

Abstract
In manual action, the relationship between a given motor command and the ensuing movement depends on the dynamics of both the arm and hand-held objects. Skilled performance relies on the brain learning both these dynamics, and previous studies have examined how people adapt to novel loads applied to either the hand or the arm. In this study, we ask whether these different kinds of load are represented independently as a result of changes in cutaneous feedback and hand-arm coordination. We used a robotic apparatus that could either apply forces to an object held in the subject's hand or directly to the segments of the arm. We tested whether subjects could retain learning of a force field applied to the hand after subsequently experiencing the opposing field applied to the arm (or vice versa), or whether retrograde interference would be observed. In separate experiments, we used force fields and torque fields that were linearly related to either hand or joint velocities, respectively. Our finding of complete interference between opposing fields suggests that loads applied to the arm and hand are not represented independently by the sensorimotor system. This interference occurred despite markedly different cutaneous inputs that were directly related to the movement task. This result suggests that the brain represents dynamics independently of these sensory inputs. In addition, we found that the rate at which subjects adapted to a given force field, specified either in hand or joint coordinates, was independent of whether the forces were applied to the hand or arm segments.
Keywords: adaptation, motor learning, motor control, arm movement, sensorimotor integration, limb dynamics
Introduction
Our ability to rapidly learn and recall a single motor task stands in sharp distinction to our inability to learn two opposing motor tasks experienced in succession. After a single exposure to a motor task, performance on the task exceeds that of novices even after a gap of several months (Shadmehr and Brashers-Krug, 1997). However, subsequent exposure to an opposing task returns performance on the first to novice levels. Learning the second task effectively extinguishes memory of the first, a phenomenon known as retrograde interference (Baddeley, 1986, 1992). In studies of motor learning, such interference has been demonstrated between opposing force fields applied to the hand (Brashers-Krug et al., 1996; Gandolfo et al., 1996; Shadmehr and Brashers-Krug, 1997; Caithness et al., 2004) or arm (Krakauer et al., 1999) during reaching movements. These studies have shown that when subjects sequentially adapt to two opposing force fields, A then B, learning of B interferes with memory of A such that, when subjects are retested on A at a later time, performance is no better than when A was first experienced.
Interference between opposing tasks suggests that the two tasks compete for the same memory resources (Krakauer et al., 1999; Wigmore et al., 2002). Previous studies have sought to determine whether opposing tasks can be represented independently in memory by attempting to identify factors that remove or attenuate interference. For example, it has been demonstrated that subjects can learn opposing force fields if the posture of the arm is altered between the two fields (Gandolfo et al., 1996) or if contextual cues are provided appropriately (Krouchev and Kalaska, 2003; Osu et al., 2004).
Here, we tested whether loads applied to the hand and arm are learned and represented separately by testing whether the interference normally observed between opposing force fields would be removed or attenuated if one field were applied to the hand and the other to the arm. Because the loads applied to the hand and arm are mechanically equivalent and require similar adjustments to shoulder and elbow muscles, we might expect opposing fields to still interfere when applied differently. However, the load-dependent cutaneous feedback experienced when moving against loads applied to the hand and arm is markedly different. Thus, we were able to test whether changes to functionally significant sensory feedback could facilitate independent learning despite the mechanical similarity in loads applied to the hand and arm. Another reason why independent learning might be expected is that loads applied to the hand require coordinated actions of the hand, wrist, and arm, whereas loads applied to the arm segments do not. We also tested, in different groups, force fields that linearly depended on hand velocity and torque fields that linearly depended on joint angular velocities. Thus, we were also able to evaluate the hypothesis that loads applied to the hand can be learned more rapidly when defined in hand coordinates compared with joint coordinates and, similarly, whether loads applied to the arm can be learned more quickly when defined in joint coordinates compared with hand coordinates.
Materials and Methods
Subjects. Eighty healthy right-handed subjects (19-27 years of age) participated in the study after providing written consent. The study was approved by the Queen's University Ethics Board. Subjects were randomly assigned to one of 10 groups (see below, Procedure) with eight in each group.
Apparatus. Subjects made reaching movements to visual targets while using a robotic device (KINARM; BKIN Technologies, Kingston, Ontario, Canada) in one of two configurations as shown in Figure 1 (Singh and Scott, 2003). In the arm configuration, torques were applied directly to the right upper arm and forearm via fiberglass braces attached to the robot. In the hand configuration, the arm braces were removed, a handle was attached to the distal link, and the subjects rested their elbow on an air sled. A Plexiglas plate was also attached to the proximal link to provide a low-friction surface for the air sled, ensuring that no torques were transmitted to the subject's arm. In the arm configuration, subjects made a fist and maintained their hand and wrist in approximately the same posture as they would when holding the handle in the hand configuration.
Figure 1.

Configurations of the KINARM robot used to apply forces to the upper and lower arm segments via arm troughs (arm configuration shown to the left) or to the hand via a handle (hand configuration shown to the right). In the hand configuration, the elbow was supported by a cushioned air puck, and subjects grasped a handle. The upper and lower arm segments did not otherwise contact the apparatus.
Procedure. Subjects performed out-and-back reaching movements, with the arm in a horizontal plane, to one of eight targets. In the first experiment, the targets were located 10 cm from the central start position and were evenly distributed in different directions. In the second experiment, targets were defined to be equidistant in resultant joint amplitude, 0.245 radians (rad; 14°) from the (same) central start position, and were distributed equally in joint space. Resultant joint amplitude was defined as the square root of the squares of the shoulder and elbow joint amplitudes.
Visual feedback of hand position, the start position, and the targets was presented in the same plane as the arm using an overhead projector and a semisilvered mirror. These were represented as white, cyan, and green circular cursors of diameter 1.2, 1.4, and 1.4 cm, respectively. To start a trial, subjects were required to return the hand cursor to the start position. A target appeared, and subjects were required to move to the target and back as accurately as they could while maintaining a movement duration of ∼500 ms. The start position cursor flashed red 500 ms after movement start, and subjects were instructed to time their movements so as to arrive back at the start position at the moment it turned red.
For the first experiment, subjects in four test groups initially performed a session of 240 trials (30 blocks of movements to each of eight targets presented in random order) under a velocity-dependent rotary force field (field A) followed, 5 min later, by a second session of 240 trials in the opposite force field (field B). One day later, these subjects experienced a third session with field A to test for retention of learning. For two test groups, fields A and B were both applied to either the hand (AhBhAh) or the arm (AaBaAa). For the other two groups, field A and B were applied in different configurations (AhBaAh and AaBhAa). Previous studies using velocity-dependent force fields applied at the hand have shown that if the opposite field B is not experienced after A, then subjects show strong retention of learning of A when tested 1 d later (Brashers-Krug et al., 1996; Shadmehr and Brashers-Krug, 1997; Caithness et al., 2004). However, because such retention has not yet been documented for velocity-dependent force fields applied to the arm, we included a control group who completed the first and third sessions only, both in the arm configuration (Aa-Aa). The second experiment was the same as the first except that we applied a torque field that linearly depended on joint angular velocity rather than a force field that linearly depended on hand velocity (see below). As in the first experiment, subjects in four test groups were examined (AhBhAh, AaBaAa, AhBaAh, and AaBhAa). We also included a control group who completed the first and third sessions only, both in the hand configuration (Ah-Ah). All subjects first completed a practice session of 64 trials with the same robot configuration and targets as used in the first test session but with the robot motors turned off. In each test session, subjects were provided with a rest period every 56 trials, and trials 230 and 240 were catch trials (with a null force field), allowing us to assess the extent of learning.
In the first experiment, the torques generated by the KINARM robot were servo-controlled at 2 kHz to create a velocity-dependent rotary force field, F, according to the following equation:
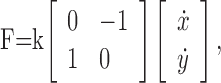 |
where F is the vector of forces in the horizontal plane acting at the handle or the center of the subject's fist in the hand and arm configurations, respectively, ẋ and ẏ are the hand velocities in the horizontal plane, and k represents the viscosity of the force field. In the first and third sessions, k was 6 N/ms-1, and in the second session, k was -6 N/ms-1. In the second experiment, we applied an angular velocity-dependent rotary torque field, T, according to the following equation:
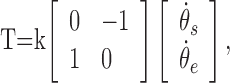 |
where T is the vector of torques acting on the arm,  and
and 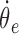 are shoulder and elbow angular velocities, and k is the angular viscosity that was set to 0.65 N/rad·s-1 for sessions 1 and 3 and -0.65 N/rad·s-1 for session 2. Note that when the elbow is in 90° flexion, this torque field is similar to the force field described above, because hand motion in one direction will lead to torques that tend to move the hand in the orthogonal direction. However, for other elbow joint angles, the two fields are distinct. Encoders on the KINARM motors measured joint angles at 200 Hz. The lengths of each subject's upper and lower arms were used to calculate the corresponding hand position.
are shoulder and elbow angular velocities, and k is the angular viscosity that was set to 0.65 N/rad·s-1 for sessions 1 and 3 and -0.65 N/rad·s-1 for session 2. Note that when the elbow is in 90° flexion, this torque field is similar to the force field described above, because hand motion in one direction will lead to torques that tend to move the hand in the orthogonal direction. However, for other elbow joint angles, the two fields are distinct. Encoders on the KINARM motors measured joint angles at 200 Hz. The lengths of each subject's upper and lower arms were used to calculate the corresponding hand position.
Analysis. Performance was measured by the area enclosed by the hand or joint movement path in experiments 1 and 2, respectively. We analyzed learning within a session by fitting the following falling exponential model to individual subject's handor joint-path areas: E(i) = α + βe-(i / τ), where E is the expected area on trial i relative to the start of a session, α is a scalar offset representing a subject's performance learning plateau, β is the gain, and τ represents the time constant of adaptation. Repeated-measures and between-subjects ANOVAs were used to compare retention levels across various sessions. p ≤ 0.05 was considered to be statistically significant.
Results
To assess performance in the first experiment, we calculated the area enclosed by the path of the hand for each movement (Wigmore et al., 2002; Caithness et al., 2004). Figure 2 shows mean hand-path area (averaged across subjects) as a function of block (averaged across the eight trials) for each group and for the first and third sessions. When subjects first encountered the force field in session 1, hand trajectories were perturbed resulting in large hand-path areas. However, subjects gradually adapted so as to produce approximately straight hand paths as indicated by small hand-path areas. Subjects who experienced the same field, applied to the arm, on day 1 and day 2 with no intervening opposing field (Aa-Aa), demonstrated clear retention of learning. That is, initial performance (average of blocks 2 and 3) was significantly better (F(1,7) = 38.57; p < 0.001) in session 3 than in session 1. In contrast, when an intervening, opposite force field was experienced in the same configuration (AhBhAh and AaBaAa), no retention of learning of the first field was observed. Specifically, initial performance in session 3 was no better (F(1,8) < 0.01; p = 0.928 for AhBhAh) or even worse (F(1,8) = 7.55; p = 0.025 for AaBaAa) than in session 1. Moreover, the initial performance in session 3 was significantly worse for both these groups compared with the control group (p < 0.005 in both cases). Critically, when different configurations were used for the opposing fields (AhBaAh and AaBhAa), subjects also failed to retain learning of the first field when retested on day 2. Initial performance in session 3 was no better than in session 1 (p > 0.52 in both cases). In other words, clear interference between the opposing force fields was observed despite the fact that one was applied to the hand and the other to the arm.
Figure 2.

Mean hand-path area results for experiment 1, in which the imposed force fields depended on the velocity of the hand. The field was applied either to the hand via a handle (subscript h) or to the arm segments via an exoskeleton (subscript a). Data are shown for sessions 1 and 3 (first and second A). A block is a set of reaches to all eight targets. Each dot represents an average across subjects, and the vertical lines represent 1 SE. Exponential curves are fitted to each session of each group. The elevated dots at the right of each panel represent the average of two catch trials delivered in blocks 29 and 30.
To test for possible differences in learning rate across the two robot configurations, we compared the first sessions of subjects in the AaBaAa and AaBhAa groups with those of subjects in the AhBhAh and AhBaAh groups. To obtain learning rates, we fit an exponential model (see Materials and Methods) to the hand-path area curves for each subject and session and determined the time constant in each case. A between-subjects ANOVA revealed no difference in learning rate (F(1,32) = 2.47; p = 0.13). To assess how well the force fields in each configuration were learned, we compared hand-path areas in catch trials (with the force field turned off) delivered late in session 1 (blocks 29 and 300) (Fig. 2). We found no differences between configurations (p > 0.05 in both cases). These results indicate that the robot configuration did not affect how quickly or how well the force fields were learned in session 1.
Although hand-path area decreased in both sessions 1 and 3 for all ABA groups and in session 1 for the A-A control group, this does not imply that hand paths necessarily became straighter. It is possible to have low path areas and curved hand paths if the out-and-back movement segments follow the same path. To verify that hand paths became straighter, we computed, for every trial, the mean perpendicular displacement (MPD) between the hand path and the straight line joining the start position and the target (averaging across all data points along the path). We then computed the average MPD for each block. To estimate the initial and final path curvature in each session, we computed, for each subject, the average MPD of blocks 2 and 3 and the average of blocks 27 and 28, respectively. The average initial and final MPD values across all sessions (excluding session 3 of the control group) were 0.68 and 0.42 cm, respectively. Separate repeated-measures ANOVAs revealed that the final MPD was reliably smaller than the initial MPD for sessions 1 and 3 of all ABA groups and for session 1 of the control group (p < 0.04 in all nine cases). Thus, hand paths become straighter across blocks within these sessions.
Because we applied forces to both the hand and arm in this study, we felt it important to test force fields specified in both hand and joint coordinates. Therefore, we ran a second experiment that was identical to the first except that we used torque fields, and target locations depended linearly on joint angular velocity (see Materials and Methods). Performance was assessed in terms of joint-path area, the area enclosed by the path of the movement in joint space. In this experiment, we also ran four test groups that differed in terms of the configurations they experienced across the three sessions, and we will refer to these groups using the same notation as in the first experiment.
Figure 3 shows mean joint-path area as a function of block for each group and for the first and third sessions. As in the first experiment, no retention of learning is evident in any of the four test groups. That is, initial performance (average of blocks 2 and 3) in session 3 was no better than in session 1 (p > 0.10 in all four cases). We also compared groups AhBhAh and AhBaAh directly with a control group, Ah-Ah, who did not perform session 2. The initial performance in session 3 was worse for both of these groups compared with the control group (p < 0.01 in both cases). In contrast to the four test groups, clear retention of learning was observed in the control group in that initial performance was better in session 3 than in session 1 (F(1,8) = 18.81; p = 0.003). Thus, the retention of learning over successive exposures to the same force field, observed for fields defined in hand coordinates, is also seen for fields defined in joint coordinates. As in the first experiment, we found no effect of configuration on the learning rates in the first session (F(1,30) = 1.78; p = 0.19).
Figure 3.

Joint-path area results for experiment 2, in which the imposed force fields depended on the angular velocities of the joints. Labels are as in Figure 2. deg, Degree.
As in the first experiment, we again compared initial and final MPD values. The average initial and final MPD values across all sessions (excluding session 3 of the A-A control group) were 0.54 and 0.32 cm, respectively. Separate repeated-measures ANOVAs revealed that the final MPD was reliably smaller than the initial MPD for sessions 1 and 3 of all ABA groups and for session 1 of the control group (p < 0.002 in all nine cases). Thus, hand paths become straighter across blocks within these sessions.
The catch trials shown in Figure 3, as well as Figure 2, clearly indicate that subjects learned the force fields applied to the hand or arm segments. In the second experiment (Fig. 3), path areas for the catch trials in session 3 were smaller than for the catch trials in session 1. We are not sure why this should be the case but can think of two possibilities. First, the order of targets within blocks was fully randomized, and therefore the catch trial movement directions varied across subjects and sessions. Thus, differences between sessions 1 and 3 could have arisen by chance variations in movement direction. Another possibility is that arm stiffness was slightly greater in session 3 than session 1. One argument against that latter possibility is that we did not see systematic differences between sessions in the first experiment (Fig. 2).
Discussion
The main aim of our study was to test whether loads applied to the hand and arm can be learned and represented separately by testing whether the interference normally observed between opposing force fields (Brashers-Krug et al., 1996; Shadmehr and Brashers-Krug, 1997; Krakauer et al., 1999; Caithness et al., 2004) would be removed or attenuated if one field were applied to the hand and the other to the arm. In agreement with these previous studies, we found complete inference between opposing force fields when the two fields were applied either to the hand or the arm. This was true for force fields defined in hand Cartesian coordinates as well as for fields defined in joint angular coordinates. However, we observed equally strong and complete interference between opposing loads when one was applied to hand and the other was applied to the arm segments. Thus, our study does not support the idea that loads applied to the hand and arm can be represented independently in motor memory. We also tested whether faster learning would be observed if the coordinate system in which the load is applied (hand or joint coordinates) and the method of application (hand or arm) were compatible. We found that learning rate was not affected by such compatibility.
Why might one expect loads applied to the hand and arm to be encoded in different ways? One possibility is that loads applied to the hand engage sensory and motor processes specialized for manual object manipulation. In object manipulation, motor actions of the arm and hand must be precisely coupled to appropriately adjust grip forces in the face of movement-dependent loads, and the motor system receives important sensory information from tactile receptors in the glabrous skin of the volar aspect of the hand (Johansson and Westling, 1984; Westling and Johansson, 1987; Flanagan and Tresilian, 1994; Flanagan and Wing, 1997). In addition, muscles stabilizing the wrist must also be coordinated with muscles driving the arm when moving hand-held loads. Given these additional sensorimotor requirements for moving hand-held loads, it is conceivable that tasks involving loads applied to the hand would be represented independently from otherwise similar tasks involving loads applied to the arm. However, our finding of strong and complete interference between opposing force fields applied to the hand and arm suggests that these loads share a common representation in motor memory. Numerous studies have shown that many neurons in primary motor cortex are active during arm movements and change their activity when moving with a mechanical load (Ashe, 1997; Scott, 2003). Whereas load representations in primary motor cortex change across behaviors contexts (Kurtzer et al., 2005), neurons that are responsive to loads applied at a single joint during reaching will also respond to more complex loads, applied to multiple joints, that incorporate the single-joint load (Gribble and Scott, 2002). Given the latter findings, the present results suggest that neurons responding to mechanical loads applied to the limb during reaching would respond in a similar manner when loads are applied to the hand.
The current study also tests whether the natural coupling between the type of load (i.e., joint- or hand-based) and sensory feedback of motor performance influences the rate of motor learning. Learning to move joint-based loads, such as those experienced when first wearing elbow or shoulder pads in hockey, must make use of sensory feedback from the proximal limb. However, when learning to manipulate hand-based loads such as tools, additional sensory feedback is provided by the hand. Through natural experience with these types of loads, one might expect that it would be more difficult to learn loads defined in hand coordinates when they are applied directly to the limb segments and vice versa. However, we have shown that learning rate was not affected by compatibility between the coordinate system in which the load is represented (hand or joint coordinates) and the method of application (hand or arm). Thus, for example, torque fields that linearly depended on joint angular velocity were learned equally quickly whether applied to the hand or the arm.
Skilled motor performance requires that the brain represent the dynamics of both the arm and hand-held objects (Kawato, 1999; Wolpert and Flanagan, 2001). It is interesting to speculate whether loads applied to the hand are treated as an external object, whereas loads applied to the arm segments are treated as a change in arm dynamics. If this were the case, our results would suggest a common coding of object and arm dynamics. Such a claim would be consistent with recent neurophysiological studies of tool use showing that body representations can expand to incorporate hand-held objects and illustrating shared neural processing to control arm movements with and without a hand-held tool (Iriki et al., 1996). However, it is not clear that the brain would interpret loads applied to the arm segments as a change in arm dynamics rather than an external object.
Previous studies have sought to determine whether opposing tasks can be represented independently in memory by attempting to identify factors that remove or attenuate interference. For example, it has been demonstrated that subjects can learn opposing force fields if the posture of the arm is altered between the two fields (Gandolfo et al., 1996). However, arbitrary sensory cues, such as color or changes in proprioceptive information from joints not involved in the movement task, are not effective contextual cues promoting independent learning of opposing force fields applied in alternation (Gandolfo et al., 1996; Seidler et al., 2001; Krouchev and Kalaska, 2003; Osu et al., 2004). Here, we tested whether functional (and nonarbitrary) changes in sensory feedback, namely changes in cutaneous feedback when applying loads to the hand versus arm segments, can facilitate independent learning of opposing dynamic tasks. Our results indicate that movement-related changes in cutaneous sensory feedback are not effective cues allowing for independent learning of opposing dynamic tasks. This suggests that internal models of force fields represent the dynamics of the task independent of the sensory consequences of the task.
Footnotes
This work was supported by the Human Frontier Science Program, the Natural Sciences and Engineering Research Council of Canada, the Canadian Institutes of Health Research, the Wellcome Trust, and the RIKEN Institute, Japan. We thank Jessica Klassen and Sean Hickman for their assistance.
Correspondence should be addressed to J. Randall Flanagan, Department of Psychology, Queen's University, Kingston, Ontario, Canada K7L 3N6. E-mail: flanagan@post.queensu.ca.
Copyright © 2005 Society for Neuroscience 0270-6474/05/255425-05$15.00/0
References
- Ashe J (1997) Force and the motor cortex. Behav Brain Res 86: 1-15. [DOI] [PubMed] [Google Scholar]
- Baddeley A (1992) Working memory. Science 255: 556-559. [DOI] [PubMed] [Google Scholar]
- Baddeley AD (1986) Working memory. Oxford: Clarendon.
- Brashers-Krug T, Shadmehr R, Bizzi E (1996) Consolidation in human motor memory. Nature 382: 252-255. [DOI] [PubMed] [Google Scholar]
- Caithness G, Osu R, Bays P, Chase H, Klassen J, Kawato M, Wolpert DM, Flanagan JR (2004) Failure to consolidate the consolidation theory of learning for sensorimotor adaptation tasks. J Neurosci 24: 8662-8671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flanagan JR, Tresilian J (1994) Grip-load force coupling: a general control strategy for transporting objects. J Exp Psychol Hum Percept Perform 20: 944-957. [DOI] [PubMed] [Google Scholar]
- Flanagan JR, Wing AM (1997) The role of internal models in motion planning and control: evidence from grip force adjustments during movements of hand-held loads. J Neurosci 17: 1519-1528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gandolfo F, Mussa-Ivaldi FA, Bizzi E (1996) Motor learning by field approximation. Proc Natl Acad Sci USA 93: 3843-3846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gribble PL, Scott SH (2002) Overlap of internal models in motor cortex for mechanical loads during reaching. Nature 417: 938-941. [DOI] [PubMed] [Google Scholar]
- Iriki A, Tanaka M, Iwamura Y (1996) Coding of modified body schema during tool use by macaque postcentral neurones. NeuroReport 7: 2325-2330. [DOI] [PubMed] [Google Scholar]
- Johansson RS, Westling G (1984) Roles of glabrous skin receptors and sensorimotor memory in automatic-control of precision grip when lifting rougher or more slippery objects. Exp Brain Res 56: 550-564. [DOI] [PubMed] [Google Scholar]
- Kawato M (1999) Internal models for motor control and trajectory planning. Current Opin Neurobiol 9: 718-727. [DOI] [PubMed] [Google Scholar]
- Krakauer JW, Ghilardi MF, Ghez C (1999) Independent learning of internal models for kinematic and dynamic control of reaching. Nat Neurosci 2: 1026-1031. [DOI] [PubMed] [Google Scholar]
- Krouchev NI, Kalaska JF (2003) Context-dependent anticipation of different task dynamics: rapid recall of appropriate motor skills using visual cues. J Neurophysiol 89: 1165-1175. [DOI] [PubMed] [Google Scholar]
- Kurtzer I, Herter TM, Scott SH (2005) Random change in cortical load representation suggests distinct control of posture and movement. Nat Neurosci 8: 498-504. [DOI] [PubMed] [Google Scholar]
- Osu R, Hirai S, Yoshioka T, Kawato M (2004) Random presentation enables subjects to adapt to two opposing forces on the hand. Nat Neurosci 7: 111-112. [DOI] [PubMed] [Google Scholar]
- Scott SH (2003) The role of primary motor cortex in goal-directed movements: insights from neurophysiological studies on non-human primates. Curr Opin Neurobiol 13: 671-677. [DOI] [PubMed] [Google Scholar]
- Seidler RD, Bloomberg JJ, Stelmach GE (2001) Context-dependent arm pointing adaptation. Behav Brain Res 119: 155-166. [DOI] [PubMed] [Google Scholar]
- Shadmehr R, Brashers-Krug T (1997) Functional stages in the formation of human long-term motor memory. J Neurosci 17: 409-419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh K, Scott SH (2003) A motor learning strategy reflects neural circuitry for limb control. Nat Neurosci 6: 399-403. [DOI] [PubMed] [Google Scholar]
- Westling G, Johansson RS (1987) Responses in glabrous skin mechanoreceptors during precision grip in humans. Exp Brain Res 66: 128-140. [DOI] [PubMed] [Google Scholar]
- Wigmore V, Tong C, Flanagan JR (2002) Visuomotor rotations of varying size and direction compete for a single internal model in motor working memory. J Exp Psychol Hum Percept Perform 28: 447-457. [DOI] [PubMed] [Google Scholar]
- Wolpert DM, Flanagan JR (2001) Motor prediction. Curr Biol 11: R729-R732. [DOI] [PubMed] [Google Scholar]