

Abstract
Although maintained by multiple arousal systems, wakefulness falters if orexin (hypocretin), orexin receptors, or orexin neurons are deficient; narcolepsy results with hypersomnolence or sudden onset of rapid eye movement sleep [or paradoxical sleep (PS)] and loss of muscle tonus. To learn how orexin neurons maintain wakefulness, we recorded neurons in head-fixed rats across the sleep-waking cycle and then labeled them with Neurobiotin to identify them by immunohistochemistry. We show that identified orexin neurons discharge during active waking, when postural muscle tone is high in association with movement, decrease discharge during quiet waking in absence of movement, and virtually cease firing during sleep, when postural muscle tone is low or absent. During PS, they remain relatively silent in association with postural muscle atonia and most often despite phasic muscular twitches. They increase firing before the end of PS and thereby herald by several seconds the return of waking and muscle tone. The orexin neurons would thus stimulate arousal, while antagonizing sleep and muscle atonia.
Keywords: arousal, hypothalamus, sleep, narcolepsy, hypocretin, REMS
Introduction
Wakefulness is maintained by many parallel and partially redundant neural systems that use multiple neurotransmitters (Jones, 2003). Lesion of any one of these systems, including the noradrenergic locus ceruleus and cholinergic brainstem or basal forebrain neurons, does not result in an inability to stay awake. Yet, loss of the peptide orexin (Orx) (also called hypocretin), its receptor, or the neurons that contain it, results in an incapacity to maintain wakefulness and a syndrome known as narcolepsy, characterized by hypersomnolence, sudden onset of rapid eye movement sleep (REMS) [also called paradoxical sleep (PS)], and/or cataplexy (Chemelli et al., 1999; Lin et al., 1999; Peyron et al., 2000; Thannickal et al., 2000). Orx neurons thus play a key role in warding off sleep, including, importantly, REMS. They may fulfill this role through their documented projections and excitatory actions on other arousal systems together with their direct stimulatory action on cortical, motor, and sympathetic systems (Peyron et al., 1998; Shirasaka et al., 1999; Bourgin et al., 2000; Eggermann et al., 2001; Eriksson et al., 2001; Bayer et al., 2002; Burlet et al., 2002; Krout et al., 2003; Yamuy et al., 2004). They would do so by releasing Orx from their terminals when they discharge. Studies assessing Orx release in the brain by microdialysis indicated that it is maximal during periods of wakefulness (Zeitzer et al., 2003) yet also indicated that it could be high during REMS (Kiyashchenko et al., 2002). Similarly, studies assessing Orx neuronal activity by examining c-Fos expression have indicated that it was high with waking (W) (Estabrooke et al., 2001) yet also appeared to be high during pharmacologically induced REMS (Torterolo et al., 2001). However, the sensitivity and temporal resolution of microdialysis for studying release of Orx and of c-Fos expression for studying activity of Orx neurons are inadequate for revealing neuronal discharge during sleep-wake states. The discharge of Orx neurons across the sleep-waking cycle can be known only by recording their activity during this cycle. Single-unit recording studies have been performed in the rat within the perifornical lateral hypothalamus where the Orx neurons are located, yet have revealed multiple cell types therein including some neurons that discharge during waking and a larger number that discharge during both waking and PS (Alam et al., 2002; Koyama et al., 2003). Thus, before undertaking the present study, the sleep-wake state-related activity of Orx neurons was uncertain, and how they could maintain wakefulness was frankly puzzling.
To determine the discharge profile of Orx neurons across the sleep-waking cycle, we recorded units during sleeping and waking in head-fixed rats (Lee et al., 2004) and then labeled the cells with Neurobiotin (Nb) using the juxtacellular technique (Pinault, 1996) for their subsequent immunohistochemical identification. We reported preliminary results of this work in abstract form (Lee and Jones, 2004).
Materials and Methods
Surgery and recording. All procedures were approved by the McGill University Animal Care Committee and the Canadian Council on Animal Care, which also meet international standards. Adult Long-Evans rats (200-250 g; Charles River, St. Constant, Quebec, Canada) were operated on under deep anesthesia (ketamine, xylazine, and acepromazine, 65:5:1 mg/kg in a mixture of 2 ml/kg initial dose and 1 mg/kg booster as needed, i.p.). EEG (epidural screws over cortex, including olfactory bulb, anterior medial prefrontal cortex, and retrosplenial cortex) and EMG electrodes (in nuccal muscles) were implanted together with a metal U-frame for holding the head by screws to a sliding carriage adapter within the stereotaxic frame, as described previously (Lee et al., 2004). Animals were maintained before and after surgery and during experimentation on a 12 h light/dark schedule with lights on from 7:00 A.M. to 7:00 P.M. After recovery from surgery (2 d), the animals were habituated to head fixation (6-9 d). Lying within a Plexiglas box, they were prevented from twisting their bodies yet were otherwise able to move their body and limbs relatively freely, as well as to sleep in a sphinx position. After adaptation, the rats were once again anesthetized (as above) and operated on by drilling holes in the skull (anteroposterior, -2.8 mm from bregma; lateral, ±1.3 mm) and opening the dura over the lateral hypothalamic area on each side. After 1 d of recovery, daily recording sessions of ∼6 h (∼11:00 A.M. to 5:00 P.M.) were performed over a maximum of 5 d. Single units were recorded using glass micropipettes (∼1 μm tip) that were filled with 0.5 m NaCl and ∼5% Nb (Vector Laboratories, Burlingame, CA), and an intracellular amplifier (Neurodata IR-283A; Cygnus Technology, Delaware Water Gap, PA). The unit signal was amplified (2000×) and filtered (0.3-3 kHz) using a CyberAmp380 (Molecular Devices, Palo Alto, CA), then acquired at 16 kHz for on-line viewing with Axoscope (version 8.1; Molecular Devices) to verify that the recorded activity was derived from a single unit. The unit signal and EEG/EMG were simultaneously acquired and digitized at 8 kHz and 250 Hz, respectively, using EEG and sleep-wake scoring software (Harmonie, version 5.2; Stellate, Montreal, Quebec, Canada). Video recording of behavior was also acquired simultaneously by the same software. On the last day of recording from one side, the last cell recorded during active wake (aW), slow-wave sleep (SWS), and PS was labeled with Nb using the juxtacellular technique, as described previously (Pinault, 1996; Manns et al., 2000; Lee et al., 2004).
Data analysis. For analysis, units were selected that had been recorded for >5 min and during at least one episode of aW, SWS, and PS. Together with the synchronized video images of behavior, records of EEG and EMG were scored by 10 s epochs for sleep-wake state as described previously (Maloney et al., 1997; Lee et al., 2004). Wake, SWS, and PS states were scored when characteristic EEG/EMG activity occupied >75% of the epoch; transitional stages into SWS and PS were scored when characteristic activity or mixed activity occurred during the epoch. Active wake was scored when movement occurred during the wake epoch, as evident by phasic EMG activity along with video images, and theta EEG activity. The unit activity was subsequently analyzed per 10 s epoch in each sleep-wake state for average discharge rate (spikes per second) and instantaneous firing frequency using the first modal peak of the interspike interval histogram. Gamma (30-58 Hz) power, delta (1-4.5 Hz) power, and theta (4.5-8 Hz) activity were measured per epoch along with EMG amplitude (30-100 Hz) for correlation with unit spike rate. Classification of units according to the state in which their maximal discharge rate occurred was performed by ANOVA followed by post hoc paired comparisons (Lee et al., 2004). All spikes were averaged across the recording for measurement of the spike duration (from first zero crossing on upward deflection to second zero crossing on second upward deflection on return to baseline, set at 10% of peak amplitude from zero). The change in firing during the transition from PS to W was determined by considering the initial increase in discharge as the time when the firing rate during three consecutive 1 s bins was at least 2 SDs greater than the mean rate during 20 s before the state transition.
Histochemistry. After the recording and labeling of units, the rat was anesthetized with an overdose of sodium pentobarbital (Somnotol; ∼100 mg/kg, i.p.) and perfused with 4% paraformaldehyde solution. After immersion in 30% sucrose for 2-3 d, brains were frozen for storage and subsequent cutting in 25-μm-thick sections. For revelation of Neurobiotin, sections were incubated for 2.5 h in cyanine 2 (Cy2)-conjugated streptavidin (1:1000; Jackson ImmunoResearch, West Grove, PA). After location of an Nb-labeled cell, the relevant section was incubated over-night at room temperature in goat anti-Orx A antiserum (1:500; Santa Cruz Biotechnology, Santa Cruz, CA) and subsequently for 2 h in Cy3-conjugated donkey anti-goat antiserum (1:1000; Jackson ImmunoResearch). Nb-labeled cells were located by epifluorescence using a Leica (Nussloch, Germany) DMLB microscope. Images of the labeled cells were acquired and their location mapped onto a computer resident atlas with the aid of Neurolucida (version 5; MicroBrightField, Williston, VT).
Results
We recorded neurons within the perifornical lateral hypothalamus in head-fixed rats that were habituated to sleep in a stereotaxic frame (Lee et al., 2004). After recording a neuron across aW, quiet waking (qW), transition to slow-wave sleep (tSWS), SWS, transition to PS (tPS), and PS, we labeled that neuron with Nb using the juxtacellular technique (Lee et al., 2004) for subsequent immunohistochemical staining of Orx. In initial sampling, we selected cells with different profiles of activity to sample all of the different cell types previously recorded and categorized in the lateral hypothalamus (Alam et al., 2002; Koyama et al., 2003). Twenty-five neurons were successfully recorded across all sleep-wake states, labeled with Nb, and found to be situated within the lateral hypothalamus in the region of the Orx neurons (Fig. 1). Six were immunopositive for Orx (Orx+) (Fig. 1, Table 1). All Nb+/Orx+ cells discharged at their maximal rate during aW and were thus classified as W-max-active (Table 1). Cells that discharged maximally during SWS (S-max-active), PS (P-max-active), or aW and PS (WP-max-active) were all Orx negative (Orx-) (Table 1) (data not shown). And cells that discharged at equivalent rates across states (wsp-eq) were Orx negative (data not shown).
Figure 1.

Orx neurons recorded and labeled in the hypothalamus. A, Image showing red, Cy3-immunostained Orx neurons in the perifornical, lateral hypothalamic area. B, Magnified image of the box in A showing an Nb-labeled cell stained with green, Cy2, and red, Cy3, thus appearing yellow as a Nb+/Orx+ cell (#c38u24). C, Plots of six Nb+/Orx+ recorded neurons on a coronal section (approximately -2.8 mm from bregma) through the lateral hypothalamic area. DMH, Dorsomedial hypothalamic nucleus; f, fornix; ic, internal capsule; LH, lateral hypothalamus; VMH, ventromedial hypothalamic nucleus; ZI, zona incerta. Scale bars: A, C, 1 mm; B, 20 μm.
Table 1.
Sleep-wake discharge of Orx-positive (Orx+) and Orx-negative (Orx−) Nb-labeled cells
|
Category of Nb + cells |
Orx+ |
Orx− |
Total |
|---|---|---|---|
| W-max | 6 | 5 | 11 |
| S-max | 0 | 1 | 1 |
| P- or WP-max | 0 | 9 | 9 |
| wsp-eq | 0 | 4 | 4 |
| Total |
6 |
19 |
25 |
Nb-labeled cells were classified individually using ANOVA followed by paired comparisons of average discharge rates (hertz) per major state (aW, SWS, and PS). Those showing state-dependent variation in the discharge rate (p < 0.05) were grouped according to the occurrence of maximal rates during aW (W-max-active), SWS (S-max-active), or PS or aW-PS (P- or WP-max-active). Others were classified as state equivalent (wsp-eq). The frequencies of cell types differ significantly between the Orx+ and Orx− cells (ϰ2 = 10.05; df = 3; p = 0.02).
The average discharge rate of the Orx+ neurons during aW was 3.17 ± 0.79 Hz (mean ± SEM; n = 6), and the instantaneous firing frequency (based on the interspike interval mode) was 12.20 ± 2.93 Hz, reflecting a relatively slow discharge in single spikes with some phasic modulation in frequency. Their average spike duration was 2.07 ± 0.08 ms. The W-max-active, Orx-negative cells (n = 5) (Table 1) showed greater variability in their discharge properties and, because of overlap, did not differ significantly as a group from the Orx+ cells in their average discharge rate (12.15 ± 7.20 Hz) or instantaneous firing frequency (40.76 ± 20.94 Hz) during active waking, nor in their average spike duration (1.72 ± 0.22 ms). Along two parameters, however, W-max-active, Orx-negative cells could be distinguished from Orx+ cells (according to values falling outside the 0.99 confidence intervals of the Orx+ cell distributions) by higher average spike rate (>6.5 Hz) and shorter or longer spike duration (<1.76 or >2.38 ms). However, there were Orx-negative cells that discharged maximally in each of the other states or equivalently across states (see above) (Table 1) that could not be distinguished from Orx+ cells by these criteria.
The average discharge rate of the Orx+ neurons decreased markedly from aW (3.17 ± 0.79 Hz; mean ± SEM; n = 6) to qW (0.64 ± 0.22; representing a significant change of rate in six of six cells; p < 0.05) (Fig. 2A). It decreased further from aW and qW to sleep (representing a significant change in rate from aW in six of six cells and from qW in three of six cells; p < 0.05). The low rate during sleep reflected a virtual silence that occurred during tSWS (0.14 ± 0.07 Hz), SWS (0.09 ± 0.05 Hz), and tPS (0.03 ± 0.02 Hz), and that continued during PS (0.24 ± 0.11 Hz; with no significant change in rate across sleep stages in five of six cells; p > 0.05). Across all sleep-wake stages (including tPS and PS), the discharge rate was negatively correlated with delta EEG power (average r = -0.47), weakly correlated with gamma EEG power (average r = 0.14), and positively correlated with EMG amplitude (average r = 0.57). Across W and SWS stages (and not including tPS and PS when Orx neurons are relatively silent despite cortical activation), the discharge rate was more strongly negatively correlated with delta power (average r = -0.64) and positively correlated with gamma power (average r = 0.49), as well as EMG amplitude (average r = 0.36). The positive correlation of discharge rate with EMG amplitude reflected the relatively high discharge rate that occurred with active waking, which was defined by phasic activity on the neck EMG recording and associated theta activity on the EEG recording along with movement of the body, limbs, whiskers, or muzzle evident on the video recording (Fig. 2B,C1) (supplemental video 1, available at www.jneurosci.org as supplemental material). It also reflected the very low discharge rate or relative silence that occurred during SWS with low muscle tone and during PS with absence of muscle tone (Fig. 2C1,C2) (supplemental video 2, available at www.jneurosci.org as supplemental material). When the neurons did discharge with one or two spikes during sleep, they usually did so at a time when a small movement or twitch occurred, although not always and not usually simultaneously. Nonetheless, the neurons remained silent during long periods of PS while prominent twitches of the whiskers and muzzle or even paws occurred (supplemental videos 2 and 3, available at www.jneurosci.org as supplemental material). In contrast, when the Orx neurons began to discharge with several spikes during PS, a brief arousal could occur with a change in EEG activity and appearance of neck muscle tone along with body movement, indicative of waking (Fig. 2C3) (supplemental video 3, available at www.jneurosci.org as supplemental material). In the transition from PS to aW, Orx+ cells consistently began to fire during PS (2-7 s) before the return of EMG activity and (up to 6 s) before the change in EEG activity seen as an attenuation of theta (in six Orx cells) (Fig. 2C3,C4) (supplemental videos 3 and 4, available at www.jneurosci.org as supplemental material). Across sleep-wake states, the spike discharge was thus positively correlated with movement, as indicative of behavioral arousal, yet could also precede as well as occur during movement and increased postural muscle tone. Uniquely across wake and SWS, the discharge was also positively correlated with gamma EEG activity, as indicative of cortical activation that can occur with attentive in addition to active waking and thus independent of movement (Maloney et al., 1997).
Figure 2.
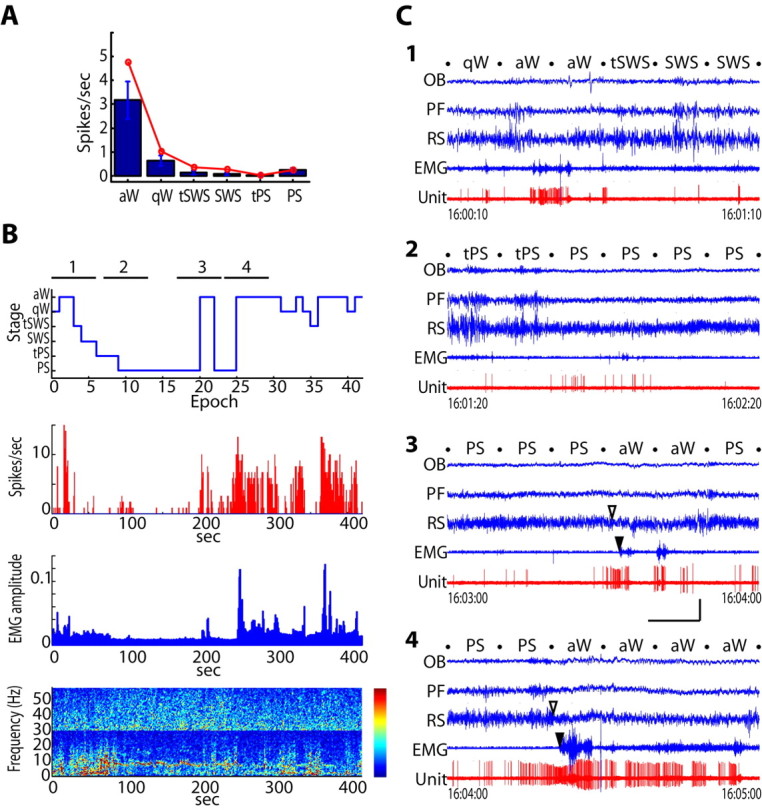
Discharge of Nb+/Orx+ neurons across sleep-wake states. A, Bar graph showing mean spike rate (in blue with SEM) that varied significantly as a function of sleep-wake stage across cells (n = 6; F = 16.05; df = 5, 25; p = 0.007). The rate during aW was significantly higher than that during qW and all sleep stages, including PS (p < 0.05). The line graph (in red) shows the discharge rate of the Nb+/Orx+ neuron (#c32u10) shown in B and C. B, Hypnogram, spike rate histogram, and EMG amplitude and EEG frequency spectra (split for different scaling from blue to red over the lower frequency range, 0-30 Hz from 1 to 100 uV, and gamma range, 30-60 Hz from 0 to 25 uV) over the recording session (of 400 s corresponding to 40 10 s epochs). C, Four 1 min segments (as indicated by time below) of unit, EEG, and EMG activity during state transitions: from qW through aW, tSWS to SWS (supplemental video 1, available at www.jneurosci.org as supplemental material) (1), from tPS to PS (supplemental video 2, available at www.jneurosci.org as supplemental material) (2), from PS through aW back to PS (supplemental video 3, available at www.jneurosci.org as supplemental material) (3), and from PS to aW (supplemental video 4, available at www.jneurosci.org as supplemental material) (4). Stages were scored per 10 s epoch as indicated by the stage names between dots marking the epochs. Note that the unit fires during aW (in 1) becomes quiet during SWS (in 1) and PS (in 2 and 3), except for occasional spikes sometimes associated with twitches (in 2) or a brief arousal (in 3). The increase in firing before arousal from PS (in 3 and 4) anticipates the transition from PS to aW judged by EEG (open arrowhead) and also EMG (filled arrowhead). Calibration: horizontal, 10 s; vertical, 1 mV for EEG, 0.5 mV for EMG, and 2 mV for unit. OB, Olfactory bulb; PF, prefrontal cortex; RS, retrosplenial cortex.
Discussion
We show for the first time that Orx neurons discharge during and preceding active waking and virtually cease firing during sleep, including SWS and, importantly, PS (or REMS). Given that Orx neurons have the capacity to sustain neural activity in the absence of synaptic input (Eggermann et al., 2003), we presume that their silence during sleep is caused by inhibitory input from GABAergic sleep-active neurons located in the basal forebrain and preoptic area or elsewhere (Henny and Jones, 2003; Modirrousta et al., 2004). Compared with other arousal systems, the Orx neurons are similar to the noradrenergic locus ceruleus neurons, which are W-On and PS-Off cells that promote cortical and behavioral arousal (Hobson et al., 1975; Aston-Jones and Bloom, 1981). They are entirely different from cholinergic brainstem and basal forebrain neurons, which are W-On and PS-On neurons that stimulate cortical activation during both W and PS, and can also promote the muscle atonia or cataplexy of PS or REMS (Mitler and Dement, 1974; El Mansari et al., 1989; Nishino et al., 1995; Lee et al., 2005). The Orx cell discharge is positively correlated with EMG activity as well as fast, gamma EEG activity during waking and negatively correlated with slow, delta EEG activity. The Orx neurons would thus stimulate both increased muscle tone and cortical activation when they fire before, as well as during, active waking. In fact, local injections of Orx in the brain have been shown to interrupt sleep and stimulate prolonged active waking (Bourgin et al., 2000; Espana et al., 2001). Given their projections and influence (Peyron et al., 1998; Krout et al., 2003), Orx neurons can directly stimulate cortical activation and muscle tone by directly exciting cortical neurons (Bayer et al., 2004) and motor neurons (Yamuy et al., 2004). Also known to play an important role in energy regulation, they moreover have the capacity to stimulate metabolism in parallel with activity through influences on the sympathetic nervous system, the hypothalamo-pituitary-adrenal axis and hypothalamo-pituitary-thyroid axis (Ida et al., 2000; Hara et al., 2001; Espana et al., 2003; Monda et al., 2003; Yamanaka et al., 2003). They would concurrently engage all other, including noradrenergic, arousal systems in the brain (Peyron et al., 1998; Bourgin et al., 2000; Eggermann et al., 2001; Eriksson et al., 2001; Bayer et al., 2002; Burlet et al., 2002; Krout et al., 2003). Indeed, the increase in discharge by the Orx neurons that precedes the cortical activity change and return of muscle tone in the transition from PS to wake, appears to anticipate that of the noradrenergic locus ceruleus neurons (Aston-Jones and Bloom, 1981). With a loss of Orx neurons, early and coordinated engagement of these multiple systems would be deficient, resulting in their inadequate recruitment and thus deficient arousal or hypersomnolence. Their absence would also likely result in an imbalance among systems under certain conditions, resulting in a predominance by the cholinergic W-On/PS-On neurons that would precipitate the loss of muscle tonus together with cortical activation and thus the sudden onset of PS or REMS, as occurs in narcolepsy (Nishino et al., 1995).
Footnotes
This work was supported by Canadian Institutes of Health Research Grant 13458 and National Institutes of Health Grant RO1 MH-60119-01A to B.E.J. We thank Lynda Mainville, Pablo Henny, Mandana Modirrousta, and Frederic Brischoux for assistance with the histochemistry and neuroanatomical analysis.
Correspondence should be addressed to Barbara E. Jones, McGill University, Montreal Neurological Institute, 3801 University Street, Montreal, Quebec, Canada H3A 2B4. E-mail: barbara.jones@mcgill.ca.
Copyright © 2005 Society for Neuroscience 0270-6474/05/256716-05$15.00/0
M.G.L. and O.K.H. contributed equally to this work.
References
- Alam MN, Gong H, Alam T, Jaganath R, McGinty D, Szymusiak R (2002) Sleep-waking discharge patterns of neurons recorded in the rat perifornical lateral hypothalamic area. J Physiol (Lond) 538: 619-631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aston-Jones G, Bloom FE (1981) Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J Neurosci 1: 876-886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayer L, Eggermann E, Saint-Mleux B, Machard D, Jones BE, Muhlethaler M, Serafin M (2002) Selective action of orexin (hypocretin) on nonspecific thalamocortical projection neurons. J Neurosci 22: 7835-7839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayer L, Serafin M, Eggermann E, Saint-Mleux B, Machard D, Jones BE, Muhlethaler M (2004) Exclusive postsynaptic action of hypocretinorex in on sublayer 6b cortical neurons. J Neurosci 24: 6760-6764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourgin P, Huitron-Resendiz S, Spier AD, Fabre V, Morte B, Criado JR, Sutcliffe JG, Henriksen SJ, de Lecea L (2000) Hypocretin-1 modulates rapid eye movement sleep through activation of locus ceruleus neurons. J Neurosci 20: 7760-7765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burlet S, Tyler CJ, Leonard CS (2002) Direct and indirect excitation of laterodorsal tegmental neurons by hypocretin/orexin peptides: implications for wakefulness and narcolepsy. J Neurosci 22: 2862-2872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chemelli RM, Willie JT, Sinton CM, Elmquist JK, Scammell T, Lee C, Richardson JA, Williams SC, Xiong Y, Kisanuki Y, Fitch TE, Nakazato M, Hammer RE, Saper CB, Yanagisawa M (1999) Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell 98: 437-451. [DOI] [PubMed] [Google Scholar]
- Eggermann E, Serafin M, Bayer L, Machard D, Saint-Mleux B, Jones BE, Muhlethaler M (2001) Orexins/hypocretins excite basal forebrain cholinergic neurones. Neuroscience 108: 177-181. [DOI] [PubMed] [Google Scholar]
- Eggermann E, Bayer L, Serafin M, Saint-Mleux B, Bernheim L, Machard D, Jones BE, Muhlethaler M (2003) The wake-promoting hypocretinorexin neurons are in an intrinsic state of membrane depolarization. J Neurosci 23: 1557-1562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El Mansari M, Sakai M, Jouvet M (1989) Unitary characteristics of presumptive cholinergic tegmental neurons during the sleep-waking cycle in freely moving cats. Exp Brain Res 76: 519-529. [DOI] [PubMed] [Google Scholar]
- Eriksson KS, Sergeeva O, Brown RE, Haas HL (2001) Orexin/hypocretin excites the histaminergic neurons of the tuberomammillary nucleus. J Neurosci 21: 9273-9279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espana RA, Baldo BA, Kelley AE, Berridge CW (2001) Wake-promoting and sleep-suppressing actions of hypocretin (orexin): basal forebrain sites of action. Neuroscience 106: 699-715. [DOI] [PubMed] [Google Scholar]
- Espana RA, Valentino RJ, Berridge CW (2003) Fos immunoreactivity in hypocretin-synthesizing and hypocretin-1 receptor-expressing neurons: effects of diurnal and nocturnal spontaneous waking, stress and hypocretin-1 administration. Neuroscience 121: 201-217. [DOI] [PubMed] [Google Scholar]
- Estabrooke IV, McCarthy MT, Ko E, Chou TC, Chemelli RM, Yanagisawa M, Saper CB, Scammell TE (2001) Fos expression in orexin neurons varies with behavioral state. J Neurosci 21: 1656-1662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hara J, Beuckmann CT, Nambu T, Willie JT, Chemelli RM, Sinton CM, Sugiyama F, Yagami K, Goto K, Yanagisawa M, Sakurai T (2001) Genetic ablation of orexin neurons in mice results in narcolepsy, hypophagia, and obesity. Neuron 30: 345-354. [DOI] [PubMed] [Google Scholar]
- Henny P, Jones BE (2003) Vesicular transporter proteins for glutamate (VgluT), GABA (VGAT) and acetylcholine (VAChT) in terminal varicosities of basal forebrain neurons projecting to the posterior lateral hypothalamus. Sleep 26: A9. [Google Scholar]
- Hobson JA, McCarley RW, Wyzinski PW (1975) Sleep cycle oscillation: reciprocal discharge by two brainstem neuronal groups. Science 189: 55-58. [DOI] [PubMed] [Google Scholar]
- Ida T, Nakahara K, Murakami T, Hanada R, Nakazato M, Murakami N (2000) Possible involvement of orexin in the stress reaction in rats. Biochem Biophys Res Commun 270: 318-323. [DOI] [PubMed] [Google Scholar]
- Jones BE (2003) Arousal systems. Front Biosci 8: S438-S451. [DOI] [PubMed] [Google Scholar]
- Kiyashchenko LI, Mileykovskiy BY, Maidment N, Lam HA, Wu MF, John J, Peever J, Siegel JM (2002) Release of hypocretin (orexin) during waking and sleep states. J Neurosci 22: 5282-5286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koyama Y, Takahashi K, Kodama T, Kayama Y (2003) State-dependent activity of neurons in the perifornical hypothalamic area during sleep and waking. Neuroscience 119: 1209-1219. [DOI] [PubMed] [Google Scholar]
- Krout KE, Mettenleiter TC, Loewy AD (2003) Single CNS neurons link both central motor and cardiosympathetic systems: a double-virus tracing study. Neuroscience 118: 853-866. [DOI] [PubMed] [Google Scholar]
- Lee MG, Jones BE (2004) Discharge of identified orexin neurons across the sleep-waking cycle. Soc Neurosci Abstr 30: 841.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee MG, Manns ID, Alonso A, Jones BE (2004) Sleep-wake related discharge properties of basal forebrain neurons recorded with micropipettes in head-fixed rats. J Neurophysiol 92: 1182-1198. [DOI] [PubMed] [Google Scholar]
- Lee MG, Hassani OK, Alonso A, Jones BE (2005) Cholinergic basal forebrain neurons burst with theta during waking and paradoxical sleep. J Neurosci 25: 4365-4369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin L, Faraco J, Li R, Kadotani H, Rogers W, Lin X, Qiu X, de Jong PJ, Nishino S, Mignot E (1999) The sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor 2 gene. Cell 98: 365-376. [DOI] [PubMed] [Google Scholar]
- Maloney KJ, Cape EG, Gotman J, Jones BE (1997) High frequency gamma electroencephalogram activity in association with sleep-wake states and spontaneous behaviors in the rat. Neuroscience 76: 541-555. [DOI] [PubMed] [Google Scholar]
- Manns ID, Alonso A, Jones BE (2000) Discharge properties of juxtacellularly labeled and immunohistochemically identified cholinergic basal forebrain neurons recorded in association with the electroencephalogram in anesthetized rats. J Neurosci 20: 1505-1518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitler MM, Dement WC (1974) Cataplectic-like behavior in cats after microinjections of carbachol in pontine reticular formation. Brain Res 68: 335-343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Modirrousta M, Mainville L, Jones BE (2004) GABAergic neurons with alpha2-adrenergic receptors in basal forebrain and preoptic area express c-Fos during sleep. Neuroscience 129: 803-810. [DOI] [PubMed] [Google Scholar]
- Monda M, Viggiano A, De Luca V (2003) Paradoxical [correction of parodoxical] effect of orexin A: hypophagia induced by hyperthermia. Brain Res 961: 220-228. [DOI] [PubMed] [Google Scholar]
- Nishino S, Tafti M, Reid MS, Shelton J, Siegel JM, Dement WC, Mignot E (1995) Muscle atonia is triggered by cholinergic stimulation of the basal forebrain: implication for the pathophysiology of canine narcolepsy. J Neurosci 15: 4806-4814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peyron C, Tighe DK, van den Pol AN, de Lecea L, Heller HC, Sutcliffe JG, Kilduff TS (1998) Neurons containing hypocretin (orexin) project to multiple neuronal systems. J Neurosci 18: 9996-10015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peyron C, Faraco J, Rogers W, Ripley B, Overeem S, Charnay Y, Nevsimalova S, Aldrich M, Reynolds D, Albin R, Li R, Hungs M, Pedrazzoli M, Padi-garu M, Kucherlapati M, Fan J, Maki R, Lammers GJ, Bouras C, Kucherlapati R, et al. (2000) A mutation in a case of early onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains. Nat Med 6: 991-997. [DOI] [PubMed] [Google Scholar]
- Pinault D (1996) A novel single-cell staining procedure performed in vivo under electrophysiological control: morpho-functional features of juxtacellularly labeled thalamic cells and other central neurons with biocytin or Neurobiotin. J Neurosci Methods 65: 113-136. [DOI] [PubMed] [Google Scholar]
- Shirasaka T, Nakazato M, Matsukura S, Takasaki M, Kannan H (1999) Sympathetic and cardiovascular actions of orexins in conscious rats. Am J Physiol 277: R1780-R1785. [DOI] [PubMed] [Google Scholar]
- Thannickal TC, Moore RY, Nienhuis R, Ramanathan L, Gulyani S, Aldrich M, Cornford M, Siegel JM (2000) Reduced number of hypocretin neurons in human narcolepsy. Neuron 27: 469-474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torterolo P, Yamuy J, Sampogna S, Morales FR, Chase MH (2001) Hypothalamic neurons that contain hypocretin (orexin) express c-fos during active wakefulness and carbachol-induced active sleep. Sleep Res Online 4: 25-32. [Google Scholar]
- Yamanaka A, Beuckmann CT, Willie JT, Hara J, Tsujino N, Mieda M, Tominaga M, Yagami K, Sugiyama F, Goto K, Yanagisawa M, Sakurai T (2003) Hypothalamic orexin neurons regulate arousal according to energy balance in mice. Neuron 38: 701-713. [DOI] [PubMed] [Google Scholar]
- Yamuy J, Fung SJ, Xi M, Chase MH (2004) Hypocretinergic control of spinal cord motoneurons. J Neurosci 24: 5336-5345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeitzer JM, Buckmaster CL, Parker KJ, Hauck CM, Lyons DM, Mignot E (2003) Circadian and homeostatic regulation of hypocretin in a primate model: implications for the consolidation of wakefulness. J Neurosci 23: 3555-3560. [DOI] [PMC free article] [PubMed] [Google Scholar]