

Abstract
Glioblastoma is the most common and malignant tumor of the CNS, with a mean survival of 14 months after diagnosis. Its unfavorable prognosis reveals the need for novel therapies. It is known that radiation can induce a systemic antitumor effect. Tumor cells produce and release microvesicles in response to cell damage such as radiation. Microvesicles contain a plethora of bioactive molecules, including antigens involved in modulation of the immune response. In this study, we characterized and evaluated irradiated C6 cell-derived microvesicles as a therapeutic vaccination in C6 malignant glioma. Cultured C6 glioma cells were irradiated with a single dose of 50 Gy to obtain the microvesicles. Subcutaneous implantation of C6 cells was performed when the tumor reached 2 cm in diameter, and non-irradiated and irradiated C6 cell-derived microvesicles were administered subcutaneously. Tumor growth, apoptosis, and immunophenotypes were determined. Reduction of tumor volume (more than 50%) was observed in the group treated with irradiated C6 cell-derived microvesicles compared with the control (p = 0.03). The percentages of infiltrative helper, cytotoxic, and regulatory T lymphocytes as well as apoptotic cells were increased in tumors from immunized rats compared with controls. These findings make microvesicle-based vaccination a promising immunotherapeutic approach against glioblastoma.
Keywords: malignant glioma, microvesicles, vaccination, antitumor immune response
Pineda et al. characterized and used irradiated C6 cell-derived microvesicles (IR-MVs) to demonstrate that IR-MV-based vaccination can elicit an antitumoral immune response against malignant glioma. They propose that IR-MV could be used as a therapeutic modality for the treatment of glioblastoma patients.
Introduction
Malignant brain tumors are devastating forms of cancer1 causing 15,000 deaths every year in the United States.2 Glioblastoma multiforme (GBM) is the most malignant and frequent primary tumor of the CNS.3 Treatment for GBM consists of extensive surgical resection followed by radiotherapy and chemotherapy.4 Radiotherapy is used as a modality to delay tumor growth.5 Radiation induces the formation of various damage-associated molecular patterns (DAMPs) that lead to activation of the innate immune system; this phenomenon is known as immunogenic cell damage. Additionally, patients with metastatic neoplasms who receive local radiotherapy show systemic immune activation; this feature is known as the abscopal effect (from the Latin ab scopus, “away from the target”).6 The therapeutic possibility of the abscopal effect was not recognized for decades until the advent of cancer immunotherapy. Clinical cases of abscopal responses are more frequent when radiation is associated with blockage of immune checkpoints such as CTLA-4 or PD-1, which provides a lasting antitumor immune response.7, 8 Nevertheless, immune activation by DNA breaks and micronucleus induction only has not been sufficient to obtain an effective therapeutic effect. Also, the liberation of extracellular vesicles (EVs) released by tumoral cells has not been fully exploited for potential therapeutic applications.9 These vesicles are particles of the lipid bilayer membrane that can be isolated from body fluids10 and are liberated by tumor cells in response to stress. In this context, microvesicles (MVs) (also referred to as microparticles or ectosomes) are a subtype of EVs released into the extracellular space by fission to the exterior of the plasma membrane; their size ranges from 200 nm to more than 1 μm.11, 12 At the site where MVs are released, phosphatidylserine, normally found on the cytoplasmic side of the membrane, is relocated to the outer layer of the membrane, whereas the topology of membrane proteins remains identical.13
The importance of MVs lies in their ability to transfer their content to other cells, either locally or systemically. The composition of MVs depends largely on the original cell, although the membrane composition of MVs might be different from the parental cell because of remodeling.14 Molecules contained in MVs from cancer cells include molecules that could participate in immunomodulation, transmembrane receptors and ligands, oncoproteins, tumor suppressor proteins, lipids, mRNA, microRNA, and genomic or mitochondrial DNA.15 These MVs can interact with various tissues, establishing pre-metastatic niches and promoting cell invasion, angiogenesis, and immune blockage.16, 17
It has been documented that ionizing radiation of glioma alters the abundance and composition of MVs, promoting a migratory phenotype.16 Recently, Baulch et al.18 demonstrated that radiation of human gliomas triggers a pro-oxidant phenotype, increasing the expression of genes involved in the regulation of cellular reprogramming and paracrine interactions that seem to be mediated by MVs, leading to tumor survival and invasiveness, with participation of metalloproteinase 2 (MMP-2). In addition, it has been shown that ionizing radiation induces the release of MVs and contributes to the formation of DAMPs,19 which can be released either passively by necrotic cells or secreted by damaged cells exposed to stress, inducing stimulation of the innate immune system through pattern recognition receptors (PRRs).20, 21 It is possible that MVs released by irradiated tumor cells might generate tumor-associated antigens (TAAs) and DAMPs within the MVs that make them immunogenic; this effect would partly explain the abscopal effect. Therefore, we investigated the capacity of immunization with MVs from C6 glioma cells to promote an antitumor immune response against malignant glioma using irradiated C6 cell-derived MVs (IR-MVs) as a therapeutic vaccination in rats with glioma and compared them with non-IR-MVs.
Results
Radiation-Induced MVs in Glioma C6 Cells
MVs from the C6 glioma cell line were obtained from cultures of C6 cells maintained under standard conditions, whereas other MVs were obtained after C6 cells received a single radiation dose of 50 Gy. The irradiated cells showed morphology changes compared with the control cells, and the irradiated cells produced more MVs (Figure 1A). The MVs were observed by transmission electron microscopy (TEM) in both groups of cells (irradiated and not irradiated); spherical particles approximately 350 nm in diameter were observed (Figure 1B).
Figure 1.

Photomicrographs of C6 Glioblastoma Cells and Microvesicles
(A) Clear-field microscopy images of C6 glioma cells (N) and irradiated C6 glioma cells (I) with a 50-Gy radiation dose. (B) Transmission electron microscopy images of non-irradiated C6 cell-derived microvesicles (MVs) and irradiated C6 cells derived MVs (IR-MVs) labeled with Annexin V-gold. The scale bars represent 300 nm.
To determine the size of the MVs, a nanoparticle tracking analysis (NTA) was performed. Figures 2A–2D show representative images of the analysis and diagrams of MV size distribution. The average size of the MVs was 395.0 ± 202.6 nm and 339.9 ± 139.0 nm for IR-MVs (Figure 2E).
Figure 2.

Size Determination of MVs
(A–D) Images of MV size distribution obtained from a video of the NanoSight nanoparticle tracking analysis [images taken from NanoSight videos of MVs (A) and IR-MVs (C)] and analysis of the MV size distribution from MVs (B) and IR-MVs (D) using the NTA 3.2 Dev Build 3.2.16 NanoSight software. (E) MV average size.
Radiation Induces Changes in the Protein Content of MVs from C6 Glioma Cells
To determine the characteristics of the MVs exposed to radiation, extraction of proteins from C6 glioma cells and MVs was carried out and analyzed by SDS-PAGE. Complete lysate from C6 glioblastoma cells had a greater amount of proteins, followed by IR-MVs and MVs; in addition, a different banding pattern was observed in three of them (Figure 3).
Figure 3.
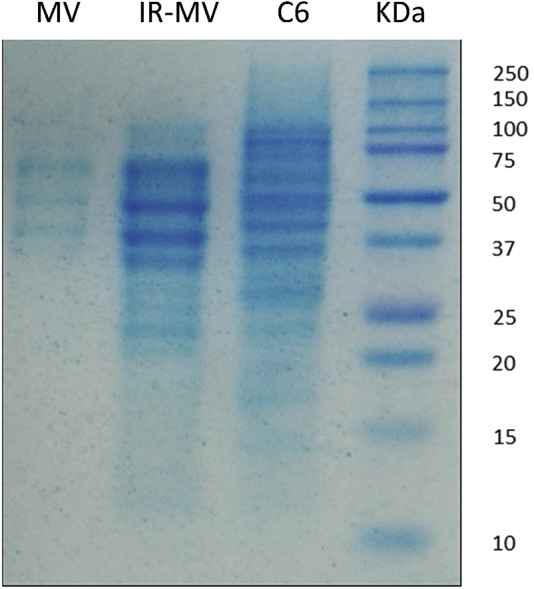
Protein Characterization of MVs
Shown is SDS-PAGE in 15% polyacrylamide gel of proteins obtained from MVs, IR-MVs, and C6 cells. KDa, molecular weight marker.
Vaccination with IR-MVs Reduces Tumor Volume
To determine whether therapeutic vaccination with MVs induces an immune response capable of reducing the tumor volume of rats implanted previously with glioma C6 cells, we administered MVs and IR-MVs to rats with a subcutaneous C6 glioma. We observed a more than 50% decrease in tumor volume in the group of rats treated with IR-MVs compared with the control and MV groups (p = 0.03). No significant changes were observed in the group of rats treated with MVs compared with the control group (Figure 4).
Figure 4.

Effect of IR-MVs on Tumor Growth Kinetics
One million viable C6 cells were implanted subcutaneously in Wistar rats. Tumor volume was determined on days 0, 7, 14, and 21 post-treatment in control rats and rats treated with MVs and IR-MVs. The results are expressed as the mean ± SEM. *p = 0.031, IR-MVs versus control.
Vaccination with IR-MVs Promotes an Antitumor Immune Response that Leads to the Death of Glioblastoma Cells by Apoptosis
To determine the mechanism of cell death induced by vaccination with MVs over the tumor cells, we determine the percentage of apoptotic and necrotic cells of tumors from rats treated with MVs or IR-MVs 21 days after vaccination (Figure 5). We found an increase of three times the percentage of apoptosis in tumor cells from rats treated with IR-MVs (p = 0.038) compared with MVs or controls.
Figure 5.

Evaluation of Apoptosis and Tumor Necrosis in Rats Treated with Irradiated and Non-IR-MVs
(A–E) Flow cytometry analysis of the percentage of viable cells (Annexin V−/PI−, A), early apoptosis (Annexin V+/PI−, B), late apoptosis (Annexin V+/PI+, C), necrotic cells (Annexin V−/PI+, D), and the total amount of apoptotic cells (E) was carried out in tumors of control group and in rats treated with MVs and IR-MVs. The results are expressed as the mean ± SEM. *p = 0.027 early apoptosis, *p = 0.022 late apoptosis, and *p = 0.038 total apoptosis for IR-MVs versus the control group. No significant differences were observed for necrosis.
Vaccination with IR-MVs Increases Helper and Cytotoxic T Lymphocyte Infiltration into the Tumor
Induction of the specific antitumor immune response is accompanied by changes in the percentage of T lymphocytes. We determined the percentage of lymphocytes in the blood, spleen, and tumor by flow cytometry. A significant increase of three times in the percentage of tumor-infiltrating CD4+ lymphocytes was seen in rats treated with IR-MVs with respect to the control (p = 0.036). No differences were observed in rats treated with MVs. No changes were observed in the percentage of CD4+ lymphocytes in the spleen or blood with respect to the controls or in any of the treated groups (Figure 6). Similarly, a significant increase of two times in the percentage of infiltrating CD8+ cells was observed in the group of rats treated with IR-MVs with respect to the controls (p = 0.036); no significant changes were seen in the group of rats treated with MVs. Also, no significant changes were observed in the percentage of CD8+ lymphocytes in the blood and spleen of immunized groups with respect to the control group (Figure 7).
Figure 6.

Comparative Study of Helper T Cells in the Blood, Spleen, and Tumors of Rats Treated with Irradiated and Non-IR-MVs
(A–C) Flow cytometry analysis of CD4+ cells in the blood (A), spleen (B), and tumors (C) after 21 days of treatment with MVs, IR-MVs, or PBS (control group). Results are expressed as the mean ± SEM. *p = 0.036, IR-MVs versus control.
Figure 7.

Comparative Study of Cytotoxic T Cells in the Blood, Spleen, and Tumors
(A–C) Flow cytometry analysis of CD8+ was carried out in the control group and in rats treated with MVs or IR-MVs. The results are expressed as the mean ± SEM. *p = 0.04, IR-MV versus control.
Regarding regulatory T cells (CD4+ CD25+ FoxP3+), a slight increase in the percentage (1.5%, p = 0.037) of these tumor-infiltrating cells was observed in the group of rats vaccinated with IR-MVs with respect to the control group. No differences were observed in the group treated with MVs or in the percentages of these cells in the blood (Figure 8). Similarly, the percentages of natural killer (NK) cells (NKR-P1) and monocytes and macrophages (CD68+) in the blood, spleen, and tumor infiltrates were not significantly different between the treated groups and the control group (data not shown).
Figure 8.

Comparative Study of Regulatory T Lymphocytes in the Blood and Tumors
(A and B) Flow cytometry analysis of CD4+/CD25+/FoxP3+ cells in the blood (A) and tumors (B) from control rats and rats treated with MVs or IR-MVs was performed. The results are expressed as the mean ± SEM. *p = 0.037, IR-MV versus control.
Discussion
Despite advances in the diagnosis and multimodal treatment of GBM, this tumor type continues to be largely refractory to current therapy, without significant progress in survival and quality of life of patients in the last two decades.22 Among brain neoplasms, glioblastoma continues to be a fatal disease, mainly because of the ability of tumor cells to modify their microenvironment and quickly adapt to adverse conditions through a high infiltrative capacity, neo-angiogenesis, induction of immunosuppression, and multi-resistance to current therapy.23 In this context, reprogramming is a mechanism that allows cells to adapt to changes in the microenvironment induced by radio- or chemotherapy.24 Several studies show that MVs from GBMs contain angiogenic proteins, several mRNAs (including those derived from mutant oncogenes), as well as microRNA and even genomic DNA,25 which make them able to suppress the immune system, increase tumor progression, promote invasiveness and metastasis, and confer multi-drug resistance.26, 27, 28, 29
Clinical studies in cancer patients have tested immunotherapy with EVs derived from neoplastic cells. In a phase I study of 15 patients with stage III/IV metastatic melanoma, EV production by autologous dendritic cells derived from monocytes loaded with the MAGE3 peptide has been demonstrated; subcutaneous administration was not accompanied by toxicity (higher than grade II).30 In another study, EVs from dendritic cells derived from monocytes loaded with lung cancer peptides were intradermally administered to 13 patients with advanced non-small-cell lung cancer (grade I–II toxicity); some patients experienced long-term stability of disease and activation of immune effectors.31 Dai et al.32 conducted a phase I clinical study in 40 patients with stage III and IV colon cancer who were treated with autologous EVs obtained from ascites fluid alone or in combination with granulocyte and monocyte colony-stimulating factor (GM-CSF); this therapy was safe and inducted specific cytotoxic T responses against the tumor, although no relevant therapeutic response was observed.32
Considering initial clinical experiences, we isolated MVs from non-radiated and irradiated C6 glioblastoma cells to investigate their immunogenic potential and determine whether subcutaneous vaccination with MVs from stressed tumoral cells could stimulate the immune system of experimental hosts and, as a consequence, reduce tumoral growth. It is important to note that this is the first study that attempted to use MVs from irradiated autologous glioma cells as a therapeutic vaccination. In a rat glioblastoma model, prophylactic vaccination with exosomes (containing MVs) induced immune system activation, stimulated humoral and cellular immunity, prevented implantation of the tumor, and a long-term memory response was obtained.9 However, the use of these exosomes for therapeutic immunization did not improve the survival of mice implanted with intracerebral glioblastoma despite the induction of both cellular and humoral immune responses. In this study, we observed a more than 50% reduction in tumor volume in rats vaccinated with IR-MVs, with no similar decrease in the group of rats vaccinated with MVs. Recent studies have shown that radiation induces a pattern of release of MVs as a response to this stress, which induces changes in cell metabolism and expression of genes that alter the phenotype of primary glioma cells.18 In the same way, the pattern of expression of antigens on MVs changes when the tumor cells are exposed to stress conditions such as ionizing radiation, promoting the generation of TAAs.18 In addition, it is known that tumor cells under a stressful stimulus express DAMPs such as heat shock proteins (HSPs), high mobility group box 1 (HMGB1) molecules, nucleotides, and uric acid, which are released together with the MV; these DAMPs are recognized by Toll-like receptors (TLRs) expressed on the surface of antigen-presenting cells (APCs), favoring their activation and maturation of immune cells.18, 19 DAMPs contained in IR-MVs facilitate the uptake of TAAs and their presentation via major histocompatibility complex (MHC) class 1 to tumor-specific cytotoxic T lymphocytes (CD8+). To determine the mechanism of cell death induced by MV vaccination, we determined the cell percentage of apoptosis and necrosis. The group of rats treated with IR-MVs showed a large increase (three times) of cells in apoptosis in comparison with rats treated with non-IR-MVs. Radiotherapy can directly induce the activation and regulation of antitumor immunity and promote immunogenic cell death.33, 34 Also, radiation induces the release of immunogenic MVs; after radiation, the tumor cells release DAMPS, particularly HSPs.35 These molecules can be recognized by cells of the innate immune system, favoring a specific antitumor immune response, promoting apoptosis of tumoral cells.36, 37 Our results show that IR-MVs could promote immunogenically mediated tumor cell death. To analyze whether treatment with IR-MVs modified immune effector cell populations, we measured the percentage of helper and cytotoxic T cells, NK cells, macrophages, and regulatory T cells in the blood, spleen, and tumor infiltrates. Our results show that rats carrying subcutaneous glioma tumors treated with IR-MVs had an increase in the percentage of tumor-infiltrating effector cells. Several studies have shown that an increase in infiltrating immune effector populations into the tumor correlates with a better prognosis.38, 39 We observed significant increases in CD4+ and CD8+ lymphocytes and a slight increase in regulatory T (Treg) cells in the tumors from animals immunized with IR-MVs. Theoretically, tumor cells express a set of antigens capable of stimulating an immune response by CD4+ and CD8+ T lymphocytes and favor their proliferation and migration to the tumor site.40 However, this does not always occur because most of the molecules contained in these MVs may also be immunosuppressive; thus, effective immunotherapy must stimulate the proliferation of antitumor effector cells with the ability to migrate to the tumor site and eliminate tumor cells effectively. According to our results, radiation promotes the release of MVs with a particular antigenic repertoire (Figure 3), which induces changes in protein content and provides an effective antitumor immune response, perhaps added by the release of TAAs. In this context, vaccination with IR-MVs favors the expansion of effector T cells with the capacity to migrate into the tumor and induce immunogenic cell death through apoptosis. It seems possible that radiation induces the release of MVs with a more extensive antigenic repertoire and expression of DAMPs that are recognized by the host’s immune system, leading to a more effective antitumor immune response.41, 42, 43 Our results warrant further studies of immunotherapy with IR-MVs as an immunotherapeutic approach. A possible application suggested by our group is to apply IR-MVs from human glioblastoma cell lines immediately after diagnosis (up to three times before surgical treatment, then continuing with the chemotherapy and radiation protocol). Currently, we are conducting new investigations, applying IR-MVs in combination with blockage of different immune checkpoints.
Conclusions
Given the relevance of MVs in intercellular communication and a potential role in the abscopal effect after tumoral radiotherapy, we showed in this study that immunization with IR-MVs could be a probable immunotherapy against malignant glioma. IR-MVs could be novel agents in therapeutic applications such as vaccination against tumors. The use of IR-MVs could be a therapeutic modality for the treatment of glioblastoma patients.
Materials and Methods
Animals
Male Wistar rats (n = 30, 230–250 g) from the animal house of the National Institute of Neurology (Mexico City) were employed throughout the study. Before they were used for the experiments, animals were housed five per cage in acrylic cages and provided with a standard commercial rat diet (Laboratory Rodent Diet 5001, PMI Feeds, Richmond, IN, USA) and water ad libitum. The housing room was maintained with constant temperature (25°C ± 3°C), humidity (50% ± 10%), and lighting (12 h light and dark cycles). All procedures with animals were carried out according to the NIH Guide for the Care and Use of Laboratory Animals and local guidelines for the ethical use of animals from the Health Ministry of Mexico. During dissection, all efforts were made to minimize animal suffering.
Cell Cultures
C6 glioma cells were acquired from the ATCC (Rockville, MD, USA). Cells were cultured under sterile conditions at 37°C in a humid atmosphere controlled with 5% CO2 in DMEM (Gibco) supplemented with 10% fetal bovine serum (FBS) (Gibco), 4 mM glutamine, 100 units/mL of penicillin, and 100 mg/mL streptomycin.
Obtention and Quantification of Glioma-Derived MVs
MVs were obtained from cultures of C6 cells either non-radiated or irradiated with 50 Gy by a linear accelerator TrueBeam Stx (Varian and Brainlab, USA). Seventy-two hours after radiation, the culture medium was collected and centrifuged at 200 × g to eliminate viable cells and cell debris. Afterward, the supernatant was centrifuged at 14,000 × g for 20 min to sediment the MVs, which were resuspended in sterile PBS. For quantification, MVs were labeled with Annexin V- fluorescein isothiocyanate (FITC) (Annexin-V-FLUOS Staining Kit, Roche), which has a high affinity for phosphatidylserine, and analyzed by flow cytometry (FACSCalibur, BD Biosciences), considering the acquisition speed (volume acquired and time) acquired in 30 s per sample, the number of events counted, and the percentage of positive events for Annexin V. For data analysis, Cell QuestPro (BD Bioscience, USA) and Flow Jo v.10 software were used.
Characterization of MVs
The size of the MVs obtained was examined using an NTA. A sample of the MVs obtained was placed on a Malvern NanoSight NS300 instrument (NanoSight, Amesbury, UK) equipped with a blue laser (488 nm) and a Complementary Metal Oxide Semiconductor (CMOS) camera, which allowed fast and automatic analysis of size distribution and concentration of nanoparticles, from 10 to 2,000 nm in diameter, according to the configuration of the instrument and the type of sample. Preparations were measured in triplicate (temperature, 22°C; viscosity, 0.95 centipose [cP]) for 10 s. The software used for capture and analysis of data was NTA 3.2 Dev Build 3.2.16.
Ultrastructural Analysis
Samples of 4 μL of MVs from C6 glioma cells not labeled and labeled previously with Annexin V coupled for 30 min to 15-nm colloidal gold particles (Biorbyt, USA) were centrifuged at 14,000 × g for 20 min at 4°C. The pellet obtained was resuspended in PBS. Then one drop was placed on nickel grids covered previously with a collodion (Parlodion) membrane (Electron Microscopy Sciences, Washington, PA, USA). The nickel grids were negatively stained with 3% uranyl acetate and observed at a magnification of 40,000× with a Jem 1400 Plus electronic TEM (Jeol, Akishima, Tokyo, Japan) at 60 kV.
Protein Contents
Protein extraction was performed with 1 × 106 C6 glioma cells, 106 MVs, and 106 IR-MVs using the ProteoJET cytoplasmic and nuclear protein extraction kit (Fermentas) and quantified by Lowry. The resultant content was analyzed using Bio-Rad protein-staining reagent. The protein contents of C6 glioblastoma cells and MVs from irradiated or non-irradiated C6 glioma cells were electrophoretically separated by 15% SDS-PAGE gels using the Precision Plus molecular weight marker (Bio-Rad, Paris). Gels were stained with Coomassie blue stain.
Malignant C6 Glioma in Rats
When C6 cell cultures reached confluence, cells were collected by trypsin treatment and resuspended in DMEM. 1 × 107 C6 cells in 500 μL of DMEM were inoculated intraperitoneally into a male Wistar rat to obtain an extensive amount of C6 cells for subcutaneous implantation of tumors. After 3 weeks, the peritoneal tumor was extracted and mechanically disaggregated; cells were resuspended in DMEM. A suspension of 1 × 107 viable cells in 500 μL of saline solution was subcutaneously inoculated into the left thigh of 12-week-old female Wistar rats. A subcutaneous tumor developed in 80% of the animals between 18 and 20 days after cell implantation; the tumors reached a diameter of 2 cm. At that size, no spontaneous involution was observed. Therefore, we only included rats that developed subcutaneous C6 gliomas of at least 2 cm in diameter.
Experimental Design in the C6 Glioma Model
Thirty rats with subcutaneous glioma were separated into three groups. The first group was injected with PBS (control group), the second group was treated with MVs, and the third group was treated with IR-MVs (1 × 108 MVs). Both PBS and treatments were emulsified with Freund adjuvant (1:1) and administered subcutaneously to the contralateral thigh of the tumor on days 0 and 7. Tumor growth was assessed once a week. Animals were sacrificed 21 days after the last injection with MVs by exsanguination (prior anesthesia with ketamine/xylazine). Tumor size was measured, and samples of tumor, blood, and spleen were collected for further analysis.
Determination of Tumor Volume
Tumors were measured weekly calibrated Vernier caliper; volume was calculated using the formula π/6 × length × width × height.
Macrophages, NK Cells, and T Lymphocyte Subpopulations
The percentage of helper (CD4+) and cytotoxic (CD8+) T lymphocytes, regulatory T cells (CD4+CD25+FoxP3+), NK killer cells (NKR-P1+), and monocytes and macrophages (CD68+) in blood, spleen, and tumor samples was determined by flow cytometry using anti-CD4-phycoerythrin (PE), anti-CD8-PE, anti-NKR-P1-FITC, anti-CD25-FITC (Biosource, Washington, USA), anti-Foxp3-APC (eBioscience, San Diego, CA), and anti-CD68 monoclonal antibodies labeled with a secondary antibody coupled to APC (Biosource, Washington, USA). Briefly, 30 μL of blood or homogenate of spleen or tumor was incubated with 5 μL of the monoclonal antibody (1:10 dilution) for 30 min. Subsequently, 200 μL of red blood cell lysis solution was added (BD Biosciences), incubated for 10 min, and washed with PBS. 200 μL of permeabilization solution was added (BD Biosciences), incubated for 10 min, washed, incubated with anti-FoxP3-APC for 30 min, and finally washed and fixed with 1% paraformaldehyde in PBS. Cells were analyzed by flow cytometry (FACSCalibur, BD Biosciences) using CellQuest Pro (BD Biosciences) and Flow Jo version 10.
Apoptosis and Necrosis Analysis
A portion of the tumor homogenate was used to evaluate apoptosis and necrosis. Cells were washed with PBS and stained with Annexin V and propidium iodide (PI) (Annexin-VFLUOS Staining Kit, Roche) in 100 μL of incubation buffer for 15 min in darkness at room temperature. Thereafter, 200 μL buffer was added and analyzed by flow cytometry (FACSCalibur, BD Biosciences) with Cell QuestPro (BD Biosciences) and Flow Jo version 10.
Statistical Analysis
The Shapiro-Wilk normality test was performed to determine parametric values. ANOVA was determined, followed by Tukey test. A value of p < 0.05 was considered significant. SPSS Statistic 23.0 software (IBM SPSS Statistics for Windows, version 23.0, Armonk, NY, USA) was used for statistical tests.
Author Contributions
B.P. conceived the experimental and theoretical design, performed data analysis, and wrote the manuscript. F.J.S.G. conceived the experimental and theoretical design and performed data analysis. N.K.O. performed cell culture and obtained, quantified, and experimentally characterized MVs. V.P.C. performed flow cytometry. A.S.R. designed the experimental C6 glioma model. S.M.-J. performed all radiation treatments. N.H.P. performed statistical analysis. A.M.N. analyzed the protein content of MVs. A.O.P. performed structural analysis of MVs. J.S. contributed to data analysis and drafting, writing, and critical revision of the paper. All authors reviewed, discussed, and agreed on the manuscript.
Conflict of Interests
The authors declare no competing interests.
Acknowledgments
This project was supported by CONACyT grant 226201. We thank Lic. Fernando Ramos for partial financial support. We thank Dra. Shantal Lizbeth Baltierra Uribe (core lab facilities, Immunology Department, ENCB-IPN) for help with the NanoSight instrument.
References
- 1.Ostrom Q.T., Gittleman H., Fulop J., Liu M., Blanda R., Kromer C., Wolinsky Y., Kruchko C., Barnholtz-Sloan J.S. CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2008-2012. Neuro-oncol. 2015;17(Suppl 4):iv1–iv62. doi: 10.1093/neuonc/nov189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Deorah S., Lynch C.F., Sibenaller Z.A., Ryken T.C. Trends in brain cancer incidence and survival in the United States: Surveillance, Epidemiology, and End Results Program, 1973 to 2001. Neurosurg. Focus. 2006;20:E1. doi: 10.3171/foc.2006.20.4.E1. [DOI] [PubMed] [Google Scholar]
- 3.Louis D.N., Perry A., Reifenberger G., von Deimling A., Figarella-Branger D., Cavenee W.K., Ohgaki H., Wiestler O.D., Kleihues P., Ellison D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: a summary. Acta Neuropathol. 2016;131:803–820. doi: 10.1007/s00401-016-1545-1. [DOI] [PubMed] [Google Scholar]
- 4.Stupp R., Mason W.P., van den Bent M.J., Weller M., Fisher B., Taphoorn M.J., Belanger K., Brandes A.A., Marosi C., Bogdahn U., European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups. National Cancer Institute of Canada Clinical Trials Group Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005;352:987–996. doi: 10.1056/NEJMoa043330. [DOI] [PubMed] [Google Scholar]
- 5.Kazda T., Dziacky A., Burkon P., Pospisil P., Slavik M., Rehak Z., Jancalek R., Slampa P., Slaby O., Lakomy R. Radiotherapy of Glioblastoma 15 Years after the Landmark Stupp’s Trial: More Controversies than Standards? Radiol. Oncol. 2018;52:121–128. doi: 10.2478/raon-2018-0023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mole R.H. Whole body irradiation; radiobiology or medicine? Br. J. Radiol. 1953;26:234–241. doi: 10.1259/0007-1285-26-305-234. [DOI] [PubMed] [Google Scholar]
- 7.Roger A., Finet A., Boru B., Beauchet A., Mazeron J.J., Otzmeguine Y., Blom A., Longvert C., de Maleissye M.F., Fort M. Efficacy of combined hypo-fractionated radiotherapy and anti-PD-1 monotherapy in difficult-to-treat advanced melanoma patients. OncoImmunology. 2018;7:e1442166. doi: 10.1080/2162402X.2018.1442166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Xu M.J., Wu S., Daud A.I., Yu S.S., Yom S.S. In-field and abscopal response after short-course radiation therapy in patients with metastatic Merkel cell carcinoma progressing on PD-1 checkpoint blockade: a case series. J. Immunother. Cancer. 2018;6:43. doi: 10.1186/s40425-018-0352-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Graner M.W., Alzate O., Dechkovskaia A.M., Keene J.D., Sampson J.H., Mitchell D.A., Bigner D.D. Proteomic and immunologic analyses of brain tumor exosomes. FASEB J. 2009;23:1541–1557. doi: 10.1096/fj.08-122184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lee Y., El Andaloussi S., Wood M.J. Exosomes and microvesicles: extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012;21(R1):R125–R134. doi: 10.1093/hmg/dds317. [DOI] [PubMed] [Google Scholar]
- 11.Cocucci E., Racchetti G., Meldolesi J. Shedding microvesicles: artefacts no more. Trends Cell Biol. 2009;19:43–51. doi: 10.1016/j.tcb.2008.11.003. [DOI] [PubMed] [Google Scholar]
- 12.Muralidharan-Chari V., Clancy J., Plou C., Romao M., Chavrier P., Raposo G., D’Souza-Schorey C. ARF6-regulated shedding of tumor cell-derived plasma membrane microvesicles. Curr. Biol. 2009;19:1875–1885. doi: 10.1016/j.cub.2009.09.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lima L.G., Chammas R., Monteiro R.Q., Moreira M.E., Barcinski M.A. Tumor-derived microvesicles modulate the establishment of metastatic melanoma in a phosphatidylserine-dependent manner. Cancer Lett. 2009;283:168–175. doi: 10.1016/j.canlet.2009.03.041. [DOI] [PubMed] [Google Scholar]
- 14.Millimaggi D., Mari M., D’Ascenzo S., Carosa E., Jannini E.A., Zucker S., Carta G., Pavan A., Dolo V. Tumor vesicle-associated CD147 modulates the angiogenic capability of endothelial cells. Neoplasia. 2007;9:349–357. doi: 10.1593/neo.07133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lee T.H., D’Asti E., Magnus N., Al-Nedawi K., Meehan B., Rak J. Microvesicles as mediators of intercellular communication in cancer--the emerging science of cellular ‘debris’. Semin. Immunopathol. 2011;33:455–467. doi: 10.1007/s00281-011-0250-3. [DOI] [PubMed] [Google Scholar]
- 16.Arscott W.T., Tandle A.T., Zhao S., Shabason J.E., Gordon I.K., Schlaff C.D., Zhang G., Tofilon P.J., Camphausen K.A. Ionizing radiation and glioblastoma exosomes: implications in tumor biology and cell migration. Transl. Oncol. 2013;6:638–648. doi: 10.1593/tlo.13640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ratajczak J., Miekus K., Kucia M., Zhang J., Reca R., Dvorak P., Ratajczak M.Z. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for horizontal transfer of mRNA and protein delivery. Leukemia. 2006;20:847–856. doi: 10.1038/sj.leu.2404132. [DOI] [PubMed] [Google Scholar]
- 18.Baulch J.E., Geidzinski E., Tran K.K., Yu L., Zhou Y.H., Limoli C.L. Irradiation of primary human gliomas triggers dynamic and aggressive survival responses involving microvesicle signaling. Environ. Mol. Mutagen. 2016;57:405–415. doi: 10.1002/em.21988. [DOI] [PubMed] [Google Scholar]
- 19.Hernández-Pedro N., Magana-Maldonado R., Ramiro A.S., Pérez-De la Cruz V., Rangel-López E., Sotelo J., Pineda B. PAMP-DAMPs interactions mediates development and progression of multiple sclerosis. Front. Biosci. (Schol. Ed.) 2016;8:13–28. doi: 10.2741/s443. [DOI] [PubMed] [Google Scholar]
- 20.Vénéreau E., Ceriotti C., Bianchi M.E. DAMPs from Cell Death to New Life. Front. Immunol. 2015;6:422. doi: 10.3389/fimmu.2015.00422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wang W., Zhang Y., Yang L., Li H. The innate immune signaling in cancer and cardiometabolic diseases: Friends or foes? Cancer Lett. 2017;387:46–60. doi: 10.1016/j.canlet.2016.06.004. [DOI] [PubMed] [Google Scholar]
- 22.Bernard-Arnoux F., Lamure M., Ducray F., Aulagner G., Honnorat J., Armoiry X. The cost-effectiveness of tumor-treating fields therapy in patients with newly diagnosed glioblastoma. Neuro-oncol. 2016;18:1129–1136. doi: 10.1093/neuonc/now102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Aldape K., Zadeh G., Mansouri S., Reifenberger G., von Deimling A. Glioblastoma: pathology, molecular mechanisms and markers. Acta Neuropathol. 2015;129:829–848. doi: 10.1007/s00401-015-1432-1. [DOI] [PubMed] [Google Scholar]
- 24.Kwon S.M., Kang S.H., Park C.K., Jung S., Park E.S., Lee J.S., Kim S.H., Woo H.G. Recurrent Glioblastomas Reveal Molecular Subtypes Associated with Mechanistic Implications of Drug-Resistance. PLoS ONE. 2015;10:e0140528. doi: 10.1371/journal.pone.0140528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Skog J., Würdinger T., van Rijn S., Meijer D.H., Gainche L., Sena-Esteves M., Curry W.T., Jr., Carter B.S., Krichevsky A.M., Breakefield X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008;10:1470–1476. doi: 10.1038/ncb1800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Janowska-Wieczorek A., Wysoczynski M., Kijowski J., Marquez-Curtis L., Machalinski B., Ratajczak J., Ratajczak M.Z. Microvesicles derived from activated platelets induce metastasis and angiogenesis in lung cancer. Int. J. Cancer. 2005;113:752–760. doi: 10.1002/ijc.20657. [DOI] [PubMed] [Google Scholar]
- 27.Liu C., Yu S., Zinn K., Wang J., Zhang L., Jia Y., Kappes J.C., Barnes S., Kimberly R.P., Grizzle W.E., Zhang H.G. Murine mammary carcinoma exosomes promote tumor growth by suppression of NK cell function. J. Immunol. 2006;176:1375–1385. doi: 10.4049/jimmunol.176.3.1375. [DOI] [PubMed] [Google Scholar]
- 28.Ginestra A., La Placa M.D., Saladino F., Cassarà D., Nagase H., Vittorelli M.L. The amount and proteolytic content of vesicles shed by human cancer cell lines correlates with their in vitro invasiveness. Anticancer Res. 1998;18(5A):3433–3437. [PubMed] [Google Scholar]
- 29.Clayton A., Mitchell J.P., Court J., Mason M.D., Tabi Z. Human tumor-derived exosomes selectively impair lymphocyte responses to interleukin-2. Cancer Res. 2007;67:7458–7466. doi: 10.1158/0008-5472.CAN-06-3456. [DOI] [PubMed] [Google Scholar]
- 30.Escudier B., Dorval T., Chaput N., André F., Caby M.P., Novault S., Flament C., Leboulaire C., Borg C., Amigorena S. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: results of the first phase I clinical trial. J. Transl. Med. 2005;3:10. doi: 10.1186/1479-5876-3-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Morse M.A., Garst J., Osada T., Khan S., Hobeika A., Clay T.M., Valente N., Shreeniwas R., Sutton M.A., Delcayre A. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J. Transl. Med. 2005;3:9. doi: 10.1186/1479-5876-3-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Dai S., Wei D., Wu Z., Zhou X., Wei X., Huang H., Li G. Phase I clinical trial of autologous ascites-derived exosomes combined with GM-CSF for colorectal cancer. Mol. Ther. 2008;16:782–790. doi: 10.1038/mt.2008.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Calzascia T., Masson F., Di Berardino-Besson W., Contassot E., Wilmotte R., Aurrand-Lions M., Rüegg C., Dietrich P.Y., Walker P.R. Homing phenotypes of tumor-specific CD8 T cells are predetermined at the tumor site by crosspresenting APCs. Immunity. 2005;22:175–184. doi: 10.1016/j.immuni.2004.12.008. [DOI] [PubMed] [Google Scholar]
- 34.Mantel F., Frey B., Haslinger S., Schildkopf P., Sieber R., Ott O.J., Lödermann B., Rödel F., Sauer R., Fietkau R., Gaipl U.S. Combination of ionising irradiation and hyperthermia activates programmed apoptotic and necrotic cell death pathways in human colorectal carcinoma cells. Strahlenther. Onkol. 2010;186:587–599. doi: 10.1007/s00066-010-2154-x. [DOI] [PubMed] [Google Scholar]
- 35.Paolini A., Pasi F., Facoetti A., Mazzini G., Corbella F., Di Liberto R., Nano R. Cell death forms and HSP70 expression in U87 cells after ionizing radiation and/or chemotherapy. Anticancer Res. 2011;31:3727–3731. [PubMed] [Google Scholar]
- 36.Garg A.D., Nowis D., Golab J., Vandenabeele P., Krysko D.V., Agostinis P. Immunogenic cell death, DAMPs and anticancer therapeutics: an emerging amalgamation. Biochim. Biophys. Acta. 2010;1805:53–71. doi: 10.1016/j.bbcan.2009.08.003. [DOI] [PubMed] [Google Scholar]
- 37.Sherman M., Multhoff G. Heat shock proteins in cancer. Ann. N Y Acad. Sci. 2007;1113:192–201. doi: 10.1196/annals.1391.030. [DOI] [PubMed] [Google Scholar]
- 38.Galon J., Angell H.K., Bedognetti D., Marincola F.M. The continuum of cancer immunosurveillance: prognostic, predictive, and mechanistic signatures. Immunity. 2013;39:11–26. doi: 10.1016/j.immuni.2013.07.008. [DOI] [PubMed] [Google Scholar]
- 39.Gajewski T.F., Woo S.R., Zha Y., Spaapen R., Zheng Y., Corrales L., Spranger S. Cancer immunotherapy strategies based on overcoming barriers within the tumor microenvironment. Curr. Opin. Immunol. 2013;25:268–276. doi: 10.1016/j.coi.2013.02.009. [DOI] [PubMed] [Google Scholar]
- 40.Schreiber R.D., Old L.J., Smyth M.J. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science. 2011;331:1565–1570. doi: 10.1126/science.1203486. [DOI] [PubMed] [Google Scholar]
- 41.Evans S.M., Putt M., Yang X.Y., Lustig R.A., Martinez-Lage M., Williams D., Desai A., Wolf R., Brem S., Koch C.J. Initial evidence that blood-borne microvesicles are biomarkers for recurrence and survival in newly diagnosed glioblastoma patients. J. Neurooncol. 2016;127:391–400. doi: 10.1007/s11060-015-2051-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lener T., Gimona M., Aigner L., Börger V., Buzas E., Camussi G., Chaput N., Chatterjee D., Court F.A., Del Portillo H.A. Applying extracellular vesicles based therapeutics in clinical trials - an ISEV position paper. J. Extracell. Vesicles. 2015;4:30087. doi: 10.3402/jev.v4.30087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Quesenberry P.J., Goldberg L.R., Aliotta J.M., Dooner M.S., Pereira M.G., Wen S., Camussi G. Cellular phenotype and extracellular vesicles: basic and clinical considerations. Stem Cells Dev. 2014;23:1429–1436. doi: 10.1089/scd.2013.0594. [DOI] [PMC free article] [PubMed] [Google Scholar]