

Abstract
The Na+/Ca2+ exchangers NCX1, NCX2, and NCX3 are vital for the control of cellular Ca2+ homeostasis. Here, we show that a doublet of downstream regulatory element sites in the promoter of the NCX3 gene mediates transcriptional repression of NCX3 by the Ca2+-modulated transcriptional repressor downstream regulatory element antagonist modulator (DREAM). Overexpression of a DREAM EF-hand mutant insensitive to Ca2+ (EFmDREAM) in hippocampus and cerebellum of transgenic mice significantly reduced NCX3 mRNA and protein levels without modifying NCX1 and NCX2 expression. Cerebellar granules from EFmDREAM transgenic mice showed increased levels of cytosolic Ca2+ and were more vulnerable to increased Ca2+ influx after partial opening of voltage-gated plasma membrane Ca2+ channels induced by increasing K+ in the culture medium but survived better in the conditions of reduced Ca2+ influx prevailing in low extracellular K+. Overexpression of NCX3 in EFmDREAM transgenic granules using a lentiviral vector restored the normal survival response to high K+ observed in wild-type granules. Thus, the downregulation of the regulator of Ca2+ homeostasis NCX3 by Ca2+-regulated DREAM is a striking example of the autoregulatory property of the Ca2+ signal in neurons.
Keywords: calcium, gene expression, cerebellar granules, EF hands, lentivirus, Na+/Ca2+ exchangers
Introduction
The Na+/Ca2+ exchangers NCX1, NCX2, and NCX3 are plasma membrane proteins crucial for the maintenance of intracellular Ca2+ homeostasis because they rapidly reduce the Ca2+ rise after neuronal excitation or heart stimulation (for review, see Philipson, 1999; Gabellini et al., 2000). The Na+/Ca2+ exchangers are differentially expressed in excitable tissues (Kofuji et al., 1992; Nicoll et al., 1996), suggesting differences in their transcriptional regulation, possibly to satisfy specific demands of Ca2+ homeostasis in various cell types.
Regulation of expression of the NCX genes has been studied in cultured cerebellar granules (Li et al., 2000), in which the levels of NCX2 protein are rapidly downregulated through a Ca2+/calcineurin-dependent mechanism induced by high extracellular K+. In contrast, membrane depolarization upregulates the expression of the NCX3 gene and fails to modify the NCX1 gene (Li et al., 2000). The human NCX3 proximal promoter contains specific enhancers for both muscle and neuronal expression as well as a cAMP response element (CRE) (Gabellini et al., 2003). Nevertheless, presence of a CRE by itself does not guarantee transcriptional activation by CRE binding protein (CREB)/CREB binding protein (CBP) in vivo (Zhang et al., 2005), and the presence of additional sites that could mediate the induction after neuronal depolarization has not been fully investigated. NCX3 is the most abundant Na+/Ca2+ exchanger in cerebellar granules, and its rapid degradation by calpains occurs in cerebellar granules exposed to excitotoxins (Bano et al., 2005). This suggests a crucial role for the NCX3 protein in the maintenance of adequate Ca2+ homeostasis in these neurons, emphasizing the importance of clarifying the mechanisms governing NCX3 expression in neurons.
A direct Ca2+-linked mechanism for the regulation of gene expression involves the Ca2+-binding protein downstream regulatory element (DRE) antagonist modulator (DREAM) (Carrion et al., 1999), which has been found to bind to specific DRE sites to repress transcription of several genes (Carrion et al., 1999; Sanz et al., 2001; Link et al., 2004; Rivas et al., 2004; Savignac et al., 2005; Scsucova et al., 2005). Binding of Ca2+ to DREAM through EF-hand motifs reduces its affinity for DNA (Carrion et al., 1999; Craig et al., 2002), leading to its detachment from the DRE sites and to derepression of target genes. DREAM-dependent transcriptional derepression is also observed after protein kinase A (PKA) activation through a mechanism that involves the interaction of DREAM with phosphorylated α-CRE modulator (α-CREM) (Ledo et al., 2000). Moreover, DREAM represses CRE-dependent transcription by a Ca2+-dependent interaction with CREB, which prevents the recruitment of CBP (Ledo et al., 2002).
Here, we show that DREAM mediates NCX3 gene repression in the cerebellum of transgenic mice overexpressing the Ca2+-insensitive EF-hand mutant EFmDREAM. As a result, transgenic cerebellar granules have increased levels of cytosolic Ca2+ and are less viable when cultured under mild membrane-depolarizing conditions. Importantly, their viability is normalized when their Ca2+ extruding ability is restored by lentiviral-mediated overexpression of NCX3. Thus, the Ca2+-modulated transcriptional repressor DREAM controls the expression of the NCX3 protein, which is fundamental for the maintenance of the Ca2+ homeostasis and viability of the neurons.
Materials and Methods
Reporter vectors. A 650 bp genomic fragment containing the minimal NCX3 promoter region and exon 1 was amplified using primers -263: 5′-GAGCTCCCAACTCGGCGG-3′, and +364 in exon 1: 5′-AAGCTTAGCGGTGACTGGAATCTACG-3′. The PCR fragment, including the doublet of DRE sites at position -7, was inserted in the pGL2Basic (Promega, Madison, WI) vector yielding reporter plasmid pNCX3. Site-directed mutagenesis at the doublet of DRE sites in the NCX3 reporter was performed using the QuickChange method (Stratagene, La Jolla, CA). The mutated NCX3 reporter includes a GT-to-CA substitution at each DRE core GTCA.
Cell culture, transfection, and reporter assays. Human carcinoma human embryonic kidney 293 (HEK293) and human neuroblastoma SH-SY5Y cells were cultured in DMEM supplemented with 10% heat-inactivated fetal calf serum. For the luciferase reporter assays, 1 × 106 cells were plated on 60 mm dishes. Six to 10 h after seeding, 5 μg of reporter plasmid and 3 μg of expression vector were coprecipitated with calcium phosphate and added to HEK293 cells. Transfection efficiency was corrected for by cotransfection with the pTKRenilla vector (Promega). Luciferase reporter assays were performed as described previously (Rivas et al., 2004). Stably transfected SH-SY5Y cells were prepared by neomycin selection in medium containing 1 mm G418 for 1 month and were fed with fresh medium every 2 d. Primary cerebellar granule cultures were prepared from 6- to 7-d-old mice as described previously (Gallo et al., 1987). Cells were plated onto 60 mm culture dishes precoated with 50 μg/ml poly-l-lysine at a density of 3 × 106 cells/plate and maintained in Neurobasal medium supplemented with B27, 2 mm glutamax, and 100 μg/ml penicillin/streptomycin (all from Invitrogen, Carlsbad, CA) in the presence of 5 (low K+) or 25 mm KCl (high K+). This culture medium is optimized for neuronal survival and minimal proliferation of glial cells, which accounted for <0.5% after 5 d in culture (Brewer, 1997). Treatments with the voltage-dependent calcium channel blocker nimodipine (3 μm) or calcineurin inhibitors FK506 (1 μm) and cyclosporin A (CsA) (10 μm) were applied immediately after plating and were left in the medium until the cells were harvested.
Transduction of cerebellar granule neurons with lentiviral vectors. Viral particles were obtained as described previously (Follenzi and Naldini, 2002). Briefly, HEK293 cells were transfected with three packaging plasmids and the vector encoding NCX3 (pRRLsin.PPT.hCMV.NCX3.Wpre) or GFP (PRRLsin.PPT.hCMV.GFPpre). After 36-60 h, the viral particles were collected and concentrated by ultracentrifugation. The virus concentration was estimated by measuring the amount of p24 protein (PerkinElmer, Wellesley, MA). Cerebellar granules were transduced 24 h after plating using 5 μg of p24/106 cells. The medium was changed 8 h after the addition of the virus, and the cells were allowed to express the protein of interest for at least 48 h before experiments. Typical infection efficiency was ∼90%, as assessed using viral delivery of GFP. The viral vectors encoding NCX3 or GFP were characterized previously (Bano et al., 2005).
Western blots. Cerebellar membrane proteins were prepared as described previously (Link et al., 2004). Protein concentration was determined using the Bradford reagent (Sigma, St. Louis, MO). Proteins were separated by 10% SDS-PAGE (20 μg/lane), electroblotted onto nitrocellulose membranes, and transiently stained with Ponceau Red. The immunostaining was performed with a specific anti-DREAM antibody (Link et al., 2004) and with peptide-specific antibodies against peptide 276-722 from rat NCX3 (Nicoll et al., 1996), against peptide 486-661 from rat NCX2, and against peptide 566-691 from rat NCX1 (Li et al., 2000). Anti-human CREB antibody (Santa Cruz Biotechnology, Santa Cruz, CA) was used at a 1:100 dilution. The reactions were revealed by ECL reagent (Amersham Biosciences, Arlington Heights, IL). Densitometric analysis was performed with the Kodak (Rochester, NY) ID Image Analysis software by evaluating the net intensity of the immunoreactive bands.
Electrophoretic mobility assay. A double-stranded oligonucleotide corresponding to the mouse NCX3 DRE site (DRENCX3) 5′-GCGCGGCTTGTCAGTCAGTGCGGCG-3′, was labeled with [γ-32P] ATP and T4 kinase and was used as probe. In addition, the mouse ICER DRE sites (DREICER) 5′-GGTGACGTCACTGTGATGTCAGTG-3′, and the Sp1 site 5′-ATTCGATCGGGGCGGGGC GAGC-3′ were used as controls. Protein extracts of SH-SY5Y cells (5-10 μg) were incubated with the radioactive oligonucleotide probe for 20 min at room temperature in reaction buffer: 10 mm HEPES, pH 7.9, 10% glycerol, 0.1 mm EDTA, 8 mm MgCl2, 1 mm dithiothreitol (DTT), 0.15 μg of poly(dI-dC) per milliliter. Protein-DNA complexes were resolved in 5% nondenaturing polyacrylamide gels and visualized by autoradiography.
Real-time quantitative PCR. Total RNA from selected mouse brain areas was prepared using Trizol (Invitrogen). Quantitative real-time PCR was performed using specific primers and TaqMan Minor Grove Binding (MGB) probes for EFmDREAM and NCX3. For EFmDREAM, the primers used were as follows: forward, 5′-CACCTATGCACACTTCCTCTTCA-3′ and reverse, 5′-ACCACAAAGTCCTCAAAGTGGAT-3′ and the probe VIC-5′-CGCCTTTGCTGCGGC-3′-MGB. For NCX3 and NCX2 quantification, specific primers and TaqMan MGB probes were supplied by Applied Biosystems (Foster City, CA). The results were normalized as indicated by parallel amplification of HPRT or β-actin. For HPRT, the primers used were as follows: forward, 5′-TTGGATACAGGCCAGACTTTGTT-3′ and reverse, 5′-CTGAAGTACTCATTATAGTCAAGGGCATA-3′ and the probe FAM-5′-TTGAAATTCCAGACA-3′-MGB. For β-actin, specific primers and TaqMan MGB probe were supplied by Applied Biosystems.
Transgenic mice. The cDNA encoding, human DREAM with two amino acid substitutions at EF hands 2, 3, and 4 (EFmDREAM) was cloned downstream of the human calcium/calmodulin-dependent protein kinase IIα (CaMKIIα) promoter (Mayford et al., 1996). The transgenic cassette was microinjected into the pronuclei of one-cell embryos (C57BL/6 × CBA F1) using standard techniques. Transgenic progeny were identified by Southern blot and qualitative PCR of tail DNA using specific primers: upstream, 5′-TTGCAGTGCACGGCAGATACACTTGCTGA-3′ and downstream, 5′-CCACTGGTGTGGGCCATAATTCAATTCGC-3′. An amplified fragment of 326 bp indicated the presence of the transgene. Founder males were backcrossed to C57BL/6 females to generate lines that were maintained as heterozygotes.
Calcium imaging. The [Ca2+]i was recorded from single-granule neurons as described previously (Mateo et al., 1998), with modifications. Briefly, the cells were loaded with 5 μm fura-2 AM (Invitrogen) for 45 min at 37°C. The coverslip was placed in a small superfusion chamber and superfused with Locke's medium (in mm: 140 NaCl, 2.5 CaCl2, 1.2 KH2PO4, 1.2 MgSO4, 5.5 glucose, and 10 HEPES), pH 7.4, containing different KCl concentrations (5, 25, or 60 mm) depending on the assay. The wavelength of the incoming light was selected using a multiple excitation microfluorescence system (PerkinElmer). Cells were excited at 340 and 380 nm, and the images were collected at 1.5 s intervals using a Nikon (Tokyo, Japan) TE-200 microscope with a 20× Plan Fluor 0.5 numerical aperture objective. Emitted light was isolated by a dichroic mirror (430 nm) and driven to an Ultrapix 2000 Mono CCD camera after passing through a 510 nm bandpass interference filter (Omega Optical, Brattle-boro, VT). The time course data represent the average light intensity in a small elliptical region within each cell. In vitro calibration of the system was performed by recording the fluorescence from small droplets of fura-2 dissolved in intracellular solution (100 mm KCl, 10 mm NaCl, 1 mm MgCl2, 2 mm CaCl2, 1-2 m fura-2, and 10 mm MOPS), pH 7.0, supplemented with 2.5 mm Ca2+ (saturating Ca2+) or 2.5 mm EGTA (zero Ca2+) and using the ratio equation derived by Grynkiewicz et al. (1985).
Results
DREAM downregulates transcription of the NCX3 promoter
To understand the regulatory mechanisms controlling NCX3 expression in vivo, we inspected the proximal regulatory region of the NCX3 gene and found several DRE elements (GTCA) that could be targeted by the transcriptional repressor DREAM and mediate Ca2+-induced derepression after neuronal depolarization (Fig. 1A). In particular, a tandem repeat of DRE elements is located immediately downstream of the TATA box at position 7 relative to the transcription start site. To investigate whether DREAM has a role in the regulation of NCX3 gene expression, we first checked the functionality of this doublet of DREs using electrophoretic mobility shift assays with nuclear extracts of SH-SY5Y cells and an oligonucleotide probe encompassing these two DREs (Fig. 1B). We observed a retarded protein-DNA complex, which could be competed by a tenfold to 100-fold excess of a nonlabeled oligonucleotide encompassing the DRENCX3 or the DREICER, which also contains two DRE sites but was unaffected by competition with the cold unrelated oligonucleotides, CRE and Sp1 (Fig. 1B and data not shown). In addition, the specific DRENCX3 retarded band was blocked by increasing concentrations of Ca2+ (10-50 μm) added to the incubation (Fig. 1B).
Figure 1.
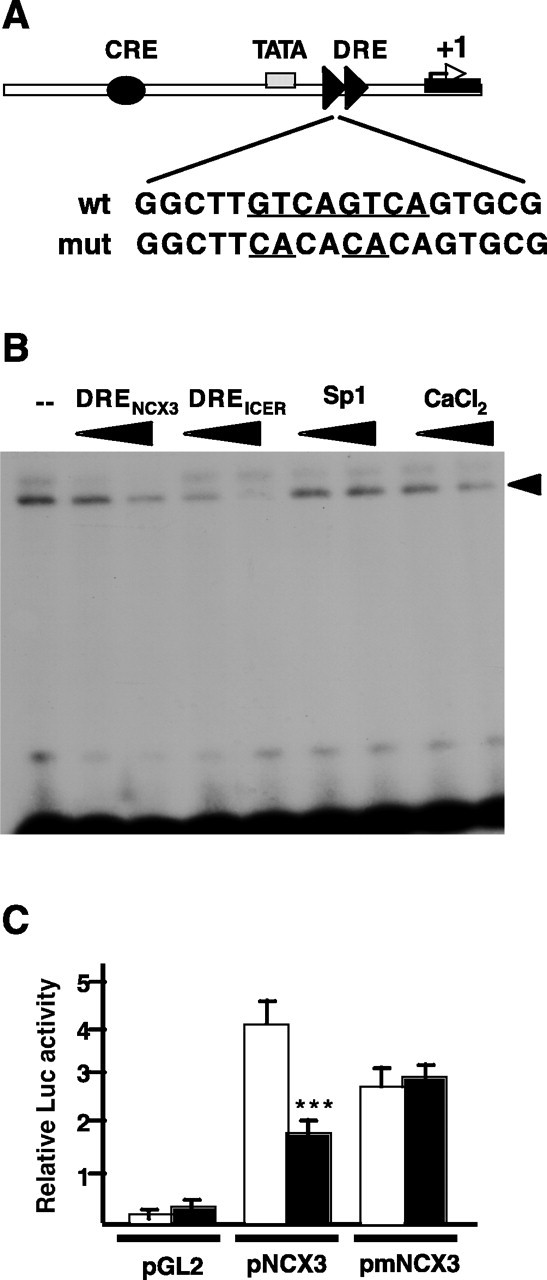
DREAM binds to and represses the NCX3 promoter. A, Schematic representation of the NCX3 promoter. The arrow marks the transcription start site (+1), the position of the tandem DREs is indicated by arrowheads, and the black ellipse represents an upstream CRE element. The sequence of the DRE doublet immediately downstream from the TATA box is depicted, as well as the mutations used to eliminate regulation by DREAM. B, Electrophoretic mobility shift analysis using nuclear extracts from SH-SY5Y cells and the doublet DRE of the NCX3 promoter as a probe. Competition with related cold oligonucleotides containing DREs (NCX3 and ICER) and lack of competition with the unrelated Sp1 oligonucleotide is shown. Addition of increasing concentrations of calcium blocked the appearance of the specific DRE-retarded band. C, Transient transfections in HEK293 cells showing repression of basal transcription from the NCX3 reporter plasmid by DREAM. Basal reporter activity (open bars) and the activity after DREAM coexpression (black bars) is shown for empty reporter (pGL2), NCX3 reporter (pNCX3), and mutated NCX3 containing the mutation of the DRE doublet (pmNCX3). Asterisks represent statistically significant differences between the means relative to corresponding controls (***p < 0.001; Student's t test). wt, Wild type; mut, mutant; Luc, luciferase.
To evaluate the contribution of the doublet DRE to the potential transcriptional repression of the NCX3 promoter by DREAM, a reporter plasmid containing the minimal NCX3 promoter and exon 1 was prepared (Fig. 1A). Transient cotransfection of the NCX3 reporter with DREAM were performed in HEK293 cells, a heterologous system that expresses neither the NCX3 nor the DREAM genes. Cotransfection with DREAM in these cells resulted in 44% lower activity of the NCX3 reporter, compared with control cells transfected with empty vector (Fig. 1C). Furthermore, to confirm that binding to the doublet of DREs next to the TATA box was responsible for the DREAM-mediated repression of the promoter, we performed parallel experiments with a NCX3 reporter bearing a double point mutation at each of the two DRE cores (Fig. 1A). The mutation completely blocked the repressor activity of DREAM on the NCX3 reporter (Fig. 1C). Thus, the doublet of DRE sites participates in the repression of the NCX3 gene by DREAM.
NCX3 is downregulated in SH-SY5Y lines stably overexpressing DREAM or EFmDREAM
SH-SY5Y cells express both the NCX3 (Gabellini et al., 2002) and DREAM (Carrion et al., 1998) proteins. To investigate whether the transcriptional downregulation by DREAM observed in the reporter assays was also operative on the endogenous NCX3 gene, we analyzed the levels of the NCX3 protein in stably transfected SH-SY5Y clones overexpressing different levels of wild-type DREAM or of the Ca2+-insensitive mutant EFmDREAM. Immunoblots with an antibody raised against peptide E276-E722 of the rat NCX3 sequence (Nicoll et al., 1996) revealed a robust immunoreactive band at ∼100 kDa, which corresponds to the mass of the human NCX3 protein (Gabellini et al., 2002). Importantly, the intensity of the band decreased by 25 and 54% in the clones with a twofold and sixfold increase in wild-type DREAM expression, respectively (Fig. 2). Overexpression of EFmDREAM produced a greater downregulation of the NCX3 protein, which was 62 and 84% in clones with only a twofold and fourfold increase in DREAM immunoreactivity, respectively (Fig. 2). The endogenous expression levels of NCX1 and NCX2 proteins in SH-SY5Y cells were lower than NCX3 levels and were not significantly changed by the overexpression of DREAM or EFmDREAM (data not shown). Furthermore, the levels of CREB protein, which were used as a control, remained constant after DREAM or EFmDREAM overexpression (Fig. 2).
Figure 2.

Downregulation of the NCX3 protein in SH-SY5Y cell lines overexpressing DREAM or EFmDREAM. Western blots for DREAM, NCX3, and CREB proteins using whole-cell extracts prepared from mock transfected SH-SY5Y cells, from two SH-SY5Y clones overexpressing DREAM, and from two SH-SY5Y clones overexpressing EFmDREAM. The fold DREAM overexpression as well as the percentage downregulation of the NCX3 protein in stably transfected clones were calculated after densitometric analysis. Expression levels of endogenous CREB were used as a control.
EFmDREAM downregulates the expression of NCX3 in mouse brain
To examine whether the downregulation of the NCX3 gene in cultured cells also occurs in vivo, we analyzed the levels of NCX3 mRNA and protein in specific brain regions from transgenic mice overexpressing EFmDREAM under the control of the CaMKIIα promoter. EFmDREAM transgenic mice were viable and developed normally to adulthood. Depending on the expression pattern of the transgene in different brain areas, several founders were selected and transgenic lines were established. Two lines, Tg-31 and Tg-33, were used in this study. Adult mice from both lines showed normal brain morphology and were not different from wild-type littermates in locomotor performance or activity. Line Tg-31 showed the predicted forebrain expression pattern for the CaMKIIα promoter (Mayford et al., 1996), with some expression of the EFmDREAM transgene in the hippocampus and no expression in the cerebellum (Fig. 3A). Line Tg-33, instead, had a broader distribution of the expression of the DREAM mutant, which was particularly high in the hippocampus but was expressed at significant levels also in the cerebellum (Fig. 3A). Quantitative real-time PCR analysis of the NCX3 transcript levels revealed a significant reduction of NCX3 mRNA in the hippocampus in both transgenic lines (Fig. 3B). Thus, the low level of EFmDREAM expression in the hippocampus of line Tg-31, which is comparable with the level of endogenous DREAM (data not shown), was enough to reduce NCX3 expression to the same extent as the decrease in the hippocampus of line Tg-33. In the cerebellum, a significant reduction of NCX3 mRNA was observed only in line Tg-33, in agreement with the cerebellar expression of the EFmDREAM transgene in this line and the absence in the Tg-31 line (Fig. 3B). To confirm the reduction in mRNA levels, the content of NCX3 protein was then quantified in cerebellar membranes purified from Tg-33 mice expressing EFmDREAM, from transgenic Tg-31 mice not expressing EFmDREAM in the cerebellum, and from wild-type mice having the same genetic background. The intensity of the 100 kDa NCX3 band decreased by ∼30% in the Tg-33 transgenic mice with respect to wild-type or Tg-31 mice (Fig. 3C). The reduction was specific for NCX3, because the expression of EFmDREAM in the cerebellum of Tg-33 mice failed to alter the levels of NCX2 and NCX1 proteins (Fig. 3C). Thus, the specificity of DREAM regulation for NCX3 was confirmed in vivo.
Figure 3.
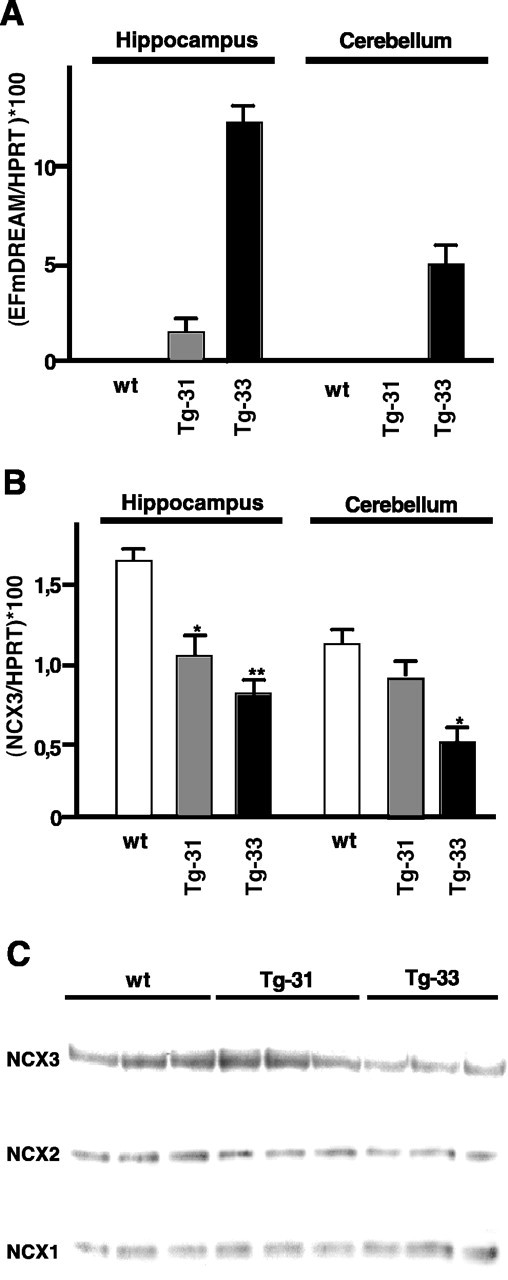
Downregulation of NCX3 mRNA and protein in hippocampus and cerebellum of mice overexpressing EFmDREAM. Quantitative real-time PCR showing the expression levels of EFmDREAM (A) and NCX3 (B), in hippocampus and cerebellum from wild-type (wt) and EFmDREAM transgenic mice. Values were normalized by the content of HPRT mRNA. Asterisks represent statistically significant differences between the means relative to corresponding controls (**p < 0.01; *p < 0.05; Student's t test). C, Western blots using cerebellar membranes from three wild-type mice, three transgenic mice not expressing EFmDREAM in the cerebellum (Tg-31), and three transgenic mice expressing EFmDREAM in the cerebellum (Tg-33) were hybridized with antibodies directed to the NCX3 and NCX2 proteins (100 kDa) and to the NCX1 protein (110 kDa).
Intracellular free [Ca2+]i in cultured cerebellar granules from EFmDREAM transgenic mice is increased
The NCX3 protein is the main Na+/Ca2+ exchanger expressed in cerebellar granules and, together with the plasma membrane calcium (PMCA) pumps, is responsible for a significant portion of the Ca2+ extrusion and thus for the control of calcium homeostasis in these neurons. In keeping with the results in whole cerebellum, the expression of NCX3 mRNA in freshly dissociated cerebellar granule neurons from Tg-33 mice was significantly reduced compared with that from wild-type and Tg-31 mice (data not shown). To investigate the physiological consequences of the NCX3 downregulation induced by the overexpression of EFmDREAM free [Ca2+]i were measured in primary cultures of Tg-33 and wild-type cerebellar granules. Because cytosolic Ca2+ levels in wild-type cerebellar granules are directly dependent on the extracellular concentration of K+ used in the culture medium (Gallo et al., 1987), we compared transgenic and wild-type granules cultured in media containing either a low (5 mm) or mildly elevated (25 mm) concentration of extracellular K+.
In resting conditions, the free [Ca2+]i in wild-type and Tg-33 granules exposed to mild membrane-depolarizing conditions was 247 ± 19 and 305 ± 18 nm, respectively (Fig. 4A). As expected, in low K+ culturing conditions, the free [Ca2+]i was lower both in wild-type granules (138 ± 13 nm) and in Tg-33 granules (205 ± 14 nm) (Fig. 4B). When a stronger membrane depolarization was induced with 60 mm extracellular K+, both transgenic and wild-type granules responded with a rapid increase in intracellular Ca2+ (Fig. 4A,B). Importantly, however, after reestablishment of either 5 or 25 mm extracellular K+ culturing conditions, a rapid decrease of free [Ca2+]i was observed in both culture types, but the decrease was less pronounced in Tg-33 granules (Fig. 4A,B). Thus, the lower expression of NCX3 in transgenic granules is associated with a reduced capacity to extrude Ca2+.
Figure 4.
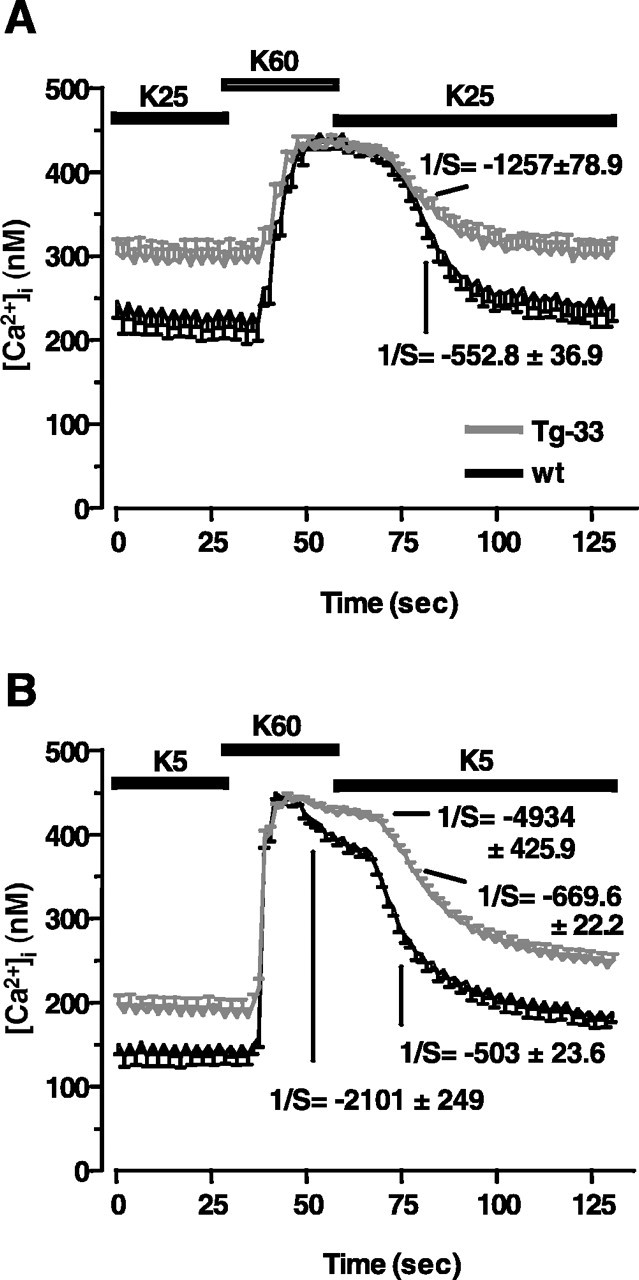
Handling of cytosolic free [Ca2+]i in transgenic cerebellar granules. A, B, Fluorimetric analysis of fura-2-loaded cerebellar granules in cultures maintained in high (A; 25 mm) or low (B; 5 mm) extracellular K+. In both cases, the Ca2+ levels were monitored before inducing their elevation with a depolarizing pulse of 60 mm K+. The kinetics of the decrease of the Ca2+ level after reestablishment of the basal KCl culturing conditions (either 5 or 25 mm) is shown as the inverse of the slope (1/S) of repolarization. Each time course data represents the mean ± SD of the calcium response recorded from at least 40 neurons. wt, Wild type.
Viability is altered in cultured cerebellar granules from EFmDREAM transgenic mice
The long-term survival of cerebellar neurons in culture is dependent on the increased level of free cytosolic Ca2+ produced by partial depolarization of the plasma membrane (Gallo et al., 1987). Thus, we compared the viability of transgenic Tg-33 cerebellar granules with that of wild-type granules in different culturing conditions. The reduced NCX3 expression was associated with reduced viability of Tg-33 cerebellar granules maintained for 3 d in 25 mm KCl compared with wild-type granules (Fig. 5A). As expected from previous work (Gallo et al., 1987), the number of surviving neurons in wild-type cultures maintained for 3 d in low extracellular K+ was lower than in high extracellular K+. However, the number of cells in Tg-33 cultures surviving in low K+, which was ∼45%, was actually higher than in wild-type cultures, in which survival was 23% (Fig. 5A,B). To better correlate the neuronal loss in the cultures with the specific disappearance of transgenic granules, we analyzed the expression level of EFmDREAM, taken as an indirect index of cell viability, in cultures maintained in different conditions and at different times in culture. The total expression level of the mutant DREAM in the cultures maintained in depolarizing conditions (25 mm KCl), decreased sharply to 40% between 0 and 3 d, reflecting the decreased percentage of transgenic neurons surviving in the depolarized cultures. The EFmDREAM mRNA content was further reduced, to 30%, in transgenic granules cultured for 6 d (Fig. 5C). In contrast, the expression of EFmDREAM in Tg-33 granules maintained at low extracellular K+ decreased less between 0 and 3 d, to 75%, and was decreased by only ∼50% after 6 d in vitro (Fig. 5C). This evidently reflects the higher percentage of surviving Tg-33 neurons maintained in low extracellular K+. To support the suggestion that the decrease in the expression of EFmDREAM mRNA in the cultures is specifically associated with the increased death of transgenic neurons and thus could be considered an index of cell viability, the cultures were exposed to nimodipine, a blocker of voltage-dependent Ca2+ channels that prevents the Ca2+ overload caused by reduced NCX3 expression in Tg-33 neurons. Nimodipine indeed totally prevented the neuronal loss and the reduction in EFmDREAM expression in Tg-33 cultures (Fig. 5C and data not shown). Together, these results suggest that the reduced ability of transgenic granules to extrude calcium as a consequence of the reduced expression of NCX3 exchanger results in their increased vulnerability to membrane depolarizing conditions. However, in the presence of low extracellular K+, the reduction in NCX3 expression in Tg-33 neurons evidently helped to maintain the level of intracellular Ca2+ needed for cell viability.
Figure 5.
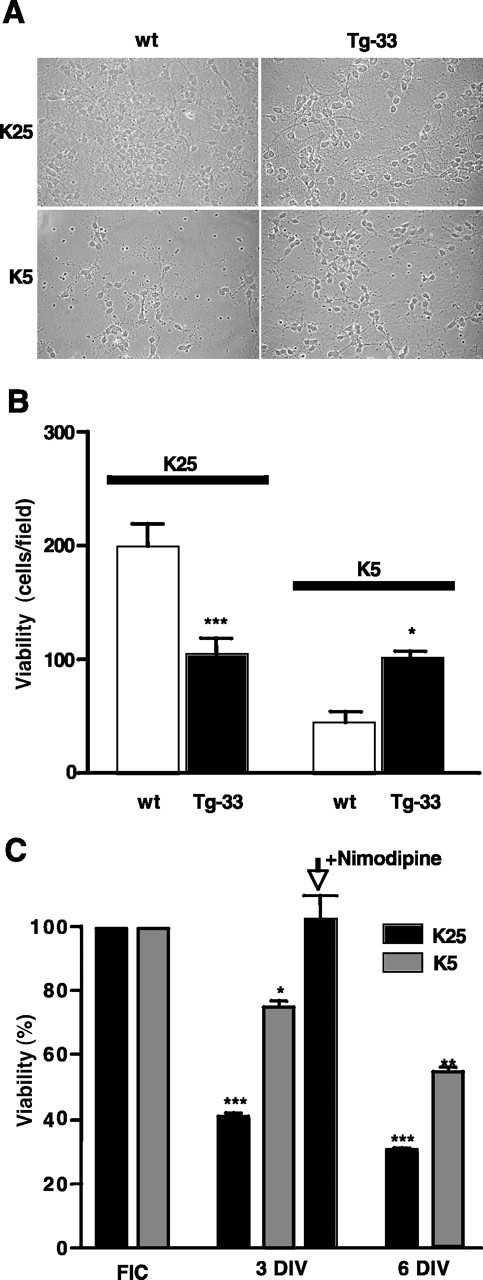
Differential effects of extracellular potassium concentrations on the viability in EFmDREAM transgenic neurons. A, Phase-contrast light micrographs of cerebellar neurons from wild-type (wt) or Tg-33 mice cultured for 3 d in vitro (3 DIV) in media containing either 25 or 5 mm KCl. B, Quantitation of cerebellar granules from wild-type or Tg-33 transgenic mice after 3 d in culture using 4′,6′-diamidino-2-phenylindole dihydrochloride (DAPI) staining. The bars represent the mean ± SEM of the total number of DAPI-labeled cells in different fields. Six different fields in four different cultures were counted. *p < 0.05; ***p < 0.001, one-way ANOVA test. C, Quantitation of the viability of transgenic granules estimated indirectly by real-time reverse transcription-PCR of EFmDREAM mRNA in cultured Tg-33 cerebellar granules maintained under depolarizing (25 mm KCl) or basal (5 mm KCl) conditions. The results from freshly isolated cells (FIC) and from cells at 3 and 6 DIV correspond to the mean ± SEM of six experiments using different cultures. Significant differences from control (EFmDREAM expression in FIC) are indicated as *p < 0.05, **p < 0.01, and ***p < 0.001 (one-way ANOVA).
Recovery of the Ca2+-extruding function restores viability in Tg-33 cerebellar granules cultured in mild membrane-depolarizing conditions
To directly test the suggestion that the reduced expression of NCX3 by EFmDREAM is determining neuronal viability, we forced the expression of NCX exchangers (NCX2 or NCX3) in cultured cerebellar granules and analyzed their viability. Two experimental approaches were used. In the first, the reduced levels of NCX3 in transgenic cerebellar granules were compensated by increasing NCX2 expression by culturing the cells in the presence of calcineurin inhibitors. The expression of the NCX2 exchanger dramatically increased in wild-type mouse cerebellar granules exposed to FK506, CsA, or a combination of both (Fig. 6A and data not shown), as described previously in cultured rat cerebellar granules (Li et al., 2000). The increase in the expression of NCX2 was also augmented in transgenic granules, in which the effect of calcineurin inhibition at day 3 was actually even more prominent (Fig. 6A). The upregulation of NCX2 resulted in the complete block of the disappearance of transgenic granules after 3 or 5 d of culture under mild depolarizing conditions, as measured by the expression levels of EFmDREAM (Fig. 6B). In the second approach, the level of NCX3 protein in transgenic cerebellar granules was increased by infecting the cultures with lentiviral particles encoding the rat NCX3 protein. Quantitative real-time PCR for total NCX3 mRNA showed a peak of NCX3 expression 2 d after the infection and still high expression levels after 4 d (Fig. 7A). No significant changes in endogenous mouse NCX3 mRNA were detected at different times after viral infection (data not shown). The increased levels of NCX3 resulted in the complete protection of transgenic cerebellar granules from the deleterious effects of high K+ concentration at 2 d after infection and in a significant protection at 4 d (Fig. 7B). Not surprisingly, the overexpression of NCX3 in transgenic granules restored their normal vulnerability to low K+ culturing conditions. At both 2 and 4 d after infection, the number of viable transgenic granules maintained in low K+, as judged by the expression levels of EFmDREAM, was significantly lower than in noninfected cultures (Fig. 7B). Nonspecific effects attributable to the viral infection procedure were controlled for by parallel infection with a lentivirus encoding the fluorescent tracer GFP, which was also useful for estimating the success of the infection protocol in each experiment. Infection with GFP-encoding lentiviral particles did not modify NCX3 expression levels or neuronal survival, regardless of culturing conditions (Fig. 7B).
Figure 6.

Induced NCX2 expression restores the viability of transgenic cerebellar granules cultured in high K+. A, Quantitative real-time PCR showing the temporal expression pattern of NCX2 mRNA in cultured cerebellar granule cells from wild-type (wt; open bars) or transgenic Tg-33 (black bars) mice cultured in the absence or presence of calcineurin inhibitors (FK506+CsA). **p < 0.01 and ***p < 0.001 (one-way ANOVA). B, Quantitation of cell viability estimated indirectly by real-time PCR of EFmDREAM expression in cerebellar transgenic granules cultured in high K+ (black bars) in the absence or presence of FK506 and CsA. Significant differences from nontreated cultures are indicated as **p < 0.01 and ***p < 0.001 (one-way ANOVA). For comparison, the expression of the transgene at different days in vitro (DIV) is shown in low K+ culturing conditions (gray bars). FIC, Freshly isolated cells.
Figure 7.

Lentiviral-induced rescue of NCX3 expression restores the viability of transgenic cerebellar granules cultured in high K+. A, Expression of NCX3 at different times after lentiviral infection. B, The viability of transgenic granules was indirectly quantified by real-time PCR of EFmDREAM expression in cerebellar transgenic granules cultured in high (black bars) or low (gray bars) K+. Results from cultures 2 or 4 d after infection with NCX3-encoding lentiviral particles (Lv-NCX3) or control virus encoding the GFP marker (Lv-GFP) are shown. Results are the mean ± SEM from three separate experiments. Significant differences from control noninfected cultures for each culturing condition are indicated as *p < 0.05, **p < 0.01, and ***p < 0.001 (one-way ANOVA). DIV postinf., Days in vitro postinfection.
Discussion
This study has shown that the Ca2+-modulated transcriptional repressor DREAM participates in the regulation of the NCX3 gene. Although the number of potential target genes regulated by DREAM has been increased recently (Sanz et al., 2001; Link et al., 2004; Rivas et al., 2004; Savignac et al., 2005; Scsucova et al., 2005), the regulation by DREAM of NCX3 is of particular interest because it describes a new self-regulatory loop by which the Ca2+ signal may control the expression, and hence the activity, of a protein that is vital for the maintenance of intracellular Ca2+ homeostasis in neurons. The work has shown that this regulatory mechanism is potentially operational in vivo, because overexpression of a Ca2+-insensitive DREAM mutant in transgenic mice downregulates NCX3 mRNA and protein levels in the cerebellum and hippocampus.
As shown previously for other Ca2+-activated genes such as c-fos, ICER, AA-NAT (arylalkylamine N-acetyltransferase), interleukin-2 (IL-2), and IL-4 (Carrion et al., 1999; Link et al., 2004; Savignac et al., 2005), in the NCX3 promoter, DREAM binds, in a Ca2+-dependent manner, to a doublet site composed by two repeats of the DRE element located downstream from the TATA box. In the cases of the prodynorphin and the fra-2 promoters, the binding of DREAM to a single DRE site downstream from the TATA box is sufficient to repress transcription (Carrion et al., 1998; Carrion et al., 1999; Link et al., 2004). Whether the presence of several GTCA core cassettes may contribute to the recruitment of a different repressosome and to a tighter repressive control of target genes is presently under investigation. Sequence analysis of the regulatory region of the NCX3 gene has revealed additional DRE sites upstream from the TATA box and within exon 1. The contribution of these sites to DREAM-mediated control of the NCX3 gene transcription has not been analyzed in the present study. The transcriptional effects of DREAM on target genes bearing DRE sites upstream from the TATA box may require the interaction between DREAM and other nucleoproteins, as shown for the thyroglobulin promoter at which DREAM binds DNA and interacts with thyroid transcription factor 1 to block its transactivating capacity (Rivas et al., 2004) and for the vitamin D-induced regulation of the p21 promoter (Scsucova et al., 2005).
A cAMP-dependent regulation of the NCX3 promoter has been associated with the presence of a CRE site (Gabellini et al., 2003). CRE-dependent transcriptional activation involves primarily the PKA- and/or Ca2+-dependent phosphorylation of Ser-133 in CREB and the recruitment of CBP (Parker et al., 1996; Goodman and Smolik, 2000). In addition, it has been shown that the Ca2+-dependent interaction between DREAM and the kinase-inducible domain in CREB blocks the recruitment of CBP and CRE-dependent transcription (Ledo et al., 2002). Because the EFmDREAM mutant has an intact leucine-charged residue rich domain, which is responsible for the interaction with CREB but does not respond to Ca2+ stimulation, it should heterodimerize with CREB and function as a dominant mutant to also block the CRE-dependent transcription of the NCX3 gene. Thus, the synergistic stimulation of the NCX3 gene by Ca2+ and cAMP signaling would be blocked by the dominant EFmDREAM. Future studies using other transgenic lines overexpressing DREAM mutants unable to interact with CREB/CREM proteins will address the relative contribution of these two major signaling pathways to the regulation of NCX3 gene expression.
The expression of the NCX2 exchanger in cultured cerebellar granules has been shown previously to also be regulated by calcium, albeit in an opposite way (i.e., the expression of NCX2 is rapidly and massively downregulated by Ca2+ in a process involving the Ca2+-dependent phosphatase calcineurin) (Li et al., 2000). Whether the downregulation of the NCX2 gene operates in vivo is presently unknown, as is the physiological meaning of the opposite regulation of the two exchangers by Ca2+. It is worth noting that, in cultured cerebellar granules, NCX2 is only responsible for about one-third of the total Na+/Ca2+ exchanger activity (Li et al., 2000), and in situ hybridization analysis has shown that the expression of NCX3 is predominant in rat cerebellum, in which it is expressed at higher levels than NCX2 and NCX1 (Canitano et al., 2002). Our results have shown that the regulation by DREAM is specific for the NCX3 gene, because the DREAM mutant does not affect the transcription of NCX1 and NCX2.
The mechanism by which the modest increase of intracellular Ca2+ induced by the partial depolarization of the plasma membrane promotes the long-term survival of cerebellar granules in culture is obscure, although the involvement of the protein kinase B pathway has been suggested (Yano et al., 1998). The increase in the set point of Ca2+ homeostasis to ∼200 nm, which is generated by the partial depolarization of the plasma membrane in the cultured neurons, is assumed to correspond in vivo to the repetitive firing of mossy neurons and evidently demands a complex reorganization of the pattern of expression of Ca2+ transporters. Changes in the expression pattern of the PMCA pumps (Guerini et al., 2000) and of the 1,4,5-inositol trisphosphate receptor (Genazzani et al., 1999) have been described, but changes involving NCXs, namely the replacement of NCX2 with NCX3, are most prominent in this reorganization. Unfortunately, no methods are at the moment available to assess possible functional differences among the NCX proteins. However, the finding that neurons expressing significantly lower levels of NCX3 as a consequence of the irreversible downregulation by mutant DREAM have special propensity to succumb when challenged with an increased entrance of Ca2+ is important. These neurons apparently become unable to buffer the increased influx of Ca2+ in a way that would lead to the set point necessary for survival without letting it deviate to unacceptably high levels. Conversely, the work presented here has shown that the reduced capacity to extrude Ca2+ helps to maintain the intracellular Ca2+ concentration at the modestly increased level that is required for survival when the neurons are hyperpolarized by the low extracellular K+ concentrations. In essence, the Ca2+-dependent effect of endogenous DREAM on the transcription of the NCX3 gene could be seen as a safety device that can be switched on or off to maintain intracellular Ca2+ at an appropriate level.
The set point of nuclear Ca2+ that directs the binding or unbinding of DREAM from DRE sites may or may not be directly related to the affinity for Ca2+ measured in vitro for recombinant DREAM (Osawa et al., 2001; Craig et al., 2002), because it could be influenced by conditions prevailing in the nucleoplasm. Among them, local microdomains of very low Ca2+ concentrations or other factors that may affect the ability of DREAM to bind Ca2+ in vivo. These uncertainties will have to be clarified by future work, but the crucial role of DREAM in the control of NCX3 gene expression and the role of NCX3 in the control of Ca2+ homeostasis, shown by the results presented here, are of considerable interest. The negative consequences of the downregulation of NCX3 function by the DREAM mutant in transgenic cerebellar granules is in line with our recent work showing that excitotoxic death of cerebellar granules is mediated by the cleavage of the NCX3 protein (Bano et al., 2005). Furthermore, transient downregulation of the NCX3 protein in cerebellar granules using specific small interfering RNA compromised neuronal Ca2+ handling, as it did in transgenic DREAM mutant cerebellar granules, transforming the Ca2+ transient elicited by nonexcitotoxic glutamate concentrations into a lethal Ca2+ overload (Bano et al., 2005). Future gene profile analysis of transgenic cerebellar neurons may reveal other genes downregulated by the expression of EFmDREAM, which, in principle, could contribute to the effects described here. Nevertheless, the functional recovery achieved by increasing NCX2 expression, or by the lentiviral-mediated overexpression of NCX3 protein, convincingly demonstrates that the reduction of NCX3 expression is a crucial determinant for the decreased viability of transgenic cerebellar granules cultured under membrane depolarizing conditions.
Footnotes
This work was supported by Human Frontier Science Program Grant RGP0156/2001B to J.R.N. and E.C., by grants from the Fundación La Caixa, Ministerio de Educación y Ciencia (MEC), Fondo de Investigación Sanitaria, and Comunidad Autonoma de Madrid to J.R.N., and by grants from the Italian Ministry of Education, University, and Research (PRIN and Foreign Investment Review Board) to E.C. R.G.-V. and M.S. were supported by the Juan de la Cierva Program from MEC and an individual Marie Curie Fellowship, respectively. We thank Dr. M. T. Miras-Portugal for access to Ca2+ fluorometric equipment and Paz Gonzalez and David Campos for technical assistance. The NCX3 antibody was a generous gift from Dr. K. D. Philipson (Department of Physiology, University of California, Los Angeles, Los Angeles, CA).
Correspondence should be addressed to either of the following: Dr. Ernesto Carafoli, Venetian Institute of Molecular Medicine, Via Orus 2, 35129 Padova, Italy, E-mail: ernesto.carafoli@unipd.it; or Jose R. Naranjo, Departamento Biologia Molecular y Celular, Centro Nacional de Biotecnologia-Consejo Superior de Investigaciones Científicas, Campus de Cantoblanco, 28049 Madrid, Spain, E-mail: naranjo@cnb.uam.es.
Copyright © 2005 Society for Neuroscience 0270-6474/05/2510822-09$15.00/0
R.G.-V., B.T., and J.B. contributed equally to this work.
References
- Bano D, Young KW, Guerin CJ, Lefeuvre R, Rothwell NJ, Naldini L, Rizzuto R, Carafoli E, Nicotera P (2005) Cleavage of the plasma membrane Na+/Ca2+ exchanger in excitotoxicity. Cell 120: 275-285. [DOI] [PubMed] [Google Scholar]
- Brewer GJ (1997) Isolation and culture of adult rat hippocampal neurons. J Neurosci Methods 71: 143-155. [DOI] [PubMed] [Google Scholar]
- Canitano A, Papa M, Boscia F, Castaldo P, Sellitti S, Taglialatela M, Annunziato L (2002) Brain distribution of the Na+/Ca2+ exchanger-encoding genes NCX1, NCX2, and NCX3 and their related proteins in the central nervous system. Ann NY Acad Sci 976: 394-404. [DOI] [PubMed] [Google Scholar]
- Carrion AM, Mellstrom B, Naranjo JR (1998) Protein kinase A-dependent derepression of the human prodynorphin gene via differential binding to an intragenic silencer element. Mol Cell Biol 18: 6921-6929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrion AM, Link WA, Ledo F, Mellstrom B, Naranjo JR (1999) DREAM is a Ca2+-regulated transcriptional repressor. Nature 398: 80-84. [DOI] [PubMed] [Google Scholar]
- Craig TA, Benson LM, Venyaminov SY, Klimtchuk ES, Bajzer Z, Prendergast FG, Naylor S, Kumar R (2002) The metal-binding properties of DREAM: evidence for calcium-mediated changes in DREAM structure. J Biol Chem 277: 10955-10966. [DOI] [PubMed] [Google Scholar]
- Follenzi A, Naldini L (2002) Generation of HIV-1 derived lentiviral vectors. Methods Enzymol 346: 454-465. [DOI] [PubMed] [Google Scholar]
- Gabellini N, Zatti A, Carafoli E (2000) The Na+/Ca2+ exchanger: structural aspects, function and regulation. In: Calcium homeostasis (Carafoli E, Krebs J, eds), pp 173-188. Berlin: Springer.
- Gabellini N, Bortoluzzi S, Danieli GA, Carafoli E (2002) The human SLC8A3 gene and the tissue-specific Na+/Ca2+ exchanger 3 isoforms. Gene 298: 1-7. [DOI] [PubMed] [Google Scholar]
- Gabellini N, Bortoluzzi S, Danieli GA, Carafoli E (2003) Control of the Na+/Ca2+ exchanger 3 promoter by cyclic adenosine monophosphate and Ca2+ in differentiating neurons. J Neurochem 84: 282-293. [DOI] [PubMed] [Google Scholar]
- Gallo V, Kingsbury A, Balazs R, Jorgensen OS (1987) The role of depolarization in the survival and differentiation of cerebellar granule cells in culture. J Neurosci 7: 2203-2213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Genazzani AA, Carafoli E, Guerini D (1999) Calcineurin controls inositol 1,4,5-trisphosphate type 1 receptor expression in neurons. Proc Natl Acad Sci USA 96: 5797-5801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodman RH, Smolik S (2000) CBP/p300 in cell growth, transformation, and development. Genes Dev 14: 1553-1577. [PubMed] [Google Scholar]
- Grynkiewicz G, Poenie M, Tsien RY (1985) A new generation of Ca2+ indicators with greatly improved fluorescence properties. J Biol Chem 260: 3440-3450. [PubMed] [Google Scholar]
- Guerini D, Wang X, Li L, Genazzani A, Carafoli E (2000) Calcineurin controls the expression of isoform 4CII of the plasma membrane Ca2+ pump in neurons. J Biol Chem 275: 3706-3712. [DOI] [PubMed] [Google Scholar]
- Kofuji P, Hadley RW, Kieval RS, Lederer WJ, Schulze DH (1992) Expression of the Na-Ca exchanger in diverse tissues: a study using the cloned human cardiac Na-Ca exchanger. Am J Physiol 263: C1241-C1249. [DOI] [PubMed] [Google Scholar]
- Ledo F, Carrion AM, Link WA, Mellstrom B, Naranjo JR (2000) DREAM-alphaCREM interaction via leucine-charged domains derepresses downstream regulatory element-dependent transcription. Mol Cell Biol 20: 9120-9126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ledo F, Kremer L, Mellstrom B, Naranjo JR (2002) Ca2+-dependent block of CREB-CBP transcription by repressor DREAM. EMBO J 21: 4583-4592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Guerini D, Carafoli E (2000) Calcineurin controls the transcription of Na+/Ca2+ exchanger isoforms in developing cerebellar neurons. J Biol Chem 275: 20903-20910. [DOI] [PubMed] [Google Scholar]
- Link WA, Ledo F, Torres B, Palczewska M, Madsen TM, Savignac M, Albar JP, Mellstrom B, Naranjo JR (2004) Day-night changes in downstream regulatory element antagonist modulator/potassium channel interacting protein activity contribute to circadian gene expression in pineal gland. J Neurosci 24: 5346-5355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mateo J, Garcia-Lecea M, Miras-Portugal MT, Castro E (1998) Ca2+ signals mediated by P2X-type purinoceptors in cultured cerebellar Purkinje cells. J Neurosci 18: 1704-1712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayford M, Bach ME, Huang YY, Wang L, Hawkins RD, Kandel ER (1996) Control of memory formation through regulated expression of a CaMKII transgene. Science 274: 1678-1683. [DOI] [PubMed] [Google Scholar]
- Nicoll DA, Quednau BD, Qui Z, Xia YR, Lusis AJ, Philipson KD (1996) Cloning of a third mammalian Na+-Ca2+ exchanger, NCX3. J Biol Chem 271: 24914-24921. [DOI] [PubMed] [Google Scholar]
- Osawa M, Tong KI, Lilliehook C, Wasco W, Buxbaum JD, Cheng HY, Penninger JM, Ikura M, Ames JB (2001) Calcium-regulated DNA binding and oligomerization of the neuronal calcium-sensing protein, calsenilin/DREAM/KChIP3. J Biol Chem 276: 41005-41013. [DOI] [PubMed] [Google Scholar]
- Parker D, Ferreri K, Nakajima T, LaMorte VJ, Evans R, Koerber SC, Hoeger C, Montminy MR (1996) Phosphorylation of CREB at Ser-133 induces complex formation with CREB-binding protein via a direct mechanism. Mol Cell Biol 16: 694-703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Philipson KD (1999) Sodium-calcium exchange. In: Calcium as a cellular regulator (Carafoli E, Klee C, eds), pp 279-294. New York: Oxford UP.
- Rivas M, Mellstrom B, Naranjo JR, Santisteban P (2004) Transcriptional repressor DREAM interacts with thyroid transcription factor-1 and regulates thyroglobulin gene expression. J Biol Chem 279: 33114-33122. [DOI] [PubMed] [Google Scholar]
- Sanz C, Mellstrom B, Link WA, Naranjo JR, Fernandez-Luna JL (2001) Interleukin 3-dependent activation of DREAM is involved in transcriptional silencing of the apoptotic Hrk gene in hematopoietic progenitor cells. EMBO J 20: 2286-2292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savignac M, Palczewska M, Pintado B, Gutierrez-Adan A, Mellstrom B, Naranjo JR (2005) Transcriptional repressor DREAM regulates T lymphocyte proliferation and cytokine gene expression. EMBO J 24: 3555-3564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scsucova S, Palacios D, Savignac M, Mellstrom B, Naranjo JR, Aranda A (2005) The repressor DREAM acts as a transcriptional activator on Vitamin D and retinoic acid response elements. Nucleic Acids Res 33: 2269-2279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yano S, Tokumitsu H, Soderling TR (1998) Calcium promotes cell survival through CaM-K kinase activation of the protein-kinase-B pathway. Nature 396: 584-587. [DOI] [PubMed] [Google Scholar]
- Zhang X, Odom DT, Koo SH, Conkright MD, Canettieri G, Best J, Chen H, Jenner R, Herbolsheimer E, Jacobsen E, Kadam S, Ecker JR, Emerson B, Hogenesch JB, Unterman T, Young RA, Montminy M (2005) Genome-wide analysis of cAMP-response element binding protein occupancy, phosphorylation, and target gene activation in human tissues. Proc Natl Acad Sci USA 102: 4459-4464. [DOI] [PMC free article] [PubMed] [Google Scholar]