

Abstract
Microbiomes impact nearly every environment on Earth by modulating the molecular composition of the environment. Temporally changing environmental stimuli and spatial organization are major variables shaping the structure and function of microbiomes. The web of interactions among members of these communities and between the organisms and the environment dictates microbiome functions. Microbial interactions are major drivers of microbiomes and are modulated by spatiotemporal parameters. A mechanistic and quantitative understanding of ecological, molecular, and environmental forces shaping microbiomes could inform strategies to control microbiome dynamics and functions. Major challenges for harnessing the potential of microbiomes for diverse applications include the development of predictive modeling frameworks and tools for precise manipulation of microbiome behaviors.
Graphical abstract
1. MICROBIOMES ARE DYNAMIC NETWORKS
Microbiomes are collections of microorganisms (bacteria, archaea, eukaryotic microbes, and viruses) that occupy and critically impact all environments on Earth by performing diverse biochemical transformations. Microbiomes have been shown to drive biogeochemical cycles;1 enhance agricultural productivity;2,3 remediate environmental pollution;4,5 and impact human health,6 nutrition,7 behavior,8 and development.9 Microbiomes can also perform metabolic activities that cannot be performed by a single monospecies population. In syntrophic communities, microbes cooperate to perform degradation reactions that would be thermodynamically unfavorable for a single organism.10 Microbial diversity is a critical variable shaping environmental states. Indeed, a reduction in the taxonomic diversity of microbiomes is associated with human gut microbiome dysbiosis,11–13 loss of resistance to invasion by pathogens,14,15 and a decreased level of organic matter deposition and nutrient cycling in soil and oceans.16,17
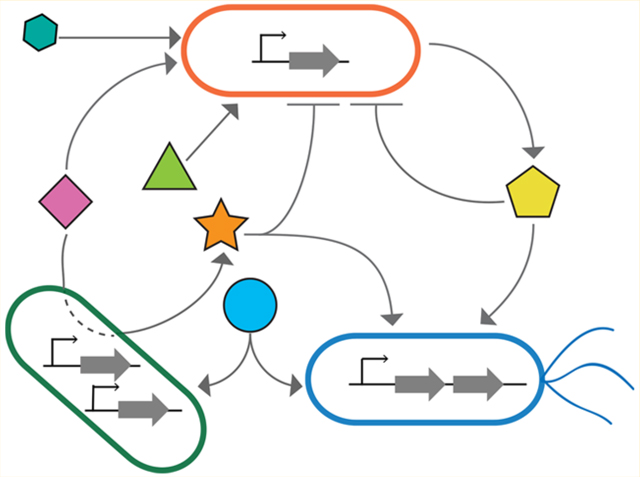
Microbiomes are highly complex and can contain hundreds to thousands of distinct species. These dense and inter-connected networks change as a function of time and spatial organization18,19 (Figure 1a,b). Environmental factors (physical and chemical parameters) and microbe—microbe interactions combine to shape the properties of microbiomes. In lakes and oceans, microbial community composition is driven by both seasonal and longer-term climate patterns.20 In the human gut, dietary substrates and drugs modify the growth and biochemical activities of human gut microbiome species.21–24 In both soils and oceans, climate change has affected microbe—microbe and plant–microbe interactions, contributing to major shifts in the patterns of organism abundance and diversity.25,26
Figure 1.
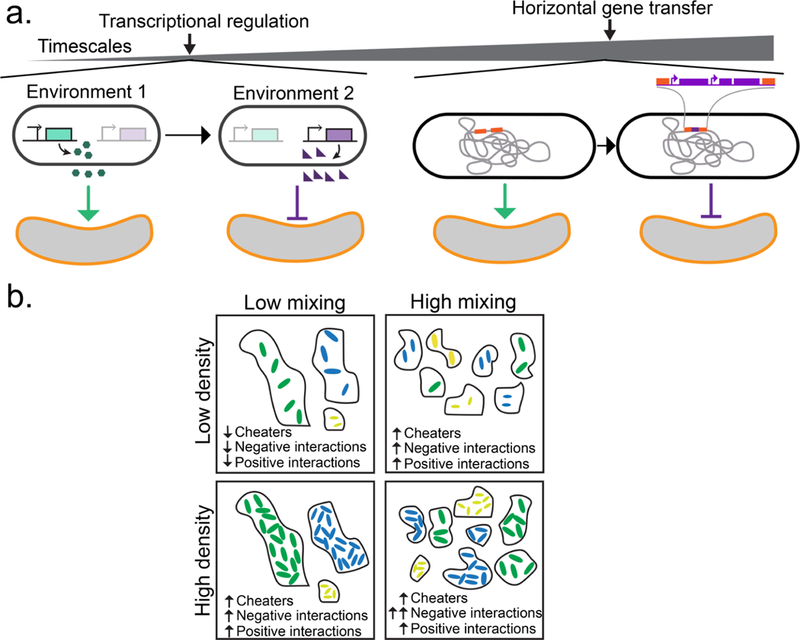
Interactions among organisms within microbiomes change through time and space. (a) Changes in intracellular network activities or changes in genotype via random mutation and/or horizontal gene transfer (HGT) can lead to changes in microbial interactions in microbiomes. Left: On short time scales, regulatory networks govern how microbes respond to their environment. In one environment, one microbe may produce molecules that lead to a net positive impact on a second microbe (green circles and green arrow), while in a second environment the same microbe may produce molecules that lead to a net negative impact on a second microbe (purple triangles and purple arrow) due to changes in transcriptional activity. Right: On longer time scales, genetic changes due to HGT can alter microbial intracellular networks and contribute to changes in ecological relationships. In natural competence (one mechanism of HGT), extracellular DNA can be taken up by a recipient organism and integrated into the chromosome via homologous recombination. When this occurs, the functional activities of a microbe can change, manifesting as a change in the ecological network from a net positive (green arrow) to a net negative interaction (purple arrow). (b) Schematic highlighting a hypothesis about how spatial mixing (x-axis) and population density (y-axis) can alter the frequency of cheating (wherein the benefit from cooperating is disproportional to the contribution toward cooperation) and the distributions of positive/negative interactions in a microbial community. Downward arrows indicate a low frequency, and upward arrows indicate a high frequency. When population densities and spatial mixing are low, microbes are less likely to encounter one another, leading to infrequent interactions and a low frequency of cheating due to low concentrations of public goods (“low density, low mixing” box). As population densities and/or spatial mixing increase, both the frequency of interactions and frequency of cheating increase (“low density, high mixing” and “high density, low mixing” boxes). In well-mixed, dense communities, negative interactions will be especially frequent because of resource competition (“high density, high mixing” box).
Interactions among organisms in microbiomes influence cell growth and biochemical activities, manifesting as positive and negative ecological relationships. Microbial interactions can originate from uptake or secretion of molecules27,28 or physical contact.29–32 The biochemical transformations performed by microbial communities are realized by biomolecular networks distributed among constituent members of the community. For example, the human gut microbiome operates as an anaerobic trophic web wherein metabolic activities are divided among distinct microbial guilds (groups of organisms that exploit the same or related resources). In this system, some organisms degrade complex carbohydrates to simpler sugars, which are fermented by other members of the ecosystem to form short-chain fatty acids.33 The fermentation products are utilized as substrates by acetogens, methanogens, and sulfate-reducing bacteria.33,34 Distributed metabolism is a feature of engineered microbiomes as well. In engineered wastewater treatment systems, well-defined guilds are responsible for each step of the nitrogen removal process.35
Microbiomes hold significant promise as systems that can be manipulated or designed to address global challenges in medicine, bioenergy, and agriculture.5 Humans have long used ecological36,37 and evolutionary38,39 processes to manipulate microbiomes. However, we currently lack the mechanistic understanding, engineering tools, and predictive modeling approaches necessary to rationally modify the properties of microbial communities and microbiomes. To harness the potential of microbiomes, a major challenge is elucidating the organizational principles of ecological networks that underlie microbiome behaviors. Computational models at different resolutions could enable prediction of microbiome dynamics and functions and inform control strategies. Tools from synthetic biology hold promise to control microbiome activities or design synthetic communities from the bottom up.40,41
2. MICROBIAL INTERACTIONS SHAPE MICROBIOME BEHAVIORS AND ARE MODULATED BY SPATIOTEMPORAL PARAMETERS
2.1. Defining Microbial Interactions.
An ecological interaction is defined as the net impact of an organism on the fitness and/or functional activities of a different organism over a given time period. Interactions can be positive (+), negative (−), or negligible (0) and emerge from the cumulative effect of molecules released into the environment and/or physical contact with other microbes as they vary through space and time. For a pair of organisms, there are six types of bidirectional interaction networks: commensalism (0/+), competition (−/−), predation or parasitism (−/+), cooperation (+/+), amensalism (0/−), or no interaction (0/0).42 The distribution of these pairwise motifs within an interaction network have been shown to influence microbiome stability.43,44
Positive interactions can arise when one organism secretes metabolites that can be utilized as a substrate by another45 or removes molecules that inhibit another’s growth (such as through degradation of antibiotics46). Negative interactions can arise because of resource competition,47 production of antimicrobial substrates,48 or toxic metabolic byproducts.49 In multispecies communities, pairwise interactions can be altered by other members of the community, representing a pairwise interaction that changes as a function of community context.50 For example, a negative pairwise interaction mediated by antibiotic production can be weakened by a third organism that exhibits antibiotic-degrading activity.51 Nevertheless, pairwise interactions are major parameters shaping the dynamics of multispecies consortia. For example, a recent study demonstrated that time-resolved measurements of single-organism growth and all pairwise communities could be used to train a dynamic model that accurately predicted the assembly of a 12-member human gut microbiome community and all single-species dropouts (all 11-member consortia) in vitro.44
2.2. Changes in Microbial Interactions Across Physiological or Evolutionary Time Scales.
Microbial interactions are context-dependent and can change as a function of environmental inputs and spatial positioning52,53 (Figure 1). On shorter time scales, microbes respond to the environment by sensing specific environmental signals and adjusting intracellular network activities accordingly. A microbe’s internal state can be altered in response to abiotic or biotic (biomolecules produced by living organisms) parameters. Organisms grown in coculture have been shown to activate biosynthetic gene clusters for secondary metabolite production that are otherwise silent in monoculture.54,55 The response of an organism to coculture growth can be quite specific: one study showed that Streptomyces coelicolor produced a unique set of molecules in coculture with each of five different actinomycetes, as revealed by mass spectrometry imaging of S. coelicolor colonies growing near each actinomycete species.56 Changes in internal network activities can also alter microbial interactions and community-level functions. For example, S. cerevisiae was shown to secrete amino acids in a nitrogen-rich environment, allowing Lactobacillus plantarum and Lactococcus lactis to grow by utilizing these subtrates.57 While these studies have illuminated changes in intracellular network activities in simple microbial communities, the network-level mechanisms through which biotic and abiotic factors combine to alter community-level functions in more complex microbiomes remain less understood.
On longer time scales, microbes can acquire and lose functional activities via horizontal gene transfer (HGT) and/or random (vertical) mutations. Evolutionary adaptations to grow and survive in microbial communities have impacted the acquisition and loss of microbial traits, including secondary metabolites such as antibiotics and signaling molecules.58,59 In particular, the evolution of antibiotic production has been shown to occur via both horizontal and vertical mutation. In an instance of horizontal gene transfer, genomic analysis showed that, when evolved in coculture with Streptomyces padanus, Rhodococcus fascians acquired a large pathway from S. padanus for the production of rhodostreptomycins,60 a class of antibiotics targeting a set of Gram-positive and Gram-negative bacteria. In an instance of vertical mutation, Streptomyces clavuligerus evolved to produce an antibiotic compound holomycin in the presence of Staphylococcus aureus61 because of several mutations affecting secondary metabolite biosynthesis.
Living in microbial communities also enables evolutionary adaptations that are not possible for organisms that live in isolation. These adaptations include (i) cross-feeding, where one species provides a molecule such as nutrient or cofactor that enables a second species to grow or survive;62,63 (ii) syntrophy, or obligate mutualism, wherein each organism performs a biochemical function essential for survival of a partner or community;64 (iii) division of labor, where different organisms specialize in specific tasks that are integrated at the community-level such as secreting virulence factors for elimination of competitive organisms,65 metabolizing available substrates,66 or biofilm formation;67 and (iv) Black Queen evolution (also called genome streamlining) where mutants lose “leaky” nonrequired functions (e.g., those that produce public goods or compounds that can benefit the community, such as extracellular cellulase) and rely on other community members to perform those functions.68
In turn, these adaptations can significantly change community-level functions, stability, diversity, and response to disturbances. For example, loss-of-function mutations have been shown to promote diversity in iron-limited communities.69 In iron-limiting conditions, some organisms produce energetically costly siderophores to scavenge iron from their environment. In these conditions, the siderophore-producer Pseudomonas aeruginosa outcompetes Burkholderia cenocepacia.69 However, when P. aeruginosa cheaters (those which do not produce siderophores but can scavenge them) are introduced into the population, competition between P. aeruginosa cheaters and noncheaters reduces the fitness advantage of noncheating P. aeruginosa, enabling coexistence with B. cenocepacia.69 A second study revealed the coexistence of cheater and noncheater P. aeruginosa populations in natural communities, suggesting cheating shapes the diversity of natural communities as well.70 In addition, community complexity can greatly alter adaptive evolution in ways that cannot be predicted from single-species evolution. For example, species in a synthetic community evolved to use waste products generated by other species, an evolutionary adaptation that may be less accessible for the same organisms grown in isolation.71
2.3. Spatial Organization Impacts Microbial Community Properties.
The spatial distance separating microbial populations is a major variable shaping the biochemical transformations that can be realized by microbiomes, as well as the metabolic efficiency of these processes (for example, in biofilms72). Spatial structuring at multiple scales influences the functions and stability of microbiomes. For example, in a mouse gastrointestinal tract, the spatial distribution of organisms in a synthetic human gut microbiome community exhibited distinct patterns at different length scales.73 On micrometer scales, species were intermixed, whereas spatial positioning was heterogeneous over longer length scales. These spatial distributions can significantly influence the functional capabilities of the gut microbiota.73 The density of cells and the level of mixing of different organisms in the environment can alter the frequency and strength of interspecies interactions (Figure 1b),53,74 and interspecies interactions may alter the spatial organization of microbiomes.75
Spatial organization can also be used to alter the outcome of long-term community dynamics. For example, spatial organization and low resource availability can prevent cheaters from dominating in the community.76,77 The precise mapping between the degree of spatial organization and community-level properties such as stability, diversity, and resilience depends on specific molecular and ecological mechanisms. For example, community stability was maximized in a synthetic consortium of soil isolates when the constituent community members were spatially separated by intermediate distances.53 Such optimal distance responses can arise because of trade-offs between positive or negative interactions that vary as a function of spatial separation.53 Exploiting this principle, bacterial signaling circuits programmed to activate only at intermediate distances from a signal sender population can generate spatial patterns.78 The quantitative contributions of spatial parameters and temporally changing inputs to microbial community behaviors can be studied using microfabrication techniques.79
3. COMPUTATIONAL MODELING OF MICROBIAL COMMUNITIES
Developing the capability to predict the response of micro-biomes to environmental inputs will allow rational control and bottom-up design of microbial communities. This section will describe modeling approaches for prediction of microbiome behaviors at different temporal scales and levels of biological organization (Figure 2).
Figure 2.
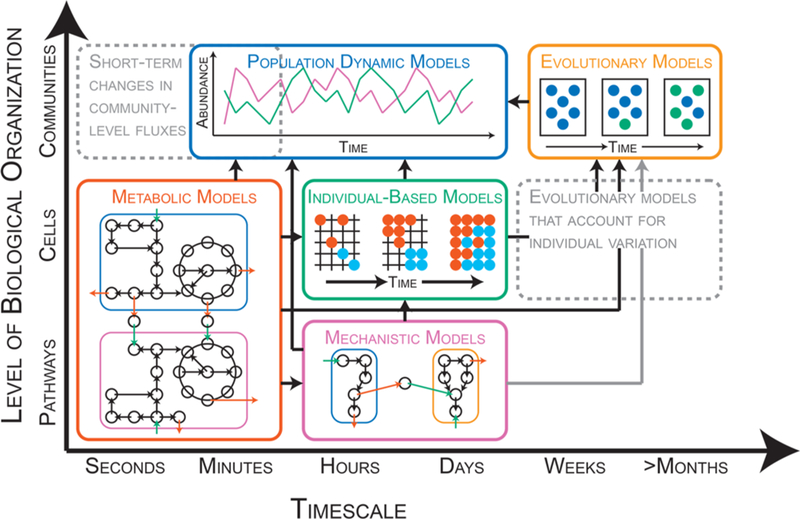
Schematic representation of modeling frameworks for processes occurring at different temporal (x-axis) and spatial (y-axis) scales. Metabolic, population-dynamic, individual-based, mechanistic, and evolutionary models are represented by the red, blue, green, purple, and orange boxes, respectively. Dashed gray boxes indicate temporal/spatial scales where modeling frameworks are currently missing. Black arrows between boxes indicate existence of publications that link modeling frameworks. Gray arrows between boxes indicate opportunities to link modeling frameworks. Metabolic models: Steady-state flux of metabolites (black circles) in and out (green arrows and red arrows, respectively) of two cell populations (blue and purple boxes) as predicted by a genome-scale metabolic network (black circles and arrows). Population-dynamic models: Change in abundance of two populations (green and purple) over time. Individual-based models: Emergence of spatial partitioning of two populations of cells (red and blue circles) on a discrete lattice, a structured arrangement of points representing a surface (black gridlines). Mechanistic models: Dynamic flux of metabolites (black circles) in and out (green arrows and red arrows, respectively) of two cell populations (blue and orange boxes) as predicted by a model of a biochemical pathway (black circles and arrows). Evolutionary models: Change in community population structure through evolutionary time. Initially, the population is homogeneous (blue circles, left-most rectangle). One cell acquires a mutation that provides a fitness advantage (green circle, middle rectangle). As a result, the community population structure changes over time (green and blue circles, right-most rectangle).
3.1. Genome-Scale Metabolic Modeling.
Genome-scale models (GEMs) can be used to predict mechanisms of metabolic interaction (Figure 2, red box).80 From a mathematical representation of an organism’s metabolic reactions, these models predict the rates and yields of metabolites consumed and secreted by each monospecies, as well as between the community and the environment.81,82 GEMs are analyzed using techniques from mathematical optimization, in which physiochemical constraints and optimality principles (i.e., maximization of an objective function, such as total community biomass) are applied to predict rates and yields.82 Although GEMs assume an intracellular steady state, they can still be used for predicting temporal dynamics, provided the steady-state assumption holds.83 These dynamic models predict metabolite and biomass concentrations, commonly by embedding steady-state models inside kinetic models of cellular growth and uptake.83 Steady-state models have been extensively used to simulate microbe—microbe and host-microbe interactions.82,84–90 Dynamic methods have been used to study interactions in the subsurface,91,92 as well as to predict emergent biosynthetic capacity,93 defined here as community production of a metabolite not produced by any of its constituent organisms growing alone.93 For example, a model of competition between the acetate-oxidizing metal reducers Rhodoferax and Geobacter correctly predicted the relative abundance of these two species under different geochemical conditions and forecasted that Geobacter is responsible for enhanced uranium reduction under conditions of low organic nitrogen.91
Despite their widespread adoption and accessibility,94,95 GEMs have a number of limitations. First, GEMs do not include regulatory information, and predicted metabolic states may not be achievable because of cellular regulation. Nonetheless, methods for incorporating regulatory networks into GEMs are emerging.96 Second, methods to analyze GEMs have an implicit requirement of evolutionary optimality.97 Optimal strategies may be unknown, change through time, or trade off with one another.98 In some cases, evolutionary optimality may not hold at all, such as in synthetic consortia composed of organisms that have not evolved in the same environment and thus have not encountered one another. Third, GEMs have many degrees of freedom and may require knowledge of the microbiome interaction network for accurate predictions.88,99 For dynamic GEMs, knowledge of growth-limiting substrates and their kinetic parameters may also be required for accurate predictions.100
3.2. Differential Equation Modeling.
Differential equation models are a broad class of models that can be applied to predict population dynamics, multisubstrate utilization, and/or intracellular network activities in microbial communities. Generalized Lotka–Volterra (gLV) models are commonly used to capture community dynamics (Figure 2, blue box). In gLV models, the abundance of an organism is a function of its intrinsic growth rate plus a term accounting for the sum of pairwise interactions with all members of the community, including self-interactions.101,102 Using techniques from dynamical systems theory, gLV models can also be used to evaluate community properties such as stability (the rate at which a community returns to an equilibrium point following a disturbance).103 Apart from interspecies interactions, gLV models have been used to simulate diverse aspects of microbial community behaviors, including assembly,104 stability,105 coexistence,44 and history-dependent dynamics.44 gLV equations have also been extended to include environmental inputs, including antibiotic administration105 and temperature and nutrient concentration.106
Despite their utility, gLV models have several limitations. First, microbes can interact both positively and negatively, and these interactions can change through both space and time. In a gLV model, the single, fixed interaction term represents the inferred cumulative impact across all of these mechanisms over a period of time. Second, some studies have shown that pairwise interactions are insufficient to fully explain community dynamics,107 meaning that gLV models may need to be extended to include higher-order interactions. Furthermore, standard gLV models may require a mechanistic representation of metabolite exchange to capture some types of pairwise interactions.107,108
Mechanistic differential equation models provide an alternative approach to overcoming some of the limitations of gLV models (Figure 2, purple box). In a mechanistic model, molecular-level details of the interaction are explicitly represented, and such models have been used to model 3-and 4-member communities with known mechanisms of molecular interaction.107,109,110 However, because of a need to characterize the mechanisms of interspecies interaction, determine the appropriate model structure, and identify model parameters, mechanistic models are significantly more difficult to develop than gLV models. As a result, mechanistic models typically describe only a few metabolic reactions performed by a small number of organisms, although some genome-scale mechanistic models have been developed.111–113
3.3. Modeling Cellular Heterogeneity Using Individual-Based Models.
Heterogeneity within microbial communities can arise from variation in microenvironments and/or variation within individuals. To capture spatial heterogeneity, partial differential equation models (PDEs) are used to model the underlying processes, such as heat and mass transfer.114 Heterogeneity within populations can be modeled using individual-based models (iBMs), which represent the activities of individual cells (or populations of cells) (Figure 2, green box). In these frameworks, system-level properties emerge from the combined activities of individuals.115,116 In many environments (such as biofilms117), microbial communities exhibit both spatial and individual heterogeneity, and PDEs and IBMs must be combined to model community behaviors.116
Recently, GEMs have been combined with IBMs. Using this approach, GEMs are used to simulate cells or populations on a lattice (a structured arrangement of points representing a surface),118–120 with differential equations describing molecular diffusion on the lattice. These models have been used to study the emergence of competition and cooperation in 2-or 3-member consortia,118 niche formation in the human microbiome,119 and spatial variation in colony size.121 A mechanistic, individual-based model of the cyanobacteria Anabaena has also been developed, wherein nitrogen metabolism within each cell was mechanistically modeled via differential equations.122
As with other modeling approaches, both PDEs and IBMs have limitations.115,116 PDE models typically do not account for phenotypic heterogeneity within members of a population.115,116 For example, the nonlinear Droop equation describes the growth rate of phytoplankton as a function of their intracellular phosphorus (P) content. Because of this nonlinearity, the growth rate of a population with average P content is greater than the average growth rate of the individual cells with a distribution of P contents.123 In addition, differential equation models assume that populations of discrete individuals can be approximated using continuous functions.115,116 In contrast, IBMs explicitly model individual organisms, enabling them to capture population heterogeneity as well as interactions between individual cells and between individual cells and their local environment.115,116 However, IBMs may result in complex and nonlinear dynamics that make parameter estimation difficult.116 Finally, integration of PDE models and IBMs poses its own challenges, such as representing processes occurring on different spatial or temporal scales.
3.4. Game-Theoretical Evolutionary Models.
The long-term outcome of changing interactions in microbial communities can be analyzed via evolutionary game theory (Figure 2, orange box).124 In this modeling framework, evolving entities (players) alter their reproductive fitness by adopting life-style strategies, or adaptations (such as cheating).124 These adaptations alter the fitness of the players, in a manner that depends on the behavior of other players in the game, as well as their relative abundance in the population.124 Apart from simulating evolutionary dynamics, evolutionary game theory can also be used to identify evolutionarily stable strategies, defined as combinations of strategies that are resistant to invasion (that is, there are no other strategies that would provide increased fitness if adopted). Additionally, evolutionary games can be transformed into gLV models,124 enabling techniques from dynamical systems theory to be applied to microbial communities on evolutionary time scales.
Evolutionary game theory was able to explain snowdrift dynamics, defined as the stable coexistence of cheaters and cooperators in a population.125 In a second example, game theory was combined with GEMs to predict evolutionarily stable interactions mediated by the exchange of specific amino acids.126 Finally, IBMs have been combined with trait-based models, in which fitness emerges from trade-offs among traits (measurable aspects of an organism’s phenotype, such as cell size or temperature sensitivity).127 Such models have been used to study the factors shaping cell size in oceanic phytoplankton128 and to highlight the importance of interspecies interactions in evolutionary outcomes.129
Despite these successes, evolutionary game theoretical models have a number of significant limitations. Using these models requires a priori knowledge of potential evolutionary strategies and their context-dependent effects on fitness.124 Furthermore, because ecological strategies and their effects on fitness are determined in advance, evolutionary game theory cannot predict the adoption of novel strategies or predict how their fitness changes in the face of a dynamic environment. Additionally, the probability that a particular strategy arises depends on the mechanism of adaptation, suggesting that evolutionary models will benefit from incorporating mechanistic information.130 Finally, evolutionary games with two players and two strategies have been studied in great detail,131 whereas microbial communities contain numerous organisms and strategies. This additional complexity can result in a breakdown of classical results from two-player games.131
4. MANIPULATING THE FUNCTIONS OF NATURAL AND SYNTHETIC COMMUNITIES
Traditional approaches for modifying the properties of microbial communities involve iterative evolution by selecting for specific phenotypes.132 Microbial communities used for wastewater treatment are a classical example: starting with a sample containing the organisms of interest, reactor engineering and environmental selection are used to enrich for strains with the desired traits,39 and metagenomic assembly/binning and metatranscriptomic profiling can be used to identify organisms potentially responsible for a function.133 However, selection-based approaches have notable limitations, including challenges in reverse engineering the mechanisms underlying changes in target function activity.134,135 In addition, because evolutionary optimization requires selection on existing traits, selection-based approaches cannot install novel capabilities. Here, we discuss how tools from synthetic biology hold promise to manipulate microbial communities or design microbial communities from the bottom up.
4.1. Harnessing Synthetic Biology to Manipulate Microbial Communities.
A core challenge in microbiome engineering is precisely altering the abundance of specific microbial populations to steer communities to desired states. Methods based on RNA-guided nucleases have been used to target specific organisms in multispecies communities. For example, CRISPR-Cas circuits delivered by bacteriophage or conjugation were used to target DNA sequences associated with antibiotic resistance or virulence, leading to selective killing of strains harboring these sequences.136,137 The host range of bacteriophage has also been modified for population editing using a modular tail fiber swapping method138 (Figure 3a). A third approach involves engineering controller organisms to selectively sense a pathogen and produce and deliver antimicrobials that inhibit the growth of the pathogen. This strategy has been used to sense and target pathogens including Salmonella, Pseudomonas aeruginosa, and Vibrio cholerae. 139–142 In CRISPR-Cas-based methods, efficient delivery and expression of the circuit components in target organisms can present challenges. Challenges in phage engineering include high mutation rates and programming phage host range to target specific populations.143 Finally, engineering a controller strain to sense and target specific organisms has a number of challenges, including ensuring the ecological and functional stability of the organism in the community for a period of time, developing tools for selective sensing, and targeting of specific organisms in the community.144,145
Figure 3.
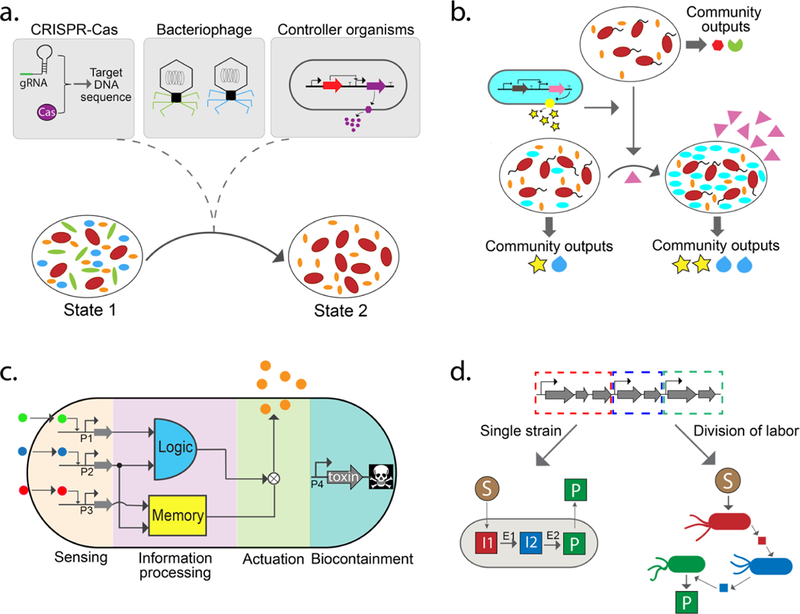
Strategies for engineering natural and synthetic communities. (a) Precise inhibition of specific organisms in natural communities via (i) CRISPR-guided nucleases that target specific DNA sequences, (ii) engineered bacteriophages, and (iii) microbes designed for production of antimicrobial compounds. (b) Introduction of an engineered organism (cyan) into a microbial community. The engineered organism can stably integrate into the community and alters community-level outputs (represented by yellow stars and blue teardrops). The abundance of the new species could be controlled through niche engineering, wherein a non-native organism is engineered to utilize a unique substrate (pink triangles) that cannot be utilized by other members of the community. (c) Design of cellular control organisms using engineered biomolecular networks to perform novel functions in complex environments. Functional modules include environmental sensors, information processing (e.g., memory and logic circuits), actuators (e.g., antimicrobial compounds), and growth control circuits for biocontainment. (d) Construction of synthetic communities from the bottom up to perform desired functions through division of labor (DOL). Top: A single biochemical pathway can be conceptually decomposed into three subpathways (red, blue, and green). Left arrow: The biochemical pathway for synthesizing a target molecule can be introduced into a single cell population. Right arrow: The biochemical pathway is partitioned among different organisms, reducing the metabolic burden on individual populations but requiring stable coexistence as a function of time. S, substrate; P, product; I, intermediate; E, enzyme.
The introduction of non-native strains (such as probiotics) to microbiomes also presents several challenges. The ecological and functional stability of a non-native strain is dependent on regulatory adaptation to the target environment, ability to secure a niche, and could potentially destabilize the resident community.146 For example, a recent human trial showed a strong negative association between colonization ability of Bifidobacterium longum AH1206 and the abundance of other B. longum strains and specific carbohydrate utilization genes that contribute to the ability of B. longum AH1206 to secure a niche in this environment.147
An engineered organism could be introduced into a microbial community to alter community-level outputs (Figure 3b). The abundance of the engineered organism could be modulated by an external parameter, such as nutrient availability. Engineering unique niches could allow stable integration of non-native organisms in microbiomes. For example, porphyran-degrading Bacteroides species were able to colonize mice fed a diet containing porphryan,148,149 and the abundance of these organisms was altered by administering different concentrations of porphyran or a pulsed diet. These studies also show that abundance of porphyran-degrading Bacteroides could be more precisely controlled in mice with fewer species in their microbiota, suggesting that community context can affect the controllability of species.148,149
An alternative to niche engineering involves genetic manipulation of the biochemical activities of resident organisms of microbiomes. For example, the microbiome-produced metabolite indolepropionic acid (IPA) is present in human serum and is associated with reduction of intestinal permeability, radical-scavenging, and neuroprotective functions.150 When a mutant symbiont Clostridium sporogenes deficient in IPA production was introduced into mice colonized with a defined, IPA-deficient microbial community, IPA levels in serum were not detected and intestinal permeability was increased compared to control mice harboring wild-type C. sporogenes.150
Finally, tools from synthetic biology can be used to engineer microbiomes by introducing novel functions. When engineering principles are applied, organisms can be designed to sense specific environmental signals (e.g., pH, temperature, and light), perform computation on these signals, and produce specific output responses151,152 (Figure 3c). Harnessing synthetic biology approaches to manipulate microbiomes presents several challenges. First, synthetic circuits must be designed to operate robustly in the presence of environmental variability and selectively respond to specific environmental inputs.153 A recent study demonstrated that a genetic circuit encoding memory of the transient inflammation signal tetrathionate in a murine Escherichia coli strain retained memory for long periods of time, as evidenced by detection of activated circuits in fecal samples six months after inflammation was induced in mice.154 A second challenge in engineering genetic circuits is minimizing fitness burden to the host cell due to competition for limited intracellular resources between the host cell and the synthetic circuit.155,156 If the metabolic costs are significant, nonfunctional mutants could rapidly outcompete the functional subpopulation.157,158 Although many advances have been made to design circuits to minimize metabolic burden,159 it is still difficult to predict the interactions between the host cell and the synthetic circuit and use this information to inform circuit design.160,161 Finally, biocontainment strategies must be developed to prevent engineered organisms from escaping into other environments.162 To implement biocontainment, toxic genes have been used to induce cell death in response to specific signals processed by multilayered logic gates. 163 However, the killing efficiency of the circuits declined over time and these circuits have not been characterized in complex environments.163
4.2. Design of Synthetic Communities from the Bottom Up.
In natural microbial communities, biochemical transformations are partitioned among diverse organisms, a phenomenon known as division of labor (DOL)164 (Figure 3d). DOL is thought to increase productivity by enabling community members to specialize at certain subtasks.165 In fact, several studies have demonstrated that synthetic consortia engineered for DOL have potential advantages compared to engineering a single strain, including modularity, the ability to exploit the unique capabilities of different strains,166 and the ability to control product rates and yields by altering community composition.167 Nevertheless, engineering DOL also has potential disadvantages, such as competition for shared resources and challenges maintaining stable coexistence of community members over time.168,169 The trade-offs between advantages and disadvantages of DOL have been recently explored using a theoretical framework.170 By analyzing 24 common architectures of metabolic pathways in which DOL can be implemented, a recent study concluded that DOL is advantageous when the increase in productivity due to DOL (as measured by product titer) is larger than the decrease in productivity arising from resource competition and limitations in cellular transport rates. The shape of the trade-off among these factors varies with metabolic pathway architecture.170 Furthermore, the authors suggest that DOL increases productivity when it reduces metabolic burden or toxicity. Metabolic burden and toxicity could be caused by expression of multiple genes in a pathway or the production of toxic intermediates.171 However, methods to predict the metabolic burden or toxicity of engineered pathways are lacking.
A core challenge for microbiome engineering is achieving stability as a function of time in response to perturbations. Equilibrium stability is a concept from dynamical systems and is defined as the rate at which a system returns to its original state (equilibrium point) following a perturbation. However, this definition does not apply to systems operating away from equilibrium. Microbial communities are highly dynamic and may not be operating at equilibrium because of temporally changing environmental stimuli. In fact, fluctuations in population sizes driven by external inputs to the system can promote species diversity over time.172 In microbial community engineering, rewiring of interactions among constituent community members may be required to achieve stability. Currently, the types (positive/negative/negligible), molecular mechanisms, and distributions of required interactions to realize stability and resilience to perturbations remain unresolved. A theoretical study showed that stability is enhanced by limiting positive interactions or globally weakening interactions.43 Bidirectional positive interactions can be destabilizing because of positive feedback loops, which renders the network sensitive to fluctuations in the abundance of organisms that are positively coupled in the network. Globally weakening interactions in the ecological network will result in a smaller change in organism abundance in response to variations in the abundances of other community members. A balance between negative and positive interactions in an ecological network can promote system stability: network topologies consisting of positive/negative (+/−) interactions can exhibit robust species coexistence by establishing a negative feedback loop.44 A different theoretical study showed that competitive interactions are stabilizing only when species exclusively consume but do not produce resources.173 The same study predicted that positive interactions are stabilizing when all pairwise interactions are reciprocated, or when production rates are sufficiently small.173
These predictions could be tested using engineered microbial communities. In addition to exploiting well-characterized mechanisms including bacterial signaling systems, cross-feeding of amino acids and metabolic byproduct utilization, microbial interactions can also be constructed by rewiring biosynthetic pathways.110,174–176 In one study, the authors used modular genetic pathway reconfiguration to design pairwise consortia with distinct interaction topologies.110 Experimental results from these pairwise consortia were used in model-guided design of three-and four-strain communities with predictable behaviors.110 In addition, spatial organization can augment community stability by excluding cheaters and invaders76,177 (Figure 2b) and could thus be harnessed as a design parameter for achieving desired community-level functions.
5. PERSPECTIVES
5.1. Challenges in Identifying Mechanisms of Ecological Interaction over Short and Long Time Scales.
Experimental approaches to determine ecological relationships and molecular mechanisms in microbial communities have largely focused on simple communities under well-defined conditions. Fabricated ecosystems that mimic the physical and chemical structure of natural environments (such as the gut-on-a-chip) can be used to determine environmental effects on interaction networks by providing investigators precise control over environmental variables.178,179 Once interactions have been identified, high-throughput genetic approaches such as Tn-Seq180 or genome-scale CRISPRi libraries181 could identity genes that enhance or weaken an interaction. Furthermore, metabolomic techniques such as mass spectrometry imaging could be used to identify molecules that may underlie the interaction.182 Isotope-labeling techniques such as 13C-metabolic flux analysis could be used to quantify intracellular fluxes,183,184 thereby linking intracellular metabolic states to interaction networks.
Long-term dynamics of microbial communities are ultimately determined by evolution. We currently have a limited understanding of ecological and evolutionary time scales of microbial communities and lack an effective strategy to predict or control the long-term evolution of microbiomes. One experimental evolution study showed that replicate communities followed a cluster of evolutionary trajectories measured by community structure, suggesting that community evolution may be partially determinstic.185 A second study showed that evolutionary outcomes of pairwise growth experiments followed simple qualitative rules, although the predictions were limited to a small set of closely related species with competitive interactions.186 In sum, these studies suggest that replication of laboratory evolution experiments is a useful tool for quantifying the forces shaping microbiome evolutionary dynamics. However, a core challenge is extracting evolutionary design principles that can be generalized across different systems. Similar experiments should be undertaken to predict community evolution patterns in more complex communities, in the presence and absence of spatial structure, and over evolutionary time scales.
5.2. Challenges in Computational Modeling of Microbiomes.
Computational models of microbes and microbial communities are often optimized for processes occurring at distinct spatial or temporal scales. However, developing a quantitative and mechanistic understanding of how molecular-level changes alter long-term evolutionary outcomes will require multiscale frameworks that can link processes occurring at different temporal or spatial scales. Some efforts are already underway, such as frameworks that embed constraint-based models of metabolism inside spatial118 or evolutionary models187 (Figure 2). Model-reduction techniques could be used to simplify mechanistic multiscale models. In addition, combining time-resolved measurements of community structure and/or function (e.g., metabolomics) with machine learning could be used to identify model structures underlying community dynamic behaviors. Such an approach has recently been used to predict metabolic pathway dynamics in engineered E. coli.188 Regardless of the modeling approach used, computational models must be rigorously validated with properly collected data in order to be useful. Validation may require new tools for spatial and temporal monitoring of metabolites, environmental parameters, and absolute abundance quantification of species abundance. Furthermore, the number of system parameters that could be modeled far exceeds the number that may be practical to infer from data. Combined experimental and computational studies could elucidate the types and frequency of data to collect, as well as the most informative way to use these data to accelerate predictive modeling.
5.3. Engineering Microbiomes or Designing Microbial Communities.
When introducing novel functions into natural microbiomes, model organisms such as E. coli may be preferred because of the availability of genetic tools. However, to persist stably in a new environment, laboratory strains must first be adapted to the abiotic and biotic properties of the target environment. If the key parameters of the target environment are known, microbes could be adaptively evolved in the lab for increased fitness before introduction to the new environment. Additionally, engineered organisms can be rapidly adapted to new environments using functional metagenomic libraries containing the genetic sequences from the resident bacterial community.189 Alternatively, microbes native to the target environment may be more suitable as an engineering chassis because of pre-existing adaptations. Thus, further development of genetic tools and parts libraries for nonmodel microbes should be encouraged, such as the promoter and ribosome-binding site libraries that have been developed for the human gut commensal Bacteroides thetaiotaomicron.190
When constructing synthetic microbial communities, a major challenge is optimizing division of labor (DOL) for community-level functions. DOL is determined in part by the metabolic burden an engineered pathway imposes on a cell across different environments. Indeed, the trade-off between engineered circuit activity and host-cell fitness depends on a changeable and uncertain environment. Coarse-grained kinetic models or genome-scale metabolic models could be used to simulate the fitness costs associated with different DOL schemes and predict functional activities associated with different configurations. Communities could be constructed and characterized, and the results used to identify new constraints and trade-offs that impact DOL.
Finally, control of microbiome structure and function requires advances in our understanding of how to manipulate complex networks of microorganisms that consist of numerous interconnected feedback loops. A control-theoretical frame-work should be developed for microbial communities to determine a minimum set of influential organisms and/or environmental inputs that need to be manipulated to steer the system to specific states. To implement such control architectures, we need modular platforms for rapidly designing microbiome editing tools to control system behaviors.
ACKNOWLEDGMENTS
We thank Yu-Yu Cheng, Ryan Clark, Jyun-Liang Lin, Erin Ostrem Loss, and James Papadopoulos for critical comments on the manuscript.
Funding
This work was supported by NIH NIGMS R35 GM124774–01 and ARO Young Investigator Award W911NF-17–1-0296.
Footnotes
The authors declare no competing financial interest.
REFERENCES
- (1).Falkowski PG, Fenchel T, and Delong EF (2008) The microbial engines that drive Earth’s biogeochemical cycles. Science (Washington, DC, U. S.) 320, 1034–1039. [DOI] [PubMed] [Google Scholar]
- (2).Busby PE, Soman C, Wagner MR, Friesen ML, Kremer J, Bennett A, Morsy M, Eisen JA, Leach JE, and Dangl JL (2017) Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 15, e2001793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Chaparro JM, Sheflin AM, Manter DK, and Vivanco JM (2012) Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 48, 489–499. [Google Scholar]
- (4).Valls M, and DeLorenzo V (2002) Exploiting the genetic and biochemical capacities of bacteria for the remediation of heavy metal pollution. FEMS Microbiol. Rev. 26, 327–338. [DOI] [PubMed] [Google Scholar]
- (5).Blaser MJ, Cardon ZG, Cho MK, Dangl JL, Donohue TJ, Green JL, Knight R, Maxon ME, Northen TR, Pollard KS, and Brodie EL (2016) Toward a Predictive Understanding of Earth’s Microbiomes to Address 21st Century Challenges. mBio 7, e00714–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Cho I, and Blaser MJ (2012) The human microbiome: at the interface of health and disease. Nat. Rev. Genet. 13, 260–270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (7).Chittim CL, Irwin SM, and Balskus EP (2018) Deciphering Human Gut Microbiota-Nutrient Interactions: A Role for Biochemistry. Biochemistry 57, 2567–2577. [DOI] [PubMed] [Google Scholar]
- (8).Cryan JF, and Dinan TG (2012) Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 13, 701–712. [DOI] [PubMed] [Google Scholar]
- (9).Arrieta M-C, Stiemsma LT, Amenyogbe N, Brown EM, and Finlay B (2014) The Intestinal Microbiome in Early Life: Health and Disease. Front. Immunol. 5, 427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Schink B (1997) Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 61, 262–280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (11).Manichanh C, Rigottier-Gois L, Bonnaud E, Gloux K, Pelletier E, Frangeul L, Nalin R, Jarrin C, Chardon P, Marteau P, Roca J, and Dore J (2006) Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut 55, 205–211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (12).Willing BP, Dicksved J, Halfvarson J, Andersson AF, Lucio M, Zheng Z, Jarnerot G, Tysk C, Jansson JK, and Engstrand L (2010) A Pyrosequencing Study in Twins Shows That Gastrointestinal Microbial Profiles Vary With Inflammatory Bowel Disease Phenotypes. Gastroenterology 139, 1844–1854. [DOI] [PubMed] [Google Scholar]
- (13).Noor SO, Ridgway K, Scovell L, Kemsley EK, Lund EK, Jamieson C, Johnson IT, and Narbad A (2010) Ulcerative colitis and irritable bowel patients exhibit distinct abnormalities of the gut microbiota. BMC Gastroenterol. 10, 134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).van Elsas JD, Chiurazzi M, Mallon CA, Elhottova D, Kristufek V, and Salles JF (2012) Microbial diversity determines the invasion of soil by a bacterial pathogen. Proc. Natl. Acad. Sci. U. S.A. 109, 1159–1164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (15).Chang JY, Antonopoulos DA, Kalra A, Tonelli A, Khalife WT, Schmidt TM, and Young VB (2008) Decreased Diversity of the Fecal Microbiome in Recurrent Clostridium difficile -Associated Diarrhea. J. Infect. Dis. 197, 435–438. [DOI] [PubMed] [Google Scholar]
- (16).Nannipieri P, Ascher J, Ceccherini MT, Landi L, Pietramellara G, and Renella G (2003) Microbial diversity and soil functions.. Eur J. Soil Sci. 54, 655–670. [Google Scholar]
- (17).Sunagawa S, Coelho LP, Chaffron S, Kultima JR, Labadie K, Salazar G, Djahanschiri B, Zeller G, Mende DR, Alberti A, Cornejo-Castillo FM, Costea PI, Cruaud C, D’Ovidio F, Engelen S, Ferrera I, Gasol JM, Guidi L, Hildebrand F, Kokoszka F, Lepoivre C, Lima-Mendez G, Poulain J, Poulos BT, Royo-Llonch M, Sarmento H, Vieira-Silva S, Dimier C, Picheral M, Searson S, Kandels-Lewis S, Bowler C, de Vargas C, Gorsky G, Grimsley N, Hingamp P, Iudicone D, Jaillon O, Not F, Ogata H, Pesant S, Speich S, Stemmann L, Sullivan MB, Weissenbach J, Wincker P, Karsenti E, Raes J, Acinas SG, Bork P, Boss E, Bowler C, Follows M, Karp-Boss L, Krzic U, Reynaud EG, Sardet C, Sieracki M, and Velayoudon D (2015) Structure and function of the global ocean microbiome. Science (Washington, DQ U. S.) 348, 1261359. [DOI] [PubMed] [Google Scholar]
- (18).Gonzalez A, King A, Robeson MS II, Song S, Shade A, Metcalf JL, and Knight R (2012) Characterizing microbial communities through space and time. Curr. Opin. Biotechnol 23, 431–436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (19).Nemergut DR, Schmidt SK, Fukami T, O’Neill SP, Bilinski TM, Stanish LF, Knelman JE, Darcy JL, Lynch RC, Wickey P, and Ferrenberg S (2013) Patterns and Processes of Microbial Community Assembly. Microbiol. Mol. Biol. Rev. 77, 342–356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (20).Fuhrman JA, Cram JA, and Needham DM (2015) Marine microbial community dynamics and their ecological interpretation. Nat. Rev. Microbiol. 13, 133–146. [DOI] [PubMed] [Google Scholar]
- (21).Tremaroli V, and Backhed F (2012) Functional interactions between the gut microbiota and host metabolism. Nature 489, 242–249. [DOI] [PubMed] [Google Scholar]
- (22).Sonnenburg ED, Zheng H, Joglekar P, Higginbottom SK, Firbank SJ, Bolam DN, and Sonnenburg JL (2010) Specificity of Polysaccharide Use in Intestinal Bacteroides Species Determines Diet-Induced Microbiota Alterations. Cell 141, 1241–1252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (23).Dethlefsen L, and Relman DA (2011) Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. U. S. A. 108, 4554–4561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (24).Maier L, Pruteanu M, Kuhn M, Zeller G, Telzerow A, Anderson EE, Brochado AR, Fernandez KC, Dose H, Mori H, Patil KR, Bork P, and Typas A (2018) Extensive impact of non-antibiotic drugs on human gut bacteria. Nature 555, 623–628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (25).Classen AT, Sundqvist MK, Henning JA, Newman GS, Moore JAM, Cregger MA, Moorhead LC, and Patterson CM (2015) Direct and indirect effects of climate change on soil microbial and soil microbial-plant interactions: What lies ahead? Ecosphere 6, No. art130. [Google Scholar]
- (26).Hutchins DA, and Fu F (2017) Microorganisms and ocean global change. Nat. Microbiol. 2, 17058. [DOI] [PubMed] [Google Scholar]
- (27).Phelan VV, Liu W-T, Pogliano K, and Dorrestein PC (2012) Microbial metabolic exchange—the chemotype-to-phenotype link. Nat. Chem. Biol. 8, 26–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (28).Braga RM, Dourado MN, and Araujo WL (2016) Microbial interactions: ecology in a molecular perspective. Braz. J. Microbiol. 47, 86–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (29).Koraimann G, and Wagner MA (2014) Social behavior and decision making in bacterial conjugation. Front. Cell. Infect. Microbiol. 4, 54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (30).Stempler O, Baidya AK, Bhattacharya S, Malli Mohan GB, Tzipilevich E, Sinai L, Mamou G, and Ben-Yehuda S (2017) Interspecies nutrient extraction and toxin delivery between bacteria. Nat. Commun. 8, 315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (31).Pande S, Shitut S, Freund L, Westermann M, Bertels F, Colesie C, Bischofs IB, and Kost C (2015) Metabolic cross-feeding via intercellular nanotubes among bacteria. Nat. Commun. 6, 6238. [DOI] [PubMed] [Google Scholar]
- (32).Konovalova A, and Søgaard-Andersen L (2011) Close encounters: contact-dependent interactions in bacteria. Mol. Microbiol. 81, 297–301. [DOI] [PubMed] [Google Scholar]
- (33).Fischbach MA, and Sonnenburg JL (2011) Eating For Two: How Metabolism Establishes Interspecies Interactions in the Gut. Cell Host Microbe 10, 336–347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (34).Abreu NA, and Taga ME (2016) Decoding molecular interactions in microbial communities. FEMS Microbiol. Rev. 40, 648–663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (35).Daims H, Taylor MW, and Wagner M (2006) Wastewater treatment: a model system for microbial ecology. Trends Biotechnol. 24, 483–489. [DOI] [PubMed] [Google Scholar]
- (36).Schroth MN, and Hancock JG (1982) Disease-Suppressive Soil and Root-Colonizing Bacteria. Science (Washington, DCU. S.) 216, 1376–1381. [DOI] [PubMed] [Google Scholar]
- (37).Young VB (2016) Therapeutic manipulation of the microbiota: past, present, and considerations for the future. Clin. Microbiol. Infect. 22, 905–909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (38).Guven D, van de Pas-Schoonen K, Schmid MC, Strous M, Jetten MSM, Sozen S, Orhon D, and Schmidt I (2004) Implementation of the Anammox Process for Improved Nitrogen Removal. J. Environ. Sci. Health, Part A: Toxic/Hazard. Subst. Environ. Eng. 39, 1729–1738. [DOI] [PubMed] [Google Scholar]
- (39).Keene NA, Reusser SR, Scarborough MJ, Grooms AL, Seib M, Santo Domingo J, and Noguera DR (2017) Pilot plant demonstration of stable and efficient high rate biological nutrient removal with low dissolved oxygen conditions. Water Res. 121, 72–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (40).Lindemann SR, Bernstein HC, Song H-S, Fredrickson JK, Fields MW, Shou W, Johnson DR, and Beliaev AS (2016) Engineering microbial consortia for controllable outputs. ISME J. 10, 2077–2084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (41).Sheth RU, Cabral V, Chen SP, and Wang HH. (2016) Manipulating Bacterial Communities by in situ Microbiome Engineering. Trends Genet. 32, 189–200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (42).Faust K, and Raes J (2012) Microbial interactions: from networks to models. Nat. Rev. Microbiol. 10, 538–550. [DOI] [PubMed] [Google Scholar]
- (43).Coyte KZ, Schluter J, and Foster KR (2015) The ecology of the microbiome: Networks, competition, and stability. Science (Washington, DQ U. S.) 350, 663–666. [DOI] [PubMed] [Google Scholar]
- (44).Venturelli OS, Carr A, Fisher G, Hsu R, Lau R, Bowen BP, Hromada S, Northen TR, and Arkin AP (2018) Deciphering microbial interactions in synthetic human gut micro-biome communities. Mol. Syst. Biol. 14, e8157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (45).Seth EC, and Taga ME (2014) Nutrient cross-feeding in the microbial world. Front. Microbiol. 5, 350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (46).Stiefel U, Tima MA, and Nerandzic MM (2015) Metallo-β-lactamase-producing Bacteroides species can shield other members of the gut microbiota from antibiotics. Antimicrob. Agents Chemother. 59, 650–653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (47).Hibbing ME, Fuqua C, Parsek MR, and Peterson SB (2010) Bacterial competition: surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 8, 15–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (48).Tyc O, van den Berg M, Gerards S, van Veen JA, Raaijmakers JM, de Boer W, and Garbeva P (2014) Impact of interspecific interactions on antimicrobial activity among soil bacteria. Front. Microbiol. 5, 567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (49).Cremer J, Arnoldini M, and Hwa T (2017) Effect of water flow and chemical environment on microbiota growth and composition in the human colon. Proc. Natl. Acad. Sci. U. S. A. 114, 6438–6443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (50).Wootton JT (1994) The Nature and Consequences of Indirect Effects in Ecological Communities. Annu. Rev. Ecol Syst. 25, 443–466. [Google Scholar]
- (51).Kelsic ED, Zhao J, Vetsigian K, and Kishony R (2015) Counteraction of antibiotic production and degradation stabilizes microbial communities. Nature 521, 516–519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (52).Liu A, Archer AM, Biggs MB, and Papin JA (2017) Growth-altering microbial interactions are responsive to chemical context. PLoS One 12, e0164919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (53).Kim HJ, Boedicker JQ, Choi JW, and Ismagilov RF (2008) Defined spatial structure stabilizes a synthetic multispecies bacterial community. Proc. Natl. Acad. Sci. U. S. A. 105, 18188–18193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (54).Bertrand S, Bohni N, Schnee S, Schumpp O, Gindro K, and Wolfender J-L (2014) Metabolite induction via microorganism co-culture: A potential way to enhance chemical diversity for drug discovery. Biotechnol. Adv. 32, 1180–1204. [DOI] [PubMed] [Google Scholar]
- (55).Netzker T, Fischer J, Weber J, Mattern DJ, Konig CC, Valiante V, Schroeckh V, and Brakhage AA (2015) Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 6, 299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (56).Traxler MF, Watrous JD, Alexandrov T, Dorrestein PC, and Kolter R (2013) Interspecies interactions stimulate diversification of the Streptomyces coelicolor secreted metabolome. mBio 4, e00459–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (57).Ponomarova O, Gabrielli N, Sévin DC, Mulleder M, Zirngibl K, Bulyha K, Andrejev S, Kafkia E, Typas A, Sauer U, Ralser M, and Patil KR (2017) Yeast Creates a Niche for Symbiotic Lactic Acid Bacteria through Nitrogen Overflow. Cell Syst. 5, 345–357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (58).Romero D, Traxler MF, Lopez D, and Kolter R (2011) Antibiotics as Signal Molecules. Chem. Rev. 111, 5492–5505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (59).Yim G, Huimi Wang H, and Davies FRS J (2007) Antibiotics as signalling molecules. Philos. Trans. R. Soc 2, 1195–1200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (60).Kurosawa K, Ghiviriga I, Sambandan TG, Lessard PA, Barbara JE, Rha C, and Sinskey AJ (2008) Rhodostreptomycins, antibiotics biosynthesized following horizontal gene transfer from Streptomyces padanus to Rhodococcus fascians. J. Am. Chem. Soc. 130, 1126–1127. [DOI] [PubMed] [Google Scholar]
- (61).Charusanti P, Fong NL, Nagarajan H, Pereira AR, Li HJ, Abate EA, Su Y, Gerwick WH, and Palsson BØ (2012) Exploiting adaptive laboratory evolution of streptomyces clavuligerus for antibiotic discovery and overproduction. PLoS One 7, e33727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (62).Hillesland KL (2018) Evolution on the bright side of life: microorganisms and the evolution of mutualism. Ann. N. Y. Acad. Sci. 1422, 88–103. [DOI] [PubMed] [Google Scholar]
- (63).D’Souza G, Shitut S, Preussger D, Yousif G, Waschina S, and Kost C (2018) Ecology and evolution of metabolic cross-feeding interactions in bacteria. Nat. Prod. Rep. 35, 455–488. [DOI] [PubMed] [Google Scholar]
- (64).Morris BEL, Henneberger R, Huber H, and Moissl-Eichinger C (2013) Microbial syntrophy: interaction for the common good. FEMS Microbiol. Rev. 37, 384–406. [DOI] [PubMed] [Google Scholar]
- (65).Diard M, Garcia V, Maier L, Remus-Emsermann MNP, Regoes RR, Ackermann M, and Hardt W-D (2013) Stabilization of cooperative virulence by the expression of an avirulent phenotype. Nature 494, 353–356. [DOI] [PubMed] [Google Scholar]
- (66).Foster RA, Sztejrenszus S, and Kuypers MMM (2013) Measuring carbon and N2 fixation in field populations of colonial and free-living unicellular cyanobacteria using nanometer-scale secondary ion mass spectrometry. J. Phycol 49, 502–516. [DOI] [PubMed] [Google Scholar]
- (67).van Gestel J, Vlamakis H, and Kolter R (2015) From Cell Differentiation to Cell Collectives: Bacillus subtilis Uses Division of Labor to Migrate. PLoS Biol. 13, e1002141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (68).Morris JJ (2015) Black Queen evolution: the role of leakiness in structuring microbial communities. Trends Genet. 31, 475–482. [DOI] [PubMed] [Google Scholar]
- (69).Leinweber A, Fredrik Inglis R, and Kummerli R (2017) Cheating fosters species co-existence in well-mixed bacterial communities. ISME J. 11, 1179–1188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (70).Butaite E, Baumgartner M, Wyder S, and Kummerli R (2017) Siderophore cheating and cheating resistance shape competition for iron in soil and freshwater Pseudomonas communities. Nat. Commun. 8, 414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (71).Lawrence D, Fiegna F, Behrends V, Bundy JG, Phillimore AB, Bell T, and Barraclough TG (2012) Species interactions alter evolutionary responses to a novel environment. PLoS Biol. 10, e1001330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (72).Tolker-Nielsen T, and Molin S (2000) Spatial Organization of Microbial Biofilm Communities. Microb. Ecol 40, 75–84. [DOI] [PubMed] [Google Scholar]
- (73).Mark Welch JL, Hasegawa Y, McNulty NP, Gordon JI, and Borisy GG (2017) Spatial organization of a model 15-member human gut microbiota established in gnotobiotic mice. Proc. Natl. Acad. Sci. U. S. A. 114, E9105–E9114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (74).van Gestel J, Weissing FJ, Kuipers OP, and Kovacs AT (2014) Density of founder cells affects spatial pattern formation and cooperation in Bacillus subtilis biofilms. ISME J. 8, 2069–2079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (75).Blanchard AE, and Lu T (2015) Bacterial social interactions drive the emergence of differential spatial colony structures. BMC Syst. Biol. 9, 59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (76).Momeni B, Waite AJ, and Shou W (2013) Spatial self-organization favors heterotypic cooperation over cheating. eLife 2, e00960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (77).Gerardin Y, Springer M, and Kishony R (2016) A competitive trade-off limits the selective advantage of increased antibiotic production. Nat. Microbiol. 1, 16175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (78).Basu S, Gerchman Y, Collins CH, Arnold FH, and Weiss R (2005) A synthetic multicellular system for programmed pattern formation. Nature 434, 1130–1134. [DOI] [PubMed] [Google Scholar]
- (79).Rusconi R, Garren M, and Stocker R (2014) Microfluidics Expanding the Frontiers of Microbial Ecology. Annu. Rev. Biophys. 43, 65–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (80).Feist AM, Herrgard MJ, Thiele I, Reed JL, and Palsson BØ (2009) Reconstruction of biochemical networks in micro-organisms. Nat. Rev. Microbiol. 7, 129–143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (81).O’Brien EJ, Monk JM, and Palsson BØ (2015) Using Genome-scale Models to Predict Biological Capabilities. Cell 161, 971–987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (82).Biggs MB, Medlock GL, Kolling GL, and Papin JA (2015) Metabolic network modeling of microbial communities. Wiley Interdiscip. Rev. Syst. Biol. Med. 7, 317–334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (83).Øyås O, and Stelling J (2018) Genome-scale metabolic networks in time and space. Curr. Opin. Syst. Biol. 8, 51–58. [Google Scholar]
- (84).Klitgord N, and Segre D (2010) Environments that induce synthetic microbial ecosystems. PLoS Comput. Biol. 6, e1001002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (85).Freilich S, Zarecki R, Eilam O, Segal ES, Henry CS, Kupiec M, Gophna U, Sharan R, and Ruppin E (2011) Competitive and cooperative metabolic interactions in bacterial communities. Nat. Commun. 2, 589. [DOI] [PubMed] [Google Scholar]
- (86).Heinken A, Sahoo S, Fleming RMT, and Thiele I (2013) Systems-level characterization of a host-microbe metabolic symbiosis in the mammalian gut. Gut Microbes 4, 28–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (87).El-Semman IE, Karlsson FH, Shoaie S, Nookaew I, Soliman TH, and Nielsen J (2014) Genome-scale metabolic reconstructions of Bifidobacterium adolescentis L2–32 and Faecali-bacterium prausnitzii A2–165 and their interaction. BMC Syst. Biol. 8, 41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (88).Shoaie S, Ghaffari P, Kovatcheva-Datchary P, Mardinoglu A, Sen P, Pujos-Guillot E, De Wouters T, Juste C, Rizkalla S, Chilloux J, Hoyles L, Nicholson JK, Dore J, Dumas ME, Clement K, Backhed F, and Nielsen J (2015) Quantifying Diet-Induced Metabolic Changes of the Human Gut Microbiome. Cell Metab. 22, 320–331. [DOI] [PubMed] [Google Scholar]
- (89).Stolyar SM, Van Dien SJ, Hillesland KL, Pinel N, Lie TJ, Leigh JA, and Stahl DA (2007) Metabolic modeling of a mutualistic microbial community. Mol. Syst. Biol. 3, 92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (90).Zomorrodi AR, and Maranas CD (2012) OptCom: A Multi-Level Optimization Framework for the Metabolic Modeling and Analysis of Microbial Communities. PLoS Comput. Biol. 8, e1002363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (91).Zhuang K, Izallalen M, Mouser P, Richter H, Risso C, Mahadevan R, and Lovley DR (2011) Genome-scale dynamic modeling of the competition between Rhodoferax and Geobacter in anoxic subsurface environments. ISME J. 5, 305–316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (92).Zomorrodi AR, Islam MM, and Maranas CD (2014) D-OptCom: Dynamic Multi-level and Multi-objective Metabolic Modeling of Microbial Communities. ACS Synth. Biol. 3, 247–257. [DOI] [PubMed] [Google Scholar]
- (93).Chiu H-C, Levy R, and Borenstein E (2014) Emergent Biosynthetic Capacity in Simple Microbial Communities. PLoS Comput. Biol. 10, e1003695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (94).Arkin AP, Cottingham RW, Henry CS, Harris NL, Stevens RL, Maslov S, Dehal P, Ware D, Perez F, Canon S, Sneddon MW, Henderson ML, Riehl WJ, Murphy-Olson D, Chan SY, Kamimura RT, Kumari S, Drake MM, Brettin TS, Glass EM, Chivian D, Gunter D, Weston DJ, Allen BH, Baumohl J, Best AA, Bowen B, Brenner SE, Bun CC, Chandonia J-M, Chia J-M, Colasanti R, Conrad N, Davis JJ, Davison BH, DeJongh M, Devoid S, Dietrich E, Dubchak I, Edirisinghe JN, Fang G, Faria JP, Frybarger PM, Gerlach W, Gerstein M, Greiner A, Gurtowski J, Haun HL, He F, Jain R, Joachimiak MP, Keegan KP, Kondo S, Kumar V, Land ML, Meyer F, Mills M, Novichkov PS, Oh T, Olsen GJ, Olson R, Parrello B, Pasternak S, Pearson E, Poon SS, Price GA, Ramakrishnan S, Ranjan P, Ronald PC, Schatz MC, Seaver SMD, Shukla M, Sutormin RA, Syed MH, Thomason J, Tintle NL, Wang D, Xia F, Yoo H, Yoo S, and Yu D (2018) KBase: The United States Department of Energy Systems Biology Knowledgebase. Nat. Biotechnol. 36, 566–569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (95).O’Brien EJ, Monk JM, and Palsson BØ (2015) Using Genome-scale Models to Predict Biological Capabilities. Cell 161, 971–987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (96).Wang Z, Danziger SA, Heavner BD, Ma S, Smith JJ, Li S, Herricks T, Simeonidis E, Baliga NS, Aitchison JD, and Price ND (2017) Combining inferred regulatory and reconstructed metabolic networks enhances phenotype prediction in yeast. PLoS Comput. Biol. 13, e1005489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (97).Varma A, and Palsson B 0. (1994) Stoichiometric flux balance models quantitatively predict growth and metabolic by-product secretion in wild-type Escherichia coli W3110. Appl Environ. Microbiol. 60, 3724–3731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (98).Schuetz R, Zamboni N, Zampieri M, Heinemann M, and Sauer U (2012) Multidimensional optimality of microbial metabolism. Science (Washington DQ U. S.) 336, 601–604. [DOI] [PubMed] [Google Scholar]
- (99).Hamilton JJ, Calixto Contreras M, and Reed JL (2015) Thermodynamics and H2 Transfer in a Methanogenic, Syntrophic Community. PLoS Comput. Biol. 11, e1004364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (100).Øyås O, and Stelling J (2018) Genome-scale metabolic networks in time and space. Curr. Opin. Syst. Biol. 8, 51–58. [Google Scholar]
- (101).May R, and McLean A (2007) Theoretical Ecology: Principles and Applications, 3rd ed., Oxford University Press, Oxford, United Kingdom. [Google Scholar]
- (102).Gonze D, Coyte KZ, Lahti L, and Faust K (2018) Microbial communities as dynamical systems. Curr. Opin. Microbiol. 44, 41–49. [DOI] [PubMed] [Google Scholar]
- (103).Åström KJ, and Murray RM (2008) Feedback Systems: An Introduction for Scientists and Engineers, 1st ed., Princeton University Press, Princeton, NJ. [Google Scholar]
- (104).Mounier J, Monnet C, Vallaeys T, Arditi R, Sarthou AS, Hélias A, and Irlinger F (2008) Microbial interactions within a cheese microbial community. Appl Environ. Microbiol. 74, 172–181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (105).Stein RR, Bucci V, Toussaint NC, Buffie CG, Ratsch G, Pamer EG, Sander C, and Xavier JB (2013) Ecological modeling from time-series inference: insight into dynamics and stability of intestinal microbiota. PLoS Comput. Biol. 9, e1003388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (106).Dam P, Fonseca LL, Konstantinidis KT, and Voit EO (2016) Dynamic models of the complex microbial metapopulation of Lake Mendota.. npj Syst. Biol. Appl. 2, 16007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (107).Momeni B, Xie L, and Shou W (2017) Lotka-Volterra pairwise modeling fails to capture diverse pairwise microbial interactions. eLife 6, e25051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (108).Vet S, de Buyl S, Faust K, Danckaert J, Gonze D, and Gelens L (2018) Bistability in a system of two species interacting through mutualism as well as competition: Chemostat vs. Lotka-Volterra equations. PLoS One 13, e0197462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (109).D’hoe K, Vet S, Faust K, Moens F, Falony G, Gonze D, Llorens-Rico V, Gelens L, Danckaert J, De Vuyst L, and Raes J (2018) Integrated culturing, modeling and transcriptomics uncovers complex interactions and emergent behavior in a three-species synthetic gut community. eLife 7, e37090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (110).Kong W, Meldgin DR, Collins JJ, and Lu T (2018) Designing microbial consortia with defined social interactions. Nat. Chem. Biol. 14, 821–829. [DOI] [PubMed] [Google Scholar]
- (111).Karr JR, Sanghvi JC, MacKlin DN, Gutschow MV, Jacobs JM, Bolival B, Assad-Garcia N, Glass JI, and Covert MW (2012) A whole-cell computational model predicts phenotype from genotype. Cell 150, 389–401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (112).Miskovic L, and Hatzimanikatis V (2010) Production of biofuels and biochemicals: In need of an ORACLE. Trends Biotechnol. 28, 391–397. [DOI] [PubMed] [Google Scholar]
- (113).Khodayari A, and Maranas CD (2016) A genome-scale Escherichia coli kinetic metabolic model satisfying flux data for multiple mutant strains. Nat. Commun. 7, 13806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (114).Edelstein-Keshet L (2005) Mathematical Models in Biology, 1st ed, SIAM, Philadelphia, PA. [Google Scholar]
- (115).Hellweger FL, and Bucci V (2009) A bunch of tiny individuals—Individual-based modeling for microbes. Ecol Modell. 220, 8–22. [Google Scholar]
- (116).Hellweger FL, Clegg RJ, Clark JR, Plugge CM, and Kreft J-U (2016) Advancing microbial sciences by individual-based modelling. Nat. Rev. Microbiol. 14, 461–471. [DOI] [PubMed] [Google Scholar]
- (117).Lardon LA, Merkey BV, Martins S, Dotsch A, Picioreanu C, Kreft JU, and Smets BF (2011) iDynoMiCS: Next-generation individual-based modelling of biofilms. Environ. Microbiol. 13, 2416–2434. [DOI] [PubMed] [Google Scholar]
- (118).Harcombe WR, Riehl WJ, Dukovski I, Granger BR, Betts A, Lang AH, Bonilla G, Kar A, Leiby N, Mehta P, Marx CJ, and Segre D (2014) Metabolic resource allocation in individual microbes determines ecosystem interactions and spatial dynamics. Cell Rep. 7, 1104–1115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (119).Bauer E, Zimmermann J, Baldini F, Thiele I, and Kaleta C (2017) BacArena: Individual-based metabolic modeling of heterogeneous microbes in complex communities. PLoS Comput. Biol. 13, e1005544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (120).Cole JA, Kohler L, Hedhli J, and Luthey-Schulten ZA (2015) Spatially-resolved metabolic cooperativity within dense bacterial colonies. BMC Syst. Biol. 9, 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (121).Chacon JM, Mobius W, and Harcombe WR (2018) The spatial and metabolic basis of colony size variation. ISME J. 12, 669–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (122).Hellweger FL, Fredrick ND, Mccarthy MJ, Gardner WS, Wilhelm SW, and Paerl HW (2016) Dynamic, mechanistic, molecular-level modelling of cyanobacteria: Anabaena and nitrogen interaction. Environ. Microbiol. 18, 2721–2731. [DOI] [PubMed] [Google Scholar]
- (123).Bucci V, Nunez-Milland D, Twining BS, and Hellweger FL (2012) Microscale patchiness leads to large and important intraspecific internal nutrient heterogeneity in phytoplankton. Aquat. Ecol. 46, 101–118. [Google Scholar]
- (124).Nowak MA (2006) Evolutionary Dynamics: Exploring the Equations of Life, 1st ed., Harvard University Press, Cambridge, MA. [Google Scholar]
- (125).Gore J, Youk H, and Van Oudenaarden A (2009) Snowdrift game dynamics and facultative cheating in yeast. Nature 459, 253–256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (126).Zomorrodi AR, and Segre D (2017) Genome-driven evolutionary game theory helps understand the rise of metabolic interdependencies in microbial communities. Nat. Commun. 8, 1563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (127).Litchman E, and Klausmeier CA (2008) Trait-Based Community Ecology of Phytoplankton. Annu. Rev. Ecol. Evol Syst. 39, 615–639. [Google Scholar]
- (128).Clark JR, Lenton TM, Williams HTP, and Daines SJ (2013) Environmental selection and resource allocation determine spatial patterns in picophytoplankton cell size. Limnol. Oceanogr. 58, 1008–1022. [Google Scholar]
- (129).Williams HTP, and Lenton TM (2007) Artificial selection of simulated microbial ecosystems. Proc. Natl. Acad. Sci. U. S. A. 104, 8918–8923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (130).van den Berg P, and Weissing FJ (2015) The importance of mechanisms for the evolution of cooperation. Proc. R. Soc. London, Ser. B 282, 20151382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (131).Broom M, and Rychtar J (2016) Nonlinear and Multiplayer Evolutionary Games In Advances in Dynamic and Evolutionary Games, pp 95–115, Springer International Publishing. [Google Scholar]
- (132).Dragosits M, and Mattanovich D (2013) Adaptive laboratory evolution -principles and applications for biotechnology. Microb. Cell Fact. 12, 64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (133).Lawson CE, Wu S, Bhattacharjee AS, Hamilton JJ, Mcmahon KD, Goel R, and Noguera DR (2017) Metabolic network analysis reveals microbial community interactions in anammox granules. Nat. Commun. 8, 15416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (134).Overmann J, Abt B, and Sikorski J (2017) Present and Future of Culturing Bacteria. Annu. Rev. Microbiol. 71, 711–730. [DOI] [PubMed] [Google Scholar]
- (135).Phillips PC (2008) Epistasis — the essential role of gene interactions in the structure and evolution of genetic systems. Nat. Rev. Genet. 9, 855–867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (136).Citorik RJ, Mimee M, and Lu TK (2014) Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat. Biotechnol. 32, 1141–1145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (137).Bikard D, Euler CW, Jiang W, Nussenzweig PM, Goldberg GW, Duportet X, Fischetti VA, and Marraffini LA (2014) Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials. Nat. Biotechnol. 32, 1146–1150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (138).Ando H, Lemire S, Pires DP, and Lu TK (2015) Engineering Modular Viral Scaffolds for Targeted Bacterial Population Editing. Cell Syst. 1, 187–196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (139).Hwang IY, Koh E, Wong A, March JC, Bentley WE, Lee YS, and Chang MW (2017) Engineered probiotic Escherichia coli can eliminate and prevent Pseudomonas aeruginosa gut infection in animal models. Nat. Commun. 8, 15028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (140).Hwang IY, Tan MH, Koh E, Ho CL, Poh CL, and Chang MW (2014) Reprogramming Microbes to Be Pathogen-Seeking Killers. ACS Synth. Biol. 3, 228–37. [DOI] [PubMed] [Google Scholar]
- (141).Palmer JD, Piattelli E, McCormick BA, Silby MW, Brigham CJ, and Bucci V (2018) Engineered Probiotic for the Inhibition of Salmonella via Tetrathionate-Induced Production of Microcin H47. ACS Infect. Dis. 4, 39–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (142).Jayaraman P, Holowko MB, Yeoh JW, Lim S, and Poh CL (2017) Repurposing a Two-Component System-Based Biosensor for the Killing of Vibrio cholerae. ACS Synth. Biol. 6, 1403–1415. [DOI] [PubMed] [Google Scholar]
- (143).Wittebole X, De Roock S, and Opal SM (2014) A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens. Virulence 5, 226–235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (144).Gupta S, Bram EE, and Weiss R (2013) Genetically Programmable Pathogen Sense and Destroy. ACS Synth. Biol. 2, 715–723. [DOI] [PubMed] [Google Scholar]
- (145).Saeidi N, Wong CK, Lo T-M, Nguyen HX, Ling H, Leong SSJ, Poh CL, and Chang MW (2011) Engineering microbes to sense and eradicate Pseudomonas aeruginosa, a human pathogen. Mol Syst. Biol. 7, 521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (146).Mallon CA, van Elsas JD, and Salles JF (2015) Microbial invasions: The process, patterns, and mechanisms. Trends Microbiol. 23, 719–729. [DOI] [PubMed] [Google Scholar]
- (147).Maldonado-Gomez MX, Martinez I, Bottacini F, O’Callaghan A, Ventura M, van Sinderen D, Hillmann B, Vangay P, Knights D, Hutkins RW, and Walter J (2016) Stable Engraftment of Bifidobacterium longum AH1206 in the Human Gut Depends on Individualized Features of the Resident Microbiome. Cell Host Microbe 20, 515–526. [DOI] [PubMed] [Google Scholar]
- (148).Shepherd ES, DeLoache WC, Pruss KM, Whitaker WR, and Sonnenburg JL (2018) An exclusive metabolic niche enables strain engraftment in the gut microbiota. Nature 557, 434–438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (149).Kearney SM, Gibbons SM, Erdman SE, and Alm EJ (2018) Orthogonal Dietary Niche Enables Reversible Engraftment of a Gut Bacterial Commensal. Cell Rep. 24, 1842–1851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (150).Dodd D, Spitzer MH, Van Treuren W, Merrill BD, Hryckowian AJ, Higginbottom SK, Le A, Cowan TM, Nolan GP, Fischbach MA, and Sonnenburg JL (2017) A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature 551, 648–652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (151).Brophy JAN, and Voigt CA (2014) Principles of genetic circuit design. Nat. Methods 11, 508–520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (152).Nandagopal N, and Elowitz MB (2011) Synthetic Biology: Integrated Gene Circuits. Science (Washington, DC, U. S.) 333, 1244–1248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (153).Venturelli OS, Egbert RG, and Arkin AP (2016) Towards Engineering Biological Systems in a Broader Context. J. Mol. Biol. 428, 928–944. [DOI] [PubMed] [Google Scholar]
- (154).Riglar DT, Giessen TW, Baym M, Kerns SJ, Niederhuber MJ, Bronson RT, Kotula JW, Gerber GK, Way JC, and Silver PA (2017) Engineered bacteria can function in the mammalian gut long-term as live diagnostics of inflammation. Nat. Biotechnol. 35, 653–658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (155).Qian Y, Huang H-H, Jimenez JI, and Del Vecchio D (2015) Resource Competition Shapes the Response of Genetic Circuits. ACS Synth. Biol. 6, 1263–1272. [DOI] [PubMed] [Google Scholar]
- (156).Weiße AY, Oyarzun DA, Danos V, and Swain PS (2015) Mechanistic links between cellular trade-offs, gene expression, and growth. Proc. Natl. Acad. Sci U. S. A. 112, E1038–E1047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (157).Sleight SC, Bartley BA, Lieviant JA, and Sauro HM (2010) Designing and engineering evolutionary robust genetic circuits. J. Biol. Eng. 4, 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (158).Sleight SC, and Sauro HM (2013) Visualization of Evolutionary Stability Dynamics and Competitive Fitness of Escherichia coli Engineered with Randomized Multigene Circuits. ACS Synth. Biol. 2, 519–528. [DOI] [PubMed] [Google Scholar]
- (159).Zhang C, Tsoi R, and You L (2016) Addressing biological uncertainties in engineering gene circuits. Integr. Biol. 8, 456–464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (160).Ceroni F, Boo A, Furini S, Gorochowski TE, Borkowski O, Ladak YN, Awan AR, Gilbert C, Stan G-B, and Ellis T (2018) Burden-driven feedback control of gene expression. Nat. Methods 15, 387–393. [DOI] [PubMed] [Google Scholar]
- (161).Ceroni F, Algar R, Stan G-B, and Ellis T (2015) Quantifying cellular capacity identifies gene expression designs with reduced burden. Nat. Methods 12, 415–418. [DOI] [PubMed] [Google Scholar]
- (162).Torres L, Kruger A, Csibra E, Gianni E, and Pinheiro VB (2016) Synthetic biology approaches to biological containment: pre-emptively tackling potential risks. Essays Biochem. 60, 393–410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (163).Chan CTY, Lee JW, Cameron DE, Bashor CJ, and Collins JJ (2016) Deadman” and “Passcode” microbial kill switches for bacterial containment. Nat. Chem. Biol. 12, 82–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (164).Zhang Z, Claessen D, and Rozen DE (2016) Understanding Microbial Divisions of Labor. Front. Microbiol. 7, 2070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (165).West SA, and Cooper GA (2016) Division of labour in microorganisms: an evolutionary perspective. Nat. Rev. Microbiol. 14, 716–723. [DOI] [PubMed] [Google Scholar]
- (166).Zhou K, Qiao K, Edgar S, and Stephanopoulos G (2015) Distributing a metabolic pathway among a microbial consortium enhances production of natural products. Nat. Biotechnol. 33, 377–383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (167).Minty JJ, Singer ME, Scholz SA, Bae C-H, Ahn J-H, Foster CE, Liao JC, and Lin XN (2013) Design and characterization of synthetic fungal-bacterial consortia for direct production of isobutanol from cellulosic biomass. Proc. Natl. Acad. Sci. U. S. A. 110, 14592–14597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (168).Brenner K, You L, and Arnold FH (2008) Engineering microbial consortia: a new frontier in synthetic biology. Trends Biotechnol. 26, 483–489. [DOI] [PubMed] [Google Scholar]
- (169).Shong J, Jimenez Diaz MR, and Collins CH (2012) Towards synthetic microbial consortia for bioprocessing. Curr. Opin. Biotechnol. 23, 798–802. [DOI] [PubMed] [Google Scholar]
- (170).Tsoi R, Wu F, Zhang C, Bewick S, Karig D, and You L (2018) Metabolic division of labor in microbial systems. Proc. Natl. Acad. Sci. U. S. A. 115, 2526–2531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (171).Yadav VG, De Mey M, Giaw Lim C, Kumaran Ajikumar P, and Stephanopoulos G (2012) The future of metabolic engineering and synthetic biology: Towards a systematic practice. Metab. Eng. 14, 233–241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (172).Huisman J, and Weissing FJ (1999) Biodiversity of plankton by species oscillations and chaos. Nature 402, 407–410. [Google Scholar]
- (173).Butler S, and O’Dwyer J (2018) Stability Criteria for Complex Microbial Communities. Nat. Commun. 9, 293605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (174).Kylilis N, Tuza ZA, Stan G-B, and Polizzi KM (2018) Tools for engineering coordinated system behaviour in synthetic microbial consortia. Nat. Commun. 9, 2677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (175).Mee MT, Collins JJ, Church GM, and Wang HH (2014) Syntrophic exchange in synthetic microbial communities. Proc. Natl. Acad. Sci. U. S. A. 111, E2149–E2156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (176).Harcombe WR (2010) Novel cooperation experimentally evolved between species. Evolution (Hoboken, NJ U. S.) 64, 2166–2172. [DOI] [PubMed] [Google Scholar]
- (177).Pande S, Kaftan F, Lang S, Svatos A, Germerodt S, and Kost C (2016) Privatization of cooperative benefits stabilizes mutualistic cross-feeding interactions in spatially structured environments. ISME J. 10, 1413–1423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (178).Kim HJ, Huh D, Hamilton G, and Ingber DE (2012) Human gut-on-a-chip inhabited by microbial flora that experiences intestinal peristalsis-like motions and flow. Lab Chip 12, 2165. [DOI] [PubMed] [Google Scholar]
- (179).Zhalnina K, Zengler K, Newman D, and Northen TR (2018) Need for Laboratory Ecosystems To Unravel the Structures and Functions of Soil Microbial Communities Mediated by Chemistry. mBio 9, e01175–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (180).van Opijnen T, Bodi KL, and Camilli A (2009) Tn-seq: High-throughput parallel sequencing for fitness and genetic interaction studies in microorganisms. Nat. Methods 6, 767–772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (181).Peters JM, Colavin A, Shi H, Czarny TL, Larson MH, Wong S, Hawkins JS, Lu CHS, Koo BM, Marta E, Shiver AL, Whitehead EH, Weissman JS, Brown ED, Qi LS, Huang KC, and Gross CA (2016) A comprehensive, CRISPR-based functional analysis of essential genes in bacteria. Cell 165, 1493–1506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (182).Petras D, Jarmusch AK, and Dorrestein PC (2017) From single cells to our planet—recent advances in using mass spectrometry for spatially resolved metabolomics. Curr. Opin. Chem. Biol. 36, 24–31. [DOI] [PubMed] [Google Scholar]
- (183).Ghosh A, Nilmeier J, Weaver DS, Adams PD, Keasling JD, Mukhopadhyay A, Petzold CJ, and Martín HG (2014) A peptide-based method for 13C Metabolic Flux Analysis in microbial communities. PLoS Comput. Biol. 10, e1003827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (184).Gebreselassie NA, and Antoniewicz MR (2015) 13C-Metabolic flux analysis of co-cultures: A novel approach. Metab. Eng. 31, 132–139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (185).Celiker H, and Gore J (2014) Clustering in community structure across replicate ecosystems following a long-term bacterial evolution experiment. Nat. Commun. 5, 4643. [DOI] [PubMed] [Google Scholar]
- (186).Friedman J, Higgins LM, and Gore J (2017) Community structure follows simple assembly rules in microbial microcosms. Nat. Ecol. Evol 1, 0109. [DOI] [PubMed] [Google Scholar]
- (187).Zomorrodi AR, and Segre D (2017) Genome-driven evolutionary game theory helps understand the rise of metabolic interdependencies in microbial communities. Nat. Commun. 8, 1563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (188).Costello Z, and Martin HG (2018) A machine learning approach to predict metabolic pathway dynamics from time-series multiomics data..npj Syst. Biol. Appl 4, 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (189).Yaung SJ, Deng L, Li N, Braff JL, Church GM, Bry L, Wang HH, and Gerber GK (2015) Improving microbial fitness in the mammalian gut by in vivo temporal functional metagenomics. Mol. Syst. Biol. 11, 788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (190).Mimee M, Tucker AC, Voigt CA, and Lu TK (2015) Programming a Human Commensal Bacterium, Bacteroides thetaio-taomicron, to Sense and Respond to Stimuli in the Murine Gut Microbiota. Cell Syst. 1, 62–71. [DOI] [PMC free article] [PubMed] [Google Scholar]