

Abstract
Bevacizumab, a monoclonal antibody targeting vascular endothelial growth factor A (VEGF-A), was used in combination with traditional chemotherapy as the first line treatment for metastatic colorectal cancer (mCRC), non-small cell lung cancer (NSCLC) and advanced ovarian cancer. However, it shows limited efficacy for human triple-negative breast cancer (TNBC). Bevacizumab shows potent anti-angiogenesis activity, meanwhile, it also increases invasive and metastatic properties of TNBC cells by activiting Wnt/β-Catenin pathway. To overcome this problem, and fully utilize its potency against cancer, further synergistic strategy is recommended to be developed, especially the concurrent use with those Wnt-targeting agents. Here, by screening a small library of traditional Chinese medicine, we identified a Chinese herb derived Oxymatrine, which could target Wnt/β-Catenin signaling and compromise the oncogenic effects of Bevacizumab. Bevacizumab was validated to induce epithelial-mesenchymal cell transformation (EMT) and cancer stem-like properties of TNBC cells in hypoxia/nutritional stress environment. On the contrary, Oxymatrine reversed the EMT phenotype and depleted the subpopulation of TNBC stem cells induced by Bevacizumab. Oxymatrine enhanced the anti-tumor effects of Bevacizumab in vivo, and holded the potential of reducing the risk of relapse and metastasis by impairing the self-renewal ability of TNBC stem cells. The underlying mechanism was elucidated: Bevacizumab stimulated Wnt/β-Catenin signaling pathway, and Oxymatrine could compromise this effect. On this foundation, factoring into the satisfactory anti-angiogenic activity and low toxicity, Oxymatrine is a good candidate for the synergistic therapy together with Bevacizumab for the treatment of TNBC.
Keywords: Oxymatrine, bevacizumab, TNBC, EMT, Wnt/β-Catenin pathway
Introduction
TNBC, characterized by the lack of expression of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2), is an aggressive subtype of breast cancer. Breast cancer incidence rate ranks the first in overall cancer in women, among them, TNBC accounts for around 10-20% [1,2]. Currently TNBC still represents the most therapeutically intractable subtype as the negative expressions of ER, PR, and HER2, and cytotoxic chemotherapy remains the standard of care for patients with metastatic TNBC. However, TNBC is clinically characterized as more aggressive and less responsive to standard treatment with a poorer overall patient prognosis [3-5].
Bevacizumab is a recombinant humanized monoclonal antibody that blocks angiogenesis by inhibiting VEGF-A approved by FDA in 2004. To date, FDA has approved its application for the treatment of mCRC, NSCLC and advanced ovarian cancer combined with the chemotherapy drugs. Meanwhile, it has also been tested by combination with the cytotoxic chemotherapy in patients with TNBC (clinical trial number NCT00528567). The result from the clinical trial showed that this combination regime increases objective response rate (ORR) and progression-free survival (PFS) vs. chemotherapy alone, however the benefit of overall survival (OS) is not enough to support its further application for this indication [6-8].
Bevacizumab, as well as some other anti-angiogenic agents, has been recently shown to be able to increase the invasive and metastatic capacity and induce EMT in breast cancer cells [9]. Bevacizumab activates Wnt/β-Catenin signaling pathway excessively, further facilitates the growth, migration and invasion of TNBC, which normally associates with the unfavorable outcome of patients [10-13]. In addition, the important angiogenesis inducing factor VEGF, a target gene of β-Catenin, is up-regulated by the activated Wnt/β-Catenin signaling, possibly leading to the incomplete blocking of tumor angiogenesis [14,15]. So it seems that the therapeutic effects of Bevacizumab against TNBC may be partially counteracted by the induced activation of Wnt/β-Catenin signaling, indicating that, to improve TNBC patient outcome, anti-angiogenic agents need to be combined with Wnt/β-Catenin signaling-targeting drugs.
TCM has a long history with various pharmacological activities and low toxicity. Thereby, those Chinese herbs hold great potential for the therapy development against cancer, and several agents like 20(S)-ginsenoside Rg3, Lupeol and Sophoridine has entered into market [16,17]. We have carried out some work about the development of Chinese herb-derived triptolide [18,19], and established a small Chinese herb library focusing on cancer therapy. Oxymatrine (C15H24N2O2), one compound in our library, is an alkaloid compound extracted from the roots of Sophora flavescens Ait. It attracts our interest as it also shows multiple biological activities like triptlide, but it is well tolerable. Oxymatrine is clinically demonstrated to display anti-inflammatory, anti-allergic, anti-virus, antifibrotic, and cardiovascular protective effects [20,21]. Recently, extensive experimental studies on Oxymatrine have demonstrated its potential applications to the treatment of various cancer [22-24], and showed that Oxymatrine can effectively repress Wnt/β-Catenin pathway in some tumors [25,26]. The potential anti-tumor activities of Oxymatrine against TNBC were verified in our preliminary study (Supplementary Figure 1).
Herein, the double-edged role of Bevacizumab for TNBC therapy deserves attention, and may be overcame by synergistic therapy. Chinese herbs are promising candidates for such design. We firstly verified the EMT and CSC phenotype caused by Bevacizumab with some routine assays like wound healing, transwell invasion, and ALDEFLUOR test. Then Oxymatrine was selected from our established Chinese herb library for further evaluation as it demonstrated good synergistic effects with Bevacizumab. In the end, the in vivo effects were also evaluated, and Wnt/β-Catenin signaling pathway were well dissected as the possible mechanism for such synergistic effects.
Materials and methods
Cell culture and reagents
Oxymatrine (purity ≥ 98%, Macklin Biochemical Co., Ltd., Shanghai, China) and Bevacizumab (Roche, Paris, France) was diluted in PBS. Docetaxel (Hengrui Medicine, Jiangsu, China) was suspended in the injection solvent and then diluted in PBS. FH535 (≥ 98%, absin, China) was suspended in DMSO. The human embryonic kidney cell line HEK293 (Cell bank of the Chinese Academy of Sciences, Shanghai, China) was cultured in DMEM high glucose medium (Gibco, New York, USA), supplemented with 10% (v/v) FBS (Gibco, Auckland, NZ). The human breast cancer cell lines MDA-MB-231/MDA-MB-468 (American Type Culture Collection, New York, USA) were maintained in L-15 medium (Gibco, New York, USA) supplemented with 10% (v/v) FBS. HUVECs (ScienCell, San Diego, CA) were maintained in endothelial culture medium (ECM, ScienCell) supplemented with 5% (v/v) FBS and 1% (v/v) endothelial cell growth supplement (ECGS, ScienCell). All cells were maintained at 37°C in a humidified atmosphere with 5% CO2.
Transwell invasion assay
1 × 104 of MDA-MB-231/MDA-MB-468 cells were suspended in serum-free medium with the addition of Oxymatrine (4 mM)/Bevacizumab (200 nM)/Oxymatrine (4 mM) + Bevacizumab (200 nM) and then plated into the upper wells of 24-well transwell chambers (Millipore, Billerica, USA) coated with 20 μL Matrigel (Corning, Bedford, USA). The lower chambers were filled with L-15 medium containing 10% (v/v) FBS. Similarly, 1 × 104 HUVECs and different concentrations of Oxymatrine (0, 1, 2 and 4 mM) were suspended in serum-free medium and added to the top 24-well transwell chambers. The bottom chambers were filled with 600 μL ECM containing 5% (v/v) FBS and 1% (v/v) ECGS. The next experimental procedure was performed as previously described [27].
Wound healing assay
3 × 104 MDA-MB-231/MDA-MB-468 cells were placed into the each well of Culture-Insert (Ibidi, Martinsried, Germany) in 24-well plate. After adherence, the Culture-Insert was removed and cells were rinsed with PBS. Oxymatrine/Bevacizumab/Oxymatrine + Bevacizumab dissolved in serum-free medium was added into the wells. After 24 h-incubation, the drug-dissolved medium was replaced by the serumfree medium. Images were taken with OLYMPUS inversion fluorescence microscope at 0, 24, 48 h after the addition of the treatments. The wound migrated distances were measured with Image-Pro-Plus program and calculated as follows: Ln = (L0-Ltime)/2.
ALDEFLUOR assay
ALDEFLUOR™ kit (STEMCELL Technologies, Canada) was used to analyze the level of the enzyme aldehyde dehydrogenase (ALDH) in cells. 5 × 105 MDA-MB-231/MDA-MB-468 cells were seeded in 6-well plates. When the confluence reached 70%-80%, cells were incubated with Oxymatrine/Bevacizumab/Oxymatrine + Bevacizumab for 24 h. Whereafter, cells were trypsinized, resuspended in ALDEFLUOR™ assay buffer and incubated with the activated ALDEFLUOR™ fluorescent reagent for 30 min at 37°C. For ALDH1 inhibitor control, diethylaminobenzaldehyde (DEAB) was added along with the fluorescent reagent. Cells were then stained with 7-AAD (BD Biosciences, San Diego, USA), green fluorescence positive cells in live cells (7-AAD) were analyzed using a BD FACSMelody flow cytometer by comparing the fluorescence intensity of DEAB-treated samples.
IF analysis
5 × 105 MDA-MB-231/MDA-MB-468 cells were seeded on cover slips in 6-well plates and allowed to adhere to the glass overnight to reach 70% confluence. Then, the cells were treated with Oxymatrine/Bevacizumab/Oxymatrine + Bevacizumab/FH535 for 24 h. Subsquently, the subcellar location of β-Catenin was detected as previously described [28]. Finally, images were taken by an Leica SP2 Confocal System at 400-times magnification.
Immunoblotting assay
5 × 105 MDA-MB-231/MDA-MB-468 cells were seeded into the wells of 6-well plates, allowed to adhere overnight to reach 70%-80% confluence, treated with Oxymatrine/Bevacizumab/Oxymatrine + Bevacizumab/FH535 (20 μM, a small molecule inhibitor of Wnt/β-Catenin pathway) for 24 h, then the whole cell protein was extracted from cells using RIPA buffer (Beyotime, Shanghai, China) and the nuclear extracts were prepared with a NE-PER Nuclear and Cytoplasmic Extraction Kit (Pierce Biotechnology, Rockford, USA). The next experimental procedure was performed as previously described [28]. The primary antibodies used included anti-β-actin, anti-Histone H3, anti-β-Catenin, anti-c-Myc, anti-Snail, anti-Slug, anti-Vimentin, anti-TCF8/ZEB1, anti-CD44, anti-E-cadherin, anti-N-cadherin (CST, MA, USA), anti-Cyclin D1, anti-VEGFA (Abcam, Cambridge, UK).
Cell xenografts in nude mice assay
5 to 6-week-old female BALB/c-nude mice were purchased from Shanghai Lab. Animal Research Center, China, maintained in a pathogen-free environment (23 ± 2°C, 55 ± 5% humidity) on a 12 h light/12 h dark cycle with food and water supplied adlibitum throughout the experimental period. All animal experiments were performed in accordance with the National Institutes of Health guide for the care and use of Laboratory animals and protocols approved by the Animal Ethics Committee of Shanghai University of Medicine and Health Sciences. Each mouse was subcutaneously inoculated with injections of 1 × 107 MDA-MB-231/MDA-MB-468 cells into its left axilla. When the tumors reached 50 mm3, mice were randomized into 8 groups (n = 5 for each treatment group), and the administration began: (1) PBS control; (2) 5 mg/kg Oxymatrine (intraperitoneal injection, once every other day); (3) 25 mg/kg Oxymatrine; (4) 25 mg/kg FH535 (intraperitoneal injection, once every other day); (5) 5 mg/kg Bevacizumab (intravenous injection, every 3 days); (6) 5 mg/kg Bevacizumab + 25 mg/kg Oxymatrine; (7) 5 mg/kg Bevacizumab + 25 mg/kg FH535; (8) 5 mg/kg Bevacizumab + 10 mg/kg Docetaxel (intravenous injection, every 3 days). Tumor development was measured periodically and the tumor volume was determined using the formula V = (length × width2)/2. The administration end at day 31 and the tumors of each group were isolated for further study.
IHC and IF analysis
Paraffin sections were cut into 5 μm sections and fixed in 4% paraformaldehyde. For IHC staining, the sections were incubated with primary antibodies anti-Ki-67 (CST, MA, USA), anti-VEGFA antibody (Abcam, Cambridge, UK), and horseradish peroxidase (HRP)-labeled secondary antibody and analyzed with the Vectastain ABC Kit (Dako, Copenhagen, Denmark). A subset of MDA-MB-231/MDA-MB-468 tumor tissues were subjected to Western blot to examine the VEGFA protein levels and show changes in VEGFA expression. For IF staining, the sections were incubated with primary antibodies anti-β-Catenin, anti-CD31 (CST, MA, USA) and Alexafluor-488/594 labeled antibody (Molecular Probes, Eugene, USA). The slides were sealed with Prolong Gold Antifade (Invitrogen, Carlsbad, USA), which contained DAPI. Subsquently, images were taken by an Leica SP2 Confocal System at 400-times magnification.
Sphere formation assay
MDA-MB-231/MDA-MB-468 tumors tissues were minced into small pieces and processed with collagenase for 2 h at 37°C. After centrifugation, the cell pellets were trypsinized and passed through an 80-μm filter to produce single-cell suspension. Then 5 × 103 dissociated tumor cells were seeded on ultralow attachment 6-well plates in serum-free medium DMEM/F12 (Gibco, Grand Island, USA) supplemented with B27 (Gibco, Grand Island, USA), 20 ng/mL human recombinant basic fibroblast growth factor (FGF)-basic, and 20 ng/mL epidermal growth factor (EGF, Sino Biological Inc., Beijing, China). The medium was changed every other day until the tumor sphere formation was observed in 2 weeks, and the mammospheres with diameter > 60 μm were counted under an OLYMPUS inversion fluorescence microscope [29].
Cell proliferation assay
HUVECs/HEK293 cells (negative control) were seeded at a density of 2000 cells/well in 96-well plates. After cultured overnight, fresh medium containing different concentrations of Oxymatrine were replenished for 48 h. Subsequently, cell viability was quantified by MTT assay and the proliferation rate was expressed as percentages of the vehicle control (Oxymatrine untreated group).
Tube formation assay
2 × 104 HUVECs were seeded into the 96-well plate coated with 50 μL Matrigel and added with 100 μL ECM supplemented with 2% (v/v) FBS, 1% (v/v) ECGS and various concentrations of Oxymatrine (0, 1, 4 and 16 mM). The next experimental procedure was performed as previously described [27].
Statistical analysis
All data were presented as the mean ± standard deviation (SD). Differences between multiple groups were analyzed by the student’s t test and p values of 0.05 or less were considered statistically significant. All figures were produced with the GraphPad Prism 5 software program.
Results
Bevacizumab induced transwell, migration of TNBC cells and increased subpopulation of putative TNBC stem-like cells under serum starvation condition
Wound healing and transwell invasion assays were performed for the evaluation of cell migration. Following Bevacizumab treatments under serum starvation, increased cell invasion (Figure 1A, 1B) and migration (Figure 1C, 1E) were detected. As increased invasion and migration are outcomes of the EMT process [30,31], which indicated Bevacizumab could induce EMT of TNBC cells in serum-deprived environment. Moreover, researchers have shown that EMT could promote stem cell properties and further generate cells with the features of CSC [32,33], conversely, CSC could accelerate tumor invasion and metastasis. In consideration of the interplay of EMT and CSC, we utilized ALDEFLUOR assay to detect the subpopulation of TNBC stem-like cells [34]. Previously, intracellular ALDH activity was identified to be a functional marker for CSCs derived from different cancer types [35-37]. As shown in Figure 1D, 1F, in comparison with the control cells, the Bevacizumab-treated TNBC cells possessed higher ALDH1+ population notably (MDA-MB-231: **P < 0.01, MDA-MB-468: ##P < 0.01), which indicated that Bevacizumab increased the subpopulation of TNBC stem-like cells remarkably under serum starvation condition.
Figure 1.

Bevacizumab promoted the invasion, migration of TNBC cells, and increased percent of ALDH1+ TNBC cells. (A) Microscopic views from transwell assay to estimate MDA-MB-231/MDA-MB-468 cells invasion following 24 h treatment of Bevacizumab (200 nM), bar = 100 μm. (B) Quantitative analysis of (A) by Image J. (C) Photomicrographs of cell migration from wound healing assay in MDA-MB-231/MDA-MB-468 cells treated with Bevacizumab, bar = 100 μm. (D) MDA-MB-231/MDA-MB-468 cells were treated with Bevacizumab for 24 h, then ALDH1+ cells were determined by Alderfluor assay and viable cells (7-AAD negative) were used for analysis. DEAB-treated cells were used as negative control. (E) Quantitative analysis of (C) by Image J. (F) Quantitative analysis of (D). Data were presented as the mean ± SD, n = 3, *P < 0.05, **P < 0.01 (MDA-MB-231 cells); #P < 0.05, ##P < 0.01 (MDA-MB-468 cells).
Oxymatrine suppressed Bevacizumab-induced transwell, migration of TNBC cells and depleted subpopulation of putative TNBC stem-like cells induced by Bevacizumab
Figure 2 demonstrated the potential of Oxymatrine to reduce the invasiveness and metastasis of TNBC cells (Oxymatrine group vs. control group). Further, the results showed that Oxymatrine indeed attenuated Bevacizumab-induced invasion and migration of MDA-MB-231 cells (**P < 0.01) and MDA-MB-468 cells (##P < 0.01), which indicated Oxymatrine strongly suppressed Bevacizumab-induced EMT of TNBC cells. We further determined the effects of Oxymatrine on the proportion of ALDH1+ cells in TNBC. Figure 3A, 3B revealed that, Oxymatrine reduced ALDH1+ population cells of TNBC significantly (Oxymatrine group vs. control group, MDA-MB-231: **P < 0.01, MDA-MB-468: ##P < 0.01), and Oxymatrine reversed the aberrantly increased proportion of ALDH1+ cells induced by Bevacizumab after treatment together with Bevacizumab (Bevacizumab + Oxymatrine group vs. Bevacizumab group, MDA-MB-231: **P < 0.01, MDA-MB-468: ##P < 0.01). The specificity of ALDEFLUOR assay was determined by treating the cells with DEAB, a specific inhibitor of ALDH. Above all, Oxymatrine not only reversed Bevacizumab induced EMT of TNBC cells, but also suppressed Bevacizumab-increased TNBC stem-like cells population.
Figure 2.
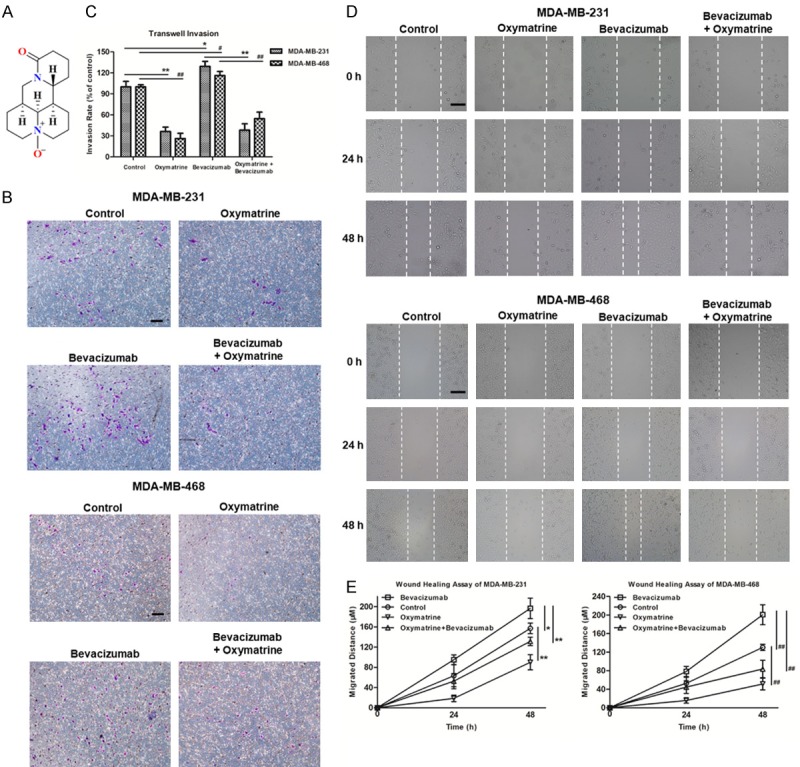
Oxymatrine inhibited Bevacizumab-induced transwell and migration of TNBC cells. (A) Chemical structure of Oxymatrine. (B) Microscopic views from transwell assay to estimate MDA-MB-231/MDA-MB-468 cells invasion following 24 h treatment of Oxymatrine (4 mM)/Bevacizumab (200 nM)/Oxymatrine (4 mM) + Bevacizumab (200 nM), bar = 100 μm. (C) Quantitative analysis of (B) by Image J. (D) Photomicrographs of cell migration from wound healing assay in MDA-MB-231/MDA-MB-468 cells treated with Oxymatrine/Bevacizumab/Oxymatrine + Bevacizumab, bar = 200 μm. (E) Quantitative analysis of (D) by Image J. The quantitative analyses of transwell invasion assay and wound healing assay were based on the mean of the 5 regions of each group. Data were presented as the mean ± SD, n = 5, *P < 0.05, **P < 0.01 (MDA-MB-231 cells); #P < 0.05, ##P < 0.01 (MDA-MB-468 cells).
Figure 3.

Oxymatrine suppressed the increased percent of ALDH1+ TNBC cells induced by Bevacizumab. (A) MDA-MB-231/MDA-MB-468 cells were treated with Oxymatrine/Bevacizumab/Oxymatrine + Bevacizumab for 24 h, then ALDH1+ cells were determined by Alderfluor assay and viable cells (7-AAD negative) were used for analysis. DEAB-treated cells were used as negative control. (B) Quantitative analysis of (A). Data were presented as the mean ± SD, n = 3, **P < 0.01 (MDA-MB-231 cells); ##P < 0.01 (MDA-MB-468 cells).
Oxymatrine inhibited EMT and Wnt/β-Catenin pathway activated by Bevacizumab in TNBC cells
Firstly, the expression level of various classical EMT markers in TNBC cells were detected by Western blot. Following treatment of Bevacizumab in serum-deprived environment, the expression level of N-cadherin and Vimentin were increased; and the expression of epithelial-associated gene E-cadherin decreased compared with control. The expression of EMT related transcription factors including ZEB, Snail, and Slug were also increased (Figure 4A, 4D). Meanwhile, Oxymatrine and FH535 exhibited an exactly opposite effect on EMT of TNBC cells compared to Bevacizumab (Figure 4A, 4D). Further, the analysis of the data from Bevacizumab + Oxymatrine group and Bevacizumab group indicated that, Oxymatrine reversed the Bevacizumab-induced increase in the expression of N-cadherin, Vimentin, ZEB, Snail and Slug, and increased E-cadherin expression in TNBC cells. Our findings strongly suggested that Oxymatrine effectively repressed Bevacizumab-induced EMT in TNBC cells.
Figure 4.

Oxymatrine inhibited EMT and Wnt/β-Catenin signaling induced by Bevacizumab. (A) Oxymatrine inhibited Bevacizumab-induced EMT in TNBC cells. MDA-MB-231/MDA-MB-468 cells were incubated with Oxymatrine/FH535 (20 μM)/Bevacizumab/Oxymatrine + Bevacizumab for 24 h. Then the results of Western blot analysis for EMT marker proteins were shown. (B) Oxymatrine repressed the accumulation of β-Catenin in the nucleus induced by Bevacizumab. IF stainings of β-Catenin (green) were shown, and nuclei were counterstained with DAPI (blue), bar = 20 μm. (C) Oxymatrine suppressed the activation of Wnt/β-Catenin pathway induced by Bevacizumab. The results of Western blot analysis for the expression of β-Catenin and downstream oncoproteins were shown. Histone H3 was used as loading control for nulear proteins, and β-actin was for cytoplasmic proteins (A, C). (D) Relative expression levels of the proteins indicated in (A). (E) Relative expression levels of nuclear β-Catenin indicated in (C). (F) Relative expression levels of the proteins indicated in (C) except nuclear β-Catenin. Data were presented as the mean ± SD, n = 3, *P < 0.05, **P < 0.01, NS: no significance.
Since Wnt/β-Catenin signaling played a crucial role in EMT, we further demonstrated that whether Bevacizumab-induced EMT involved the activation of Wnt/β-Catenin signaling [10,38]. IF staining of β-Catenin was performed to determine the localization of β-Catenin, and thereby the effect of Bevacizumab on nuclear β-Catenin accumulation was verified [39]. Figure 4B showed that, compared to the control group, Bevacizumab promoted the translocation of β-Catenin from the cytoplasm to the nucleus in TNBC cells. Figure 4C, 4E further confirmed the increased β-Catenin level in nucleus (Bevacizumab group vs. control group). Intriguingly, both cytoplasmatic and nuclear β-Catenin expression level of Bevacizumab-treated TNBC cells were improved compared to the control. Bevacizumab also induced the expression of c-Myc, Cyclin D1 and CD44, downstream components of the Wnt/β-Catenin pathway (Figure 4C, 4F). To sum up, the nuclear accumulation of β-Catenin, along with the increased expression level of total β-Catenin protein and downstream components strongly indicated Bevacizumab activated Wnt/β-Catenin pathway remarkably. On the contrary, Oxymatrine exhibited the potency of disrupting Wnt/β-Catenin pathway (vs. control group), which was similar to FH535 (Figure 4B, 4C). After making a further comparative study of Bevacizumab + Oxymatrine group and Bevacizumab group, we found β-Catenin staining was attenuated in the nuclei and remained in the cytoplasm, and total β-Catenin protein level declined notably in Bevacizumab + Oxymatrine group (Figure 4B, 4C). Furthermore, the expression of Wnt/β-Catenin downstream oncoproteins (c-Myc, Cyclin D1, CD44) was repressed in Bevacizumab + Oxymatrine group (vs. Bevacizumab group, Figure 4C, 4F). It was noteworthy that, VEGF, an important target gene of Wnt/β-Catenin pathway and also a direct target of Bevacizumab, was down-regulated by Bevacizumab and Oxymatrine synergistically (Figure 4C, 4F). In the immunoblotting assay, Histone H3 was used as an internal control for nuclear protein and β-actin was used as the control for cytoplasm protein. The comprehensive analysis of IF and Western blot indicated that Oxymatrine markedly suppressed Wnt/β-Catenin pathway activated by Bevacizumab, and thus inhibited EMT in TNBC cells.
Oxymatrine enhanced the anti-tumor activity of Bevacizumab against TNBC in vivo
Since Oxymatrine was demonstrated to suppress Wnt/β-Catenin pathway, inhibit EMT and deplete subpopulation of CSC effectively in vitro, TNBC cell xenografts in nude mice assay was executed to investigate the anti-tumor capacity of Oxymatrine in vivo. The tumor photographs (Figure 5A, 3 tumors of each group were shown, the other 2 not shown were treated for further study in the fresh condition) and tumor growth curves (Figure 5B, 5C) of each group showed that, Oxymatrine inhibited the tumor growth in MDA-MB-231/MDA-MB-468 xenografts. Notably, the high dose (25 mg/kg) of Oxymatrine achieved 62.16 ± 5.82% MDA-MB-231 tumor growth inhibition (Figure 5B) and 62.54 ± 4.29% MDA-MB-468 tumor growth inhibition (Figure 5C) compared to the PBS control. After making a further comparative study of the data from Bevacizumab group and Bevacizumab + Oxymatrine group, we found Bevacizumab + Oxymatrine significantly reduced tumor growth when compared either to PBS control (MDA-MB-231: **P < 0.01, MDA-MB-468: ##P < 0.01) or Bevacizumab alone treatment group (MDA-MB-231: *P < 0.05, MDA-MB-468: #P < 0.05). Figure 5B, 5C verified that FH535 showed a similar effect with Oxymatrine. What differentiated FH535 from Oxymatrine was that, the anti-tumor synergistic effect of FH535 was weaker than Oxymatrine when combined with Bevacizumab; as showed in Figure 5C, there was no significant difference between the MDA-MB-468 tumor volume of Bevacizumab group and Bevacizumab + FH535 group at the endpoint of administration. Moreover, let it be noted that Bevacizumab + Oxymatrine exhibited slightly better anti-proliferative activity than Bevacizumab + Docetaxel, which was one of the relatively effective clinical therapies of TNBC.
Figure 5.

Oxymatrine combined with Bevacizumab significantly inhibited tumor growth in MDA-MB-231/MDA-MB-468 xenografts. A. Representative images of isolated tumors from MDA-MB-231 (the left 3 rows)/MDA-MB-468 (the right 3 rows)-tumor bearing nude mice. B, C. MDA-MB-231/MDA-MB-468 tumor growth curves of each group under different treatments. Data were given as the mean ± SD (n = 5), *P < 0.05, **P < 0.01 (MDA-MB-231 tumor); #P < 0.05, ##P < 0.01, NS: no significance (MDA-MB-468 tumor). D, E. IHC staining of Ki-67 on paraffin sections of MDA-MB-231/MDA-MB-468 xenografted tumor. Ki-67+ cells were identified with an anti-Ki-67 antibody (brown staining), bar = 20 μm. F. Ki-67 positive cells of tumor tissues were counted using Image-Pro-Plus program, and the percent of Ki-67 positive cells of each group was shown. Data were given as the mean ± SD (n = 3), *P < 0.05, **P < 0.01.
Transplanted tumor tissues were harvested for IHC at the end of the treatment period. IHC demonstrated that there was a significant decrease in the numbers and intensity of cell proliferation marker Ki-67 in Bevacizumab + Oxymatrine group compared to PBS control or Bevacizumab group (Figure 5D-F), which was consistent with the results of the inhibition of TNBC tumor growth. In conclusion, Oxymatrine could enhance the anti-tumor effects of Bevacizumab against TNBC effectively.
Oxymatrine abated Wnt/β-Catenin pathway of TNBC activated by Bevacizumab in vivo and weakened the self-renew capacity of TNBC cells enhanced by Bevacizumab
We have verified that Oxymatrine indeed abated Wnt/β-Catenin pathway in TNBC cells under conditions with and without Bevacizumab treatment in vitro. To extend these observations to the mouse models, we examined the expression and localization of β-Catenin in MDA-MB-231/MDA-MB-468 tumors by IF. As shown in Figure 6A, 6B, β-Catenin was primarily detected in the cytoplasm of tumor cells from PBS control group. In contrast, cells within Bevacizumab-treated tumors displayed distinct nuclear localization of β-Catenin (typically as the white arrow indicated). The activated Wnt/β-Catenin pathway was probably induced by Bevacizumab via generating intratumoral hypoxia in human TNBC xenografts [11,30]. Compared to Bevacizumab group, we found Oxymatrine inhibited the accumulation of nuclear β-Catenin and reduced total expression level of β-Catenin, which were in agreement with the data in vitro. Hence, we came to conclusion that Oxymatrine could abate Wnt/β-Catenin pathway of TNBC activated by Bevacizumab in vivo.
Figure 6.
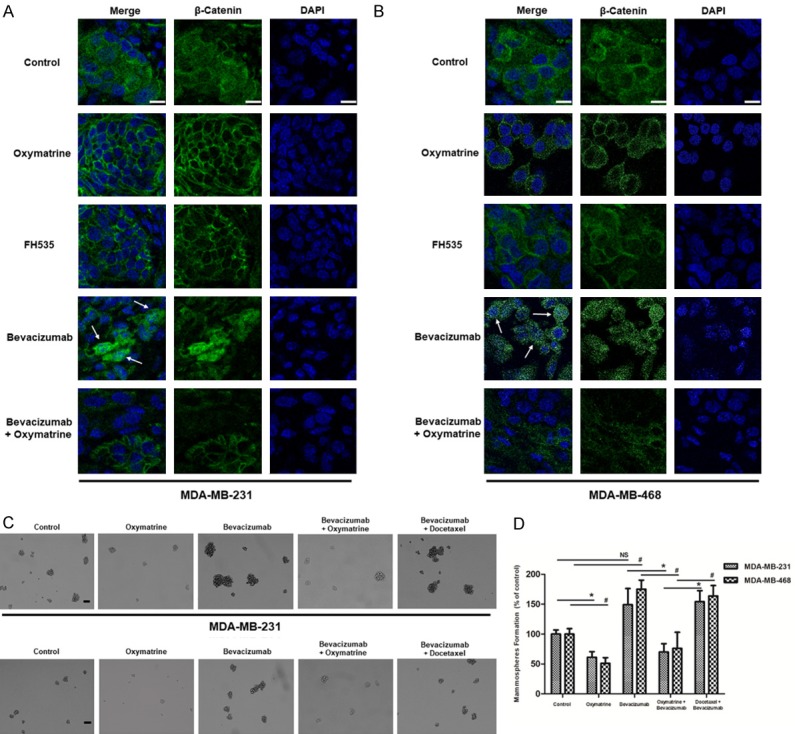
Oxymatrine repressed Bevacizumab-induced accumulation of β-Catenin in nucleus of MDA-MB-231/MDA-MB-468-tumor xenografts, and reduced mammosphere formation of tumor cells. A, B. IF staining of β-Catenin (green) on paraffin sections of MDA-MB-231/MDA-MB-468 xenografted tumor. Nuclei were counterstained with DAPI (blue), bar = 10 μm. C. 5 × 103 single cells isolated from MDA-MB-231/MDA-MB-468 xenografts were cultured in low attachment 6 well culture plates for 2 weeks to form the mammospheres, bar = 100 μm. D. Quantitation of the mammosphere-forming efficiency. The numbers of mammospheres (> 60 μm) were recorded after 2 weeks of culture. Data were presented as the mean ± SD, n = 3, *P < 0.05, NS: no significance (MDA-MB-231 cells); #P < 0.05 (MDA-MB-468 cells).
Multiple researches have verified that Wnt/β-Catenin pathway played a key role in the behavior of CSCs, especially the self-renew capacity of cancer cells. Then, mammosphere formation assay was executed to evaluate the self-renew capacity of TNBC cells. Figure 6C, 6D showed an increase in mammosphere formation in Bevacizumab treatment group compared to the control, which indicated Bevacizumab could enhance the self-renew capacity of TNBC cells. It was also observed in Figure 6C, 6D that, Oxymatrine attenuated mammosphere formation capacity of TNBC cells under conditions with and without Bevacizumab treatment. Moreover, compared to Bevacizumab + Docetaxel group, TNBC cells of Bevacizumab + Oxymatrine group exhibited significantly weaker mammosphere formation capacity (MDA-MB-231: *P < 0.05, MDA-MB-468: #P < 0.05), which highlighted the advantage of Oxymatrine when combined with Bevacizumab. Above all, Oxymatrine suppressed Bevacizumab-enhanced self-renew capacity of TNBC cells, which predicted Oxymatrine could prevent TNBC recurrence and metastasis after bevacizumab treatment.
Oxymatrine exhibited potential anti-angiogenic activity in vivo and in vitro
Tumor angiogenesis is one of the important components of tumor microenvironment, playing an oncogenic role in tumorigenesis, also is the target of the anti-angiogenic agent. The CD31 staining (Figure 7A, 7B) was executed to evaluate tumor angiogenesis of each treatment group. We were delighted to find that Oxymatrine treatment reduced the blood vessel density effectively (Oxymatrine group vs. PBS control). The remarkably reduced intensity of CD31 staining in Bevacizumab + Oxymatrine group indicated that the density of tumor neovascularization was significantly lower in Bevacizumab + Oxymatrine group compared to Bevacizumab group or Oxymatrine group, further illustrating the synergistic anti-angiogenesis synergistic effect of Bevacizumab and Oxymatrine.
Figure 7.

Oxymatrine combined with Bevacizumab reduced markers of angiogenesis in MDA-MB-231/MDA-MB-468-tumor xenografts. A, B. IF staining of CD31 on paraffin sections of MDA-MB-231/MDA-MB-468 implanted tumor. The CD31+ blood vessels were identified with an anti-CD31 antibody (red), and nuclei were counterstained with DAPI (blue), bar = 25 μm.
VEGF, a key pro-angiogenic factor, was then detected by IHC and Western blot. As an important target gene of Wnt/β-Catenin pathway, VEGF was supposed to be down regulated by Oxymatrine. Figure 8A-D corroborated the speculation that the expression level of VEGFA declined after Oxymatrine treatment. Compared to other treatment groups, Bevacizumab + Oxymatrine group exhibited the lowest expression of VEGFA, which was consistent with the results of CD31 staining.
Figure 8.

Oxymatrine exhibited potential anti-angiogenic activity in vivo and in vitro. (A, B) IHC staining of VEGFA (brown staining) on paraffin sections of MDA-MB-231/MDA-MB-468 xenografted tumor, bar = 20 μm. (C) A subset of MDA-MB-231/MDA-MB-468 tumor tissues were subjected to Western blot to examine the VEGFA protein levels. β-actin was used as loading control for proteins. (D) Relative expression levels of VEGFA protein after gray scanning. Data were presented as the mean ± SD, n = 3, *P < 0.05, **P < 0.01, NS: no significance. (E) The viability of HUVECs was assessed by MTT assay at 48 h after treatment with different concentrations of Oxymatrine (ranging from 0 to 8 mg/mL), data were presented as the mean ± SD, n = 3, *P < 0.05, **P < 0.01, vs. the previous group. (F) Microscopic views from transwell assay to estimate HUVECs invasion following 24 h treatment of different concentrations of Oxymatrine (0, 1, 2, and 4 mM), bar = 100 μm. (G) Quantitative analysis of (F) by Image J. (H) Photomicrographs of HUVECs tube formation treated by different concentrations of Oxymatrine for 8 h, bar = 100 μm. (I) Quantitative analysis of (H) by Image J. The quantitative analyses of transwell invasion and HUVECs tube formation (a complete polygon was considered as a tube) were based on the mean of the 5 regions of each group. Data were presented as the mean ± SD, n = 5, *P < 0.05, **P < 0.01, vs. the previous group.
To further verify the anti-angiogenic potential of Oxymatrine, we studied its effects on the proliferation, transwell invasion, and tube formation of HUVECs. Figure 8E showed that, Oxymatrine effectively inhibited the growth of HUVECs in a dose-dependent manner in vitro. After 48 h effect of Oxymatrine, the IC50 value was 0.51 mM. However, no inhibition on HEK293 was seen. As shown in Figure 8F, 8G, the invasive cells to the lower chamber were significantly reduced with increasing concentration of Oxymatrine, and the inhibitory rate of 4 mM Oxymatrine on the invasion of HUVECs reached 76.2 ± 3.43%. The maturation of endothelial cells into a capillary tube, a critical early step of angiogenesis, was gradually abrogated with increasing concentrations of Oxymatrine, and the inhibitory rate of 4 mM Oxymatrine on the tube formation of HUVECs reached 92.8 ± 1.89% (Figure 8H, 8I). Oxymatrine did not show any cytotoxicity in the transwell and tube formation assays (data not shown). In conclusion, Oxymatrine exhibited superior anti-angiogenic activities in vivo and in vitro, and enhanced the anti-angiogenic effect when combined with Bevacizumab.
Discussion
Anti-angiogenic therapy has been demonstrated to hold significant potential for the treatment of various cancers, including TNBC. However, the efficacy of such therapy, especially in TNBC patients, has been called into question, as recent clinical trials reveal only limited effectiveness of anti-angiogenic agents in prolonging patient survival [40]. Conley SJ et al [11], found that anti-angiogenic agents increased the population of CSCs by generating intratumoral hypoxia in human TNBC xenografts, further demonstrated hypoxia-driven CSC stimulation limited the effectiveness of antiangiogenic agents. Huang W et al [30], showed that Bevacizumab induced EMT and promoted tumor invasion in malignancies, which compromised its efficacy severely. Sun H et al [40], indicated that anti-angiogenic treatment induced the formation of vasculogenic mimicry (VM) channels via causing acute hypoxic stress in TNBC; this effect might influence eventual tumor progression, invasion and angiogenesis rebound. Furthermore, there has been increasing evidence for that, the Wnt/β-Catenin pathway involved in EMT/CSC was activated in Sunitinib/Bevacizumab-treated TNBC cells under hypoxia/serum deprivation condition in vitro and in Sunitinib/Bevacizumab-treated mouse xenografts [41,42]. These studies suggest that to improve patient outcome, anti-angiogenic agents need to be combined with Wnt/β-Catenin pathway/CSC-targeting drugs. Additional approaches for targeting the hypoxic tumor microenvironment are being investigated in pre-clinical and clinical studies, which also have potential for producing synthetic lethality when combined with anti-angiogenic therapy [43,44].
Through a series of experiments, Marjon PL et al [45], found that TNBC cell line was sensitive to serum deprivation rather than hypoxia. There’s more evidence that hypoxia was just one of the triggering factors for the production of tumor specific growth factors (TSGFs) [46,47]. Thus, in this study, we utilized cell serum starvation model to confirm Bevacizumab treatment induced TNBC cells transwell, migration in vitro. Figure 1A, 1C revealed that, compared to serum starvation control group, invasion and migration of TNBC cells were enhanced after Bevacizumab treatment, which could be due to the expression of invasive factors and transfer factors stimulated by the ablation of VEGF pathway [48]. Figures 1D and 4 indicated that, in serum-deprived environment, Bevacizumab increased the subpopulation of TNBC stem-like cells and promoted EMT of TNBC via activating Wnt/β-Catenin pathway, which probably accounted for the invasion and migration of TNBC cells. Similarly, Wnt/β-Catenin pathway was also activated in Bevacizumab-treated mouse xenografts (Figure 6A, 6B), mainly due to the generation of tumor hypoxia [49]. As a key regulator of breast CSC self-renewal [38], activated Wnt/β-Catenin pathway further enhanced mammosphere formation capacity of TNBC cells (Figure 6C).
This study showed that, Oxymatrine reversed the Bevacizumab-activated Wnt/β-Catenin pathway and Bevacizumab-induced EMT in TNBC effectively; in addition, Oxymatrine exhibited potent anti-angiogenic activities, which implied Oxymatrine was a potential candidate for the drug combination with anti-angiogenic agents in the treatment of TNBC. Figures 2, 3 suggested Oxymatrine inhibited the invasion, migration of TNBC cells, and decreased the subpopulation of TNBC stem-like cells through repressing EMT and Wnt/β-Catenin pathway stimulated by Bevacizumab. Furthermore, in vivo (Figure 6A, 6B), Wnt/β-Catenin pathway was disrupted remarkably in the Bevacizumab + Oxymatrine group compared to the Bevacizumab group, which was consistent with cell studies. Moreover, Figures 7 and 8A-D clearly demonstrated Oxymatrine strengthened the anti-angiogenic potency of Bevacizumab in a synergistic mechanism. In view of the above, Bevacizumab + Oxymatrine treatment achieved a better curative effect, which embodied in tumor growth curves (Figure 5B, 5C) and tumor proliferation (Figure 5D, 5E). The enhanced TNBC CSC self-renewal capacity portended tumor recurrence and metastasis, going straight to the heart of Bevacizumab’s problem, which was also an key problem during the clinical application of Docetaxel. Notably, Figure 6C verified that Oxymatrine suppressed Bevacizumab-enhanced self-renew capacity of TNBC cells, indicating the combination of Oxymatrine and Bevacizumab possibly reduced the risk of tumor recurrence and metastasis caused by Bevacizumab treatment alone.
Since anti-angiogenic agents increased TNBC CSCs via the generation of tumor hypoxia, which was demonstrated to be mediated by hypoxia-inducible factor 1α (HIF-1α), an important upstream factor of the β-Catenin response [38,50,51]; Oxymatrine was hypothesized to stimulate TNBC CSC behavior as a direct effect of the hypoxia induced by the angiogenic inhibition. While, mammosphere formation assay (Figure 6C) exhibited a contrary result that, Oxymatrine negatively regulated TNBC stem cell self-renewal. Figure 6A, 6B indicated the effects of Oxymatrine was at least partly regulated by blocking β-Catenin signaling; but the effects of Oxymatrine on the critical factor HIF-1α needed to be further investigated, on account of that HIF-1α was reported to enhance β-Catenin signaling in hypoxic embryonic stem cells and be involved in EMT.
The findings presented here provided additional evidence suggesting that Oxymatrine was sufficient to inhibit Bevacizumab-induced cancer stem-like properties and EMT with a suppressive effect on Wnt/β-Catenin signaling. The combination of Oxymatrine and Bevacizumab achieved better anti-tumor and anti-angiogenic effects, further reduced the risk of TNBC recurrence and metastasis induced by Bevacizumab treatment alone. In conclusion, the concurrent use of Oxymatrine and Bevacizumab provides an effective candidate for the clinical treatment of TNBC, and lays foundation for the synergistic strategy of combining anti-angiogenic agents with Wnt/CSC-targeting agents.
Acknowledgements
This study was supported by the National Natural Science Foundation of China (NSFC81703401, NSFC81801833, NSFC 81830052 and NSFC81530053), “Chen Guang” project supported by Shanghai Municipal Education Commission and Shanghai Education Development Foundation (18CG72), and Shanghai Key Laboratory of Molecular Imaging (18DZ2260400).
Disclosure of conflict of interest
None.
Abbreviations
- ALDH
aldehyde dehydrogenase
- CSC
cancer stem cell
- DEAB
diethylaminobenzaldehyde
- ECGS
endothelial cell growth supplement
- ECM
endothelial culture medium
- EGF
epidermal growth factor
- EMT
epithelial-mesenchymal cell transformation
- FGF
fibroblast growth factor
- HIF-1α
hypoxia-inducible factor 1α
- HUVECs
human umbilical vascular endothelial cells
- IF
immunofluorescence
- IHC
immunohistochemistry
- mCRC
metastatic colorectal cancer
- NSCLS
non-small cell lung cancer
- ORR
objective response rate
- OS
overall survival
- PFS
progression-free survival
- TCM
traditional Chinese medicine
- TNBC
triple-negative breast cancer
- TSGF
tumor specific growth factor
- VEGF-A
vascular endothelial growth factor A
Supporting Information
References
- 1.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69:7–34. doi: 10.3322/caac.21551. [DOI] [PubMed] [Google Scholar]
- 2.Sikov WM, Berry DA, Perou CM, Singh B, Cirrincione CT, Tolaney SM, Kuzma CS, Pluard TJ, Somlo G, Port ER, Golshan M, Bellon JR, Collyar D, Hahn OM, Carey LA, Hudis CA, Winer EP. Impact of the addition of carboplatin and/or bevacizumab to neoadjuvant once-per-week paclitaxel followed by dose-dense doxorubicin and cyclophosphamide on pathologic complete response rates in stage II to III triple-negative breast cancer: CALGB 40603 (Alliance) J. Clin. Oncol. 2015;33:13–21. doi: 10.1200/JCO.2014.57.0572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bardia A, Mayer IA, Vahdat LT, Tolaney SM, Isakoff SJ, Diamond JR, O’Shaughnessy J, Moroose RL, Santin AD, Abramson VG, Shah NC, Rugo HS, Goldenberg DM, Sweidan AM, Iannone R, Washkowitz S, Sharkey RM, Wegener WA, Kalinsky K. Sacituzumab govitecan-hziy in refractory metastatic triple-negative breast cancer. N Engl J Med. 2019;380:741–751. doi: 10.1056/NEJMoa1814213. [DOI] [PubMed] [Google Scholar]
- 4.Carey L, Winer E, Viale G, Cameron D, Gianni L. Triple-negative breast cancer: disease entity or title of convenience? Nat Rev Clin Oncol. 2010;7:683–692. doi: 10.1038/nrclinonc.2010.154. [DOI] [PubMed] [Google Scholar]
- 5.Cameron D, Brown J, Dent R, Jackisch C, Mackey J, Pivot X, Steger GG, Suter TM, Toi M, Parmar M, Laeufle R, Im YH, Romieu G, Harvey V, Lipatov O, Pienkowski T, Cottu P, Chan A, Im SA, Hall PS, Bubuteishvili-Pacaud L, Henschel V, Deurloo RJ, Pallaud C, Bell R. Adjuvant bevacizumab-containing therapy in triple-negative breast cancer (BEATRICE): primary results of a randomised, phase 3 trial. Lancet Oncol. 2013;14:933–942. doi: 10.1016/S1470-2045(13)70335-8. [DOI] [PubMed] [Google Scholar]
- 6.Dieras V, Campone M, Yardley DA, Romieu G, Valero V, Isakoff SJ, Koeppen H, Wilson TR, Xiao Y, Shames DS, Mocci S, Chen M, Schmid P. Randomized, phase II, placebo-controlled trial of onartuzumab and/or bevacizumab in combination with weekly paclitaxel in patients with metastatic triple-negative breast cancer. Ann Oncol. 2015;26:1904–1910. doi: 10.1093/annonc/mdv263. [DOI] [PubMed] [Google Scholar]
- 7.Miles DW, Dieras V, Cortes J, Duenne AA, Yi J, O’Shaughnessy J. First-line bevacizumab in combination with chemotherapy for HER2-negative metastatic breast cancer: pooled and subgroup analyses of data from 2447 patients. Ann Oncol. 2013;24:2773–2780. doi: 10.1093/annonc/mdt276. [DOI] [PubMed] [Google Scholar]
- 8.Miller K, Wang M, Gralow J, Dickler M, Cobleigh M, Perez EA, Shenkier T, Cella D, Davidson NE. Paclitaxel plus bevacizumab versus paclitaxel alone for metastatic breast cancer. N Engl J Med. 2007;357:2666–2676. doi: 10.1056/NEJMoa072113. [DOI] [PubMed] [Google Scholar]
- 9.Hurwitz H, Fehrenbacher L, Novotny W, Cartwright T, Hainsworth J, Heim W, Berlin J, Baron A, Griffing S, Holmgren E, Ferrara N, Fyfe G, Rogers B, Ross R, Kabbinavar F. Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N Engl J Med. 2004;350:2335–2342. doi: 10.1056/NEJMoa032691. [DOI] [PubMed] [Google Scholar]
- 10.Gurney A, Axelrod F, Bond CJ, Cain J, Chartier C, Donigan L, Fischer M, Chaudhari A, Ji M, Kapoun AM, Lam A, Lazetic S, Ma S, Mitra S, Park IK, Pickell K, Sato A, Satyal S, Stroud M, Tran H, Yen WC, Lewicki J, Hoey T. Wnt pathway inhibition via the targeting of Frizzled receptors results in decreased growth and tumorigenicity of human tumors. Proc Natl Acad Sci U S A. 2012;109:11717–11722. doi: 10.1073/pnas.1120068109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Conley SJ, Gheordunescu E, Kakarala P, Newman B, Korkaya H, Heath AN, Clouthier SG, Wicha MS. Antiangiogenic agents increase breast cancer stem cells via the generation of tumor hypoxia. Proc Natl Acad Sci U S A. 2012;109:2784–2789. doi: 10.1073/pnas.1018866109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Linke F, Harenberg M, Nietert MM, Zaunig S, von Bonin F, Arlt A, Szczepanowski M, Weich HA, Lutz S, Dullin C, Janovska P, Krafcikova M, Trantirek L, Ovesna P, Klapper W, Beissbarth T, Alves F, Bryja V, Trumper L, Wilting J, Kube D. Microenvironmental interactions between endothelial and lymphoma cells: a role for the canonical WNT pathway in Hodgkin lymphoma. Leukemia. 2017;31:361–372. doi: 10.1038/leu.2016.232. [DOI] [PubMed] [Google Scholar]
- 13.Zhang L, Wang H, Li C, Zhao Y, Wu L, Du X, Han Z. VEGF-A/neuropilin 1 pathway confers cancer stemness via activating wnt/beta-Catenin axis in breast cancer cells. Cell Physiol Biochem. 2017;44:1251–1262. doi: 10.1159/000485455. [DOI] [PubMed] [Google Scholar]
- 14.Zhang L, Xiong W, Xiong Y, Liu H, Liu Y. 17 beta-Estradiol promotes vascular endothelial growth factor expression via the Wnt/beta-catenin pathway during the pathogenesis of endometriosis. Mol Hum Reprod. 2016;22:526–535. doi: 10.1093/molehr/gaw025. [DOI] [PubMed] [Google Scholar]
- 15.Paez-Ribes M, Allen E, Hudock J, Takeda T, Okuyama H, Vinals F, Inoue M, Bergers G, Hanahan D, Casanovas O. Antiangiogenic therapy elicits malignant progression of tumors to increased local invasion and distant metastasis. Cancer Cell. 2009;15:220–231. doi: 10.1016/j.ccr.2009.01.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Saleem M, Maddodi N, Abu Zaid M, Khan N, bin Hafeez B, Asim M, Suh Y, Yun JM, Setaluri V, Mukhtar H. Lupeol inhibits growth of highly aggressive human metastatic melanoma cells in vitro and in vivo by inducing apoptosis. Clin Cancer Res. 2008;14:2119–2127. doi: 10.1158/1078-0432.CCR-07-4413. [DOI] [PubMed] [Google Scholar]
- 17.Wang XJ, Zhou RJ, Zhang N, Jing Z. 20(S)-ginsenoside Rg3 sensitizes human non-small cell lung cancer cells to icotinib through inhibition of autophagy. Eur J Pharmacol. 2019;850:141–149. doi: 10.1016/j.ejphar.2019.02.023. [DOI] [PubMed] [Google Scholar]
- 18.Zhou ZL, Yang YX, Ding J, Li YC, Miao ZH. Triptolide: structural modifications, structure-activity relationships, bioactivities, clinical development and mechanisms. Nat Prod Rep. 2012;29:457–475. doi: 10.1039/c2np00088a. [DOI] [PubMed] [Google Scholar]
- 19.Li Z, Zhou ZL, Miao ZH, Lin LP, Feng HJ, Tong LJ, Ding J, Li YC. Design and synthesis of novel C14-hydroxyl substituted triptolide derivatives as potential selective antitumor agents. J Med Chem. 2009;52:5115–5123. doi: 10.1021/jm900342g. [DOI] [PubMed] [Google Scholar]
- 20.Liu Y, Xu Y, Ji W, Li X, Sun B, Gao Q, Su C. Anti-tumor activities of matrine and oxymatrine: literature review. Tumour Biol. 2014;35:5111–5119. doi: 10.1007/s13277-014-1680-z. [DOI] [PubMed] [Google Scholar]
- 21.Lu ML, Xiang XH, Xia SH. Potential signaling pathways involved in the clinical application of oxymatrine. Phytother Res. 2016;30:1104–1112. doi: 10.1002/ptr.5632. [DOI] [PubMed] [Google Scholar]
- 22.Wu J, Cai Y, Li M, Zhang Y, Li H, Tan Z. Oxymatrine promotes s-phase arrest and inhibits cell proliferation of human breast cancer cells in vitro through mitochondria-mediated apoptosis. Biol Pharm Bull. 2017;40:1232–1239. doi: 10.1248/bpb.b17-00010. [DOI] [PubMed] [Google Scholar]
- 23.Li W, Yu X, Tan S, Liu W, Zhou L, Liu H. Oxymatrine inhibits non-small cell lung cancer via suppression of EGFR signaling pathway. Cancer Med. 2018;7:208–218. doi: 10.1002/cam4.1269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lin B, Li D, Zhang L. Oxymatrine mediates Bax and Bcl-2 expression in human breast cancer MCF-7 cells. Pharmazie. 2016;71:154–157. [PubMed] [Google Scholar]
- 25.Zhang Y, Piao B, Zhang Y, Hua B, Hou W, Xu W, Qi X, Zhu X, Pei Y, Lin H. Oxymatrine diminishes the side population and inhibits the expression of beta-catenin in MCF-7 breast cancer cells. Med Oncol. 2011;28(Suppl 1):S99–107. doi: 10.1007/s12032-010-9721-y. [DOI] [PubMed] [Google Scholar]
- 26.Song MQ, Zhu JS, Chen JL, Wang L, Da W, Zhu L, Zhang WP. Synergistic effect of oxymatrine and angiogenesis inhibitor NM-3 on modulating apoptosis in human gastric cancer cells. World J Gastroenterol. 2007;13:1788–1793. doi: 10.3748/wjg.v13.i12.1788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Xie W, Li D, Zhang J, Li Z, Acheampong DO, He Y, Wang Y, Chen Z, Wang M. Generation and characterization of a novel human IgG1 antibody against vascular endothelial growth factor receptor 2. Cancer Immunol Immunother. 2014;63:877–888. doi: 10.1007/s00262-014-1560-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Xie W, Zhang Y, He Y, Zhang K, Wan G, Huang Y, Zhou Z, Huang G, Wang J. A novel recombinant human Frizzled-7 protein exhibits anti-tumor activity against triple negative breast cancer via abating Wnt/beta-catenin pathway. Int J Biochem Cell Biol. 2018;103:45–55. doi: 10.1016/j.biocel.2018.08.004. [DOI] [PubMed] [Google Scholar]
- 29.Xu Z, Wang Z, Jia X, Wang L, Chen Z, Wang S, Wang M, Zhang J, Wu M. MMGZ01, an anti-DLL4 monoclonal antibody, promotes nonfunctional vessels and inhibits breast tumor growth. Cancer Lett. 2016;372:118–127. doi: 10.1016/j.canlet.2015.12.025. [DOI] [PubMed] [Google Scholar]
- 30.Huang W, Zhang C, Cui M, Niu J, Ding W. Inhibition of bevacizumab-induced epithelial-mesenchymal transition by BATF2 overexpression involves the suppression of wnt/beta-catenin signaling in glioblastoma cells. Anticancer Res. 2017;37:4285–4294. doi: 10.21873/anticanres.11821. [DOI] [PubMed] [Google Scholar]
- 31.Gupta GP, Massague J. Cancer metastasis: building a framework. Cell. 2006;127:679–695. doi: 10.1016/j.cell.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 32.Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, Campbell LL, Polyak K, Brisken C, Yang J, Weinberg RA. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133:704–715. doi: 10.1016/j.cell.2008.03.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lo JF, Yu CC, Chiou SH, Huang CY, Jan CI, Lin SC, Liu CJ, Hu WY, Yu YH. The epithelial-mesenchymal transition mediator S100A4 maintains cancer-initiating cells in head and neck cancers. Cancer Res. 2011;71:1912–1923. doi: 10.1158/0008-5472.CAN-10-2350. [DOI] [PubMed] [Google Scholar]
- 34.Qiu M, Peng Q, Jiang I, Carroll C, Han G, Rymer I, Lippincott J, Zachwieja J, Gajiwala K, Kraynov E, Thibault S, Stone D, Gao Y, Sofia S, Gallo J, Li G, Yang J, Li K, Wei P. Specific inhibition of Notch1 signaling enhances the antitumor efficacy of chemotherapy in triple negative breast cancer through reduction of cancer stem cells. Cancer Lett. 2013;328:261–270. doi: 10.1016/j.canlet.2012.09.023. [DOI] [PubMed] [Google Scholar]
- 35.Bhola NE, Balko JM, Dugger TC, Kuba MG, Sanchez V, Sanders M, Stanford J, Cook RS, Arteaga CL. TGF-beta inhibition enhances chemotherapy action against triple-negative breast cancer. J Clin Invest. 2013;123:1348–1358. doi: 10.1172/JCI65416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Liu P, Kumar IS, Brown S, Kannappan V, Tawari PE, Tang JZ, Jiang W, Armesilla AL, Darling JL, Wang W. Disulfiram targets cancer stem-like cells and reverses resistance and cross-resistance in acquired paclitaxel-resistant triple-negative breast cancer cells. Br J Cancer. 2013;109:1876–1885. doi: 10.1038/bjc.2013.534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chang WW, Hu FW, Yu CC, Wang HH, Feng HP, Lan C, Tsai LL, Chang YC. Quercetin in elimination of tumor initiating stem-like and mesenchymal transformation property in head and neck cancer. Head Neck. 2013;35:413–419. doi: 10.1002/hed.22982. [DOI] [PubMed] [Google Scholar]
- 38.Korkaya H, Paulson A, Charafe-Jauffret E, Ginestier C, Brown M, Dutcher J, Clouthier SG, Wicha MS. Regulation of mammary stem/progenitor cells by PTEN/Akt/beta-catenin signaling. PLoS Biol. 2009;7:e1000121. doi: 10.1371/journal.pbio.1000121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yang L, Wu X, Wang Y, Zhang K, Wu J, Yuan YC, Deng X, Chen L, Kim CC, Lau S, Somlo G, Yen Y. FZD7 has a critical role in cell proliferation in triple negative breast cancer. Oncogene. 2011;30:4437–4446. doi: 10.1038/onc.2011.145. [DOI] [PubMed] [Google Scholar]
- 40.Sun H, Zhang D, Yao Z, Lin X, Liu J, Gu Q, Dong X, Liu F, Wang Y, Yao N, Cheng S, Li L, Sun S. Anti-angiogenic treatment promotes triple-negative breast cancer invasion via vasculogenic mimicry. Cancer Biol Ther. 2017;18:205–213. doi: 10.1080/15384047.2017.1294288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ebos JM, Lee CR, Cruz-Munoz W, Bjarnason GA, Christensen JG, Kerbel RS. Accelerated metastasis after short-term treatment with a potent inhibitor of tumor angiogenesis. Cancer Cell. 2009;15:232–239. doi: 10.1016/j.ccr.2009.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Schomber T, Zumsteg A, Strittmatter K, Crnic I, Antoniadis H, Littlewood-Evans A, Wood J, Christofori G. Differential effects of the vascular endothelial growth factor receptor inhibitor PTK787/ZK222584 on tumor angiogenesis and tumor lymphangiogenesis. Mol Cancer Ther. 2009;8:55–63. doi: 10.1158/1535-7163.MCT-08-0679. [DOI] [PubMed] [Google Scholar]
- 43.Vasudev NS, Goh V, Juttla JK, Thompson VL, Larkin JM, Gore M, Nathan PD, Reynolds AR. Changes in tumour vessel density upon treatment with anti-angiogenic agents: relationship with response and resistance to therapy. Br J Cancer. 2013;109:1230–1242. doi: 10.1038/bjc.2013.429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.McIntyre A, Harris AL. Metabolic and hypoxic adaptation to anti-angiogenic therapy: a target for induced essentiality. EMBO Mol Med. 2015;7:368–379. doi: 10.15252/emmm.201404271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Marjon PL, Bobrovnikova-Marjon EV, Abcouwer SF. Expression of the pro-angiogenic factors vascular endothelial growth factor and interleukin-8/CXCL8 by human breast carcinomas is responsive to nutrient deprivation and endoplasmic reticulum stress. Mol Cancer. 2004;3:4. doi: 10.1186/1476-4598-3-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mazzone M, Dettori D, de Oliveira RL, Loges S, Schmidt T, Jonckx B, Tian YM, Lanahan AA, Pollard P, de Almodovar CR, De Smet F, Vinckier S, Aragones J, Debackere K, Luttun A, Wyns S, Jordan B, Pisacane A, Gallez B, Lampugnani MG, Dejana E, Simons M, Ratcliffe P, Maxwell P, Carmeliet P. Heterozygous deficiency of PHD2 restores tumor oxygenation and inhibits metastasis via endothelial normalization. Cell. 2009;136:839–851. doi: 10.1016/j.cell.2009.01.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kallergi G, Markomanolaki H, Giannoukaraki V, Papadaki MA, Strati A, Lianidou ES, Georgoulias V, Mavroudis D, Agelaki S. Hypoxia-inducible factor-1alpha and vascular endothelial growth factor expression in circulating tumor cells of breast cancer patients. Breast Cancer Res. 2009;11:R84. doi: 10.1186/bcr2452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Li A, Dubey S, Varney ML, Dave BJ, Singh RK. IL-8 directly enhanced endothelial cell survival, proliferation, and matrix metalloproteinases production and regulated angiogenesis. J Immunol. 2003;170:3369–3376. doi: 10.4049/jimmunol.170.6.3369. [DOI] [PubMed] [Google Scholar]
- 49.Liu L, Zhu XD, Wang WQ, Shen Y, Qin Y, Ren ZG, Sun HC, Tang ZY. Activation of beta-catenin by hypoxia in hepatocellular carcinoma contributes to enhanced metastatic potential and poor prognosis. Clin Cancer Res. 2010;16:2740–2750. doi: 10.1158/1078-0432.CCR-09-2610. [DOI] [PubMed] [Google Scholar]
- 50.Mazumdar J, O’Brien WT, Johnson RS, LaManna JC, Chavez JC, Klein PS, Simon MC. O2 regulates stem cells through Wnt/beta-catenin signalling. Nat Cell Biol. 2010;12:1007–1013. doi: 10.1038/ncb2102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mendez O, Zavadil J, Esencay M, Lukyanov Y, Santovasi D, Wang SC, Newcomb EW, Zagzag D. Knock down of HIF-1alpha in glioma cells reduces migration in vitro and invasion in vivo and impairs their ability to form tumor spheres. Mol Cancer. 2010;9:133. doi: 10.1186/1476-4598-9-133. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.