

Abstract
The cingulate cortex is richly innervated by dopaminergic projections and plays a critical role in attentional control (AC). Evidence indicates that dopamine enhances the neurophysiological signal-to-noise ratio and that dopaminergic tone in the frontal cortex is critically dependent on catechol-O-methyltransferase (COMT). A functional polymorphism (val158met) in the COMT gene accounts for some of the individual variability in executive function mediated by the dorsolateral prefrontal cortex. We explored the effect of this genetic polymorphism on cingulate engagement during a novel AC task. We found that the COMT val158met polymorphism also affects the function of the cingulate during AC. Individuals homozygous for the high-activity valine (“val”) allele show greater activity and poorer performance than val/methionine (“met”) heterozygotes, who in turn show greater activity and poorer performance than individuals homozygous for the low-activity met allele, and these effects are most evident at the highest demand for AC. These results indicate that met allele load and presumably enhanced dopaminergic tone improve the “efficiency” of local circuit processing within the cingulate cortex and thereby its function during AC.
Keywords: attention, cognitive control, catechol-O-methyltransferase, genetics, dopamine, dorsal cingulate, fMRI, variable attentional control task
Introduction
Cognitive control, the capacity to flexibly direct and allocate resources to a goal by selecting and integrating relevant contextual information, is critical for higher mental abilities. Two important aspects of cognitive control include monitoring performance in the face of conflicts and deciding when and how to allocate control and attentional resources and thus biasing lower-level “centers” for the selection of specific features. Increasing the level of perceptual conflict increases the demand for attentional control. Previous functional imaging studies in humans have demonstrated that attentional control is associated with the activity of a network of brain regions, including the dorsal and anterior part of the cingulate cortex (Carter et al., 1998, 2000; Botvinick et al., 1999; Casey et al., 2000; MacDonald et al., 2000; Durston et al., 2003; Weissman et al., 2003; Kerns et al., 2004), the dorsolateral prefrontal cortex (DLPFC) (Botvinick et al., 1999; Carter et al., 2000; MacDonald et al., 2000; Durston et al., 2003; Kerns et al., 2004), and the parietal cortex (Casey et al., 2000; Durston et al., 2003). More specifically, the cingulate cortex is implicated in detecting and/or monitoring conflict between stimuli (Botvinick et al., 1999; Carter et al., 2000; MacDonald et al., 2000; Durston et al., 2003; Kerns et al., 2004), whereas the dorsolateral prefrontal cortex and the parietal cortex are implicated in implementing control (Botvinick et al., 1999; Carter et al., 2000; MacDonald et al., 2000; Durston et al., 2003; Kerns et al., 2004) and orienting attention (Desimone and Duncan, 1995; Casey et al., 2000; Corbetta and Shulman, 2002; Durston et al., 2003). Recently, Durston et al. (2003), using a task with parametrically increasing level of cognitive conflict, demonstrated that the peak of neuronal activity in the cingulate is much more likely to be associated with conflict monitoring than other regions in the network.
The cingulate cortex is richly innervated by dopaminergic projections from the mesencephalic ventrotegmental area (VTA) (Seamans and Yang, 2004). Dysfunction of the dopaminergic system has been implicated in the attentional control deficits that are characteristic of neuropsychiatric disorders such as schizophrenia, Parkinson's disease, and attention deficit hyperactivity disorder, as well as advanced aging (Nieoullon, 2002). Converging evidence from studies in experimental animals and neuroimaging studies in humans indicates that dopamine (DA) spatially tunes the neuronal signal during executive processing and enhances the cortical neurophysiological signal-to-noise ratio (Daniel et al., 1991; Mattay et al., 1996; Seamans et al., 2001a,b; Gonzalez-Burgos et al., 2002; Seamans and Yang, 2004). Additional support for the role of dopamine in optimizing cortical activity comes from recent studies assessing the contribution of catechol-O-methyltransferase (COMT) (Egan et al., 2001; Mattay et al., 2003; Bertolino et al., 2004) and other DA-related genes (Fossella et al., 2002; Fan et al., 2003). Converging evidence indicates that methylation by COMT is an important mechanism for cortical dopamine inactivation in the prefrontal cortex, where the dopamine transporter has relatively little density or impact (Sesack et al., 1998; Moron et al., 2002; Shen et al., 2004). Pharmacological COMT inhibitors, which increase extracellular dopamine in the prefrontal cortex (Tunbridge et al., 2004), also improve working memory performance (Liljequist et al., 1997) and attentional set shifting (Tunbridge et al., 2004) in animals. Similarly, studies in COMT knock-out mice have shown allele load-dependent specific increases in PFC dopamine levels and no change in other catecholamines (Gogos et al., 1998).
Recent evidence also indicates that a functional polymorphism in the gene for COMT accounts for some of the individual variability across healthy controls on tests of executive cognition (Goldberg and Weinberger, 2004). The human COMT gene is located on the long arm of chromosome 22 and contains a common functional polymorphism with a methionine (“met”) to valine (“val”) substitution at codon 158 (val158met) (Mannisto and Kaakkola, 1999). The met allele is associated with low enzymatic activity, whereas the val allele is associated with high activity (Weinshilboum et al., 1999; Chen et al., 2004). Therefore, it has been hypothesized that individuals homozygous for the met allele compared with individuals homozygous for the val allele have increased cortical dopamine signaling (Weinberger et al., 2001) possibly associated with more efficient modulation of cortical processing. Consistent with this model, several groups (Egan et al., 2001; Bilder et al., 2002; Bertolino et al., 2004) have demonstrated in humans that individuals homozygous for the val allele have relatively attenuated performance on tests of executive cognition, along with inefficient prefrontal cortical activity when compared with met carriers. Collectively, these results suggest that COMT plays an important role in modulating DA levels in the PFC and that the val158met polymorphism is a genetic factor accounting for variability across individuals in prefrontal cortex function.
Previously, the effects of this polymorphism have been primarily investigated on the function of the DLPFC. Its effects on the function of other regions in the prefrontal cortex, such as the cingulate, have been unexplored. The present study was designed to evaluate the potential effect of the COMT val158met polymorphism on the response of the cingulate during attentional control processing, when conflicting information is presented. To this end, we developed a “variable attentional control” (VAC) task, which was specifically designed to vary the level of attentional control-based global-local dimensions and congruence of the stimuli across three levels of attentional control (low, intermediate, and high). Using functional magnetic resonance imaging (fMRI) during this task, we found main effects of task difficulty in several brain regions prominently implicated in attentional control processes, including the dorsal cingulate, the dorsolateral prefrontal, and the parietal cortices. This was associated with a significant detriment in performance, reflected by a decrease in accuracy and an increase in reaction time (RT). Based on the putative cortical tuning effects of dopamine, we hypothesized that individuals homozygous for the COMT val allele (i.e., subjects with relatively lower cortical dopamine levels) would exhibit an inefficient neurophysiological response in the cingulate, along with poor performance relative to individuals homozygous for the met allele. Our results support this hypothesis.
Materials and Methods
Subjects
Twenty-three normal subjects [12 males; age ± SE, 26.3 ± 1.1; intelligence quotient (IQ) ± SE, 104.7 ± 1.6; eight val/val (five males; age ± SE, 26.7 ± 1.9; IQ ± SE, 103.0 ± 2.9), eight val/met (five males; age ± SE, 25.0 ± 1.1; IQ ± SE, 105.4 ± 3.7), and seven met/met (two males; age ± SE, 27.3 ± 2.9; IQ ± SE, 104.8 ± 2.6)], who had undergone extensive clinical evaluation, were recruited. Inclusion criteria were the absence of any neurological and psychiatric disorders and any other medical condition, absence of any pharmacological treatment that could influence cerebral metabolism or blood flow, absence of drug abuse, age of <45 years, and handedness >0.8, as measured with the Edinburgh Handedness Inventory (Oldfield, 1971). Genotype groups were matched for age, IQ, handedness, and gender (χ = 2.24; df = 2; p = 0.3).
All of these subjects underwent fMRI while performing the VAC task. All subjects gave written informed consent to the study after the procedure was fully explained to them. The protocol was approved by the National Institute of Mental Health Institutional Review Board.
Variable attentional control task
Subjects performed a behavioral task (Fig. 1) that was specifically designed to increase perceptual conflict. Briefly, each stimulus was composed of arrows of three different sizes (1 large, 6 medium, and 42 small arrows) pointing either to the right or to the left; seven small arrows were embedded in each medium-sized arrow, and six medium-sized arrows were embedded in the large arrow. The direction of the arrows in each particular size was always the same
Figure 1.

VAC task. See Materials and Methods for explanation.
Subjects were instructed by a cue word (BIG, MEDIUM, or SMALL) displayed above each stimulus to press a button corresponding to the direction of the large, medium, or small arrows (either right or left). To increase the required level of conflict monitoring, the direction of the arrows was congruent or incongruent across all three sizes. This resulted in the following conditions: (1) Low level of attentional control (LOW): all three sizes of arrows were congruent in direction with each other, and the stimuli were cued with the word BIG. (2) Intermediate level of attentional control (INT): the big arrow was incongruent in direction to the small and the medium arrows; the cue was either BIG or SMALL. (3) High level of attentional control (HIGH): the medium-sized arrows were incongruent in direction to the big and the small arrows; the cue was either SMALL or MEDIUM. In addition, a simple bold arrow pointing to either the left or right was used as a sensorimotor control condition with no conflict.
Subjects were instructed to press as quickly and accurately as possible and to move their thumb to a small plastic knob placed in the center of the array of buttons between responses. All subjects were trained on the task before the fMRI session. Only subjects who demonstrated a linear RT response to attentional control were included.
Each stimulus was presented for 800 ms, and the order of the stimuli was randomly distributed across the session (Friston et al., 1999). The randomization of the sequence of stimulus presentation was established according to the stochastic design option in Statistical Parametric Mapping 99 (SPM99) software (Wellcome Department of Imaging Neuroscience, University College London, London, UK). The total number of stimuli was 241: 50 HIGH (25 stimuli of each of the two stimulus types that subtended this level of conflict), 68 INT (34 stimuli of each of the two stimulus types that subtended this level of conflict), 57 LOW, and 66 simple bold arrows (sensorimotor control condition) (Fig. 1); the total duration of the task was 10 min, 8 s. A fixation cross-hair was presented during the interstimulus interval, which ranged from 2000 to 6000 ms.
Blood oxygen level-dependent fMRI
Blood oxygen level-dependent (BOLD) fMRI was performed on a General Electric (Milwaukee, WI) Signa 3T scanner (gradient-echo planar-imaging sequence; repetition time, 2000 ms; echo time, 28 ms; 26 interleaved slices; thickness, 4 mm; gap, 1 mm; voxel size, 3.75 × 3.75 × 5; scan repetitions, 300; flip angle, 90°; field of view, 24 cm; matrix, 64 × 64) while subjects performed the VAC task. The first four scans were discarded to allow for signal saturation. Stimuli were presented via a back-projection system, and the responses were recorded through a fiberoptic response box, which allowed the measurement of the accuracy and reaction time for each trial.
Data analysis
Behavioral data. Behavioral data [accuracy (percentage correct) and reaction time for correct responses] were compared for each level of attentional control using analysis of covariance (ANCOVA) with COMT genotype as a grouping factor and gender as a covariate of no interest. We opted to covary for gender because several studies have shown a sexually dimorphic effect of COMT on dopamine levels and behavior (Gogos et al., 1998; Karayiorgou et al., 1999; Alsobrook et al., 2002; Goudreau et al., 2002; Rybakowski et al., 2002; Kremer et al., 2003; Chen et al., 2004; Domschke et al., 2004; Ono et al., 2004; Sazci et al., 2004; Shifman et al., 2004); furthermore, estrogen hormones have been shown to modulate COMT activity (Weinshilboum et al., 1999). Fisher's least significant difference (LSD) method was used for post hoc analyses. Accuracy was measured as the percentage of correct responses relative to the total number of stimuli for each level of attentional control.
fMRI analysis. Whole-brain image analysis was completed using SPM99 (www.fil.ion.ucl.ac.uk/spm) and the event-related module. Images for each subject were realigned to the first volume in the time series to correct for head motion, slice time corrected, and spatially normalized into a standard stereotactic space (Montreal Neurological Institute template) using a 12 parameter affine model. Spatial smoothing was also applied with a Gaussian filter set at 12 mm full-width at half-maximum to minimize noise and residual differences in gyral anatomy. After realignment, data sets were also screened for high quality (scan stability) as demonstrated by small motion correction (<2 mm translation, <1.5° rotation).
fMRI responses were modeled using a canonical hemodynamic response function (hrf), ratio normalized to the whole-brain global mean to control for systematic differences in global activity, and temporally filtered using an SPM default high-pass filter based on the frequency of the events and an hrf-shaped low-pass filter. Vectors were created for each condition using the timing of correct responses for each stimulus type. The timing of the incorrect responses and the residual movement were also modeled as regressors of no interest. Predetermined condition effects at each voxel were created using a t statistic, producing a statistical image for the contrasts: HIGH > baseline (HIGH), INT > baseline (INT), LOW > baseline (LOW), and HIGH > INT > LOW (HIGH > INT > LOW).
A second-level random-effects model that accounts for both scan-to-scan and subject-to-subject variability was then used for group analyses.
To evaluate the main effect of the increasing level of attentional control, ANCOVA with gender as covariate [p < 0.05; family-wise error (FWE) rate, corrected, k = 8] was performed across the three levels of attentional control (HIGH > INT > LOW). To evaluate the main effect of COMT genotype, ANCOVA with gender as covariate (p < 0.05; FWE rate, corrected, k = 8) was performed across the three genotypes (val/val > val/met > met/met). To evaluate the interaction between COMT genotype and increasing level of attentional control, ANCOVA with gender as covariate (p < 0.05; FWE rate, corrected, k = 8) across genotypes (val/val > val/met > met/met and vice versa) was performed comparing the contrast images from the contrast HIGH > INT > LOW. These analyses were constrained by a mask based on the main effect of the task obtained by combining the HIGH, INT, and LOW group activation maps (p < 0.001).
To compare brain activation across COMT genotypes within each level of attentional control (HIGH, INT, and LOW) ANCOVA across genotypes (val/val > val/met > met/met and vice versa), with gender as a covariate, was performed. The analysis was constrained by a mask generated by combining the group activation maps for the three genotypes for each of the three levels of attentional control (p < 0.001, uncorrected).
Brodmann's areas were assigned to activated clusters using the Talairach Daemon Client (http://ric.uthscsa.edu/projects/talairachdaemon.html) after converting the Montreal Neurological Institute coordinates of the local maxima in the activated clusters to Talairach coordinates (http://www.mrc-cbu.cam.ac.uk/Imaging/Common/mnispace.shtml).
Region-of-interest analysis. Additional analysis was performed using MarsBaR to extract signal change from the cingulate region, the area that showed a significant interaction of genotype by increasing level of attentional control. The cingulate region of interest (ROI) was identified in the activated cluster from the contrast evaluating the interaction between level of attentional control and genotype (val/val > val/met > met/met). This ROI was applied to each group activation map (HIGH > baseline, INT > baseline, and LOW > baseline) for each genotype.
Additionally, to explore the contribution of neuronal activity in the DLPFC to behavior (see below), we also extracted BOLD signal change from the significantly activated cluster in this region that showed a main effect of increase in level of attentional control.
Regression analysis. To examine the relationship between COMT genotype, mean BOLD response in the cingulate cortex and DLPFC (extracted as described above), IQ, and accuracy, we performed a hierarchical multiple-regression analysis in which variables are entered (or removed) from the linear equation on the basis of their ability to improve R2 at each successive step in the model (SAS Institute, Cary, NC).
Genotype determination. DNA was extracted by standard methods. COMT val108/158met genotype was determined by 5′ exonuclease allelic discrimination TaqMan assay (Livak, 1999) that uses the 5′ nuclease activity of TaqDNA polymerase to detect a fluorescent reporter signal generated after PCRs.
Results
Behavioral data
Accuracy
There was a main effect of increasing level of attentional control (F(2,38) = 9.25; p < 0.0006), a main effect of genotype approaching significance (F(2,19) = 3.39; p = 0.055), and an interaction between level of attentional control and genotype (F(4,38) = 4.90; p < 0.003) (Fig. 2). Post hoc analysis with Fisher's LSD of the main effect of increasing level of attentional control showed a significant difference across all three levels (HIGH > INT, p < 0.000001; HIGH > LOW, p < 0.000001; INT > LOW, p < 0.0006).
Figure 2.

Effect of COMT genotype on accuracy during the VAC task. A statistically significant effect of COMT genotype was found on accuracy (percentage correct) only at the highest level of attentional control. A met allele load effect was found with individuals homozygous for the met allele performing significantly better than the val/met (vm) heterozygotes and the val/val (vv) homozygotes. mm, met/met. Error bars represent SEM.
Post hoc analysis of the main effect of genotype showed a significant difference between homozygote met/met and val/val genotypes (met/met > val/val; p < 0.022) and no difference between met/met and val/met (p = 0.30) or between val/met and val/val (p = 0.14). Post hoc analysis of the significant interaction between increasing level of attentional control and genotype indicated a statistically significant difference for accuracy to HIGH between met/met and val/val (met/met > val/val; p < 0.0009), between val/met and val/val (val/met > val/val; p < 0.022), and a trend toward significance (p = 0.1) between met/met and val/met. No significant differences between genotypes were found in accuracy for INT and LOW levels of conflict (all p values > 0.6).
Reaction time
There was a main effect of increasing level of attentional control (F(2,38) = 21.90; p < 0.00001), no effect of genotype (F(2,19) = 1.11; p > 0.35), and no interaction between demand of attentional control and genotype (F(4,38) = 0.20; p > 0.93) (Table 1). Post hoc analysis with Fisher's LSD of the main effect of level of conflict showed a statistically significant difference across all three levels (all p values < 0.000001).
Table 1.
Effect of COMT genotype on RT (in milliseconds ± SEM) during the VAC task
|
COMT genotype |
LOW |
INT |
HIGH |
|---|---|---|---|
| val/val | 770.9 ± 60.5 | 929.3 ± 47.2 | 1114.8 ± 43.9 |
| val/met | 691.6 ± 35.6 | 839.1 ± 51.3 | 1016.6 ± 58.8 |
|
met/met
|
726.5 ± 26.0 |
864.1 ± 23.8 |
1074.3 ± 42.8 |
Although there was a significant main effect of increasing level of attentional control on RT, there was no significant effect of COMT genotype or a significant interaction between demand of attentional control and COMT genotype on RT. For details, see Results.
Imaging data
Effect of increasing level of attentional control
There was a main effect of level of attentional control with greater activation in HIGH in several brain regions, including the parietal cortex [Brodmann's area 7/40 (BA7/40)] bilaterally, the right dorsolateral prefrontal cortex (BA46), the medial vallum of the prefrontal cortex (supplementary motor cortex, BA6), the dorsal cingulate (BA32) bilaterally, and the right premotor cortex (BA6) (Fig. 3A).
Figure 3.
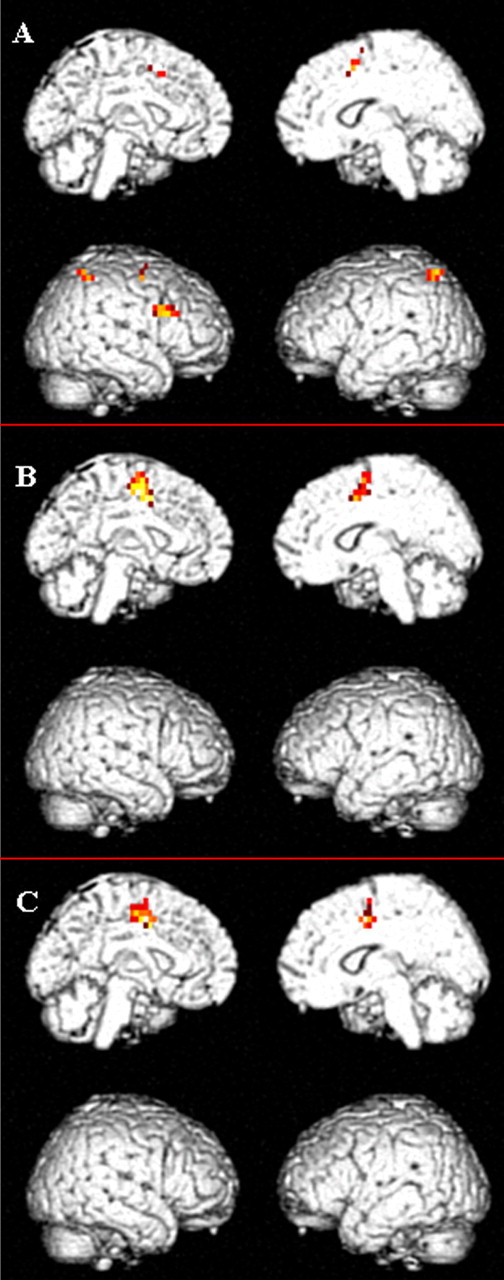
Effect of attentional load and COMT genotype on BOLD response during the VAC task. There were main effects of increasing level of attentional control (A) and COMT genotype (B) and a significant interaction between increasing level of attentional control and COMT genotype (C) on brain activation during the VAC task (p < 0.05; FWE rate, corrected). A, In addition to the dorsal cingulate, the dorsolateral prefrontal cortex, premotor, and supplementary motor areas showed a main effect of increasing level of attentional control. B, The main effect of COMT genotype (val/val > val/met > met/met) was restricted only to the dorsal cingulate [local maxima in BA24 (-4, -2, 41)]. Note that the local maxima for the cluster that showed a significant COMT genotype by level of attentional control is similar to the one that showed a main effect of COMT genotype [BA24 (-4, -2, 41].
Effect of COMT genotype
There was a main effect of COMT genotype in the dorsal cingulate [BA24 (x, y, z: -4, -2, 41); Z = 6.01; k = 66] with greatest activation for val/val individuals, followed by val/met, and then met/met individuals. There were no significant differences in the inverse contrast (met/met > val/met > val/val) (Fig. 3B).
Interaction of COMT genotype by level of attentional control
There was a significant interaction between increasing level of attentional control (contrast, HIGH > INT > LOW) and COMT genotype (val/val > val/met > met/met) in the dorsal cingulate [BA24 (-4, -2, 41); Z = 3.62; k = 57]. There was no significant difference in the inverse contrast (met/met > val/met > val/val) (Fig. 3C).
Effect of COMT genotype within each level of attentional control
High attentional control demand. There was a main effect of genotype (val/val > val/met > met/met) that was again restricted only to the dorsal cingulate region [local maxima in BA24 (-8, -2, 41); k = 23; Z = 3.33; p < 0.001] (Fig. 4A). No significant differences were found on the inverse contrast (met/met > val/met > val/val).
Figure 4.

Effect of COMT genotype on BOLD response at intermediate and high demands of the VAC task. There was a main effect of COMT genotype (val/val > val/met > met/met) on brain activation during the VAC task (p < 0.001, uncorrected). A, High demand [local maxima in BA24 (-8, -2, 41)]. B, Intermediate demand [local maxima in BA24 (-4, -2, 41)].
Intermediate attentional control demand. ANCOVA showed increased activation across genotypes (val/val > val/met > met/met) only in the dorsal cingulate [local maxima in BA24 (-4, 2, 41); k = 9; Z = 3.62; p < 0.001] (Fig. 4B). No significant differences were found on the inverse contrast (met/met > val/met > val/val).
Low attentional control demand. ANCOVA at a significance threshold level similar to that used above did not show differential activation across genotypes in both directions (either val/val > val/met > met/met or met/met > val/met > val/val). However, at a lower statistical threshold (p < 0.002), differential activation was found only in the dorsal cingulate [local maxima in BA24 (-4-5, 46); k = 9; Z = 2.81] in the contrast val/val > val/met > met/met (Fig. 5).
Figure 5.

BOLD signal change in dorsal cingulate during the VAC task. For illustrative purposes, BOLD signal change is plotted from a representative voxel in the cluster in BA24/32 that showed a significant interaction for COMT genotype by increasing level of attentional control. vv, val/val; vm, val/met; mm, met/met.
Regression analysis
To assess the relative predictive power of regional neurophysiology and COMT genotype on accuracy performance during the critical high-demand condition, we regressed the following variables onto accuracy: COMT genotype, IQ, DLPFC BOLD signal, cingulate BOLD signal, and interaction terms composed of genotype by cingulate and genotype by DLPFC BOLD signal. Using a variant of stepwise regression designed to maximize “explained variance,” we found that only two variables entered at significant levels: COMT genotype, which accounted for 33% of the accuracy variance at HIGH (standardized β = 0.85; p = 0.0017), and BOLD signal change in the dorsal cingulate, which accounted for 9% of the accuracy variance at HIGH (standardized β = 0.47; p = 0.059).
For both INT and LOW conditions, no variables or combination of variables accounted for a significant proportion of the variance (R2 < 0.12). Most importantly, BOLD signal change in the dorsolateral prefrontal cortex did not contribute significantly to the variance in accuracy across individuals during this task at any level of load (p = 0.2).
Discussion
We report convergent findings that implicate an effect of the COMT val158met functional polymorphism on the function of the normal dorsal cingulate cortex. First, there is a met allele load effect on the performance of the VAC task, a task that demands an increasing level of attentional control for which the cingulate has been particularly implicated. Individuals homozygous for the high-activity val allele perform relatively worse on this task (decreased accuracy) when compared with val/met heterozygotes, which in turn perform worse when compared with individuals homozygous for the low-activity met allele. It is noteworthy that these groups of normal volunteers were matched on age, IQ, and other demographic factors.
The imaging data show that, although increasing the level of task difficulty is associated with an increase in the activity of a network of brain regions that include the dorsal cingulate cortex, the dorsolateral prefrontal cortex, and the parietal cortex, the effect of the COMT val158met polymorphism during the task is especially robust in the dorsal cingulate, where activity and performance covary. Here again, there is a met allele load effect, with individuals homozygous for the val allele showing much more activity than the val/met heterozygotes, who in turn show greater activity than the individuals homozygous for the met allele. This suggests that met allele load, presumably by modulating dopaminergic tone, improves the “efficiency” in the physiological response of the cingulate cortex during conflict monitoring. Interestingly, this effect of the COMT val158met polymorphism is most evident at the highest level of conflict, presumably when demands for efficient processing are greatest.
The effect of the COMT val158met polymorphism on accuracy during the VAC task is consistent with the basic science literature detailed above (Liljequist et al., 1997; Tunbridge et al., 2004) and with several studies in healthy humans and in patients with schizophrenia, albeit in the context of working memory tasks (Egan et al., 2001; Bilder et al., 2002; Malhotra et al., 2002; Goldberg et al., 2003; Bertolino et al., 2004). The lack of the COMT genotype effect on behavioral reaction time may appear counter-intuitive, but it is consistent with recent studies in animals suggesting that the effect of dopamine signaling in the medial prefrontal cortex during attention is more evident for discriminative performance than for speed (Chudasama and Robbins, 2004). In addition, results from lesion studies suggest that lesions of the dorsal pregenual cingulate cortex induce deficits in response accuracy (Passetti et al., 2002; Chudasama et al., 2003), whereas lesions of the ventral infralimbic region of the medial prefrontal cortex increase premature responding during attention. These results are also consistent with data from human studies (Goldberg et al., 2003; Bertolino et al., 2004) suggesting that response latency is not specifically influenced by dopaminergic signaling in cingulate during cognition. Consistent with the behavioral data, our imaging data show a specific and exclusive effect of COMT val158met genotype on activity in the dorsal cingulate region during this specific task. Given the abundant dopaminergic projections to the cingulate, we had hypothesized that COMT genotype would affect level of activity in this region. In the dorsal cingulate, val homozygotes have higher activation at the highest level of conflict relative to met homozygotes. Thus, at the highest level of conflict, val homozygotes increased activation in the dorsal cingulate despite impaired performance (accuracy); this region of the dorsal cingulate has been implicated previously in conflict during tasks involving hand movements (Paus, 2001). Collectively, the imaging and behavioral data suggest less-efficient cortical processing of the stimuli and less-efficient allocation of attentional resources in individuals homozygous for the val allele. We use the term efficiency here to represent an empirical inverse relationship between performance and BOLD response, which has been reported in the context of a variety of cognitive control tasks (Rypma and D'Esposito, 1999; Callicott et al., 2000).
The effect of the COMT val158met polymorphism became more apparent as demand for attentional control increased. These findings are consistent with those of Mattay et al. (2003) and Bertolino et al. (2004), albeit in the DLPFC in the context of working memory performance, showing inefficient information processing primarily at high task demand in individuals homozygous for the val allele relative to the other two groups. Not surprisingly, they suggest that COMT genotype effects are most evident when processing load stress-specific neural systems, perhaps limiting redundancy in the processing parameters.
Several studies have found error-related activity in the cingulate cortex, suggesting that it is implicated in cognitive processes associated with erroneous performance. In this regard, there is an ongoing debate in the literature about the specific function of the cingulate during erroneous performance. It is unlikely that the effect of the COMT val158met genotype on cingulate activity that we observed is related to error-related activity, because we analyzed activity related to only correct responses. Although the effect of COMT genotype on error-related activity in the cingulate poses an interesting question, the number of errors committed, although statistically different across the genotype groups, were few (∼4%), precluding the exploration of this issue in a statistically meaningful way.
In the present study, we did not find an effect of the COMT val158met polymorphism on DLPFC activity. Previous studies using working memory tasks have shown an effect of this polymorphism primarily in the DLPFC, with no significant effect in other brain regions in the working memory network, including the cingulate and parietal regions. In this study, we used a task that was specifically designed to manipulate the level of conflict monitoring required and thereby the level of activity in the cingulate. In this experimental context, we find an effect of the COMT val158met polymorphism only on the activity of the cingulate. We also find that brain activity predicts task performance only in the cingulate. We believe that this supports the notion that the conflict-monitoring aspects of this task are the critical processes engaged and that the load on DLPFC processing is minimal. Evidence suggests that the dorsal cingulate is primarily involved in handling conflict (Carter et al., 1998, 2000; Botvinick et al., 1999; Casey et al., 2000; MacDonald et al., 2000; Durston et al., 2003; Weissman et al., 2003; Kerns et al., 2004) and that the dorsolateral prefrontal cortex is primarily involved in executive cognition, updating (Goldberg et al., 2003), and working memory function (Callicott et al., 1999). Furthermore, we also found, using hierarchical multiple-regression analysis, that only COMT genotype and BOLD signal change in the cingulate contributed significantly to the variance in accuracy across individuals during this task; DLPFC activity did not contribute significantly to this variance. Together, these data suggest that COMT val158met polymorphism modulates dopaminergic signaling and thereby brain activity in a task- and region-specific manner. This notion is consistent with the observations of Mattay et al. (1996), who noted that amphetamine, a monoaminergic drug, in humans modulated activity in the prefrontal cortex during executive cognition and in the hippocampus during episodic memory processing, and of Chudasama and Robbins (2004), who reported that cortical infusions of a dopamine receptor D1 agonist [SKF 81297 ((±)-6-chloro-7,8-dihydroxy-1-phenyl-2,3,4,5-tetrahydro-1M-3-benzazepine] in rats differentially regulate attention and working memory performance.
In summary, these results strongly suggest that, in the context of attentional control, variation in dopamine signaling mediated by COMT is critical for processes that engage specific subregions of the prefrontal cortex (e.g., conflict management in the dorsal cingulate). These results add to evidence from pharmacological studies that dopaminergic modulation influences the efficiency of executive attention. Furthermore, they may represent a viable approach to understanding individual differences in attentional control and possibly neuropsychiatric disorders with complex genetic etiologies that show attentional deficits.
Footnotes
We thank Roberta Rasetti, Guilna Alce, Brad Zoltick, and Jessica Cohen for research assistance.
Correspondence should be addressed to Daniel R. Weinberger, Clinical Brain Disorders Branch, National Institute of Mental Health, National Institutes of Health, Department of Health and Human Services, Building 10, Center Drive, Room 4S-235, Bethesda, MD 20892-1379. E-mail: weinberd@mail.nih.gov.
Copyright © 2005 Society for Neuroscience 0270-6474/05/255038-08$15.00/0
G.B. and V.S.M. contributed equally to this work.
References
- Alsobrook II JP, Zohar AH, Leboyer M, Chabane N, Ebstein RP, Pauls DL (2002) Association between the COMT locus and obsessive-compulsive disorder in females but not males. Am J Med Genet 114: 116-120. [DOI] [PubMed] [Google Scholar]
- Bertolino A, Caforio G, Blasi G, De Candia M, Latorre V, Petruzzella V, Altamura M, Nappi G, Papa S, Callicott JH, Mattay VS, Bellomo A, Scarabino T, Weinberger DR, Nardini M (2004) Interaction of COMT (Val(108/158)Met) genotype and olanzapine treatment on prefrontal cortical function in patients with schizophrenia. Am J Psychiatry 161: 1798-1805. [DOI] [PubMed] [Google Scholar]
- Bilder RM, Volavka J, Czobor P, Malhotra AK, Kennedy JL, Ni X, Goldman RS, Hoptman MJ, Sheitman B, Lindenmayer JP, Citrome L, McEvoy JP, Kunz M, Chakos M, Cooper TB, Lieberman JA (2002) Neurocognitive correlates of the COMT Val(158)Met polymorphism in chronic schizophrenia. Biol Psychiatry 52: 701-707. [DOI] [PubMed] [Google Scholar]
- Botvinick M, Nystrom LE, Fissell K, Carter CS, Cohen JD (1999) Conflict monitoring versus selection-for-action in anterior cingulate cortex. Nature 402: 179-181. [DOI] [PubMed] [Google Scholar]
- Callicott JH, Mattay VS, Bertolino A, Finn K, Coppola R, Frank JA, Goldberg TE, Weinberger DR (1999) Physiological characteristics of capacity constraints in working memory as revealed by functional MRI. Cereb Cortex 9: 20-26. [DOI] [PubMed] [Google Scholar]
- Callicott JH, Bertolino A, Mattay VS, Langheim FJ, Duyn J, Coppola R, Goldberg TE, Weinberger DR (2000) Physiological dysfunction of the dorsolateral prefrontal cortex in schizophrenia revisited. Cereb Cortex 10: 1078-1092. [DOI] [PubMed] [Google Scholar]
- Carter CS, Braver TS, Barch DM, Botvinick MM, Noll D, Cohen JD (1998) Anterior cingulate cortex, error detection, and the online monitoring of performance. Science 280: 747-749. [DOI] [PubMed] [Google Scholar]
- Carter CS, Macdonald AM, Botvinick M, Ross LL, Stenger VA, Noll D, Cohen JD (2000) Parsing executive processes: strategic vs. evaluative functions of the anterior cingulate cortex. Proc Natl Acad Sci USA 97: 1944-1948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey BJ, Thomas KM, Welsh TF, Badgaiyan RD, Eccard CH, Jennings JR, Crone EA (2000) Dissociation of response conflict, attentional selection, and expectancy with functional magnetic resonance imaging. Proc Natl Acad Sci USA 97: 8728-8733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J, Lipska BK, Halim N, Ma QD, Matsumoto M, Melhem S, Kolachana BS, Hyde TM, Herman MM, Apud J, Egan MF, Kleinman JE, Weinberger DR (2004) Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): effects on mRNA, protein, and enzyme activity in postmortem human brain. Am J Hum Genet 75: 807-821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chudasama Y, Robbins TW (2004) Dopaminergic modulation of visual attention and working memory in the rodent prefrontal cortex. Neuropsychopharmacology 29: 1628-1636. [DOI] [PubMed] [Google Scholar]
- Chudasama Y, Passetti F, Rhodes SE, Lopian D, Desai A, Robbins TW (2003) Dissociable aspects of performance on the 5-choice serial reaction time task following lesions of the dorsal anterior cingulate, infralimbic and orbitofrontal cortex in the rat: differential effects on selectivity, impulsivity and compulsivity. Behav Brain Res 146: 105-119. [DOI] [PubMed] [Google Scholar]
- Corbetta M, Shulman GL (2002) Control of goal-directed and stimulus-driven attention in the brain. Nat Rev Neurosci 3: 201-215. [DOI] [PubMed] [Google Scholar]
- Daniel DG, Goldberg TE, Gibbons RD, Weinberger DR (1991) Lack of a bimodal distribution of ventricular size in schizophrenia: a Gaussian mixture analysis of 1056 cases and controls. Biol Psychiatry 30: 887-903. [DOI] [PubMed] [Google Scholar]
- Desimone R, Duncan J (1995) Neural mechanisms of selective visual attention. Annu Rev Neurosci 18: 193-222. [DOI] [PubMed] [Google Scholar]
- Domschke K, Freitag CM, Kuhlenbaumer G, Schirmacher A, Sand P, Nyhuis P, Jacob C, Fritze J, Franke P, Rietschel M, Garritsen HS, Fimmers R, Nothen MM, Lesch KP, Stogbauer F, Deckert J (2004) Association of the functional V158M catechol-O-methyl-transferase polymorphism with panic disorder in women. Int J Neuropsychopharmacol 7: 183-188. [DOI] [PubMed] [Google Scholar]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, Watts R, Ulug AM, Casey BJ (2003) Parametric manipulation of conflict and response competition using rapid mixed-trial event-related fMRI. NeuroImage 20: 2135-2141. [DOI] [PubMed] [Google Scholar]
- Egan MF, Goldberg TE, Kolachana BS, Callicott JH, Mazzanti CM, Straub RE, Goldman D, Weinberger DR (2001) Effect of COMT Val108/158 Met genotype on frontal lobe function and risk for schizophrenia. Proc Natl Acad Sci USA 98: 6917-6922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan J, Fossella J, Sommer T, Wu Y, Posner MI (2003) Mapping the genetic variation of executive attention onto brain activity. Proc Natl Acad Sci USA 100: 7406-7411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fossella J, Sommer T, Fan J, Wu Y, Swanson JM, Pfaff DW, Posner MI (2002) Assessing the molecular genetics of attention networks. BMC Neurosci 3: 14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friston KJ, Zarahn E, Josephs O, Henson RN, Dale AM (1999) Stochastic designs in event-related fMRI. NeuroImage 10: 607-619. [DOI] [PubMed] [Google Scholar]
- Gogos JA, Morgan M, Luine V, Santha M, Ogawa S, Pfaff D, Karayiorgou M (1998) Catechol-O-methyltransferase-deficient mice exhibit sexually dimorphic changes in catecholamine levels and behavior. Proc Natl Acad Sci USA 95: 9991-9996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg TE, Weinberger DR (2004) Genes and the parsing of cognitive processes. Trends Cogn Sci 8: 325-335. [DOI] [PubMed] [Google Scholar]
- Goldberg TE, Egan MF, Gscheidle T, Coppola R, Weickert T, Kolachana BS, Goldman D, Weinberger DR (2003) Executive subprocesses in working memory: relationship to catechol-O-methyltransferase Val158Met genotype and schizophrenia. Arch Gen Psychiatry 60: 889-896. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Burgos G, Kroner S, Krimer LS, Seamans JK, Urban NN, Henze DA, Lewis DA, Barrionuevo G (2002) Dopamine modulation of neuronal function in the monkey prefrontal cortex. Physiol Behav 77: 537-543. [DOI] [PubMed] [Google Scholar]
- Goudreau JL, Maraganore DM, Farrer MJ, Lesnick TG, Singleton AB, Bower JH, Hardy JA, Rocca WA (2002) Case-control study of dopamine transporter-1, monoamine oxidase-B, and catechol-O-methyl transferase polymorphisms in Parkinson's disease. Mov Disord 17: 1305-1311. [DOI] [PubMed] [Google Scholar]
- Karayiorgou M, Sobin C, Blundell ML, Galke BL, Malinova L, Goldberg P, Ott J, Gogos JA (1999) Family-based association studies support a sexually dimorphic effect of COMT and MAOA on genetic susceptibility to obsessive-compulsive disorder. Biol Psychiatry 45: 1178-1189. [DOI] [PubMed] [Google Scholar]
- Kerns JG, Cohen JD, MacDonald III AW, Cho RY, Stenger VA, Carter CS (2004) Anterior cingulate conflict monitoring and adjustments in control. Science 303: 1023-1026. [DOI] [PubMed] [Google Scholar]
- Kremer I, Pinto M, Murad I, Muhaheed M, Bannoura I, Muller DJ, Schulze TG, Reshef A, Blanaru M, Gathas S, Goichman R, Rietschel M, Dobrusin M, Bachner-Melman R, Nemanov L, Belmaker RH, Maier W, Ebstein RP (2003) Family-based and case-control study of catechol-O-methyltransferase in schizophrenia among Palestinian Arabs. Am J Med Genet B Neuropsychiatr Genet 119: 35-39. [DOI] [PubMed] [Google Scholar]
- Liljequist R, Haapalinna A, Ahlander M, Li YH, Mannisto PT (1997) Catechol O-methyltransferase inhibitor tolcapone has minor influence on performance in experimental memory models in rats. Behav Brain Res 82: 195-202. [DOI] [PubMed] [Google Scholar]
- Livak KJ (1999) Allelic discrimination using fluorogenic probes and the 5′ nuclease assay. Genet Anal 14: 143-149. [DOI] [PubMed] [Google Scholar]
- MacDonald III AW, Cohen JD, Stenger VA, Carter CS (2000) Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science 288: 1835-1838. [DOI] [PubMed] [Google Scholar]
- Malhotra AK, Kestler LJ, Mazzanti C, Bates JA, Goldberg T, Goldman D (2002) A functional polymorphism in the COMT gene and performance on a test of prefrontal cognition. Am J Psychiatry 159: 652-654. [DOI] [PubMed] [Google Scholar]
- Mannisto PT, Kaakkola S (1999) Catechol-O-methyltransferase (COMT): biochemistry, molecular biology, pharmacology, and clinical efficacy of the new selective COMT inhibitors. Pharmacol Rev 51: 593-628. [PubMed] [Google Scholar]
- Mattay VS, Berman KF, Ostrem JL, Esposito G, Van Horn JD, Bigelow LB, Weinberger DR (1996) Dextroamphetamine enhances “neural network-specific” physiological signals: a positron-emission tomography rCBF study. J Neurosci 16: 4816-4822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattay VS, Goldberg TE, Fera F, Hariri AR, Tessitore A, Egan MF, Kolachana B, Callicott JH, Weinberger DR (2003) Catechol O-methyltransferase val158-met genotype and individual variation in the brain response to amphetamine. Proc Natl Acad Sci USA 100: 6186-6191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moron JA, Brockington A, Wise RA, Rocha BA, Hope BT (2002) Dopamine uptake through the norepinephrine transporter in brain regions with low levels of the dopamine transporter: evidence from knock-out mouse lines. J Neurosci 22: 389-395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieoullon A (2002) Dopamine and the regulation of cognition and attention. Prog Neurobiol 67: 53-83. [DOI] [PubMed] [Google Scholar]
- Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9: 97-113. [DOI] [PubMed] [Google Scholar]
- Ono H, Shirakawa O, Nushida H, Ueno Y, Maeda K (2004) Association between catechol-O-methyltransferase functional polymorphism and male suicide completers. Neuropsychopharmacology 29: 1374-1377. [DOI] [PubMed] [Google Scholar]
- Passetti F, Chudasama Y, Robbins TW (2002) The frontal cortex of the rat and visual attentional performance: dissociable functions of distinct medial prefrontal subregions. Cereb Cortex 12: 1254-1268. [DOI] [PubMed] [Google Scholar]
- Paus T (2001) Primate anterior cingulate cortex: where motor control, drive and cognition interface. Nat Rev Neurosci 2: 417-424. [DOI] [PubMed] [Google Scholar]
- Rybakowski JK, Borkowska A, Czerski PM, Hauser J (2002) Eye movement disturbances in schizophrenia and a polymorphism of catechol-O-methyltransferase gene. Psychiatry Res 113: 49-57. [DOI] [PubMed] [Google Scholar]
- Rypma B, D'Esposito M (1999) The roles of prefrontal brain regions in components of working memory: effects of memory load and individual differences. Proc Natl Acad Sci USA 96: 6558-6563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sazci A, Ergul E, Kucukali I, Kilic G, Kaya G, Kara I (2004) Catechol-O-methyltransferase gene Val108/158Met polymorphism, and susceptibility to schizophrenia: association is more significant in women. Brain Res Mol Brain Res 132: 51-56. [DOI] [PubMed] [Google Scholar]
- Seamans JK, Yang CR (2004) The principal features and mechanisms of dopamine modulation in the prefrontal cortex. Prog Neurobiol 74: 1-58. [DOI] [PubMed] [Google Scholar]
- Seamans JK, Durstewitz D, Christie BR, Stevens CF, Sejnowski TJ (2001a) Dopamine D1/D5 receptor modulation of excitatory synaptic inputs to layer V prefrontal cortex neurons. Proc Natl Acad Sci USA 98: 301-306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seamans JK, Gorelova N, Durstewitz D, Yang CR (2001b) Bidirectional dopamine modulation of GABAergic inhibition in prefrontal cortical pyramidal neurons. J Neurosci 21: 3628-3638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sesack SR, Hawrylak VA, Matus C, Guido MA, Levey AI (1998) Dopamine axon varicosities in the prelimbic division of the rat prefrontal cortex exhibit sparse immunoreactivity for the dopamine transporter. J Neurosci 18: 2697-2708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen HW, Hagino Y, Kobayashi H, Shinohara-Tanaka K, Ikeda K, Yamamoto H, Yamamoto T, Lesch KP, Murphy DL, Hall FS, Uhl GR, Sora I (2004) Regional differences in extracellular dopamine and serotonin assessed by in vivo microdialysis in mice lacking dopamine and/or serotonin transporters. Neuropsychopharmacology 29: 1790-1799. [DOI] [PubMed] [Google Scholar]
- Shifman S, Bronstein M, Sternfeld M, Pisante A, Weizman A, Reznik I, Spivak B, Grisaru N, Karp L, Schiffer R, Kotler M, Strous RD, Swartz-Vanetik M, Knobler HY, Shinar E, Yakir B, Zak NB, Darvasi A (2004) COMT: a common susceptibility gene in bipolar disorder and schizophrenia. Am J Med Genet B Neuropsychiatr Genet 128B: 61-64. [DOI] [PubMed] [Google Scholar]
- Tunbridge EM, Bannerman DM, Sharp T, Harrison PJ (2004) Catechol-O-methyltransferase inhibition improves set-shifting performance and elevates stimulated dopamine release in the rat prefrontal cortex. J Neurosci 24: 5331-5335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinberger DR, Egan MF, Bertolino A, Callicott JH, Mattay VS, Lipska BK, Berman KF, Goldberg TE (2001) Prefrontal neurons and the genetics of schizophrenia. Biol Psychiatry 50: 825-844. [DOI] [PubMed] [Google Scholar]
- Weinshilboum RM, Otterness DM, Szumlanski CL (1999) Methylation pharmacogenetics: catechol O-methyltransferase, thiopurine methyltransferase, and histamine N-methyltransferase. Annu Rev Pharmacol Toxicol 39: 19-52. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Giesbrecht B, Song AW, Mangun GR, Woldorff MG (2003) Conflict monitoring in the human anterior cingulate cortex during selective attention to global and local object features. NeuroImage 19: 1361-1368. [DOI] [PubMed] [Google Scholar]