACCCAAGGACACCAAA | |
|
| |
| p65 | F: AATGCTGTGCGGCTCTGCTTC |
| R:CCGTGAAATACACCTCAATGTCCTCT | |
|
| |
| PPAR-γ | F: CGCCCAGGTTTGCTGAATGTG |
| R: AGGGAAATGTTGGCAGTGGCTC | |
|
| |
| LXRα | F: TGATGTTCCCACGGATGCTAATG |
| R: TTTGCCCTTCTCAGTCTGTTCCAC | |
|
| |
| TLR4 | F: CACAGACTTGCGGGTTCTACATC |
| R:GGACTTCTAAACCAGCCAGACCTTG | |
|
| |
| β-actin | F:GACTTAGTTGCGTTACACCCTTTCTTG |
| R:CTGTCACCTTCACCGTTCCAGTTTT | |
2.6. Western Blotting Analysis
Protein levels of IκBα, p-IκBα, p65, p-p65, PPAR-γ, LXRα, and TLR4 in HGFs were characterized by Western blotting. HGFs were seeded in a 6-well plate at a density of 1×106 cells/well and pretreated with quercetin (final concentration at 5 μM, 10 μM, and 20 μM, respectively) for 1 h, followed by LPS stimulation for 30 min. The cells were washed three times using cold PBS buffer before the total proteins were isolated using cold radio immunoprecipitation assay (RIPA, 150 mM NaCl, 50 mM Tris–HCl, pH 7.2, 1% Triton X-100, 0.1% SDS) (Solarbio, Beijing, China) lysis buffer supplemented with protease inhibitor of phenylmethylsulfonyl fluoride (PMSF) (final concentration at 1 mM) and phosphatase inhibitor cocktail (Cwbiotech, Jiangsu, China). Protein concentrations were measured using a BCA kit (Solarbio, Beijing, China) based on corresponding standard curves. Samples (30 μg) were loaded to each well and separated on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Then the proteins were transferred onto polyvinylidene difluoride (PVDF) membrane (Solarbio, Beijing, China) at 300 mA under 4°C for 110 min. Under constant shaking, the membrane was blocked with 5% FBS (diluted in PBS) for 2 h and then incubated with corresponding rabbit primary monoclonal antibodies (diluted in 5% FBS) including IκBα (1:500), p-IκBα (1:500), p65 (1:500), p-p65 (1:500), β-actin (1:500), PPAR-γ (1:500), LXRα (1:1000) and TLR4 (1:200) at 4°C overnight. After being washed with cold PBS three times (5 min each), the membrane was incubated with HRP-conjugated secondary anti-rabbit antibodies diluted in 5% FBS (1:2000) for 2 h. After washing the membrane with cold PBS three times (5 min each), protein bands were visualized by ECL with a chemiluminescence gel imaging system (FluorChem Q, ProteinSimple, CA, USA). Band densities of proteins were analyzed using an ImageJ Gel Analysis tool (NIH, Bethesda, MD, United States) and normalized against β-actin which was used as an internal reference. The results were expressed as fold changes in relative densities to control.
2.7. Statistical Analyses
All experiments were performed three times independently and the results were presented as means ± SD. Data was analyzed by one-way ANOVA. Student–Newman–Keuls (SNK) post hoc test was employed to determine the significant difference among groups. All statistical analyses were carried out using SPSS software of version 19.0 (SPSS Inc., Chicago, IL, USA). P < 0.05 was considered statistically significant.
3. Results
3.1. Effects of Quercetin on Cell Viability of HGFs
Effects of quercetin on cell viability of HGFs were evaluated by MTT assay. As is shown in Figure 1, quercetin at 5 μM, 10 μM, and 20 μM, respectively, exerted no significant cytotoxic effects on cell viability of HGFs. Thus the effects of quercetin on LPS-stimulated HGFs in the present work are not attributed to the nonspecific cytotoxicity. Therefore, quercetin at these concentrations was used in the subsequent study.
Figure 1.
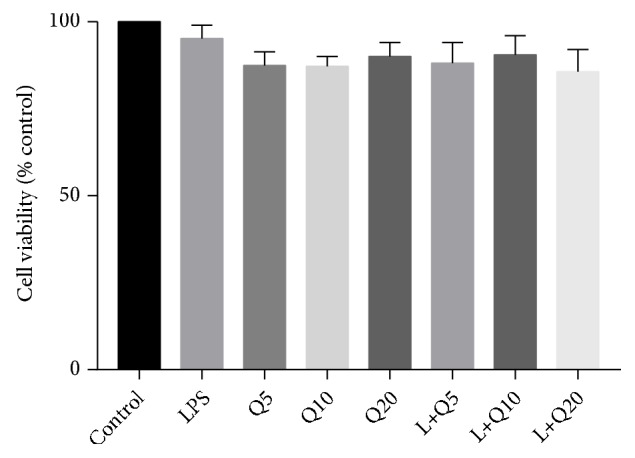
Effects of quercetin on cell viability of HGFs. Cells were cultured with quercetin at different final concentration of 5 μM, 10 μM, and 20 μM, respectively, with and without LPS (final concentration at 1 μg/mL) for 24 h. The values of three independent experiments are presented as means ± SD.
3.2. Effects of Quercetin on Production of IL-1β, IL-6, IL-8, and TNF-α in LPS-Stimulated HGFs
To investigate the anti-inflammatory effects of quercetin on LPS-stimulated HGFs, production of IL-1β, IL-6, IL-8, and TNF-α was detected by ELISA assay (Figure 2). Levels of these inflammatory mediators were significantly upregulated by LPS stimulation, compared with the control group. However, quercetin suppressed the LPS-induced production of IL-1β, IL-6, IL-8, and TNF-α in a dose-dependent manner (Figure 2).
Figure 2.

Quercetin inhibited production of IL-1β, IL-6, IL-8, and TNF-α in LPS-stimulated HGFs. HGFs were pretreated with quercetin at different final concentration of 5 μM, 10 μM and 20 μM for 1 h, respectively, followed by LPS stimulation (final concentration at 1 μg/mL) for 24 h. The values of three independent experiments are presented as means ± SD. ## P<0.01 vs. control group; ∗∗ P<0.01 versus LPS group.
3.3. Effects of Quercetin on mRNA Levels of IL-1β, IL-6, IL-8, TNF-α, IκBα, p65, PPAR-γ, LXRα, and TLR4 in LPS-Stimulated HGFs
mRNA levels of IL-1β, IL-6, IL-8, TNF-α, IκBα, p65, PPAR-γ, LXRα, and TLR4 in LPS-stimulated HGFs were detected by RT-qPCR. As is shown in Figure 3, LPS stimulation significantly upregulated the mRNA levels of IL-1β, IL-6, IL-8, TNF-α, p65, IκBα, and TLR4 but downregulated that of PPAR-γ, which could be suppressed by quercetin dose-dependently.
Figure 3.

Effects of quercetin on mRNA levels of IL-1β, IL-6, IL-8, TNF-α, IκBα, p65, PPAR-γ, LXRα and TLR4 in LPS-stimulated HGFs. HGFs were pretreated with quercetin at different final concentration of 5 μM, 10 μM, and 20 μM for 1 h, respectively, followed by LPS stimulation (final concentration at 1 μg/mL) for 3 h. A: IL-1β; B: IL-6; C: IL-8; D: TNF-α; E: p65; F: IκBα; G: TLR4; H: LXRα; I: PPAR-γ. The values of three independent experiments are presented as means ± SD. #P<0.05 versus control group, ##P<0.01 versus control group; ∗P<0.05 versus LPS group, ∗∗P<0.01 versus LPS group.
3.4. Effects of Quercetin on TLR4 Expression and NF-κB Activation in LPS-Stimulated HGFs
TLR4 serves as the main receptor of LPS. The import role of NF-κB signaling pathway in regulating the production of inflammatory mediators has been reported previously. In the present work, we investigated the effects of quercetin on TLR4 expression and NF-κB activation in LPS-stimulated HGFs. The results exhibited that LPS upregulated TLR4 expression and the phosphorylation of p65 and IκBα significantly, which could be inhibited by quercetin in a dose-dependent manner, however (Figure 4).
Figure 4.

Effects of quercetin on TLR4 expression and NF-κB activation in LPS-stimulated HGFs. HGFs were pretreated with quercetin at different final concentration of 5 μM, 10 μM, and 20 μM for 1 h, respectively, followed by LPS stimulation (final concentration at 1 μg/mL) for 30 min. The values of three independent experiments are presented as means ± SD. #P<0.05 versus control group; ∗∗P<0.01 versus LPS group.
3.5. Effects of Quercetin on Expression of PPAR-γ and LXRα in LPS-Stimulated HGFs
Individual activation of PPAR-γ and LXRα has been suggested to exert anti-inflammatory effects. In the present work, we investigated the effects of quercetin on expression of PPAR-γ and LXRα. The results showed that the expression of PPAR-γ was upregulated significantly, with no significant change in that of LXRα (Figure 5). This suggests that the anti-inflammation effects of quercetin in LPS-stimulated HGFs are through the activation of PPAR-γ.
Figure 5.

Effects of quercetin on expression of PPAR-γ and LXRα in LPS-stimulated HGFs. HGFs were pretreated with quercetin at different final concentration of 5 μM, 10 μM, and 20 μM for 1 h, respectively, followed by LPS stimulation (final concentration at 1 μg/mL) for 30 min. The values of three independent experiments are presented as means ± SD. ∗∗P<0.01 versus LPS group.
3.6. Anti-Inflammatory Effects of Quercetin on LPS-Stimulated HGFs Are PPAR-γ-Dependent
Whether the anti-inflammatory effects of quercetin are PPAR-γ-dependent was investigated. The results showed that the inhibitory effects of quercetin on the production of IL-1β, IL-6, IL-8 and TNF-α were reversed by the PPAR-γ antagonist of GW9662 (Figure 6).
Figure 6.

PPAR-γ antagonist of GW9662 reversed the anti-inflammatory effects of quercetin on LPS-stimulated HGFs. The values of three independent experiments are presented as means ± SD. ##P<0.01 versus control group; ∗∗P<0.01 versus LPS group.
4. Discussion
It has been reported that inflammation plays an important part in the pathogenesis of periodontal disease [24]. Therefore, controlling inflammation is conducive to the treatment of periodontal disease [25]. Previous studies demonstrated the ability of many natural compounds to treat periodontal disease [26, 27]. Quercetin, a natural flavonol rich in fruits, vegetables and some berries, and so on, has exhibited good anti-inflammatory effects previously [20, 21]. In the present work, we investigated the anti-inflammatory effects of quercetin on LPS-stimulated HGF in vitro. The results showed that quercetin attenuated the production of inflammatory mediators of IL-1β, IL-6, IL-8 and TNF-α by suppressing NF-κB signaling pathway.
The Gram-negative bacterium of P. gingivalis serves as the most important etiologic factor in periodontal disease [28]. As a crucial virulence factor for periodontitis, P. gingivalis LPS could induce the release of inflammatory mediators aforementioned in various cells such as HGFs, and result in a series of inflammatory reactions [29]. IL-1β can promote the production of IL-6 and its overproduction may initiate and facilitate the breakdown of connective tissue [30]. IL-6 contributes to the pathogenesis of periodontal diseases via inducing osteoclastogenesis, tissue destruction and bone resorption [31]. As a major chemoattractant, IL-8 can recruit neutrophils which can cause the destruction of normal periodontal tissues by releasing metalloproteinases [32]. Generated in early period of inflammation, TNF-α brings oxidative damage to periodontal tissues due to its most effectiveness in inducing superoxide production in HGFs [33]. Moreover, IL-1β, IL-8 and TNF-α exert dose- and time-dependent synergistic effects on the upregulation of IL-6, hundred times more potent than LPS dose [34]. It has been well documented regarding the association between these inflammatory mediators and the pathogenesis of periodontal disease [35]. And previous studies showed attenuated pathological process of periodontal disease by inhibiting these inflammatory mediators [36]. In the present study, the mRNA and protein levels of IL-1β, IL-6, IL-8 and TNF-α were significantly upregulated by the noncytotoxic concentration of LPS but down-regulated by noncytotoxic concentrations of quercetin, suggesting the anti-inflammatory capability of quercetin. And these inflammatory mediators can be down-regulated effectively by all the three concentrations of quercetin (5 μM, 10 μM, and 20 μM, respectively). Similarly, pretreatment with quercetin at 20 μM for 24 h could significantly attenuated trauma-induced TNF-α increase in H9c2 cells and pretreatment with quercetin at higher than 20 μM exerted cytotoxicity [37]. However, quercetin was also reported to exert anti-inflammatory effects at dosage of as high as 100 μM in pulmonary epithelial (A549) and N9 microglial cells [38, 39].
The vital role of NF-κB signaling pathway in regulating inflammatory responses has been presented previously [40]. Periodontal inflammation may be relieved via the inhibition of NF-κB activation [25]. We investigated the effects of quercetin on NF-κB activation to study its anti-inflammation mechanism. The results showed that quercetin dose-dependently inhibited the LPS-induced NF-κB activation and elevated TLR4 level. It has been reported that TLR4 could mediate the activation of NF-κB in LPS-stimulated HGFs [41], which is consistent with the present work. TLR4 can induce the activation of NF-κB and mitogen-activated protein kinases (MAPK) signaling pathways, both of which can predominately modulate the expression of inflammatory cytokines [40, 42]. In this study, IκBα was used as an indicator of the status of NF-κB activation [43]. Normally, NF-κB is bound to its inhibitors (IκB family) and locates in cytoplasm in an inactive form [44]. Under the stimulation of LPS, IκB proteins are phosphorylated and degraded (Figure 4), and NF-κB p65 may translocate from cytoplasm into the nucleus to upregulate the inflammatory mediators [45]. The present work demonstrated that quercetin suppressed LPS-induced NF-κB activation and IκBα phosphorylation in a dose-dependent manner (Figure 4). Similar anti-inflammatory effects of quercetin via inhibiting NF-κB activation were also reported in other cell types [46, 47]. As to the mechanism of natural bioactive products against inflammation induced by LPS, some researchers support the direct inhibition of p65 nuclear translocation [48], while others claim the suppression of IκBa phosphorylation and degradation [49]. Whether the translocation of NF-κB p65 can also be directly inhibited by quercetin in LPS-stimulated HGFs needs further verification. It is reported that quercetin could down-regulate the expression of inflammatory mediators through suppressing other signaling pathways such as MAPK [50]. Therefore, whether any other signaling pathway plays a role in the anti-inflammatory effects of quercetin in LPS-stimulated HGFs also needs further clarification.
PPAR-γ is a nuclear receptor and capable of regulating bone metabolism and inflammation [51]. The progression of experimental periodontitis in rats was attenuated significantly by the PPAR-γ agonist of rosiglitazone in a previous study [52]. Moreover, PPAR-γ could be activated by many natural products [53–55]. LXRα, a member of nuclear hormone superfamily, has been reported to be an important anti-inflammatory transcription factor involved in the development of inflammatory disease [56]. Agonists of LXRα have been shown to exert anti-inflammatory activity [57]. In addition, the activation of PPAR-γ or LXRα has been demonstrated to attenuate the inflammatory responses induced by LPS via suppressing the activation of NF-κB [58, 59]. To clarify the anti-inflammatory mechanism of quercetin, its effects on the expression of PPAR-γ and LXRα were characterized. The results showed that quercetin significantly upregulated the expression of PPAR-γ, while exerting no significant effects on that of LXRα. In addition, the inhibitory effects of quercetin on the production of IL-1β, IL-6, IL-8 and TNF-α can be reversed by GW9662 which is a PPAR-γ antagonist. This is consistent with a previous study exploring the anti-inflammatory effects of asiatic acid on LPS-stimulated HGFs [60]. Therefore, the results suggest that quercetin attenuates LPS-induced inflammatory response in HGFs by activating PPAR-γ, which subsequently suppressed the activation of NF-κB. Indeed, more blocking assays are needed in our future work to confirm whether quercetin pretreatment could inhibit the nuclear translocation of NF-κB p65 by immunofluorescence staining and the activation of NF-κB using a luciferase reporter gene assay. Moreover, a NF-κB agonist can be used to confirm whether it could reverse the anti-inflammatory effects of quercetin on LPS-stimulated HGFs.
5. Conclusion
In conclusion, our results demonstrated that quercetin attenuated the production of inflammatory mediators of IL-1β, IL-6, IL-8, and TNF-α in P. gingivalis LPS-treated HGFs. The anti-inflammatory mechanism was through activating PPAR-γ, which subsequently suppressed the activation of NF-κB induced by LPS. More in vivo studies are needed to evaluate the anti-inflammatory property of quercetin. The present work suggested a promising therapeutic potential for quercetin in treating periodontal disease.
Acknowledgments
This work was funded by Shandong Provincial Natural Science Foundation, China [Grant no. ZR2017LC021], Projects of Medical and Health Technology Development Program in Shandong Province [Grant no. 2016WS0675], Shandong Provincial Award Foundation for Youth and Middle-aged Scientist [Grant no. BS2010SW034], National Natural Science Foundation of China [Grant no. 81870237], and Science and Technology Innovation Fund for College Students of Weifang Medical University [Grant no. KX2018039].
Contributor Information
Xia Wang, Email: hawangxia@163.com.
Tianliang Zhang, Email: burke1981@163.com.
Data Availability
The data used to support the findings of this study are included within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- 1.Sun H. Y., Jiang H., Du M. Q., et al. The prevalence and associated factors of periodontal disease among 35 to 44-year-old Chinese adults in the 4th national oral health survey. Chinese Journal of Dental Research. 2018;21(4):241–247. doi: 10.3290/j.cjdr.a41082. [DOI] [PubMed] [Google Scholar]
- 2.Holt S., Ebersole J., Felton J., Brunsvold M., Kornman K. Implantation of Bacteroides gingivalis in nonhuman primates initiates progression of periodontitis. Science. 1988;239(4835):55–57. doi: 10.1126/science.3336774. [DOI] [PubMed] [Google Scholar]
- 3.Ardila C. M., Guzmán I. C. Comparison of serum amyloid a protein and C-reactive protein levels as inflammatory markers in periodontitis. Journal of Periodontal & Implant Science. 2015;45(1):14–22. doi: 10.5051/jpis.2015.45.1.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Emani S., Gunjiganur G. V., Mehta D. S. Determination of the antibacterial activity of simvastatin against periodontal pathogens, Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans: An in vitro study. Contemporary Clinical Dentistry. 2014;5(3):377–382. doi: 10.4103/0976-237X.137959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Alhogail S., Suaifan G. A., Bizzarro S., et al. On site visual detection of Porphyromonas gingivalis related periodontitis by using a magnetic-nanobead based assay for gingipains protease biomarkers. Microchimica Acta. 2018;185(2):p. 149. doi: 10.1007/s00604-018-2677-x. [DOI] [PubMed] [Google Scholar]
- 6.Yucel-Lindberg T., Båge T. Inflammatory mediators in the pathogenesis of periodontitis. Expert Reviews in Molecular Medicine. 2013;15, article e7:22. doi: 10.1017/erm.2013.8. [DOI] [PubMed] [Google Scholar]
- 7.Wang Q., Sun L., Gong Z., Du Y. Veratric acid inhibits LPS-induced IL-6 and IL-8 production in human gingival fibroblasts. Inflammation. 2016;39(1):237–242. doi: 10.1007/s10753-015-0243-9. [DOI] [PubMed] [Google Scholar]
- 8.Naruishi K., Nagata T. Biological effects of interleukin-6 on Gingival Fibroblasts: Cytokine regulation in periodontitis. Journal of Cellular Physiology. 2018;233(9):6393–6400. doi: 10.1002/jcp.26521. [DOI] [PubMed] [Google Scholar]
- 9.Scheres N., Crielaard W. Gingival fibroblast responsiveness is differentially affected by Porphyromonas gingivalis : implications for the pathogenesis of periodontitis. Molecular Oral Microbiology. 2013;28(3):204–218. doi: 10.1111/omi.12016. [DOI] [PubMed] [Google Scholar]
- 10.Goutoudi P., Diza E., Arvanitidou M. Effect of periodontal therapy on crevicular fluid interleukin-6 and interleukin-8 levels in chronic periodontitis. International Journal of Dentistry. 2012;2012:8. doi: 10.1155/2012/362905.362905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Muniz F. W., Nogueira S. B., Mendes F. L., et al. The impact of antioxidant agents complimentary to periodontal therapy on oxidative stress and periodontal outcomes: A systematic review. Archives of Oral Biolog. 2015;60(9):1203–1214. doi: 10.1016/j.archoralbio.2015.05.007. [DOI] [PubMed] [Google Scholar]
- 12.Jian C. X., Li M. Z., Zheng W. Y., et al. Tormentic acid inhibits LPS-induced inflammatory response in human gingival fibroblasts via inhibition of TLR4-mediated NF-κB and MAPK signalling pathway. Archives of Oral Biolog. 2015;60(9):1327–1332. doi: 10.1016/j.archoralbio.2015.05.005. [DOI] [PubMed] [Google Scholar]
- 13.Herath T. D. K., Darveau R. P., Seneviratne C. J., Wang C.-Y., Wang Y., Jin L. Tetra- and penta-acylated lipid a structures of Porphyromonas gingivalis LPS differentially activate TLR4-mediated NF-kappa B signal transduction cascade and immunoinflammatory response in human gingival fibroblasts. PLoS ONE. 2013;8(3) doi: 10.1371/journal.pone.0058496.e58496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Heming M., Gran S., Jauch S. L., et al. Peroxisome Proliferator-Activated Receptor-γ Modulates the Response of Macrophages to Lipopolysaccharide and Glucocorticoids. Frontiers in Immunology. 2018;9:p. 893. doi: 10.3389/fimmu.2018.00893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang J., Xiao C., Wei Z., Wang Y., Zhang X., Fu Y. Activation of liver X receptors inhibit LPS-induced inflammatory response in primary bovine mammary epithelial cells. Veterinary Immunology and Immunopathology. 2018;197:87–92. doi: 10.1016/j.vetimm.2018.02.002. [DOI] [PubMed] [Google Scholar]
- 16.Hajishengallis G. Periodontitis: from microbial immune subversion to systemic inflammation. Nature Reviews Immunology. 2014;15(1):30–44. doi: 10.1038/nri3785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Oppermann R. V., Gomes S. C., Cavagni J., Cayana E. G., Conceição E. N. Response to proximal restorations placed either subgingivally or following crown lengthening in patients with no history of periodontal disease. International Journal of Periodontics and Restorative Dentistry. 2016;36(1):116–124. doi: 10.11607/prd.2015. [DOI] [PubMed] [Google Scholar]
- 18.de Souza J. A. C., Rossa Junior C., Garlet G. P., Nogueira A. V. B., Cirelli J. A. Modulation of host cell signaling pathways as a therapeutic approach in periodontal disease. Journal of Applied Oral Science. 2012;20(2):128–138. doi: 10.1590/S1678-77572012000200002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Li Y., Yao J., Han C., et al. Quercetin, inflammation and immunity. Nutrients. 2016;8(3, article no. 167) doi: 10.3390/nu8030167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lee K. M., Hwang M. K., Lee D. E., Lee K. W., Lee H. J. Protective effect of quercetin against arsenite-induced COX-2 expression by targeting PI3K in rat liver epithelial cells. Journal of Agricultural and Food Chemistry. 2010;58(9):5815–5820. doi: 10.1021/jf903698s. [DOI] [PubMed] [Google Scholar]
- 21.Ribeiro D., Freitas M., Tomé S. M., et al. Flavonoids inhibit COX-1 and COX-2 enzymes and cytokine/chemokine production in human whole blood. Inflammation. 2015;38(2):858–870. doi: 10.1007/s10753-014-9995-x. [DOI] [PubMed] [Google Scholar]
- 22.Cheng W.-C., Huang R.-Y., Chiang C.-Y., et al. Ameliorative effect of quercetin on the destruction caused by experimental periodontitis in rats. Journal of Periodontal Research. 2010;45(6):788–795. doi: 10.1111/j.1600-0765.2010.01301.x. [DOI] [PubMed] [Google Scholar]
- 23.Napimoga M. H., Clemente-Napimoga J. T., Macedo C. G., et al. Quercetin inhibits inflammatory bone resorption in a mouse periodontitis model. Journal of Natural Products. 2013;76(12):2316–2321. doi: 10.1021/np400691n. [DOI] [PubMed] [Google Scholar]
- 24.Ji S., Choi Y. S., Choi Y. Bacterial invasion and persistence: critical events in the pathogenesis of periodontitis? Journal of Periodontal Research. 2015;50(5):570–585. doi: 10.1111/jre.12248. [DOI] [PubMed] [Google Scholar]
- 25.Kirkwood K. L., Cirelli J. A., Rogers J. E., Giannobile W. V. Novel host response therapeutic approaches to treat periodontal diseases. Periodontology 2000. 2007;43(1):294–315. doi: 10.1111/j.1600-0757.2006.00166.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Josino Soares D., Walker J., Pignitter M., et al. Pitanga (Eugenia uniflora L.) fruit juice and two major constituents thereof exhibit anti-inflammatory properties in human gingival and oral gum epithelial cells. Food & Function. 2014;5(11):2981–2988. doi: 10.1039/c4fo00509k. [DOI] [PubMed] [Google Scholar]
- 27.Elburki M. S., Rossa C., Guimaraes M. R., et al. A novel chemically modified curcumin reduces severity of experimental periodontal disease in rats: initial observations. Mediators of Inflammation. 2014;2014:10. doi: 10.1155/2014/959471.959471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lin F., Hsiao F., Huang C., et al. Porphyromonas gingivalis GroEL induces osteoclastogenesis of periodontal ligament cells and enhances alveolar bone resorption in rats. PLoS ONE. 2014;9(7) doi: 10.1371/journal.pone.0102450.e102450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ding P. H., Darveau R. P., Wang C. Y., Jin L. 3LPS-binding protein and its interactions with P. gingivalis LPS modulate pro-inflammatory response and Toll-like receptor signaling in human oral keratinocytes. PLoS One. 2017;12(4) doi: 10.1371/journal.pone.0173223.e0173223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Boch J. A., Wara-Aswapati N., Auron P. E. Interleukin 1 signal transduction current concepts and relevance to periodontitis. Journal of Periodontal Research. 2001;80(2):400–407. doi: 10.1177/00220345010800020101. [DOI] [PubMed] [Google Scholar]
- 31.Bakker A. D., Kulkarni R. N., Klein-Nulend J., Lems W. F. IL-6 alters osteocyte signaling toward osteoblasts but not osteoclasts. Journal of Dental Research. 2014;93(4):394–399. doi: 10.1177/0022034514522485. [DOI] [PubMed] [Google Scholar]
- 32.Noh M. K., Jung M., Kim S. H., et al. Assessment of IL-6, IL-8 and TNF-α levels in the gingival tissue of patients with periodontitis. Experimental and Therapeutic Medicine. 2013;6(3):847–851. doi: 10.3892/etm.2013.1222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kim H. K., Park H. R., Lee J. S., Chung T. S., Chung H. Y., Chung J. Down-regulation of iNOS and TNF-α expression by kaempferol via NF-κB inactivation in aged rat gingival tissues. Biogerontology. 2007;8(4):399–408. doi: 10.1007/s10522-007-9083-9. [DOI] [PubMed] [Google Scholar]
- 34.Kent L. W., Rahemtulla F., Michalek S. M. Interleukin (IL)-1 and Porphyromonas gingivalis lipopolysaccharide stimulation of IL-6 production by fibroblasts derived from healthy or periodontally diseased human gingival tissue. Journal of Periodontology. 1999;70(3):274–282. doi: 10.1902/jop.1999.70.3.274. [DOI] [PubMed] [Google Scholar]
- 35.Hou L., Liu C., Rossomando E. F. Crevicular interleukin‐1β in moderate and severe periodontitis patients and the effect of phase I periodontal treatment. Journal of Clinical Periodontology. 1995;22(2):162–167. doi: 10.1111/j.1600-051X.1995.tb00128.x. [DOI] [PubMed] [Google Scholar]
- 36.Yu H., Sun C., Argraves K. M. Periodontal inflammation and alveolar bone loss induced by Aggregatibacter actinomycetemcomitans is attenuated in sphingosine kinase 1-deficient mice. Journal of Periodontal Research. 2016;51(1):38–49. doi: 10.1111/jre.12276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Jing Z., Wang Z., Li X., et al. Protective effect of quercetin on posttraumatic cardiac injury. Scientific Reports. 2016;6:p. 30812. doi: 10.1038/srep30812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Geraets L., Moonen H. J. J., Brauers K., Wouters E. F. M., Bast A., Hageman G. J. Dietary flavones and flavonoles are inhibitors of poly(ADP-ribose) polymerase-1 in pulmonary epithelial cells. Journal of Nutrition. 2007;137(10):2190–2195. doi: 10.1093/jn/137.10.2190. [DOI] [PubMed] [Google Scholar]
- 39.Bureau G., Longpré F., Martinoli M.-G. Resveratrol and quercetin, two natural polyphenols, reduce apoptotic neuronal cell death induced by neuroinflammation. Journal of Neuroscience Research. 2008;86(2):403–410. doi: 10.1002/jnr.21503. [DOI] [PubMed] [Google Scholar]
- 40.Zhao Z., Tang X., Zhao X., et al. Tylvalosin exhibits anti-inflammatory property and attenuates acute lung injury in different models possibly through suppression of NF-κB activation. Biochemical Pharmacology. 2014;90(1):73–87. doi: 10.1016/j.bcp.2014.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Li L., Sun W., Wu T., Lu R., Shi B. Caffeic acid phenethyl ester attenuates lipopolysaccharide-stimulated proinflammatory responses in human gingival fibroblasts via NF-κB and PI3K/Akt signaling pathway. European Journal of Pharmacology. 2017;794:61–68. doi: 10.1016/j.ejphar.2016.11.003. [DOI] [PubMed] [Google Scholar]
- 42.Syam S., Bustamam A., Abdullah R., et al. β Mangostin suppress LPS-induced inflammatory response in RAW 264.7 macrophages in vitro and carrageenan-induced peritonitis in vivo. Journal of Ethnopharmacology. 2014;153(2):435–445. doi: 10.1016/j.jep.2014.02.051. [DOI] [PubMed] [Google Scholar]
- 43.Hsuan C., Hsu H., Tseng W., et al. Glossogyne tenuifolia extract inhibits TNF-α-induced expression of adhesion molecules in human umbilical vein endothelial cells via blocking the NF-kB signaling pathway. Molecules. 2015;20(9):16908–16923. doi: 10.3390/molecules200916908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cai L., Wang Z., Meyer J. M., Ji A., van der Westhuyzen D. R. Macrophage SR-BI regulates LPS-induced pro-inflammatory signaling in mice and isolated macrophages. Journal of Lipid Research. 2012;53(8):1472–1481. doi: 10.1194/jlr.M023234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Manna S. K. Double-edged sword effect of biochanin to inhibit nuclear factor kappaB: Suppression of serine/threonine and tyrosine kinases. Biochemical Pharmacology. 2012;83(10):1383–1392. doi: 10.1016/j.bcp.2012.02.011. [DOI] [PubMed] [Google Scholar]
- 46.Bahar E., Kim J., Yoon H. Quercetin attenuates manganese-induced neuroinflammation by alleviating oxidative stress through regulation of apoptosis, iNOS/NF-κB and HO-1/Nrf2 pathways. International Journal of Molecular Sciences. 2017;18(9) doi: 10.3390/ijms18091989.E1989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chen H., Lu C., Liu H., et al. Quercetin ameliorates imiquimod-induced psoriasis-like skin inflammation in mice via the NF-κB pathway. International Immunopharmacology. 2017;48:110–117. doi: 10.1016/j.intimp.2017.04.022. [DOI] [PubMed] [Google Scholar]
- 48.Cho M. S., Park W. S., Jung W.-K., et al. Caffeic acid phenethyl ester promotes anti-inflammatory effects by inhibiting MAPK and NF-κB signaling in activated HMC-1 human mast cells. Pharmaceutical Biology. 2014;52(7):926–932. doi: 10.3109/13880209.2013.865243. [DOI] [PubMed] [Google Scholar]
- 49.Ha J., Choi H.-S., Lee Y., Lee Z. H., Kim H.-H. Caffeic acid phenethyl ester inhibits osteoclastogenesis by suppressing NFκB and downregulating NFATc1 and c-Fos. International Immunopharmacology. 2009;9(6):774–780. doi: 10.1016/j.intimp.2009.03.001. [DOI] [PubMed] [Google Scholar]
- 50.Zou Y., Wei H. K., Xiang Q., Wang J., Zhou Y., Peng J. Protective effect of quercetin on pig intestinal integrity after transport stress is associated with regulation oxidative status and inflammation. Journal of Veterinary Medical Science. 2016;78(9):1487–1494. doi: 10.1292/jvms.16-0090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bensinger S. J., Tontonoz P. Integration of metabolism and inflammation by lipid-activated nuclear receptors. Nature. 2008;454(7203):470–477. doi: 10.1038/nature07202. [DOI] [PubMed] [Google Scholar]
- 52.Lu Y., Zhou Q., Shi Y., et al. SUMOylation of PPARγ by rosiglitazone prevents LPS-induced NCoR degradation mediating down regulation of chemokines expression in renal proximal tubular cells. PLoS ONE. 2013;8(11) doi: 10.1371/journal.pone.0079815.e79815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Liu Q., Wang C. Y., Liu Z., et al. Hydroxysafflor yellow A suppresses liver fibrosis induced by carbon tetrachloride with high-fat diet by regulating PPAR-γ/p38 MAPK signaling. Pharmaceutical Biology. 2014;52(9):1085–1093. doi: 10.3109/13880209.2013.877491. [DOI] [PubMed] [Google Scholar]
- 54.Wei D., Huang Z. Anti-inflammatory effects of triptolide in LPS-induced acute lung injury in mice. Inflammation. 2014;37(4):1307–1316. doi: 10.1007/s10753-014-9858-5. [DOI] [PubMed] [Google Scholar]
- 55.Lim H. A., Lee E. K., Kim J. M., et al. PPARγ activation by baicalin suppresses NF-κB-mediated inflammation in aged rat kidney. Biogerontology. 2012;13(2):133–145. doi: 10.1007/s10522-011-9361-4. [DOI] [PubMed] [Google Scholar]
- 56.Myhre A. E., Ågren J., Dahle M. K., et al. Liver X receptor is a key regulator of cytokine release in human monocytes. Shock. 2008;29(4):468–474. doi: 10.1097/SHK.0b013e31815073cb. [DOI] [PubMed] [Google Scholar]
- 57.Fu Y., Hu X., Cao Y., Zhang Z., Zhang N. Saikosaponin a inhibits lipopolysaccharide-oxidative stress and inflammation in Human umbilical vein endothelial cells via preventing TLR4 translocation into lipid rafts. Free Radical Biology & Medicine. 2015;89(89):777–785. doi: 10.1016/j.freeradbiomed.2015.10.407. [DOI] [PubMed] [Google Scholar]
- 58.Wang D., Liu M., Wang Y., et al. Synthetic LXR agonist T0901317 attenuates lipopolysaccharide-induced acute lung injury in rats. International Immunopharmacology. 2011;11(12):2098–2103. doi: 10.1016/j.intimp.2011.09.002. [DOI] [PubMed] [Google Scholar]
- 59.Zhang Y.-F., Zou X.-L., Wu J., Yu X.-Q., Yang X. Rosiglitazone, a Peroxisome Proliferator-Activated Receptor (PPAR)-gamma Agonist, Attenuates Inflammation Via NF-kappa B Inhibition in Lipopolysaccharide-Induced Peritonitis. Inflammation. 2015;38(6):2105–2115. doi: 10.1007/s10753-015-0193-2. [DOI] [PubMed] [Google Scholar]
- 60.Hao C., Wu B., Hou Z., et al. Asiatic acid inhibits LPS-induced inflammatory response in human gingival fibroblasts. International Immunopharmacology. 2017;50:313–318. doi: 10.1016/j.intimp.2017.07.005. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data used to support the findings of this study are included within the article.