

Abstract
GABAergic septohippocampal neurons play a major role in the generation of hippocampal theta rhythm, but modulatory factors intervening in this function are poorly documented. The neuropeptide somatostatin (SST) may be one of these factors, because nearly all hippocampal GABAergic neurons projecting to the medial septum/diagonal band of Broca (MS-DB) express SST. In this study, we took advantage of the high and selective expression of the SST receptor sst2A in MS-DB to examine its possible role on theta-related activity. Immunohistochemical experiments demonstrated that sst2A receptors were selectively targeted to the somatodendritic domain of neurons expressing the GABAergic marker GAD67 but were not expressed by cholinergic neurons. In addition, a subpopulation of GABAergic septohippocampal projecting neurons expressing parvalbumin (PV) also displayed sst2A receptors. Using in vivo juxtacellular recording and labeling with neurobiotin, we showed that a number of bursting and nonbursting neurons exhibiting high discharge rates and brief spikes were immunoreactive for PV or GAD67 and expressed the sst2A receptor. Microiontophoresis applications of SST and the sst2A agonist octreotide (OCT) showed that sst2A receptor activation decreased the discharge rate of both nonbursting and bursting MS-DB neurons and lessened the rhythmic activity of the latter. Finally, intraseptal injections of OCT and SST in freely moving rats reduced the power of hippocampal EEG in the theta band. Together, these in vivo experiments suggest that SST action on MS-DB GABAergic neurons, through sst2A receptors, represents an important modulatory mechanism in the control of theta activity.
Keywords: neuromodulation, neuropeptide, GABAergic, theta rhythm, medial septum, somatostatin
Introduction
The medial septum-diagonal band of Broca complex (MS-DB) plays a pivotal role in the generation and maintenance of the hippocampal theta rhythm, a periodic field potential implicated in attention and memory (Buzsaki, 2002). The septohippocampal pathway is composed of cholinergic and GABAergic neurons (Kohler et al., 1984; Frotscher and Leranth, 1985), the latter containing the calcium-binding protein parvalbumin (PV) (Freund, 1989). Both cell types exhibit a rhythmically bursting (RB) activity that is considered the principal generator of hippocampal theta rhythm (King et al., 1998; Brazhnik and Fox, 1999; Morris et al., 1999; Borhegyi et al., 2004). The cholinergic component modulates the amplitude of the hippocampal theta, whereas the GABAergic one paces its frequency (Lee et al., 1994; Apartis et al., 1998). GABAergic-PV neurons project to GABAergic hippocampal interneurons that, in turn, phasically disinhibit pyramidal cells (Freund and Antal, 1988; Toth et al., 1997). Reciprocally, GABAergic hippocamposeptal neurons send projections to GABAergic septohippocampal neurons, thus forming an inhibitory loop that reinforces theta-related activity in the septohippocampal system (Toth et al., 1993).
Although the role of the GABAergic component is now better understood, the regulation of the septohippocampal rhythmic activity is a complex mechanism that is not yet clarified. Several in vitro studies have demonstrated that neurotransmitters such as glutamate (Armstrong and MacVicar, 2001), serotonin (Alreja, 1996), noradrenalin (Alreja and Liu, 1996), and neuropeptides including endorphin (Alreja et al., 2000), neurotensin (Matthews, 1999), and hypocretin (Wu et al., 2004) might influence the rhythmogenesis. The neuropeptide somatostatin (SST) is a good candidate to play a role in the regulation of the rhythmic activity, because nearly all hippocampal GABAergic neurons that project to the MS-DB express SST (Gulyas et al., 2003), in agreement with previous experiments that demonstrate SST axon terminals in this structure (Johansson et al., 1984). Thus, the present study aimed to define the cellular targets of SST in the MS-DB and its potential neuromodulatory role on theta activity as well as the identity of the SST receptors by which such effects might be mediated. Because the sst2A receptor accounts for an important facet of brain somatostatinergic transmission (Dournaud et al., 1996, 1998, 2000; Schindler et al., 1997) and because it appears to be the unique subtype expressed in the MS-DB (Helboe et al., 1998; Handel et al., 1999; Schindler et al., 1999; Stroh et al., 1999; Schreff et al., 2000), the sst2A receptor is likely to transduce the hippocamposeptal SST actions. Using immunohistochemical approaches, we first determined the distribution of sst2A receptors in the MS-DB and the neurochemical identity of cells bearing this receptor. Juxtacellular recordings and labeling were performed in vivo to characterize relationships between firing patterns and neurotransmitter phenotypes of sst2A-expressing neurons. We then examined the effects of exogenous activation of sst2A receptors on MS-DB cells using extracellular recordings in urethane-anesthetized animals. Finally, we analyzed the effects of microinjection of sst2A agonists in the MS-DB on theta activity using electroencephalogram (EEG) recordings.
Materials and Methods
Animals
Male Sprague Dawley rats (270-300 g body weight; Iffa-Credo, L'Arbresle, France) were housed and used in conformity with the local Animal Care and Use Committee.
Primary antibodies
The sst2A receptor was immunolocalized by using a fully characterized antiserum raised in rabbit against the C-terminal segment 330-369 of the human protein (Helboe et al., 1997; Helboe and Moller, 1999; Csaba et al., 2001, 2002, 2003). Goat antisera were used to detect choline acetyltransferase (ChAT) (AB144P; 1:100; Chemicon, Temecula, CA) and somatostatin-14 (SST) (D20; 1:500; Santa Cruz Biotechnology, Santa Cruz, CA). Mouse monoclonal antibodies were used to detect the 67 KDa form of glutamic acid decarboxylase (GAD67) (MAB5406; 1:2000; Chemicon), the microtubule-associated protein type 2 (MAP2) (M4403; 1:100; Sigma, St. Louis, MO), the calcium-binding protein parvalbumin (P3088; 1:500; Sigma), and synaptophysin (MAB368; 1:500; Chemicon).
Immunohistochemistry
For immunohistochemical studies, a total of 17 animals were deeply anesthetized with sodium pentobarbital (60 mg/kg, i.p.; Sanofi Pharmaceuticals, Toulouse, France) and perfused through the ascending aorta with 600 ml of 4% paraformaldehyde in 0.1 m phosphate buffer (PB), pH 7.4. For GAD67 detection, animals received one injection of colchicine (100 μg in 10 μl) into the lateral ventricle (Paxinos and Watson, 1986) 48 h before they were killed. Brains were cryoprotected in 30% sucrose in PB (4°C, 24 h), frozen in liquid isopentane at -40°C, coronally sectioned at a thickness of 30 μm at the level of the MS-DB [anteroposterior (AP), 9.6 to 9.2] (Paxinos and Watson, 1986), and collected in PB. For all experiments, free-floating sections were rinsed in Tris-buffered saline (TBS), pH 7.4, and blocked by preincubation in 5% normal donkey serum (NDS) in TBS containing 0.3% Triton X-100 for 30 min. Sections were rinsed in TBS after each incubation. Primary antibodies were incubated overnight at room temperature. To illustrate the distribution of sst2A-expressing cells in the MS-DB, some sections were processed by using the immunoperoxidase method with the tyramide amplification system (TSA-Indirect; NEN Life Sciences, Boston, MA) as described previously (Csaba et al., 2002). For single-, double-, and triple-labeling immunofluorescence microscopy, the rabbit anti-sst2A antiserum (1:2000) was detected using a 45 min incubation in a 1:500 biotinylated donkey anti-rabbit IgG (Jackson ImmunoResearch, West Grove, PA) solution followed by a 20 min incubation in a 1:6000 streptavidin cyanine 3 (Cy3) (Jackson ImmunoResearch) solution. Other primary antibodies were detected using appropriate fluorochrome-conjugated secondary antibodies (1:100; Alexa 488/633; Molecular Probes, Eugene, OR). Control incubations included omission of the primary antibodies to test the non-specific binding of secondary antibodies and incubation with one primary but with secondary antibodies to demonstrate the absence of cross-labeling in double- and triple-labeling experiments. Sections were analyzed by confocal laser-scanning microscopy using a TCS SP2 confocal imaging system equipped with Ar 488 nm, HeNe 543 nm, and HeNe 633 nm lasers (Leica, Heidelberg, Germany). Digital images were collected from a single optical plane using a 100× Plan-Apochromat oil immersion lens (numerical aperture, 1.4). The pinhole setting was 0.75 Airy unit for all images. Double and triple fluorescence images were acquired in sequential mode to avoid potential contamination by linkage-specific fluorescence emission cross-talk. Colors of digital images were chosen to ensure the best illustration rendering. To estimate the overlap between sst2A-expressing cells and GAD67, ChAT, or PV-immunoreactive cell populations in the MS-DB, a profile count was performed in double-labeled sections. Three sections (AP, 9.6, 9.4, 9.2) were analyzed per animal under epifluorescence illumination. Results are expressed as the mean ± SEM of five animals.
Electrophysiology
Unit activity recordings and analysis. Rats were anesthetized with urethane (1.3 g/kg, i.p.) and placed in a stereotaxic frame. A catheter was inserted in the peritoneal cavity to give small additional doses of urethane if necessary to keep the rat at a level of anesthesia optimal for recording the hippocampal theta rhythm. In addition to the general anesthesia, all wounds and pressure points were infiltrated with lidocaine before surgery. Body temperature was maintained with a heating pad controlled by a rectal thermal probe. A monopolar electrode to record EEG was placed in the left dorsal hippocampus [AP, 5; lateral (L), 3; dorsoventral, 6.6] (Paxinos and Watson, 1986). A screw placed on the occipital crest served as reference. The skull was opened above the MS-DB (AP, 9; L, 0), and the central sinus was ligatured and cut. Recorded neurons were located in the midline part of MS-DB, 6000-7500 μm below the brain surface (Paxinos and Watson, 1986), an area that contains a high proportion of PV neurons (Knapp et al., 2000). Extracellular activity was recorded with conventional amplification methods and collected on a personal computer via a Cambridge Electronics Design (Cambridge, UK) interface using the Spike 2 software (sampling rate, 1 kHz) in parallel with analog-to-digital samplings of filtered EEG (bandpass, 1-70 Hz; sampling rate, 100 Hz). The analysis of unit activity included the following: (1) the average discharge rate (number of spikes/s); (2) the duration of spike (measured from the peak of initial negative phase of the action potential to the peak of its subsequent positive phase) (Brazhnik and Fox, 1999); and (3) the autocorrelation histogram, calculated to characterize the RB pattern of activity. If recurrent events such as rhythmic burst of action potentials occur, then the autocorrelation histogram shows periodic sinusoidal-like density peaks (Perkel et al., 1967). An “index” of rhythmic activity (average value of peaks minus average value of troughs divided by average discharge rate) was used to classify cells as RB or non-RB neurons (Apartis et al., 1998). Cells with an index <0.95 were classified as nonrhythmic. The higher index denotes the more regular RB activity. Although an unquestionable classification of neurons cannot be inferred from extracellular signal (Matthews and Lee, 1991; King et al., 1998), an attempt was made to distinguish putative GABAergic neurons on the basis of their firing properties (Brazhnik and Fox, 1999; Borhegyi et al., 2004). Neurons with brief spike (spike duration, ≤0.5 ms), high average discharge rate (≥12 spike/s), and 4-15 spikes/burst in the case of RB activity were assumed to be GABAergic and subsequently recorded.
Juxtacellular recording and labeling. To characterize the neurochemical identity of recorded neurons, juxtacellular labelings with neurobiotin (Vector Laboratories, Burlingame, CA) were performed in 10 rats. After the detection of a unit, the discharge rate, spike duration, and spontaneous or induced RB activity were examined on-line. If these parameters matched those of presumed GABAergic neurons, the electrode was progressively advanced to reach close proximity of the cell. Pulses of anodal current (200 ms; 50% duty cycle) were delivered by way of the microelectrode (filled with 0.5 m sodium acetate and 5% neurobiotin). The current amplitude was increased incrementally from 0.4 to a maximum of 10 nA until the cell firing became entrained (Pinault, 1996). For neurochemical characterization of juxtacellularly labeled neurons, 40-μm-thick septal sections were first incubated in streptavidin Cy3 (1:16,000) for 30 min to reveal neurobiotin. Labeled cells were subsequently processed for immunohistochemistry using anti-sst2A receptor and anti-PV or anti GAD67 antibodies detected with appropriate fluorochrome-conjugated secondary antibodies (1:100; Alexa 488/633; Molecular Probes).
Microiontophoresis. Microiontophoresis experiments were performed in 18 rats. Extracellular activity was recorded using micropipettes (filled with 1 m NaCl) attached to a multibarreled micropipette. Solution for testing by iontophoresis included the sst2A agonist octreotide (OCT) (2 mm, pH 5.5; SMS 201-995; gift from Dr. D. Hoyer, Novartis, Basel, Switzerland), SST (3 mm, pH 5.5; Peninsula Laboratories, San Carlos, CA), glutamate (0.2 m, pH 8; Sigma), and GABA (0.1 m, pH 5; Sigma). Peptides were dissolved in 165 mm NaCl, and other compounds were dissolved in distilled water. Application currents for peptides ranged from 40 to 110 nA, 20 to 40 s. To prevent drug diffusion, retaining currents (5-10 nA) were applied between periods of ejection. Average discharge rates of an individual neuron were determined before, during, and after drug ejection (five episodes of 10 s each time). A significant effect of a drug was defined as a change of at least 25% with respect to the basal discharge rate (Stone, 1985). Data were considered for additional analyses on a condition that the hippocampal EEG remained unchanged both in control conditions and during drug applications. Discharge rates and indexes of rhythmic activity were compared using Student's paired t test. p < 0.05 was considered statistically significant.
EEG recordings and analysis
Rats (n = 12) were anesthetized with pentobarbital (60 mg/kg, i.p.) and placed in a stereotaxic apparatus. Rats were implanted with a guide cannula (5.5 mm 26 gauge stainless-steel tube) placed 1 mm above the MS-DB (AP, 9.5; L, 1.8; right side, angle of 12° off the vertical). A stylet was inserted in the cannula and remained there except during intraseptal injection. Electrodes to record hippocampal EEG were placed as described above and soldered to a connector. The guide cannula and the connector were embedded in a mound of dental cement and jointed to the rat skull. Cephalosporin (cefuroxime, 60 mg/kg; GlaxoSmithKline, Brentford, UK) was administered intramuscularly every 2 d during each week. During the recovery period (7 d), rats were gradually habituated to the recording conditions. They were placed in a Plexiglas chamber under dimmed lighting and connected to a rotating collector by a flexible cable. Hippocampal theta activity was observed during type 1 behavior (exploratory walking, climbing, rearing up on the chamber wall). Elements of the rat's behavior were continuously scored and stored together with EEG. On the day of experiment, hippocampal theta was recorded before and for 90 min after intracerebral injection of sst2A agonists. Injected compounds were as follows: OCT (0.5 nmol/0.25 μl or 1 nmol/0.5 μl; injection rate, 0.12 μl/min), SST (0.5 nmol/0.25 μl or 1 nmol/0.5 μl; injection rate, 0.12 μl/min), or saline (0.5 μl; injection rate, 0.12 μl/min) as control. OCT, SST, or saline was given in a random order at 3 d intervals. At the end of the experiment, rats were deeply anesthetized and perfused as indicated above. Diffusion spread OCT, and SST was assessed by immunohistochemistry using a rabbit polyclonal anti-OCT antibody (Novartis) and D20 anti-SST-14 antibody. Both substances diffused in the MS-DB, sparing adjoining regions (data not shown).
Seven samples of theta activity (7 s each) obtained during strictly similar behavior were selected before and 10, 20, 30, 40, 60, and 90 min after OCT or SST injections and analyzed off-line (Cambridge Electronic Design). The power spectrum of EEG was computed with a fast Fourier transform over a range of 0-20 Hz with 0.1 Hz of resolution. The power of the signal was obtained for the bands 1-6 Hz, 6-9 Hz (theta frequency), and 9-20 Hz and for the total band (1-20 Hz). Relative power was calculated to obtain information on the proportional contribution of each band (expressed as a percentage) to total power. The peak frequency in the 6-9 Hz range was calculated. Averaged values before and at different times after OCT, SST, or saline injections were compared using Student's paired t test. p < 0.05 was considered statistically significant.
Results
Regional and cellular localization of sst2A receptor immunoreactivity in the MS-DB
Microscopic examination of immunoreacted sections with the sst2A receptor antiserum using both chromogenic and fluorescent techniques demonstrated that numerous immunoreactive cell profiles displaying the morphological features of neurons were apparent throughout the rostrocaudal extent of the MS-DB (Fig. 1A,B). In comparison with the MS, sst2A receptor-positive neurons were, however, less numerous in the DB. Receptor-positive neurons ranged from small size (10-15 μm) with a fusiform or oval shape in the MS to medium (15-25 μm) or large (> 25 μm) size with a polygonal shape in the DB (Fig. 1C-E). At the cellular level, single confocal microscopic optical sections demonstrated a preferential distribution of the sst2A receptor labeling at the periphery of somatodendritic cell profiles (Fig. 1F). Receptor immunoreactivity also decorated the edge of medium and small size processes scattered in the neuropil (Fig. 1G,H). To further characterize cellular compartments expressing sst2A receptors, we examined by confocal laser-scanning microscopy double-labeled sections with the sst2A antiserum and with either a marker of dendritic processes, MAP2, or a marker of presynaptic specializations, synaptophysin. As illustrated in Figure 1 (I-K), all of the sst2A receptor-immunoreactive processes also expressed MAP2 labeling. In contrast, sst2A receptor immunoreactivity never appeared colocalized with synaptophysin, although synaptophysin-immunoreactive puncta were often found in close apposition with tangential or cross-sectioned sst2A receptor-positive elements (Fig. 1L-N). Together, these results suggest that the sst2A receptor is targeted to postsynaptic sites in MS-DB neurons at the plasma membrane of cell bodies and proximal and distal dendrites.
Figure 1.
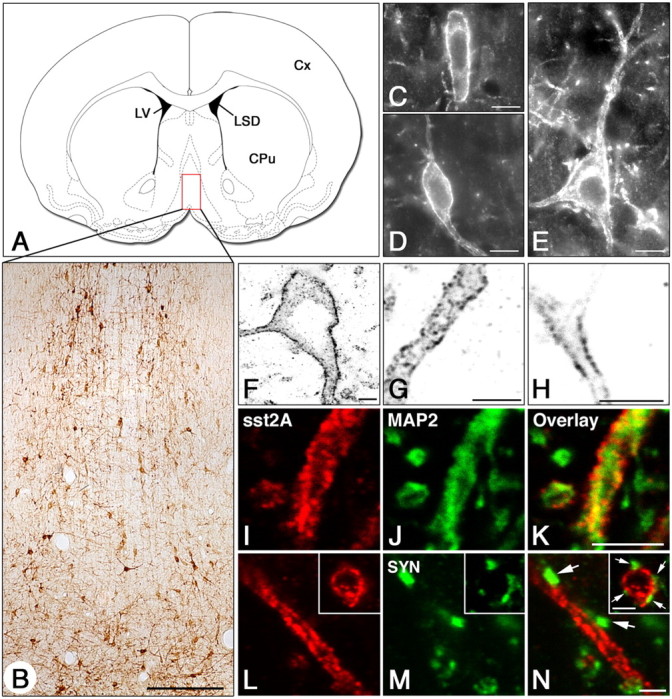
Regional and cellular localization of the sst2A receptor in the rat MS-DB complex as visualized by immunohistochemistry. A, Schematic representation of a frontal section (interaural 9.48) illustrating the localization of the MS-DB. The red frame depicts the region of the MS-DB shown in B. B, Low-power magnification of sst2A-immunoreactive cells using chromogenic labeling. Note that numerous neuronal cell bodies and processes are distributed throughout the dorsoventral extent of the nucleus. C, D, Representative examples of the numerous sst2A-positive fusiform cells found in the MS-DB using fluorescence labeling. E, Albeit less numerous, immunoreactive cells for the sst2A receptor also displayed a polygonal shape. F-H, As evidenced in confocal single-optical sections (false inverted black and white colors), sst2A receptor immunoreactivity decorates the plasmic membrane of somata (F), proximal dendrites (F), and distal processes of medium (G) and small (H) sizes. I-K, Processes immunoreactive for the sst2A receptor (sst2A; I) also express MAP2 immunoreactivity (MAP2; J) as evidenced by the yellow signal observed in the overlay panel (Overlay; K). L-N, In sections double labeled for sst2A (L) and synaptophysin (SYN; M), no colocalization is observed between the two markers (N). Note, however, that SYN-immunoreactive elements are closely apposed to both sst2A-immunoreactive elongated and cross-sectioned (L-N, boxes) processes (N, arrows). Scale bars: B, 100 μm; C-E, 10 μm; (in K) F-K, 4 μm; (in N) L-N, 2 μm. CPu, Caudate-putamen; Cx, cerebral cortex; LSD, dorsolateral septum; LV, lateral ventricle.
GABAergic but not cholinergic neurons express the sst2A receptor
Because the pattern of localization of sst2A receptor-expressing cells in the MS-DB corresponded well to that of both GABAergic and cholinergic neurons, we first investigated whether subpopulations of sst2A receptor-bearing cells expressed the GABAergic marker GAD67 or the cholinergic marker ChAT. As illustrated in Figure 2A, of 2648 sst2A receptor positive-cells analyzed, all expressed GAD67. Conversely, of 9775 GAD67 cells in the MS and 4217 in the DB, 23.4 ± 3.8 and 15.1 ± 2.7% were also positive for the sst2A receptor, respectively. In contrast, of 2701 sst2A-expressing cells analyzed in double-labeled sections with ChAT, colocalization of both markers was not observed, although both cell populations were often observed intermingled throughout the MS-DB complex (Fig. 2B). Having demonstrated that a subpopulation of GABAergic cells expressed the sst2A receptor, we performed double-immunolabeling experiments with the calcium-binding protein PV (Fig. 2C), a well established marker of GABAergic neurons projecting to the hippocampus (Freund, 1989). Quantitative analysis revealed that of 1915 sst2A receptor-positive cells in the MS and 798 in the DB, 35.8 ± 3.1 and 12.2 ± 4%, also expressed PV immunoreactivity, respectively. Conversely, of 3035 PV-immunoreactive neurons in the MS and 1246 in the DB, 25.3 ± 4.7 and 8.0 ± 2.7% also expressed sst2A receptor immunoreactivity, respectively. We next performed triple-immunolabeling experiments to determine whether sst2A receptor/PV neurons were innervated by endogenous SST. As illustrated in the MS (Fig. 2D), confocal microscopic examination of triple-labeled sections revealed that numerous SST-immunoreactive varicosities were situated in close apposition with cell bodies and dendrites of PV/sst2A cells. Together, these results suggest that sst2A receptors are well poised to regulate the activity of both GABAergic septal interneurons and septohippocampal projecting neurons but not cholinergic neurons.
Figure 2.
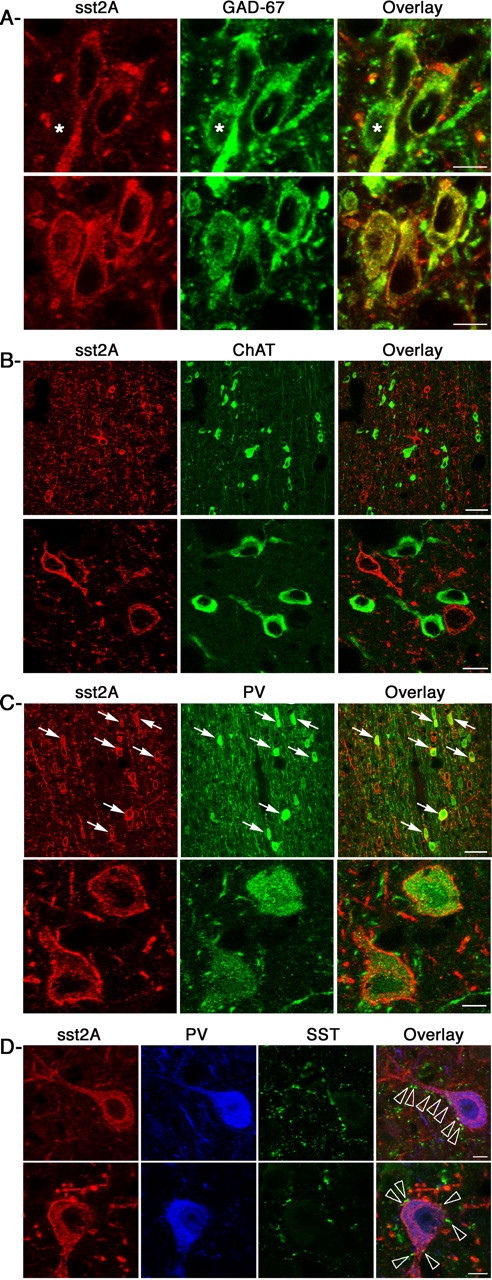
Neurochemical characterization of sst2A-expressing cells in the MS. A, Sections double labeled with the sst2A receptor (sst2A; red) and GAD67 (green) reveal that all sst2A-immunoreactive cells also display GAD67 immunoreactivity. GAD67-positive cells do not all display sst2A receptors (stars in top row). B, Low (top row) and high (bottom row) magnifications of confocal microscopic images of double-labeled sections for sst2A (red) and ChAT (green) immunoreactivities demonstrate that both markers are not expressed by the same cells. C, Sections double labeled with the sst2A receptor (red) and parvalbumin (PV; green) reveal that a subpopulation of sst2A-immunoreactive cells also expresses PV (top row, arrows), as also illustrated at a higher magnification in the bottom row. D, Confocal images of triple-labeled sections with sst2A (red), PV (blue), and somatostatin (SST; green) illustrate that numerous SST-immunoreactive axon-like terminals are located in close apposition with the somatodendritic domain of sst2A/PV neurons (overlay, arrowheads). Scale bars: A, 10 μm; B, 40 μm (top row), 8 μm (bottom row); C, 40 μm (top row), 4 μm (bottom row); D, 8 μm.
Recorded neurons express the sst2A receptor
Thirteen neurons with high average discharge rate (30.2 ± 4 spikes/s) and short spike duration (0.40 ± 0.02 ms) were labeled with neurobiotin. They were located in the midline part of MS-DB. Seven of these 13 neurons were PV positive. Four of the seven PV-positive neurons (three RB and one non-RB; 26.0 ± 8 spikes/s; spike duration, 0.39 ± 0.01 ms) also expressed sst2A receptors (Fig. 3). Of the six neurons labeled with neurobiotin but PV negative, three were found to express GAD67 immunoreactivity (three RB; 30 ± 7 spikes/s; spike duration, 0.40 ± 0.01 ms). Of these three GAD67 neurons, two expressed the sst2A receptors (Fig. 4). Thus, of 10 neurons expressing PV or GAD67, six displayed sst2A receptor immunoreactivity and discharge profiles characteristic of GABAergic cells (Brazhnik and Fox, 1999; Borhegyi et al., 2004). These experiments confirmed that MS-DB neurons expressing sst2A receptors are GABAergic. Three of the 13 labeled neurons were not found PV or GAD67 positive, although they displayed high discharge rate, brief spike, and sustained RB activity. This might be attributed to weak penetration of the antibodies in neurobiotin-filled cells (Morris et al., 1999).
Figure 3.
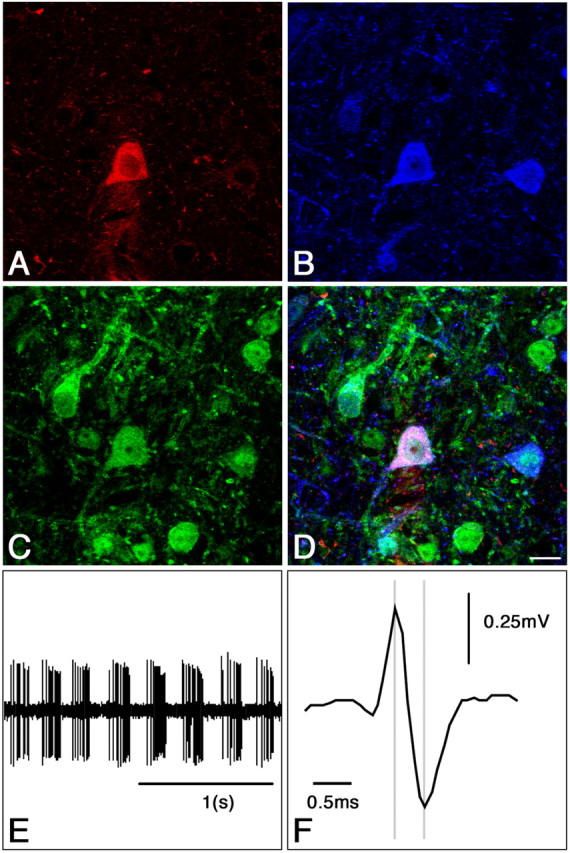
Immunohistochemical identification and electrophysiological characterization of a single juxtacellular labeled PV/sst2A neuron in the rat MS-DB. A, Single confocal section showing a recorded neuron labeled with neurobiotin (red). This neuron was also immunoreactive for parvalbumin (B, blue) and sst2A receptors (C, green) as illustrated in D. E, Extracellular recording from the neurobiotin-labeled neuron shown in A-D. The discharge profile is characterized by a theta-related bursting activity (4.0 Hz) and a high discharge rate (45 spike/s). F, Expanded scale from E indicating the short duration of the spike (0.38 ms). Scale bar: (in D) A-D, 10 μm.
Figure 4.
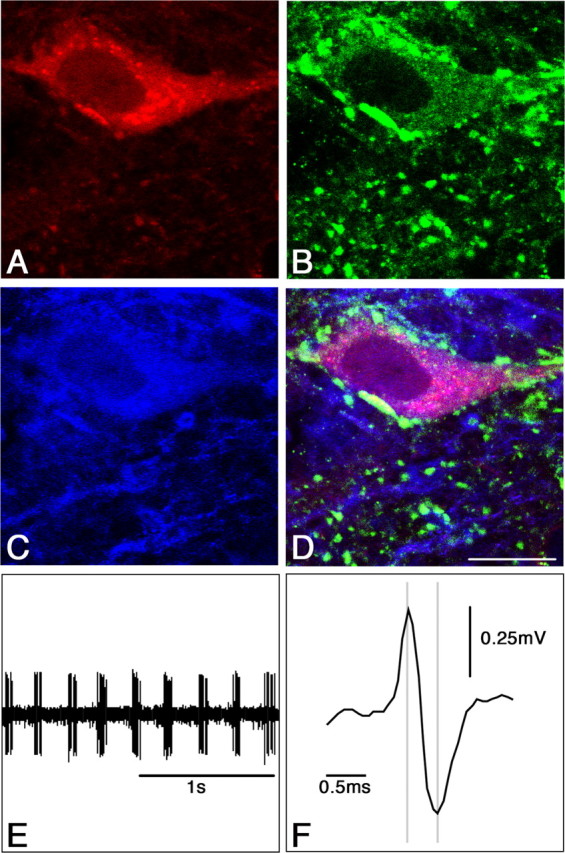
Immunohistochemical identification and electrophysiological characterization of a single juxtacellular labeled GAD67/sst2A neuron in the rat MS-DB. A, Single confocal section showing a recorded neuron labeled with neurobiotin (red). This neuron was also immunoreactive for GAD67 (B, green) and sst2A receptors (C, blue) as illustrated in D. E, Extracellular recording from the neurobiotin-labeled neuron shown in A-D. The discharge profile is characterized by a theta-related bursting activity (4.5 Hz) and a high discharge rate (24 spike/s). F, Expanded scale from E indicating the short duration of the spike (0.39 ms). Scale bar: (in D) A-D, 10 μm.
Effects of OCT on MS-DB neurons
To further analyze the role of the sst2A receptor on MS-DB GABAergic neurons, we first investigated the effects of microiontophoretic applications of the sst2A agonist OCT. The agonist was applied on a total number of 41 neurons (20 RB neurons and 21 non-RB neurons). OCT applications were selectively performed on neurons with characteristics of putative GABAergic cells (see above). To ensure that an absence of response to the drug was not caused by a space between the tips of the multibarrel micropipette and the recording electrode, only neurons responding to applications of glutamate or GABA were retained for statistical analysis. Six of 20 RB neurons (31%) responded to OCT applications by a decrease of the discharge rate (29.0 ± 3.0 vs 16.2 ± 2.0 spikes/s; mean decrease, 45.0 ± 4.0%; p < 0.01) (Fig. 5A). Additionally, the RB activity became disorganized (Fig. 5B,C). Bursts, less numerous and intermingled with isolated spikes, were significantly shorter (114 ± 10 vs 73 ± 13 ms; p < 0.01). OCT induced a marked decrease of the index of rhythmic activity (5.6 ± 0.9 before vs 2.0 ± 0.3 during application; p < 0.01). RB activity, however, was not fully abolished as shown by the residual periodic density peaks of the autocorrelation histogram (Fig. 5D). The index of rhythmic activity returned to control value (5.8 ± 0.8) with the recovery of basal activity. Five of the 21 non-RB neurons (24%) responded to OCT application by a decrease of the discharge rate (14.0 ± 2.1 vs 6.3 ± 1.8 spikes/s; mean decrease, 56.8 ± 6.8%; p < 0.01). The effect of OCT was equivalent in non-RB compared with RB cells (p = 0.17). In a number of cases, both in RB and non-RB neurons, the amplitude of the spikes increased slightly. Although this effect can result from various postsynaptic mechanisms, we can hypothesize that OCT hyperpolarized the postsynaptic membrane, as previously observed in vitro (Twery and Gallagher, 1989; Sun et al., 2002). Excitatory effects of OCT were never observed.
Figure 5.

Illustration of the effects of iontophoretic OCT application on the firing pattern of a bursting neuron in the MS-DB. A, During application of OCT (80 nA, 25 s), the discharge rate decreases by 57%. B, C, RB activity is impaired (C, expanded time scale corresponding to the stars in B). Bursts, composed of a reduced number of spikes, are less numerous. Note that the spike amplitude is increased, suggesting a hyperpolarizing effect of OCT. D, Autocorrelation histograms of representative 10 s periods of unit activity before (left), during (middle), and after (right) OCT application. Rhythmically bursting activity is considerably reduced but not completely abolished as shown by the residual peaks in the middle autocorrelation histogram.
Effects of SST on MS-DB neurons
Applications of SST were performed on a total of 30 neurons with properties of putative GABAergic cells (13 RB neurons and 17 non-RB neurons). Four of the 13 RB neurons (31%) responded to SST application by a decrease of the discharge rate (23.8 ± 8.0 vs 12.4 ± 5.0 spikes/s; mean decrease, 51.3 ± 4.0%; p < 0.05) (Fig. 6A). As observed after OCT applications, SST altered RB activity (Fig. 6C,D). Rarefied bursts were shorter (106 ± 8.0 vs 63 ± 7.0 ms; p < 0.01), and the index of rhythmic activity shifted from 4.3 ± 0.8 before to 1.2 ± 0.3 during SST application (p < 0.01). Four of the 17 non-RB neurons (23.5%) responded to SST application by a decrease of the discharge rate (13.0 ± 1.9 vs 5.2 ± 1.1 spikes/s; mean decrease, 60.5 ± 5.5%; p < 0.01) (Fig. 7). The effects of SST on RB and non-RB neurons did not differ significantly (p = 0.25). As for OCT, excitatory effects of SST were never observed. Comparison of the effects of OCT and SST on average discharge rate of RB and non-RB neurons did not show significant differences (p = 0.35 and p = 0.69, respectively). Effects of OCT and SST on rhythmic activity index were not statistically different (p = 0.58).
Figure 6.

Illustration of the effects of iontophoretic SST application on the firing pattern of a bursting neuron in the MS-DB. A, During application of SST (80 nA, 20 s), the discharge rate decreases by 70%. B, C, RB activity is impaired (C, expanded timescale corresponding to the stars in B). Bursts, composed of a reduced number of spikes, are less numerous. D, Autocorrelation histograms of representative 10 s periods of unit activity before (left), during (middle), and after (right) OCT application. Rhythmically bursting activity is considerably reduced but not completely abolished as shown by the residual peaks in the middle autocorrelation histogram.
Figure 7.

Illustration of the inhibitory effect of SST on the firing pattern of a nonbursting neuron in MS-DB. Note that the discharge rate decreases by 70% during iontophoretic application of SST (80 nA, 40 s) and recovers progressively after application.
Neurons that did not respond to OCT and/or SST (23 RB and 29 non-RB neurons) did not differ from responsive neurons in terms of discharge rate (p = 0.66 and p = 0.41, respectively) and spike duration (p = 0.44 and p = 0.48, respectively).
Effects of intraseptal injections of OCT and SST on hippocampal EEG
To investigate the physiological role of the sst2A receptor in the medial septum, the effects of microinjection of sst2A agonists in the MS-DB on hippocampal theta activity were analyzed using electroencephalogram recordings. The effects of OCT (0.5 nmol) were studied in a first group of rats (n = 5). In control conditions, the relative power in the theta band (6-9 Hz) was 70.1 ± 1.8% and was 15.7 ± 1.2% in the 1-6 Hz band and 13.3 ± 0.6% in the 9-20 Hz band. In OCT-treated rats, theta activity was altered. It lost its characteristic regularity and was intermingled with brief bouts of slow waves (Fig. 8). Ten minutes after OCT, the power in the theta band was 56.3 ± 4.0% of the total power (p < 0.05 compared with preinjection value) (Fig. 9A). The effect was maximal 20 min after injection (52.2 ± 2.0% of the total power; p < 0.001). Return to control values was observed 90 min after injection. After OCT injection, peaks of the power spectrum, although markedly reduced in terms of energy, did not change significantly in terms of frequency (7.1 ± 0.2 Hz in control vs 6.9 ± 0.2, 7.3 ± 0.2, and 7.4 ± 0.2 Hz at 10, 30, and 60 min, respectively). Conversely to the change found in the theta band, the power in the 1-6 Hz band increased to reach 28.8 ± 2.0% of the total power 20 min after injection (p < 0.01) and then gradually returned to control values. The power of hippocampal EEG in the 9-20 Hz band and in the total 1-20 Hz band was never modified by OCT. The effect of OCT was dose dependent, as shown by the larger amplitude of the changes in power values (p < 0.01) induced by the higher dose of 1 nmol (n = 4) (Fig. 9A). In companion experiments, rats received intraseptal injections of SST (0.5 nmol, n = 5; 1 nmol, n = 3). In control conditions, the relative power in the theta band was 76 ± 2.0% and was 10 ± 1.0% in the 1-6 Hz band and 14.8 ± 1.0% in the 9-20 Hz band. The effects of SST on hippocampal EEG were similar to those of OCT (Fig. 9B) (i.e., a significant but reversible decrease of the theta power). Changes induced by OCT and SST, compared at different times after injection, did not differ significantly (p = 0.4).
Figure 8.

Illustration of the effect of intraseptal OCT infusion on hippocampal theta in a freely moving rat. A, The relative power spectrum of hippocampal EEG in control condition is 10% in the 1-6 Hz band and 71% in the 6-9 Hz (theta) band. B, Twenty minutes after intraseptal infusion of OCT (0.5 nmol), the relative power increases to 43% in the 1-6 Hz band and decreases to 38% in the theta band (6-9 Hz). Note that the theta rhythm is less steady after OCT (sample in B).
Figure 9.

Time course of changes in hippocampal EEG after intraseptal OCT (A) or SST (B) infusions. Values are the mean ± SEM percentage of the power in the 1-6 and 6-9 Hz ranges relative to the total power. For both OCT and SST, note the opposite variations of the relative power between 1-6 and 6-9 Hz frequencies (top and bottom, respectively). Note the absence of effect of saline, the larger effect of the higher doses, and the progressive return to control values (*p < 0.05; **p < 0.01; ***p < 0.001).
Discussion
Recent investigations have contributed to improve our understanding of the intrinsic mechanisms underlying the bursting activity in the MS-DB (Sotty et al., 2003; Borhegyi et al., 2004; Henderson et al., 2004). However, less is known about the action of factors that might intervene to modulate this rhythmicity, as observed for example during specific behavior and arousal states (Sweeney et al., 1992; King et al., 1998). Several peptides are present in the MS-DB and might be considered potential neuromodulators of the medial septal activity. Based on the high and selective expression of the somatostatin receptor sst2A in the MS-DB, we examined the possible role of this receptor on theta-related activity in the septohippocampal pathway. The regional and cellular localization of the sst2A receptor was first defined. Double- and triple-labeling immunohistochemical experiments, using complementary cellular and neurochemical markers, demonstrated that the sst2A receptor was selectively targeted to the somatodendritic domain of a large number of neurons expressing the GABAergic marker GAD67, whereas neurons expressing the cholinergic marker ChAT did not express this receptor subtype. Furthermore, a subpopulation of GABAergic septohippocampal projecting neurons, expressing PV, also expressed the sst2A receptor and was innervated by SST. These results, together with the fact that virtually all hippocampal GABAergic neurons projecting to the MS-DB expressed SST, led us to hypothesize that SST, acting through sst2A receptors, might modulate the GABAergic neuronal activity in the MS-DB and consequently affect the theta-related rhythmicity. In agreement with our immunohistochemical data, in vivo experiments demonstrated that SST and OCT decreased the discharge rate of a subpopulation of MS-DB neurons and lessened the rhythmic activity of bursting neurons. Juxtacellular recordings and labelings showed that neurons with high discharge rate and brief spikes were immunoreactive for PV, in accordance with a recent study (Borhegyi et al., 2004), or GAD67, and that these neurons expressed the sst2A receptor. Finally, intraseptal injections of OCT and SST in freely moving rats reduced the power of hippocampal EEG in the theta band in a dose-dependent manner.
Two previous in vivo studies also reported that SST decreases neuronal activity in the rabbit lateral vestibular nucleus (Chan-Palay et al., 1982) and in the rat cortex and hippocampus (Mancillas et al., 1986). Noteworthy, these two latter regions express high levels of sst2A receptors (Dournaud et al., 1996; Schindler et al., 1997). In vitro studies have shown that SST exerts predominantly inhibitory effects through the opening of different types of K+ channels (Schweitzer et al., 1998; Sun et al., 2002). In addition, the inhibitory effect of SST might also be caused by the reduction of different types of high-voltage-activated Ca2+ currents through mechanisms involving G-proteins [Viana and Hille (1996), their references]. The in vivo extracellular approach does not allow the establishment of which K+ and Ca2+ conductances are modified in MS-DB neurons. It is, however, plausible that the activation of postsynaptic sst2A receptors may induce membrane hyperpolarization that decreases the probability of the neuron to produce an action potential. In agreement with this hypothesis, we observed that the reduction of firing level was sometimes accompanied by an increase in action potential size evocative of hyperpolarization. As observed in hippocampus, cortex (Mancillas et al., 1986), and hypothalamus (Peineau et al., 2003), the activation of sst2A receptors might also lower the discharge rate of MS-DB GABAergic neurons by reducing the postsynaptic excitatory effects of glutamate. This effect might have important functional consequences, because NMDA receptors in the MS-DB are likely to modulate the amplitude of hippocampal theta (Leung and Shen, 2004).
The rhythmic activity of many neurons in the CNS is regulated by various ionic conductances, including the hyperpolarization-activated current Ih (Luthi and McCormick, 1998). It has been demonstrated recently in vitro that septohippocampal neurons expressing GAD mRNA exhibit bursting activity and an important Ih (Sotty et al., 2003; Morris et al., 2004). Furthermore, the selective blockade of Ih current in the MS-DB decreases the spontaneous firing of GABAergic neurons in septal slices (Xu et al., 2004) and reduces the frequency of hippocampal theta in freely moving and urethane anesthetized rats (Kocsis and Li, 2004). One might hypothesize that activation of sst2A receptors impairs the RB activity of MS-DB neurons via an action on Ih current. However, this hypothesis does not agree with the facts that SST both decreases burst discharges in thalamic reticular neurons without any change in Ih current (Sun et al., 2002) and hyperpolarizes intrinsically bursting subicular neurons without any effect on inwardly rectifying currents (Greene and Mason, 1996). Additional intracellular in vitro studies are clearly needed to unravel cellular mechanisms by which SST alters the firing of both RB and non-RB MS-DB neurons.
As previously mentioned, the septohippocampal pathway consists mainly of cholinergic and GABAergic neurons that exhibit RB activity in the in vivo conditions (Brazhnik and Fox, 1997; Apartis et al., 1998). It is now well established that the hippocampal theta critically depends on the bursting activity of the GABAergic septohippocampal neurons. These neurons send rhythmic inputs to GABAergic hippocampal interneurons that, in turn, phasically disinhibit the pyramidal cells (Freund and Antal, 1988; Toth et al., 1997). GABAergic hippocamposeptal neurons, which express SST in their large majority, project to GABAergic septohippocampal neurons (Gulyas et al., 2003). This dual projection forms a loop that strengthens synchronous activity in the septohippocampal system (Toth et al., 1993). It has also been shown that rhythmic inhibitory inputs from local GABAergic neurons trigger RB activity in septal cholinergic neurons, leading a greater number of MS-DB cells to discharge rhythmically (Brazhnik and Fox, 1997). Our results show that the power of hippocampal theta rhythm is significantly reduced after intraseptal infusion of sst2A agonists. This finding is consistent with the inhibitory effect of OCT and SST on a subpopulation of rhythmic and tonic GABAergic neurons of the MS-DB. Interestingly, after SST and OCT applications, the rhythmically bursting activity of MS-DB neurons, although markedly damped, is still distinguishable with a preserved frequency. This suggests that, in contrast to their effects on mechanisms that sustain discharge rate, SST and OCT do not markedly alter those that sustain oscillatory activity. Considering the key role of GABAergic neurons in generating hippocampal theta, one might assume that a decrease of RB activity, even in a limited number of them, has an extensive repercussion in the septal network and induces a powerful reduction of hippocampal theta activity. In addition, our immunocytochemical and electrophysiological results suggest that the sst2A receptor activation might also influence theta rhythm through inhibition of nonbursting septohippocampal neurons, a population of GABAergic neurons that could also control the excitability of hippocampal interneurons (Knapp et al., 2000). Last, the sst2A receptor might influence theta activity by reducing the disinhibitory role of GABAergic interneurons involved in the control of GABAergic and cholinergic MS-DB networks. Based on our results, it is not possible to know whether endogenously released SST contributes to the generation of theta activity. This study, however, strongly suggests that released SST, acting on different types of MS-DB GABAergic neurons through postsynaptic sst2A receptors, represents an important modulatory factor in the control of theta activity. Such a physiological role for a neuropeptide receptor strengthens the importance of the neuropeptidergic transmission in the mammalian brain. Dissecting intracellular pathways recruited by the sst2A receptor in the inhibition of the MS-DB GABAergic neuronal activity will represent an intriguing issue for future in vitro studies.
Footnotes
This work was supported by the Institut National de la Santé et de la Recherche Médicale and European Union Grant QLG-1999-0098. Z.C. was a recipient of a fellowship from the Ligue Française Contre l'Epilepsie. We are grateful to L. Helboe for the gift of the antiserum against the sst2A receptor.
Correspondence should be addressed to Dr. Pascal Dournaud, Institut National de la Santé et de la Recherche Médicale, U549, Centre Paul Broca, 2 ter rue d'Alésia, 75014 Paris, France. E-mail: dournaud@broca.inserm.fr.
Z. Csaba's present address: Neuroendocrine Research Laboratory, Hungarian Academy of Sciences and Department of Human Morphology and Developmental Biology, Semmelweis University, H-1094 Budapest, Hungary.
Copyright © 2005 Society for Neuroscience 0270-6474/05/252032-10$15.00/0
References
- Alreja M (1996) Excitatory actions of serotonin on GABAergic neurons of the medial septum and diagonal band of Broca. Synapse 22: 15-27. [DOI] [PubMed] [Google Scholar]
- Alreja M, Liu W (1996) Noradrenaline induces IPSCs in rat medial septal/diagonal band neurons: involvement of septohippocampal GABAergic neurons. J Physiol (Lond) 494: 201-215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alreja M, Shanabrough M, Liu W, Leranth C (2000) Opioids suppress IPSCs in neurons of the rat medial septum/diagonal band of Broca: involvement of μ-opioid receptors and septohippocampal GABAergic neurons. J Neurosci 20: 1179-1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Apartis E, Poindessous-Jazat FR, Lamour YA, Bassant MH (1998) Loss of rhythmically bursting neurons in rat medial septum following selective lesion of septohippocampal cholinergic system. J Neurophysiol 79: 1633-1642. [DOI] [PubMed] [Google Scholar]
- Armstrong JN, MacVicar BA (2001) Theta-frequency facilitation of AMPA receptor-mediated synaptic currents in the principal cells of the medial septum. J Neurophysiol 85: 1709-1718. [DOI] [PubMed] [Google Scholar]
- Borhegyi Z, Varga V, Szilagyi N, Fabo D, Freund TF (2004) Phase segregation of medial septal GABAergic neurons during hippocampal theta activity. J Neurosci 24: 8470-8479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brazhnik ES, Fox SE (1997) Intracellular recordings from medial septal neurons during hippocampal theta rhythm. Exp Brain Res 114: 442-453. [DOI] [PubMed] [Google Scholar]
- Brazhnik ES, Fox SE (1999) Action potentials and relations to the theta rhythm of medial septal neurons in vivo Exp Brain Res 127: 244-258. [DOI] [PubMed] [Google Scholar]
- Buzsaki G (2002) Theta oscillations in the hippocampus. Neuron 33: 325-340. [DOI] [PubMed] [Google Scholar]
- Chan-Palay V, Ito M, Tongroach P, Sakurai M, Palay S (1982) Inhibitory effects of motilin, somatostatin, [Leu]enkephalin, [Met]enkephalin, and taurine on neurons of the lateral vestibular nucleus: interactions with gamma-aminobutyric acid. Proc Natl Acad Sci USA 79: 3355-3359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csaba Z, Bernard V, Helboe L, Bluet-Pajot MT, Bloch B, Epelbaum J, Dournaud P (2001) In vivo internalization of the somatostatin sst2A receptor in rat brain: evidence for translocation of cell-surface receptors into the endosomal recycling pathway. Mol Cell Neurosci 17: 646-661. [DOI] [PubMed] [Google Scholar]
- Csaba Z, Simon A, Helboe L, Epelbaum J, Dournaud P (2002) Neurochemical characterization of receptor-expressing cell populations by in vivo agonist-induced internalization: insights from the somatostatin sst2A receptor. J Comp Neurol 454: 192-199. [DOI] [PubMed] [Google Scholar]
- Csaba Z, Simon A, Helboe L, Epelbaum J, Dournaud P (2003) Targeting sst2A receptor-expressing cells in the rat hypothalamus through in vivo agonist stimulation: neuroanatomical evidence for a major role of this subtype in mediating somatostatin functions. Endocrinology 144: 1564-1573. [DOI] [PubMed] [Google Scholar]
- Dournaud P, Gu YZ, Schonbrunn A, Mazella J, Tannenbaum GS, Beaudet A (1996) Localization of the somatostatin receptor SST2A in rat brain using a specific anti-peptide antibody. J Neurosci 16: 4468-4478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dournaud P, Boudin H, Schonbrunn A, Tannenbaum GS, Beaudet A (1998) Interrelationships between somatostatin sst2A receptors and somatostatin-containing axons in rat brain: evidence for regulation of cell surface receptors by endogenous somatostatin. J Neurosci 18: 1056-1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dournaud P, Slama A, Beaudet A, Epelbaum A (2000) Somatostatin receptors. In: Handbook of chemical neuroanatomy, Vol 16, Peptide receptors, Part I (Quirion R, Björklund A, Hökfelt T, eds), pp 1-43. Amsterdam: Elsevier.
- Freund TF (1989) GABAergic septohippocampal neurons contain parvalbumin. Brain Res 478: 375-381. [DOI] [PubMed] [Google Scholar]
- Freund TF, Antal M (1988) GABA-containing neurons in the septum control inhibitory interneurons in the hippocampus. Nature 336: 170-173. [DOI] [PubMed] [Google Scholar]
- Frotscher M, Leranth C (1985) Cholinergic innervation of the rat hippocampus as revealed by choline acetyltransferase immunocytochemistry: a combined light and electron microscopic study. J Comp Neurol 239: 237-246. [DOI] [PubMed] [Google Scholar]
- Greene JR, Mason A (1996) Neuronal diversity in the subiculum: correlations with the effects of somatostatin on intrinsic properties and on GABA-mediated IPSPs in vitro J Neurophysiol 76: 1657-1666. [DOI] [PubMed] [Google Scholar]
- Gulyas AI, Hajos N, Katona I, Freund TF (2003) Interneurons are the local targets of hippocampal inhibitory cells which project to the medial septum. Eur J Neurosci 17: 1861-1872. [DOI] [PubMed] [Google Scholar]
- Handel M, Schulz S, Stanarius A, Schreff M, Erdtmann-Vourliotis M, Schmidt H, Wolf G, Hollt V (1999) Selective targeting of somatostatin receptor 3 to neuronal cilia. Neuroscience 89: 909-926. [DOI] [PubMed] [Google Scholar]
- Helboe L, Moller M (1999) Immunohistochemical localization of somatostatin receptor subtypes sst1 and sst2 in the rat retina. Invest Ophthalmol Vis Sci 40: 2376-2382. [PubMed] [Google Scholar]
- Helboe L, Moller M, Norregaard L, Schiodt M, Stidsen CE (1997) Development of selective antibodies against the human somatostatin receptor subtypes sst1-sst5. Brain Res Mol Brain Res 49: 82-88. [DOI] [PubMed] [Google Scholar]
- Helboe L, Stidsen CE, Moller M (1998) Immunohistochemical and cytochemical localization of the somatostatin receptor subtype sst1 in the somatostatinergic parvocellular neuronal system of the rat hypothalamus. J Neurosci 18: 4938-4945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henderson Z, Fiddler G, Saha S, Boros A, Halasy K (2004) A parvalbumin-containing, axosomatic synaptic network in the rat medial septum: relevance to rhythmogenesis. Eur J Neurosci 19: 2753-2768. [DOI] [PubMed] [Google Scholar]
- Johansson O, Hokfelt T, Elde RP (1984) Immunohistochemical distribution of somatostatin-like immunoreactivity in the central nervous system of the adult rat. Neuroscience 13: 265-339. [DOI] [PubMed] [Google Scholar]
- King C, Recce M, O'Keefe J (1998) The rhythmicity of cells of the medial septum/diagonal band of Broca in the awake freely moving rat: relationships with behaviour and hippocampal theta. Eur J Neurosci 10: 464-477. [DOI] [PubMed] [Google Scholar]
- Knapp JA, Morris NP, Henderson Z, Matthews RT (2000) Electrophysiological characteristics of non-bursting, glutamate decarboxylase messenger RNA-positive neurons of the medial septum/diagonal band nuclei of guinea-pig and rat. Neuroscience 98: 661-668. [DOI] [PubMed] [Google Scholar]
- Kocsis B, Li S (2004) In vivo contribution of h-channels in the septal pacemaker to theta rhythm generation. Eur J Neurosci 20: 2149-2158. [DOI] [PubMed] [Google Scholar]
- Kohler C, Chan-Palay V, Wu JY (1984) Septal neurons containing glutamic acid decarboxylase immunoreactivity project to the hippocampal region in the rat brain. Anat Embryol (Berl) 169: 41-44. [DOI] [PubMed] [Google Scholar]
- Lee MG, Chrobak JJ, Sik A, Wiley RG, Buzsaki G (1994) Hippocampal theta activity following selective lesion of the septal cholinergic system. Neuroscience 62: 1033-1047. [DOI] [PubMed] [Google Scholar]
- Leung LS, Shen B (2004) Glutamatergic synaptic transmission participates in generating the hippocampal EEG. Hippocampus 14: 510-525. [DOI] [PubMed] [Google Scholar]
- Luthi A, McCormick DA (1998) H-current: properties of a neuronal and network pacemaker. Neuron 21: 9-12. [DOI] [PubMed] [Google Scholar]
- Mancillas JR, Siggins GR, Bloom FE (1986) Somatostatin selectively enhances acetylcholine-induced excitations in rat hippocampus and cortex. Proc Natl Acad Sci USA 83: 7518-7521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews RT (1999) Neurotensin depolarizes cholinergic and a subset of non-cholinergic septal/diagonal band neurons by stimulating neurotensin-1 receptors. Neuroscience 94: 775-783. [DOI] [PubMed] [Google Scholar]
- Matthews RT, Lee WL (1991) A comparison of extracellular and intracellular recordings from medial septum/diagonal band neurons in vitro Neuroscience 42: 451-462. [DOI] [PubMed] [Google Scholar]
- Morris NP, Harris SJ, Henderson Z (1999) Parvalbumin-immunoreactive, fast-spiking neurons in the medial septum/diagonal band complex of the rat: intracellular recordings in vitro Neuroscience 92: 589-600. [DOI] [PubMed] [Google Scholar]
- Morris NP, Fyffe RE, Robertson B (2004) Characterisation of hyperpolarization-activated currents (I(h)) in the medial septum/diagonal band complex in the mouse. Brain Res 1006: 74-86. [DOI] [PubMed] [Google Scholar]
- Paxinos G, Watson C (1986) The rat brain in stereotaxic coordinates atlas. Sydney: Academic.
- Peineau S, Potier B, Petit F, Dournaud P, Epelbaum J, Gardette R (2003) AMPA-sst2 somatostatin receptor interaction in rat hypothalamus requires activation of NMDA and/or metabotropic glutamate receptors and depends on intracellular calcium. J Physiol (Lond) 546: 101-117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkel DH, Gerstein GL, Moore GP (1967) Neuronal spike trains and stochastic point processes. I. The single spike train. Biophys J 7: 391-418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinault D (1996) A novel single-cell staining procedure performed in vivo under electrophysiological control: morpho-functional features of juxtacellularly labeled thalamic cells and other central neurons with biocytin or Neurobiotin. J Neurosci Methods 65: 113-136. [DOI] [PubMed] [Google Scholar]
- Schindler M, Sellers LA, Humphrey PP, Emson PC (1997) Immunohistochemical localization of the somatostatin SST2(A) receptor in the rat brain and spinal cord. Neuroscience 76: 225-240. [DOI] [PubMed] [Google Scholar]
- Schindler M, Humphrey PP, Lohrke S, Friauf E (1999) Immunohistochemical localization of the somatostatin sst2(b) receptor splice variant in the rat central nervous system. Neuroscience 90: 859-874. [DOI] [PubMed] [Google Scholar]
- Schreff M, Schulz S, Handel M, Keilhoff G, Braun H, Pereira G, Klutzny M, Schmidt H, Wolf G, Hollt V (2000) Distribution, targeting, and internalization of the sst4 somatostatin receptor in rat brain. J Neurosci 20: 3785-3797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schweitzer P, Madamba SG, Siggins GR (1998) Somatostatin increases a voltage-insensitive K+ conductance in rat CA1 hippocampal neurons. J Neurophysiol 79: 1230-1238. [DOI] [PubMed] [Google Scholar]
- Sotty F, Danik M, Manseau F, Laplante F, Quirion R, Williams S (2003) Distinct electrophysiological properties of glutamatergic, cholinergic and GABAergic rat septohippocampal neurons: novel implications for hippocampal rhythmicity. J Physiol (Lond) 551: 927-943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stone TW (1985) Microiontophoresis and pressure ejection. In: IBRO handbook series: methods in neurosciences, Vol 8 (Smith AD, ed), p 214. New York: Wiley. [Google Scholar]
- Stroh T, Kreienkamp HJ, Beaudet A (1999) Immunohistochemical distribution of the somatostatin receptor subtype 5 in the adult rat brain: predominant expression in the basal forebrain. J Comp Neurol 412: 69-82. [DOI] [PubMed] [Google Scholar]
- Sun QQ, Huguenard JR, Prince DA (2002) Somatostatin inhibits thalamic network oscillations in vitro: actions on the GABAergic neurons of the reticular nucleus. J Neurosci 22: 5374-5386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweeney JE, Lamour Y, Bassant MH (1992) Arousal-dependent properties of medial septal neurons in the unanesthetized rat. Neuroscience 48: 353-362. [DOI] [PubMed] [Google Scholar]
- Toth K, Borhegyi Z, Freund TF (1993) Postsynaptic targets of GABAergic hippocampal neurons in the medial septum-diagonal band of broca complex. J Neurosci 13: 3712-3724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toth K, Freund TF, Miles R (1997) Disinhibition of rat hippocampal pyramidal cells by GABAergic afferents from the septum. J Physiol (Lond) 500: 463-474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Twery MJ, Gallagher JP (1989) Somatostatin hyperpolarizes neurons and inhibits spontaneous activity in the rat dorsolateral septal nucleus. Brain Res 497: 315-324. [DOI] [PubMed] [Google Scholar]
- Viana F, Hille B (1996) Modulation of high voltage-activated calcium channels by somatostatin in acutely isolated rat amygdaloid neurons. J Neurosci 16: 6000-6011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu M, Zaborszky L, Hajszan T, van den Pol AN, Alreja M (2004) Hypocretin/orexin innervation and excitation of identified septohippocampal cholinergic neurons. J Neurosci 24: 3527-3536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C, Datta S, Wu M, Alreja M (2004) Hippocampal theta rhythm is reduced by suppression of the H-current in septohippocampal GABAergic neurons. Eur J Neurosci 19: 2299-2309. [DOI] [PubMed] [Google Scholar]