

Abstract
The influence of activation of glutamate receptor (GluR) on outward K+ current in cultured neonate rat hippocampal astrocytes was investigated. Patch-clamp analysis of K+ channel currents in cultured astrocytes identified the existence of 71 ± 6 and 161 ± 11 pS single-channel K+ currents that were sensitive to changes in voltage and [Ca2+]i and blocked by external TEA but not by charybdotoxin, iberiotoxin, apamin, or 4-aminopyridine. Reverse transcriptase (RT)-PCR and Northern blot analysis revealed transcripts of the Ca2+-activated K+ channel (KCa) β4-subunit (β4) (KCNMB4) in cultured astrocytes. Expression of the metabotropic glutamate receptor (mGluR) subtypes mGluR1 and mGluR5 and the ionotropic glutamate receptor (iGluR) subtypes iGluR1 and iGluR4 were detected by RT-PCR and immunofluorescence analysis in cultured astrocytes. The mGluR agonistsl-glutamate and quisqualate increased the open state probability (NPo) of the 71 and 161 pS K+ channel currents that were prevented by the mGluR receptor antagonists 1-aminoindan-1,5-dicarboxylic acid orl-(+)-2-amino-3-phosphonopropionic acid and not by the iGluR antagonists (+)-5-methyl-10,11-dihydro-5H-dibenzo [a,d] cyclohepten-5,10-imine maleate or CNQX. Activation of the two types of K+ channel currents by mGluR agonists was attenuated by pertussis toxin and by inhibition of phospholipase C (PLC) or cytochrome P450 arachidonate epoxygenase. These results indicate that brain astrocytes contain the KCNMB4 transcript and express two novel types of KCa channels that are gated by activation of a G-protein coupled metabotropic glutamate receptor functionally linked to PLC and cytochrome P450 arachidonate epoxygenase activity.
Keywords: neonate rat hippocampus, cultured astrocytes, excitatory amino acids, glutamate receptors, patch-clamp recording, Ca2+-activated K+ channels, KCNMB4, neuronal activity
Introduction
Astrocytes were originally considered to be only of structural importance (Matsas and Tsacopoulos, 1999). However, recent studies have indicated that astrocytes take up and metabolize neurotransmitters, buffer changes in extracellular ion concentration, and serve as intermediates in the cross talk between neurons and blood vessels (Kandel et al., 2000). Glutamate receptors (GluRs) are the most ubiquitous receptor type expressed in neuronal synapses and astrocytes. The excitatory neurotransmitter glutamate released from activated neurons stimulates two types of glutamate receptors, ionotropic glutamate receptors (iGluRs), which are ligand-gated ion channels and modulate synaptic response, and metabotropic glutamate receptors (mGluRs) (Nakanishi, 1992). So far, eight mGluRs (mGluR1–mGluR8) have been identified on the basis of sequence homology and pharmacological profile (Nakanishi, 1992). mGluR1 and mGluR5 are coupled to phosphoinositide hydrolysis, whereas other mGluRs are primarily coupled to a downregulation of cAMP formation (Nakanishi, 1992). Several studies have demonstrated expression of some of iGluR subunits in astrocytes, and the expression appears to vary among brain regions (Burnashev et al., 1992; Steinhauser and Gallo, 1996; Iino et al., 2001). Electrophysiologic studies of astrocytes support a functional role for some of the iGluR subtypes (Burnashev et al., 1992; Muller et al., 1992, 1997; Holzwarth et al., 1994; Robert and Magistretti, 1997; Schipke et al., 2001). However, the electrophysiological consequences of mGluR activation have not been well studied in astrocytes.
The excitability of astrocytes to stimulation by excitatory amino acids, other neurotransmitters, and humeral factors is a function of changes in ion channel activity. Astrocytes in culture express an array of voltage-gated ion channels, including as yet incompletely characterized Ca2+-activated K+ channels (KCa) (Barres et al., 1990; Hansson et al., 1994). The KCa channels are ubiquitous in a variety of tissue types and are involved in diverse physiological functions. Thus, vascular smooth muscle KCa channels are important regulators of vascular tone (Brayden and Nelson, 1992; Gebremedhin et al., 1992), and in neuronal cells they contribute to membrane excitability (Lee et al., 1995). In the presynaptic terminals, KCa channels are colocalized with voltage-dependent Ca2+channels and play a critical role in the regulation of transmitter release (Lee et al., 1995). To our knowledge, there is no evidence of identification of KCa channel currents in cultures of neonatal rat hippocampal astrocytes. Furthermore, no evidence exists that describes the influence of agonist-induced modulation of GluRs on the gating properties of KCa channel current in rat hippocampal astrocytes using the patch-clamp technique. The goals of the present studies were to (1) examine the type of GluR subtypes expressed in astrocytes, (2) identify and characterize the properties of native KCa channel currents in cultured astrocytes, and (3) determine the signal transduction pathways mediating activation of KCa channel currents by GluR stimulation. The pathways that were investigated pharmacologically include G-protein coupling to phospholipase C (PLC) and cytochrome P450 arachidonate epoxygenase activity. The latter was investigated because epoxyeicosatrienoic acids (EETs) are catalytically formed from arachidonic acid in cultured astrocytes by cytochrome P450 epoxygenase and stimulate KCa channel currents in vascular smooth muscle (Gebremedhin et al., 1992).
Materials and Methods
Cell culture. Astrocytes were cultured from cerebral cortices and hippocampi of 1- to 2-d-old Sprague Dawley rat brains under aseptic conditions as described previously (Alkayed et al., 1996). Briefly, brain tissue was cut into small pieces and transferred to a sterile dish containing 20 U/ml papain (Worthington, Freehold, NJ) and 0.15 mg/ml cysteine (Sigma, St. Louis, MO) dissolved in Earle's balanced salt solution (Invitrogen, Carlsbad, CA). The tissue pieces were incubated at 37°C for 40 min with gentle agitation and then washed three times in the feeding medium, which contained DMEM (Invitrogen) with 10% fetal bovine serum (ICN Biomedicals, Cleveland, OH) and 1% penicillin–streptomycin solution (Sigma). The tissue was then dissociated by triturating with a flame-narrowed Pasteur pipette. The cell suspension was diluted with feeding medium and seeded into 75 cm2 culture flasks (Costar, Cambridge, MA) at an initial density of 2 × 105 cells per square centimeter. Cells were incubated at 37°C in a 95 and 5% mixture of atmospheric air and CO2, respectively. The medium was changed after 2 d and subsequently twice per week. Confluent monolayers of 10- to 14-d-old primary cultures of rat hippocampal astrocytes were studied. The cells in culture contain >99% astrocytes as revealed by positive reaction of the cells to glial fibrillary acidic protein.
Indirect immunofluorescence for glutamate receptors.Confluent cultured astrocytes adherent to glass slides were fixed with 4% paraformaldehyde, blocked in PBS, pH 7.4, containing 0.1% BSA for 1 hr at room temperature, and incubated with polyclonal antibodies specific for mGluR1, mGluR5, GluR1 (Chemicon, Temecula, CA), and GluR4 (PharMingen, San Diego, CA) at dilutions of 1:100, 1:80, 1:40, and 1:50, respectively, overnight at 4°C. Astrocytes incubated without primary antibody served as controls. The astrocytes were then rinsed with PBS and incubated with fluorescein-conjugated goat anti-rabbit IgG (Alexa Fluor 488; Molecular Probes, Eugene, OR) at a dilution of 1:80 for 1 hr in the dark at room temperature. Slides were rinsed and mounted under coverslips. Fluorescent labeling was observed with a Nikon (Tokyo, Japan) E600 microscope equipped with epifluorescence using specific filters for fluorescein. Pictures were taken using a digital camera attached to the microscope.
Reverse transcription-PCR. Total RNA from 10- to 14-d-old astrocytes in culture was isolated using Trizol (Invitrogen). The RNA was treated with DNase I (Invitrogen) before PCR. Reverse transcription (RT) was performed using gene-specific primers and a Superscript one-step PCR kit (Invitrogen). The RT-PCR was performed by mixing reaction buffer with 1 μl of RNA (∼1 μg), the gene-specific primers at final concentration of 0.2 μm, and enzymes according to the instructions from the manufacturers. PCR was run as follows: 94°C for 2 min, followed by 35 cycles (94°C, 30 sec; 55°C, 30 sec; and 72°C, 1 min) and a final extension step (72°C, 7 min). Reactions omitting reverse transcriptase or DNA polymerase were used as control for contaminations. PCR products were run on 2% agarose gel and stained with ethidium bromide, and pictures were taken under UV light. The gene-specific primers used were 5′-GGACGAGATCAGACAACCAG-3′ (sense) and 5′-TCGTACCACCATTTGCTTTTCA-3′ (antisense) for GluR1, 5′-GAAGGACCCAGTGACCAGC-3′ (sense) and 5′-TCGTACCACCATTTGTTTTTCA-3′ (antisense) for GluR4, 5′-GACCCTACCTTTTCGAACCC-3′ (sense) and 5′-GGCTTCCCAATTATGGAGACC-3′ (antisense) for mGluR1, and 5′-GCAGGATGCACAGCAACAGG-3′ (sense) and 5′-GGCTGGATCTCTGCGAAGGT-3′ (antisense) for mGluR5. The specific primers used for amplification of rat KCa β4-subunit (KCNMB4) were designed from sequences with the GenBank accession number AY028605. The primers used for KCNMB4 were 5′-GATGGCGAAGCTCAGGGTGTCT-3′ (sense) and 5′-CTCCTCCCCGTTAAGAGAACT-3′ (antisense).
Northern blot analysis. Twenty-five micrograms of total RNA isolated from cultured astrocytes was electrophoresed in a 1.0% agarose/formaldehyde gel and transferred to a nylon membrane. The amplified PCR product of KCNMB4 was cloned into pCRII-topo TA cloning vector (Invitrogen), and the insert was sequenced. The insert was cut out from the plasmid with EcoRI (Promega, Madison, WI), gel purified, and labeled with [32P]CTP (3000 Ci/mm) (Amersham Biosciences, Arlington Heights, IL) using a random primer method (CTP labeling beads; Amersham Biosciences). The membrane was hybridized at 68°C overnight in ExpressHyp solution (Clontech, Cambridge, UK) and washed twice for 30 min in 2× SSC/0.05% SDS (1× SSC contains 150 mm NaCl and 15 mm sodium citrate, pH 7.0) at room temperature, once for 60 min in 0.5× SSC/0.1% SDS at 50°C, and once for 15 min in 0.1× SSC/0.1% SDS at 50°C before the radioactive signal was registered in a Typhoon 9400 (Amersham Biosciences).
KCa channel current recordings.Single-channel K+ currents were recorded at room temperature from cell-attached and excised inside-out membrane patches of primary cultures of rat brain astrocytes using the patch-clamp technique as described previously (Hamill et al., 1981; Gebremedhin et al., 1996). Briefly, recording pipettes were fabricated from borosilicate glass, pulled on a 2-stage micropipette puller (PC-84), and heat-polished under a microscope (MF-83 heat polisher; Narishige, Tokyo, Japan). The recording pipettes were mounted on a three-way hydraulic micromanipulator (Narishige) for placement of the tips on the cell membrane. High-resistance seals (>1 GΩ) were established by applying a slight suction between fire-polished pipette tips (3-10 MΩ) and cell membranes. The offset potentials between pipette and bath solution were corrected with an offset circuit before each experiment. Pipette potential was clamped, and single-channel currents were recorded through a List EPC-7 patch-clamp amplifier (List Biologic, Campbell, CA). The amplifier output was low-pass filtered at 1 kHz with an eight pole Bessel filter (Frequency Devices, Haverhill, MA). Current signals were digitized at a sampling rate of 2.5 kHz. Single-channel currents were analyzed using a pClamp software package (pClamp version 5.5 and 6.04; Axon Instruments, Foster City, CA) to determine event frequency, mean current amplitudes, and open state probability. The mean open state probability (NPo) was expressed asNPo = I/i, whereI is the time averaged current, N is the number of channels, i is the amplitude of the unitary current, andPo is the probability of a channel being open (Aldrich and Yellen, 1983). Slope conductance was determined by fitting the unitary current–voltage relationship using least square linear regression. Macroscopic KCa currents were measured after forming a tight seal (10–20 GΩ), and the cell membrane was ruptured by applying pulsatile suction until there was a large increase in capacitive current, indicating access to the interior of the cell. Capacitive transients were not electronically cancelled by introduction of fast and slow capacitance compensation and series resistance.
Outside-out membrane patch. Isolation of an outside-out membrane patch from astrocytes was performed after gigaseal (10–20 GΩ) formation and patch rupture using pipette solution containing 150 mm KCl, 3 mm HEPES, and low Ca2+ (<10−6m) achieved by buffering with 3 mm BAPTA, pH 7.2, and after withdrawal of the pipette tip from the cell (Hamill et al., 1981). Single-channel K+ currents were recorded from outside-out membrane patches bathed in normal physiological salt solution (PSS) at an approximate membrane potential of −70 mV, and the effects of the various K+ channel blockers were studied by adding into the bath.
Patch-clamp solutions. Pipette solutions for both cell-attached and excised inside-out patches contained (in mm): 145 KCl, 1.8 CaCl2, 1.1 MgCl2, and 5 HEPES, with the final pH adjusted to 7.2 with KOH. During recording from cell-attached patches, the bath solution was normal PSS, whereas for excised inside-out patches and some cell-attached patches it was composed of (in mm): 145 KCl, 1.8 CaCl2, 1.1 MgCl2, 5 HEPES, and 10 EGTA, with pH adjusted to 7.2 with KOH. This resulted in a calculated final [Ca2+]i of 10−7m (Godt, 1974). The bath was contained in a volume of 1 ml that was continually exchanged with fresh solution at a rate of 2 ml/min by gravitational flow. To study the sensitivity of inside-out patches of cultured astrocytes, the [Ca2+]i was calculated using a computer program (Godt, 1974). In some experiments the solution bathing the excised inside-out membrane patches was exchanged with a solution in which KCl was reduced to 40 mm by substituting with equimolar amounts of NaCl to study the relative selectivity of the channels for Na+ and K+.
Whole-cell K+ currents were recorded from cultured astrocytes bathed in a normal PSS using a recording pipette solution of the following composition (in mm): 145 KCl, 1 MgCl2, 1.8 CaCl2, 5 EGTA, 2 dipotassium adenosine triphosphate, and 10 HEPES, pH 7.2.
Drugs and chemicals. Dithiothreitol, EGTA, 1,2-BAPTA, TEA chloride, 4-aminopyridine (4-AP), l-glutamate, miconazole,l-nitro-l-arginine-methyl-ester, soybean trypsin inhibitor, and pertussis toxin (PTX) were purchased from Sigma. Papain and collagenase type II were purchased from Worthington. U-73122 was obtained from Biomol (Plymouth Meeting, PA). (+)-5-methyl-10,11-dihydro-5H-dibenzo [a,d] cyclohepten-5,10-imine maleate (MK-801), CNQX,l-(+)-2-amino-3-phosphonopropionic acid (l-AP-3), 1-aminoindan-1,5-dicarboxylic acid (AIDA), and quisqualate were purchased from Tocris Cookson (Bristol, UK). Iberiotoxin (IBX), apamin, and charybdotoxin (CTX) were obtained from Alomone Labs (Jerusalem, Israel). Final concentrations of iberiotoxin, charybdotoxin, or apamin were prepared by diluting aliquots of frozen stock solutions (1–4 μm) in the bath or pipette solutions.
Statistical analysis. Data are presented as mean ± SEM. Differences between mean values were assessed using a Student'st test or ANOVA for multiple comparisons. p< 0.05 was considered statistically significant.
Results
Identification of single-channel KCa currents
Voltage sensitivity
Representative tracings of single-channel K+ currents recorded from excised inside-out membrane patches of cultured rat brain astrocytes at different patch potentials using symmetrical KCl (145 mm) solution are presented in Figure1A. The amplitudes and opening frequencies of single-channel K+ currents through both the small- and large-amplitude current-conducting channels increased in response to changes in patch potential between −60 and 60 mV when recorded in excised inside-out membrane patches bathed in symmetrical KCl (145 mm) solution (Fig. 1A). The single-channel slope conductance averaged 71 ± 6 pS (n = 4–26 cells) for the small-amplitude single-channel current and 161 ± 11 pS (n = 10–35 cells) for the large-amplitude single-channel current (Fig.1B). The voltage sensitivity of both the 71 and 161 pS K+ channels was determined by measuringNPo over the patch potential range of −60 to 60 mV. As summarized in Figure2A, theNPo of both K+ channel types increased during step depolarization of the excised inside-out membrane patches of cultured astrocytes.
Fig. 1.

Current–voltage relationship of K+ channel currents in cultured astrocytes.A, Representative tracings of voltage-dependent openings of two different amplitude single-channel K+currents recorded from excised inside-out patches of cultured brain astrocytes at various patch potentials using symmetrical KCl (145 mm). c represents the closed state of the channel. B, Mean current–voltage relationship of the two single-channel K+ current types determined in 4–18 cells revealed unitary slope conductance of 71 ± 5 and 161 ± 9 pS for the small- and large-amplitude K+ currents, respectively. Vertical lines represent mean ± SEM.
Fig. 2.

Voltage sensitivity and selectivity for K+ of the two types of K+channels in cultured astrocytes. A, Bar graphs depicting voltage-sensitivities of the openings of the 71 and 161 pS single-channel K+ currents recorded from excised inside-out patches of cultured brain astrocytes using symmetrical KCl (145 mm) containing 0.1 μm[Ca2+]i. Increases in patch potential from −60 to 60 mV in steps of 20 mV progressively increased theNPo of the 71 and 161 pS K+ channels. Vertical lines represent mean ± SEM. The asterisk denotes a significant difference (p < 0.05; n= 4–20 patches) from the value measured at a patch potential of −60 mV. B, K+ selectivity.a, Examples of single-channel K+current tracings recorded from inside-out patches of cultured rat brain astrocytes at a patch potential (PP) of 60 mm using symmetrical KCl (145 mm) containing 0.1 μm Ca2+ and a bath solution in which the concentration of K+ was reduced to 40 mm by equimolar replacement with Na+(bottom). Reduction of the concentration of K+ in the bath significantly reduced the unitary amplitude of both the 71 and 161 pS K+ channel types. c represents the closed state of the channel.S, Small; L, large.b, Bar graphs depicting significant reduction of the mean unitary current amplitudes of the 71 and 161 pS single-channel K+ currents revealing high selectivity for K+ over Na+ of both K+ channel types (n = 5;p < 0.05).
Selectivity for K+
The current–voltage relationship curves of both the 71 and the 161 pS single-channel K+ currents had reversal potentials near 0 mV (Fig. 1B) when recorded from excised inside-out membrane patches of cultured astrocytes using symmetrical KCl (145 mm) solution. As depicted in Figure 2B, reduction of the concentration of K+ from 145 to 40 mmin the bathing solution by equimolar replacement with Na+ significantly reduced the unitary current amplitudes of the 71 and 161 pS K+and resulted in a shift of the reversal potentials. Thus, when K+ in the bath was reduced from 145 to 40 mm, the observed reversal potentials were 27.9 and 32.5 mV (EK = 33.5 mV) for the 71 and 161 pS K+ channels, respectively. These shifts in reversal potential are consistent with those of channels highly selective for K+.
Sensitivity to [Ca2+]i
The Ca2+ dependence of the openings of the 71 and 161 pS K+ channels was examined by varying the concentration of Ca2+ on the cytosolic surface of astrocytic inside-out membrane patches during recording at patch potentials of 40 and −40 mV. When [Ca2+]i was elevated from 0.001 to 0.01 μm and then to 0.1 μm, the NPo of the 71 pS K+ channel increased from 0.001 ± 0.0004 − 0.0024 ± 0.005 and then to 0.0086 ± 0.0005, whereas the NPo of the 161 pS K+ channel increased from 0.0015 ± 0.0007 − 0.0041 ± 0.0005 and then to 0.0096 ± 0.003 during recording at 40 mV, respectively, (n = 5–7 for each group; *p < 0.05). These data indicate that both the 71 and 161 pS single-channel K+currents in astrocyte membranes are activated by elevation of [Ca2+]i in the submicromolar range. As depicted in Fig.3, A and B, activation of both the 71 and 161 pS single-channel K+ currents by increases in [Ca2+]i is voltage dependent. The two channel types exhibited a similar trend of activation to changes in voltage and [Ca2+]i, in that both channel types displayed more increased opening at the depolarizing patch potential at all [Ca2+]i studied. In a separate study, application of 1 mm ATP on the cytoplasmic surface of inside-out membrane patches did not affect the NPo of both the 71 pS (0.0011 ± 0.0004 before and 0.0012 ± 0.0001 after) and 161 pS (0.0015 ± 0.0002 before and 0.0013 ± 0.0005 after;p > 0.05; n = 4 for all groups) K+ channel currents recorded at a patch potential of 40 mV, suggesting that neither of these two K+ channel types are ATP-sensitive K+ channels.
Fig. 3.

Calcium sensitivity of K+channel currents in cultured astrocytes. Calcium-dependent activation of the 71 and 161 pS single-channel K+ currents recorded from inside-out membrane patches of cultured astrocytes is shown. A, Representative single-channel K+ currents recorded at patch potentials of 40 mV (left) and −40 mV (right) and free [Ca2+]i of 0.001, 0.01, and 0.1 μm. The frequency of openings of the 71 and 161 pS K+ channel currents were both Ca2+ and voltage dependent. crepresents the closed state of the channel. B,Bar graphs depicting changes in theNPo of the 71 and 161 pS single-channel K+ currents in response to step increases in free [Ca2+]i from 0.001 to 0.01 to 0.1 μm during recording at patch potentials of 40 mV and −40 mV. The NPo of both the 71 and 161 pS K+ channels significantly increased in response to increases in free [Ca2+] in a voltage-dependent manner. The asterisk denotes significance difference (p < 0.05) from control (n = 4–5 patches).
Effects of K+ channel inhibitors
The effects of known KCa channel inhibitors TEA (1 mm), CTX (300 nm), IBX (300 nm), or apamin (300 nm), and that of the delayed rectifier K+ channel blocker 4-AP (1 mm) (Blatz and Magleby, 1986; Hermann and Erxleben, 1987; Lang and Ritchie, 1987) were studied on the openings of two types of single-channel KCa currents in outside-out membrane patches of cultured astrocytes by application into the bath. At a membrane potential of −70 mV, two types of single-channel KCa currents were recorded from the outside-out patches of astrocytes, which were not sensitive to the blocker effects of an externally applied 300 nm concentration of CTX, IBX, or apamin, as well as 1 mm 4-AP (data not shown). However, external application of TEA (1 mm) reversibly blocked the opening frequency and NPo of the two types of single-channel KCa currents recorded from the outside-out patches (Fig.4A,B). In a previous study, the insensitivity of neuronal KCa channels to the toxin blockers CTX or IBX has been attributed to the high level of expression of the human KCNMB4 homolog in the brain (Meera et al., 2000). To examine whether astrocytes contain KCNMB4 mRNA, we investigated the expression of KCNMB4 in cultured astrocytes. As depicted in Figure 4C, RT-PCR (a) and Northern blot analysis (b) detected KCNMB4 transcripts in cultured astrocytes, which could provide a molecular explanation for the lack of sensitivity to CTX, IBX, or apamin of the two types of KCa channels in astrocytes.
Fig. 4.

Effects of different KCa channel blockers on two types of single-channel KCa currents in outside-out membrane patches of cultured astrocytes recorded using a pipette solution containing 150 mm KCl, 3 mmHEPES, and 0.1 μm Ca2+ at pH 7.2.A, Single-channel KCa currents were recorded from outside-out patches at approximately −70 mV before (control) and after TEA, CTX, IBX, or apamin was added to the bath. External application of TEA (1 mm) reversibly blocked (left) the openings of both the small- and the large-conductance KCa channel currents, whereas the openings of the two types of KCa channel currents were resistant to external application of CTX (300 nm), IBX (300 nm), or apamin (300 nm), as shown in the respective panels. C−, Closed;S, small; L, large.B, Bar graphs depicting a summary of the effects of external application of TEA (1 mm), CTX (300 nm), IBX (300 nm), or apamin (300 nm) on the NPo of the small- and large-conductance KCa single-channel currents recorded from outside-out patches of cultured astrocytes. TEA, but not CTX, IBX, or apamin, reversibly reduced the NPo of the two types of single-channel currents. The asteriskdenotes significant difference (p < 0.05;n = 4–5). C, Detection by RT-PCR (a) and Northern blot analysis (b) of the expression of KCNMB4 transcripts in cultured brain astrocytes. MM, Molecular marker.
Externally applied apamin (300 nm), either alone (Fig.4A,B) or in combination with charybdotoxin (300 nm) or iberiotoxin (300 nm) (data not shown), also failed to alter the openings of both types of single-channel KCacurrents in outside-out membrane patches of cultured astrocytes, ruling out the possibility that these two types of K+ channels in astrocytes represent the small or the intermediate conductance KCa channel current reported in other tissues. Together, these findings indicate that brain astrocytes in culture express two types of novel single-channel KCa currents and contain transcript of the human KCa β4-subunit KCNMB4 homolog that could contribute to the insensitivity of these two KCa channel types to the toxin blockers CTX and IBX. These two KCa channels in astrocytes appear different from other KCa channels identified in various tissues (Farley and Rudy, 1988; Reinhart et al., 1989; Nelson and Quayle, 1995; Gebremedhin et al., 1996).
To further investigate whether the macroscopic KCa channel current recorded from cultured astrocytes displays properties similar to those of the astrocytic single-channel KCa currents, the effects of TEA (1 mm), CTX (300 nm), IBX (300 nm), or apamin (300 nm) were re-examined on an outward macroscopic K+ current recorded from cultured astrocytes. External application of TEA (1 mm) induced reversible inhibition of the astrocytic macroscopic KCa current, whereas external application of CTX, IBX, or apamin had no effect on this macroscopic KCa current, consistent with the lack of effect of these toxin blockers on the single-channel KCacurrents recorded from the outside-out membrane patches of astrocytes (n = 4–5) (Fig.5Ba–Bd). As depicted in Figure 5A, b and c, lowering extracellular [Ca2+] significantly reduced the magnitude of the macroscopic current and markedly depressed (p < 0.05; n = 4–5) the normalized macroscopic current–voltage relationship curve. These findings indicate that the astrocytic macroscopic current displays properties of a Ca2+-dependent K+ channel current, which appears to be carried through the same KCa channels identified in culturedastrocytes using the single-channelcurrent-recording technique.
Fig. 5.

A, Effects of low external Ca2+ concentration on the action of glutamate on macroscopic KCa channel current in cultured astrocytes.a, Control outward macroscopic KCa current recorded from cultured astrocytes during step depolarization from −60 to 80 mV in steps of 10 mV increments from a holding potential of −70 mV. b, Lowering external [Ca2+] significantly reduced the magnitude of macroscopic KCacurrents compared with the control (a).c, Application of glutamate (300 μm) to the bath was unable to elicit any effect in low-Ca2+bathing media. Replacement of Ca2+ to the bath restored the magnitude of the macroscopic KCa current to control level (d). e, Line graphs depicting the current–voltage relationship of astrocytic macroscopic KCa current in external bath solution containing a physiological concentration of Ca2+ (open circles), in low-Ca2+ external bath solution (filled triangles), and after addition of glutamate (300 μm, open triangles) to the bath. B, Effects of different KCa channel blockers on macroscopic K+ current recorded from cultured astrocytes. Application of TEA (1 mm) to the bath significantly reduced the macroscopic current (b), whereas addition of 300 nmcharybdotoxin, iberiotoxin, or apamin or 1 mm 4-AP (data not shown) had no effect on the magnitude of the macroscopic KCa current (c) compared with the control (a). d, Bar graphs depicting a summary of the effects of TEA, CTX, IBX, and apamin on the normalized peak macroscopic KCa current recorded from cultured astrocytes. Only application of TEA (1 mm) significantly reduced the macroscopic KCacurrent. The asterisk denotes significant difference (p < 0.05; n = 5 for each group).
Expression of glutamate receptor subtypes
To determine the types of glutamate receptor subtypes expressed in primary cultures of rat brain astrocytes, RT-PCR and immunofluorescence analysis studies were performed using gene-specific primers and subtype-specific antibodies, respectively. RT-PCR analysis amplified PCR bands of the expected size for the ionotropic iGluR1 (631 bp) and iGluR4 (625 bp) and for the metabotropic mGluR1 (570 bp) and mGluR5 (407 bp), whereas no expression of the mGluR7 (634 bp) glutamate receptor subtype was detected (Fig.6A). No such RT-PCR products were observed when the reaction was performed without RNA template or when reverse transcription was omitted. As depicted in Figure 6B, the results of immunofluorescence studies using subtype-specific antibodies against GluR1, GluR4, mGluR1, and mGluR5 confirmed the expression of the protein for ionotropic iGluR1 and iGluR4 and the metabotropic mGluR1 (570 bp) and mGluR5 (407 bp) glutamate receptors, albeit a stronger signal of the later subtypes detected in cultured brain astrocytes. Together, these results suggest that primary cultures of astrocytes express the ionotropic iGluR1 and iGluR4 and the metabotropic mGluR1 and mGluR5 receptor subtypes at the protein and transcript level.
Fig. 6.

RT-PCR and immunofluorescence analysis of glutamate receptor subtype expression in cultured astrocytes.A, Total RNA was isolated from 10- to 14-d-old astrocytes in culture, and mRNA expression for GluR1, GluR4, mGluR1, mGluR5, and mGluR7 was examined by RT-PCR using gene-specific primers. The PCR analysis revealed gene expression of the GluR1, GluR4, mGluR1, and mGluR5 subtypes, but not for the mGluR7 subtype in astrocytes that was detectable in brain, which was used as a positive control.GAPDH, Glyceraldehyde-3-phosphate dehydrogenase.B, Immunofluorescence analysis using subtype-specific antibodies detected the expression of ionotropic glutamate receptor subtypes iGluR1 and iGluR4 (a) and metabotropic glutamate receptor subtypes mGluR1 and mGluR5 in cultured brain astrocytes (b).
Effect of glutamate receptor activation on the KCachannel currents in cultured hippocampal astrocytes
Application of l-glutamate to the bath increased theNPo of the 71 and 161 pS single-channel KCa currents recorded from cell-attached patches of cultured rat brain astrocytes at 60 mV using symmetrical KCl (145 mm) solution. Thus, cumulative addition of glutamate to the bath markedly increased the frequency of openings of both the 71 and 161 pS single-channel KCa currents (Fig.7A) and significantly increased the NPo of the 71 pS KCa channel current from the controlNPo value of 0.003 ± 0.0002 − 0.004 ± 0.002 at 30 μm to 0.015 ± 0.004 at 100 μm and then to 0.022 ± 0.003 at 300 μm; theNPo of the 161 pS KCa channel current was increased from the control NPo value of 0.0034 ± 0.0004 − 0.003 ± 0.0001 at 30 μm to 0.015 ± 0.004 at 100 μm and then to 0.025 ± 0.004 at 300 μm(n = 5–9 cells for each group) (Fig. 7B). The glutamate-evoked increase in NPoof the two KCa channel types was mimicked by a structurally different mGluR agonist, quisqualate. It increased theNPo of the 71 pS KCa channel current from the controlNPo of 0.003 ± 0.004 − 0.0047 ± 0.0004 at 30 μm to 0.012 ± 0.004 at 100 μm and then to 0.022 ± 0.005 at 300 μm. It also increased theNPo of the 161 pS KCa channel current from the controlNPo of 0.0033 ± 0.00031 − 0.0037 ± 0.0003 at 30 μm to 0.015 ± 0.005 at 100 μm and then to 0.028 ± 0.008 at 300 μm (n = 4–10 cells for each group) (Fig.8A).
Fig. 7.
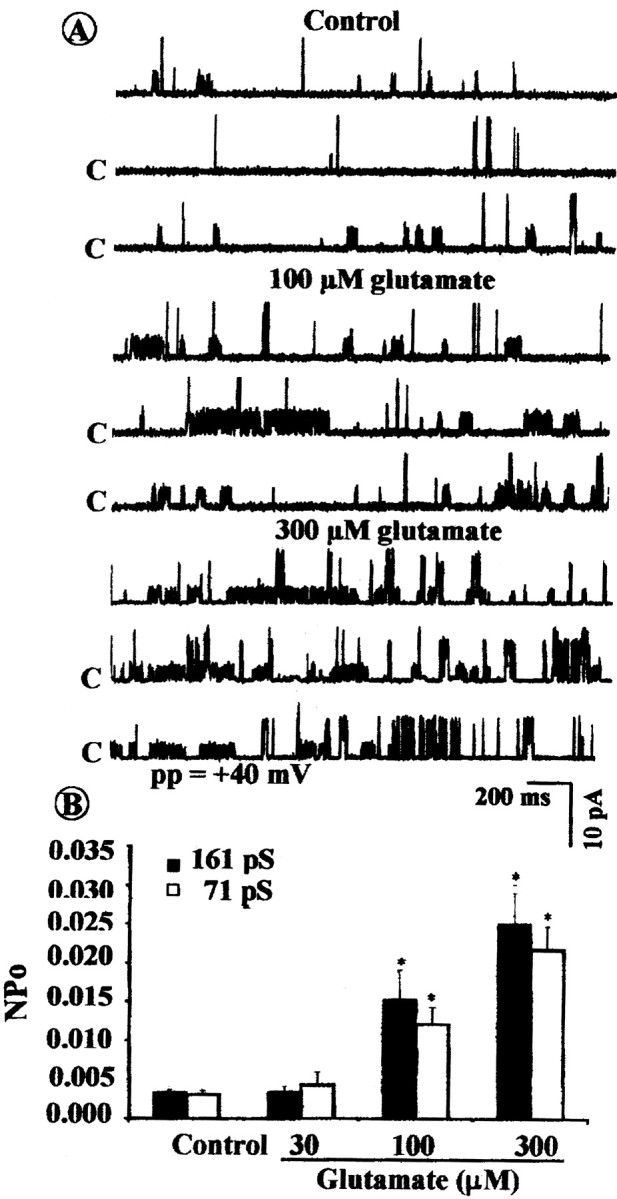
Effects of glutamate on the openings of the 71 and 161 pS single-channel KCa currents in cell-attached patches of cultured astrocytes recorded at a patch potential (pp) of 60 mV using symmetrical KCl (145 mm) solution. A, Representative tracings of single-channel KCa currents under control conditions and after addition of 100 or 300 μm glutamate to the bath.C represents the closed state of the channel.B, Summary of effects of glutamate (30–300 μm) on the NPo of the 71 pS (open bars) and the 161 pS (filled bars) single-channel KCa currents. Glutamate beginning at 100 μm significantly increased the opening frequencies and NPo of both the 71 and 161 pS KCa channel currents. Vertical linesrepresent mean ± SEM. The asterisk denotes significant difference from the respective controls (p < 0.05; n = 7–12 cells).
Fig. 8.

A, Effects of the mGluR agonist quisqualate on the KCa single-channel activity in cultured astrocytes. Application of quisqualate (30–300 μm) increased the NPo of the 71 pS (open bars) and the 161 pS (shaded bars) KCa channel currents recorded from cell-attached patches of cultured brain astrocytes at a patch potential of 60 mV using symmetrical KCl (145 mm) solution. Quisqualate induced a concentration-related significant increase in theNPo of the 71 and 161 pS KCachannel currents. Vertical lines represent mean ± SEM. The asterisk denotes significant difference (p < 0.05) from the control.B, Effects of the iGluR antagonists MK-801 and CNQX and the mGluR antagonists l-AP-3 and AIDA on the glutamate-induced increase in NPo of the KCa channel currents. Application of MK-108 (100 μm) or CNQX (100 μm) had no effect (*p > 0.05), whereas addition of the mGluR antagonists l-AP-3 (100 μm) or AIDA (100 μm) significantly blocked the glutamate-induced increase in NPo of both the 71 and 161 pS single-channel KCa currents (*p < 0.05) (n = 4 for each group). #p> 0.05 compared with control.
To determine the types of glutamate receptor involved in mediating the glutamate-induced activation of the two KCachannel types in cultured astrocytes, the effects of two mechanistically different antagonists of ionotropic glutamate receptors, MK-801 and CNQX, were examined. Application of the iGluR antagonist MK-801 (100 μm) or CNQX (100 μm) had no effect on the glutamate-induced increase in the NPo of single-channel KCa currents (Fig. 8B). In contrast, the glutamate-evoked enhancement of theNPo of KCasingle-channel currents in cultured astrocytes was completely attenuated by the mGluR5 antagonist AIDA (100 μm) and by the mixed mGluR1 and mGluR5 antagonist l-AP-3 (100 μm) (Fig. 8B).
Application of glutamate (300 μm) or quisqualate (300 μm) to the bath also significantly enhanced (p < 0.05) the magnitude of the macroscopic KCa current in cultured astrocytes (Fig.9B,C) that was completely attenuated in the presence of the KCa channel blocker TEA (1 mm) (Fig. 9D). Figure 9Edepicts a summary of the glutamate- and quisqualate-evoked enhancement, as well as inhibition by the KCachannel blocker TEA (1 mm) of the normalized peak macroscopic KCa current in cultured astrocytes recorded during a 10 mV step depolarization between −60 and 80 mV. Both glutamate and quisqualate significantly increased the magnitude of the macroscopic KCa current at almost all positive membrane potentials studied compared with the control, whereas application of TEA to the bath significantly inhibited the control current. In an attempt to examine whether glutamate activates astrocytic KCa channels independent of external Ca2+, the effect of glutamate (300 μm) on the macroscopic KCa current recorded from cultured astrocytes was determined after removal of Ca2+ from the external recording solution. Lowering the extracellular [Ca2+] significantly reduced the mean peak outward K+ current. In the absence of external Ca2+, glutamate failed to enhance the magnitude of the outward macroscopic KCa current or alter the current–voltage relationship curve in cultured brain astrocytes (Fig.5Ab,c). As shown in Figure 5d, replacement of external Ca2+ restored the magnitude of peak macroscopic KCa current to the control level (Fig. 5a). These findings indicate that the opening of the macroscopic KCa channel current in cultured brain astrocytes is dependent on the availability of external Ca2+.
Fig. 9.

Effects of externally applied mGluR agonists glutamate and quisqualate on whole-cell (macroscopic) KCacurrents recorded from cultured astrocytes. A–D, Macroscopic KCa currents recorded under control conditions (A) and after application to the bath of 300 μm glutamate (B), 300 μm quisqualate (C), and 300 μm glutamate (Glut) or 300 μm quisqualate (Quis) (D) in the presence of 1 mm TEA.E, Summary of mean peak current–voltage relationship demonstrating a significant enhancement of the peak macroscopic KCa current by glutamate or quisqualate and inhibition of the control current by TEA (1 mm). (*p< 0.05; n = 5 cells).
In a separate group of experiments, pretreatment of cultured astrocytes with PTX, an inhibitor of G-proteins of the Gi/Go subtype, prevented the ability of glutamate to increase theNPo of the two types of KCa channel currents in cultured astrocytes, indicating that the effects of glutamate on the astrocytic KCa channel currents are attributable to activation of mGluR coupled to PTX-sensitive G-proteins (Fig.10). In PTX-pretreated astrocytes, the other mGluR agonist quisqualate (300 μm) also failed to increase the NPo of both the 71 and 161 pS KCa channel currents. Thus, theNPo for the 71 pS KCa channel changed from 0.0032 ± 0.0004 − 0.0037± 0.0002, whereas theNPo of the 161 pS KCa channel changed from 0.0034 ± 0.0003 − 0.0040 ± 0.002 in response to quisqualate application in control and PTX-treated astrocytes, respectively (n = 4–5 cells; p > 0.05). To determine whether the glutamate-induced activation of mGluR is linked to PLC and has an influence on the glutamate-induced increased activities of both the 71 and 161 pS single-channel KCa currents, the effects of the PLC inhibitor U-73122 on the glutamate-induced activation of the KCa single-channel currents in cultured astrocytes were examined. As summarized in Figure 10, the glutamate-induced increase in the NPoof both the 71 and 161 pS KCa single-channel currents in cell-attached patches of cultured astrocytes was significantly attenuated by preapplication of the PLC inhibitor U-73122 (30 μm; n = 4–5; *p < 0.005), indicating the involvement of PLC in the glutamate-induced increased activities of the two types of astrocytic KCa channel currents.
Fig. 10.

Effects of the G-protein inhibitor PTX (1 μg/ml), the PLC inhibitor U-73122 (30 μm), the cytochrome P450 arachidonate epoxygenase inhibitor miconazole (5 μm), and the nitric oxide synthase inhibitorl-NAME (100 μm) on the glutamate-induced increase in NPo of the 71 and 161 pS single-channel KCa currents in cultured brain astrocytes. Pretreatment of cultured astrocytes with either PTX for 2 hr or U-73122 or miconazole for 20 min significantly attenuated the glutamate-evoked increase in NPo of the 71 and 161 pS single-channel KCa currents recorded from cell-attached patches of cultured astrocytes at a patch potential of 60 mV using symmetrical KCl (145 mm) solution. Vertical lines represent mean ± SEM. The asteriskdenotes significant difference (p < 0.05) from control (n = 4–5 for each group).#p < 0.05.
Glutamate has been shown previously to induce the release of cytochrome P450 arachidonate epoxygenase-derived EETs from cultured brain astrocytes, which activate arterial KCa channels and induce cerebral vasodilation (Gebremedhin et al., 1992; Alkayed et al., 1996; Nithipatikom et al., 2001). To examine whether the actions of glutamate on KCa channel currents in astrocytes are mediated by release of endogenously formed EETs, the effect of the cytochrome P450 epoxygenase inhibitor miconazole (5 μm) was studied. As depicted in Figure 10, the glutamate-induced increase in the NPoof both the 71 and 161 pS KCa single-channel currents in cell-attached patches of brain astrocytes was blunted by a 30 min pretreatment of cultured astrocytes with miconazole (n = 5; *p < 0.05). In contrast, blockade of nitric oxide (NO) synthase withN-ω-nitro-l-arginine methyl ester (l-NAME) (100 μm; n = 5) had no effect on the glutamate-evoked increase in the NPoof both the 71 and 161 pS KCa single-channel currents (Fig. 10). Miconazole, U-73222, andl-NAME, at the concentrations used, had no influence on the openings of KCa channel currents in excised inside-out membrane patches, ruling out any direct effect of these blockers on KCa channel activity (data not shown). Together, these findings suggest that stimulation by glutamate of a G-protein-coupled mGluR activates KCachannel currents in astrocytes via a PLC- and cytochrome P450 arachidonate epoxygenase-dependent pathway.
Discussion
We have identified and characterized two novel types of K+-selective single-channel KCa currents with a unitary conductance of 71 ± 5 and 161 ± 9 pS in membranes of primary cultures of neonatal rat brain astrocytes using symmetrical KCl (145 mm) solution and the patch-clamp technique. These two KCa channels were sensitive to blockade by TEA but were insensitive to 4-AP, a blocker of the delayed rectifier K+ channel, and to IBX, CTX, or apamin, known inhibitors of the KCa channel current in different tissues (Blatz and Magleby, 1986; Hermann and Erxleben, 1987;Lang and Ritchie, 1987; Talvenheimo et al., 1988; Brayden and Nelson, 1992; Gebremedhin et al., 1996). One obvious difference observed between the two KCa channel types identified in the membranes of cultured astrocytes was that they represent small and large conductance levels. This difference in conductance levels did not appear to be attributed to the existence of subconductance states, in that open and closed states of both the small- and large-conductance KCa channels were distinct and frequent in all membrane patches studied and displayed direct transitions from the main open states to the closed states (Fox, 1987). The properties of these two types of KCa channel currents identified in membranes of astrocytes in the present study are similar if not identical to the previously reported KCa channel phenotypes in brain plasma vesicles (Reinhart et al., 1989) and to the KCa channel currents described in a variety of tissue types (Farley and Rudy, 1988; Reinhart et al., 1989).
Although the existence and distribution of an array of voltage-gated ion channels in glial cells and astrocytes have been reported previously (Barres et al., 1990), very little is known about the expression and functional role of KCa channels in astrocytes (Barres et al., 1990). To our knowledge, the present finding is the first description of the identification of 71 and 161 pS KCa channel currents in the membranes of cultured astrocytes. Because the activities of both the 71 and 161 pS KCa channels are not sensitive to elevated concentrations of ATP, it is unlikely that these KCa channels represent an ATP-sensitive K+ channel (KATP) (data not shown).
An interesting property of the 71 and 161 pS KCasingle-channel currents observed in cultured astrocytes was that both the 71 and 161 pS KCa channels were insensitive to IBX, CTX, and apamin toxins known to inhibit KCa channel currents, despite their dual regulation by voltage and increases in cytosolic [Ca2+]i. KCa channel types resistant to inhibition by KCa channel blocker toxins such as CTX and IBX also have been found previously in rat brain plasma vesicles (Reinhart et al., 1989). However, the cause or the mechanism that makes the KCa channel currents insensitive to IBX and CTX in either the brain plasma vesicles (Reinhart et al., 1989) or cultured astrocytes of the present study remains unknown. In the present study, we also found that cultured astrocytes express the KCa channel β4-subunit (β4) KCNMB4 at a transcriptional level. KCNMB4 has been detected previously in the brain and represents the molecular mechanism that renders the neuronal KCa channel α-subunit insensitive to IBX and CTX (Meera et al., 2000). The detection of the KCNMB4 transcript in cultured astrocytes could provide a possible explanation for the lack of sensitivity to CTX and IBX of the astrocytic KCa channel currents identified in the present study. The insensitivity of both the 71 and 161 pS KCa single-channel currents to the toxin blockers of the KCa channel could also indicate that these two KCa channel types native in astrocytes might be associated with the β4-subunit (β4).
The KCa channels are important regulators of arterial muscle reactivity and the development of pressure-induced myogenic constriction in the cerebral circulation and in other arterial beds (Brayden and Nelson, 1992; Gebremedhin et al., 1992; Nelson and Quayle, 1995; Gebremedhin et al., 1996), whereas in neuronal cells they contribute to coordination of membrane excitability (Lee et al., 1995) and to regulation of the resting potential and control of spontaneous impulse generation (Johansson et al., 2001). In presynaptic nerve terminals, KCa channels are colocalized with voltage-dependent Ca2+ channels and have been suggested to play a critical role in the regulation of transmitter release (Marrion and Tavalin, 1998). In cochlear hair cells, KCa channels contribute to electrical tuning of cochlear hair cells that determines the cochlear resonant frequency (Krishnan et al., 1999). Although the functional role of the two types of KCa channel currents identified in cultured astrocytes in the normal physiology of astrocytes in vivo is yet to be understood, they may be involved in the regulation of membrane potential of these cells and serve as target membrane molecules for endogenous modulatory influences. Furthermore, by permitting the efflux of K+ from these cells, KCa channels may also contribute to the spatial K+ buffering capabilities of brain astrocytes (Paulson and Newman, 1987), especially if activated by calcium waves propagated through gap junctions forming an astrocyte syncytium. The finding that glutamate activates astrocytic KCa channel currents via stimulation of mGluR is intriguing, and suggests that the two KCa channel types may serve as a possible site through which brain astrocytes sense neuronal activity and relay the signal to cerebral microvessels in the vicinity.
The results of the present study also demonstrated that neonate rat brain astrocytes in culture express ionotropic glutamate receptor subtypes, iGluR1 and iGluR4, and metabotropic glutamate receptor subtypes, mGluR1 and mGluR5, at the protein and transcript level. However, using specific pharmacological blockers, we found that inhibition of mGluR but not iGluR attenuated the glutamate-induced increased openings of the 71 and the 161 pS single-channel KCa currents in cultured astrocytes, thus suggesting a functional coupling between mGluR and the two KCa channel types in astrocytes, which could determine the communication between activated neurons and astrocytes. Such coupling not only demonstrates that astrocytic KCa channels can be activated by stimulation of mGluR, but also may help to understand the physiological functions of astrocytes. Interestingly, in their recent elegant study, Isaacson and Murphy (2001) discovered the functional coupling between iGluR and KCa channels in rat olfactory bulb granule cells, which they suggested to have a modulatory role on synaptic transmission. Therefore, it appears that the coupling between glutamate receptor subtypes and KCa channels could be cell-type specific.
The fact that inhibition of G-proteins of the Gi/Go subtype with PTX abrogated the glutamate-induced activation of the two KCa channel current types in astrocytes suggests that the mGluR subtypes expressed in cultured astrocytes are coupled through G-proteins to a variety of signal transduction systems, such as activation of PLC. Moreover, the reduction of the response to glutamate by a PLC inhibitor implicates the contribution of phosphatidylinositol hydrolysis, which in addition to releasing other second messengers also liberates arachidonic acid from membrane phospholipids (Dennis et al., 1991) that may contribute to activation of KCachannel types invoked by stimulation of glutamate receptor.
Because inhibition of cytochrome P450 epoxygenase but not nitric oxide synthase attenuated the glutamate-induced activation of the two astrocytic KCa channel types, it appears that the release of P450 arachidonate epoxygenase-derived EETs rather than NO is primarily required to mediate the glutamate-evoked increase in the activities of the two KCa channel types. This observation is consistent with our previous findings that demonstrated the ability of glutamate to stimulate release of EETs from astrocytes (Alkayed et al., 1997). In vivo, EETs may act to increase KCa channel activity directly or may act indirectly as a calcium influx factor (Rziglinski et al., 1999) to replenish Ca2+ stores necessary to sustain an increased Ca2+ level for KCa channel activation. The results of the present study in astrocytes together with the previous work on isolated vascular smooth muscle cells (Gebremedhin et al., 1992) implicate the ability of EETs to act on KCa channels. A recently emerging physiological role of astrocytes is related to their capacity to express the cytochrome P450 enzyme of the 2C11 gene family (Alkayed et al., 1996), which catalyzes the epoxidation of arachidonic acid into four regioisomers of EETs. These regioisomeric EETs cause cerebral vasodilation through activation of KCa channel current (Gebremedhin et al., 1992), which also appears to be a major molecular target for the EETs in various cell types (Gebremedhin et al., 1992; Campbell et al., 1996;Baron et al., 1997). The role of cytochrome P450 2C11 epoxygenase that catalyzes the formation of the EETs in the regulation of cerebral blood flow (CBF) has become more relevant, in that inhibition of this enzyme in vivo reduced the increase in CBF induced by glutamate and neuronal activation (Alkayed et al., 1997;Bhardwaj et al., 2000). These findings resulted in the inclusion of the EETs to the list of endogenous products alleged to mediate functional hyperemia in the brain (Harder et al., 1998, 2002).
In conclusion, the expression of functional KCachannels as well as formation of P450 arachidonate epoxygenase-derived EETs, which activate these channels in astrocytes could form part of an intrinsic mechanism that links neuronal activity and regional cerebral blood flow. Further investigation of the molecular transduction mechanisms of the neuronal regulatory components of regional cerebral blood flow will lead to a greater understanding of how disruption of these normal mechanisms may lead to certain pathological disorders such as stroke and Alzheimer's disease.
Footnotes
This work was supported in part by National Heart, Lung, and Blood Institute Grants HL3833-16 and HL59996-01, Veterans Affairs Merit Review Grants 3440-02P and 3440-03N, and grants from the Wenner–Grens Foundation to J.B. We thank Jayashree Narayanan and Kris Hoefert for excellent technical assistance.
Correspondence should be addressed to Dr. Debebe Gebremedhin, Medical College of Wisconsin, Department of Physiology, 8701 Watertown Plank Road, Milwaukee, WI 53226. E-mail: mariyei@mcw.edu.
References
- 1.Aldrich RW, Yellen G. Analysis of nonstationary channel kinetics. In: Sakmann B, Neher E, editors. Single channel recording. Plenum; New York: 1983. pp. 287–299. [Google Scholar]
- 2.Alkayed NJ, Narayanan J, Gebremedhin D, Medhora M, Roman RJ, Harder DR. Molecular characterization of an arachidonic acid epoxygenase in rat brain astrocytes. Stroke. 1996;27:971–979. doi: 10.1161/01.str.27.5.971. [DOI] [PubMed] [Google Scholar]
- 3.Alkayed NJ, Eric KB, Narayanan J, Petrie KA, Kohler-Cabot AE, Harder DR. Role of P-450 arachidonic acid epoxygenase in the response of cerebral blood flow to glutamate in rats. Stroke. 1997;28:1066–1072. doi: 10.1161/01.str.28.5.1066. [DOI] [PubMed] [Google Scholar]
- 4.Baron A, Frieden M, Beny JL. Epoxyeicosatrienoic acids activate a high-conductance, Ca2+-activated K+ channel in pig coronary artery endothelial cells. J Physiol (Lond) 1997;504:537–543. doi: 10.1111/j.1469-7793.1997.537bd.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Barres BA, Chun LLY, Corey DP. Ion channel in vertebral glia. Annu Rev Neurosci. 1990;13:441–474. doi: 10.1146/annurev.ne.13.030190.002301. [DOI] [PubMed] [Google Scholar]
- 6.Bhardwaj A, Northington FJ, Carhuapoma JR, Falck JR, Harder DR, Traystman RJ, Koehler RC. P-450 epoxygenase and NO synthase inhibitors reduce cerebral blood flow response to N-methyl-d-aspartate. Am J Physiol. 2000;279:H1616–H1624. doi: 10.1152/ajpheart.2000.279.4.H1616. [DOI] [PubMed] [Google Scholar]
- 7.Blatz AL, Magleby KL. Single apamin blocked Ca2+-activated K+ channels of small conductance in cultured rat skeletal muscle. Nature. 1986;323:718–720. doi: 10.1038/323718a0. [DOI] [PubMed] [Google Scholar]
- 8.Brayden JE, Nelson MT. Regulation of arterial tone by activation of calcium-dependent potassium channels. Science. 1992;256:532–535. doi: 10.1126/science.1373909. [DOI] [PubMed] [Google Scholar]
- 9.Burnashev N, Khodorova A, Jonas P, Helm PJ, Wisden W, Monyer H, Seeburg PH, Sakmann B. Calcium-permeable AMPA-kainate receptors in fusiform cerebellar glial cells. Science. 1992;256:566–570. doi: 10.1126/science.1317970. [DOI] [PubMed] [Google Scholar]
- 10.Campbell WB, Gebremedhin D, Pratt PF, Harder DR. Identification of epoxyeicosatrienoic acids as endothelium-derived hyperpolarizing factors. Circ Res. 1996;78:415–423. doi: 10.1161/01.res.78.3.415. [DOI] [PubMed] [Google Scholar]
- 11.Dennis ED, Rhee SG, Billah MM, Hannun YA. Role of phospholipases in generating lipid second messengers in signal transduction. FASEB J. 1991;5:2068–2077. doi: 10.1096/fasebj.5.7.1901288. [DOI] [PubMed] [Google Scholar]
- 12.Farley J, Rudy B. Multiple types of voltage-dependent Ca2+-activated K+ channels of large conductance in rat brain synaptosomal membranes. Biophys J. 1988;53:919–934. doi: 10.1016/S0006-3495(88)83173-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fox AJ. Ion channel subconductance states. J Membr Biol. 1987;97:1–8. doi: 10.1007/BF01869609. [DOI] [PubMed] [Google Scholar]
- 14.Gebremedhin D, Ma YH, Falck JR, Roman RJ, VanRollins M, Harder DR. Mechanism of action of cerebral epoxyeicosatrienoic acids on cerebral arterial smooth muscle. Am J Physiol. 1992;263:H519–H525. doi: 10.1152/ajpheart.1992.263.2.H519. [DOI] [PubMed] [Google Scholar]
- 15.Gebremedhin D, Kaldunski M, Jacobs ER, Harder DR, Roman RJ. Coexistence of two types of Ca2+-activated K+ channels in rat renal arterioles. Am J Physiol. 1996;270:F69–F81. doi: 10.1152/ajprenal.1996.270.1.F69. [DOI] [PubMed] [Google Scholar]
- 16.Godt RE. Calcium-activated tension of skinned muscle fibers of the frog. J Gen Physiol. 1974;63:722–739. doi: 10.1085/jgp.63.6.722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch-clamp technique for high-resolution current recording from cells and cell-free membrane patches. Pflügers Arch. 1981;391:85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- 18.Hansson E, Johansson BB, Westergren I, Rönnbäck L. Glutamate-induced swelling of single astroglial cells in primary culture. Neuroscience. 1994;63:1057–1066. doi: 10.1016/0306-4522(94)90572-x. [DOI] [PubMed] [Google Scholar]
- 19.Harder DR, Alkayed NJ, Lange AR, Gebremedhin G, Roman RJ. Functional hyperemia in the brain: hypothesis for astrocyte-derived vasodilator metabolites. Stroke. 1998;28:229–234. doi: 10.1161/01.str.29.1.229. [DOI] [PubMed] [Google Scholar]
- 20.Harder DR, Zhang C, Gebremedhin D. Astrocytes function in matching blood flow to metabolic activity. News Physiol Sci. 2002;17:27–31. doi: 10.1152/physiologyonline.2002.17.1.27. [DOI] [PubMed] [Google Scholar]
- 21.Hermann A, Erxleben C. Charybdotoxin selectively blocks small calcium-activated K+ channels in Aplysia neurons. J Gen Physiol. 1987;90:27–47. doi: 10.1085/jgp.90.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Holzwarth JA, Gibbons SJ, Brorson JR, Philipson LH, Miller RJ. Glutamate receptor agonists stimulate diverse calcium responses in different types of cultured rat cortical glial cells. J Neurosci. 1994;14:879–891. doi: 10.1523/JNEUROSCI.14-04-01879.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Iino M, Goto K, Kakegawa W, Okado H, Sudo M, Ischiuchi S, Miwa A, Takayasu Y, Saito I, Tszuki K, Ozawa S. Glia-synapse interaction through Ca2+-permeable AMPA receptors in Bergmann glia. Science. 2001;292:926–929. doi: 10.1126/science.1058827. [DOI] [PubMed] [Google Scholar]
- 24.Isaacson JS, Murphy GJ. Glutamate-mediated extrasynaptic inhibition: direct coupling of NMDA receptors to Ca2+-activated K+ channels. Neuron. 2001;51:1027–1034. doi: 10.1016/s0896-6273(01)00428-7. [DOI] [PubMed] [Google Scholar]
- 25.Johansson S, Druzin M, Haage D, Wang M-D. The functional role of a bicuculline-sensitive Ca2+-activated K+ current in rat medial preoptic neurons. J Physiol (Lond) 2001;532:625–635. doi: 10.1111/j.1469-7793.2001.0625e.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kandel ER, Schwartz JH, Jessell TM. Principle of neural science, Ed 4. McGraw-Hill; New York: 2000. [Google Scholar]
- 27.Krishnan R, Timothy HM, Guo-Jian J, Hakim H, Paul AF. A molecular mechanism for electrical tuning of cochlear hair cells. Science. 1999;283:215–217. doi: 10.1126/science.283.5399.215. [DOI] [PubMed] [Google Scholar]
- 28.Lang DG, Ritchie AK. Large and small conductance calcium-activated potassium channels in the GH3 anterior pituitary cell line. Pflügers Arch. 1987;410:614–622. doi: 10.1007/BF00581321. [DOI] [PubMed] [Google Scholar]
- 29.Lee K, Rowe ICM, Ashford LMJ. Characterization of an ATP-modulated large conductance Ca2+-activated K+ channel present in rat cortical neurons. J Physiol (Lond) 1995;488:319–337. doi: 10.1113/jphysiol.1995.sp020969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Marrion NV, Tavalin SJ. Selective activation of Ca2+-activated K+ channels by colocalized Ca2+ channels in hippocampal neurons. Nature. 1998;395:900–905. doi: 10.1038/27674. [DOI] [PubMed] [Google Scholar]
- 31.Matsas R, Tsacopoulos M. The functional roles of glial cells in health and disease: dialogue between glia and neurons. Kluwer Academic/Plenum; New York: 1999. [Google Scholar]
- 32.Meera P, Wallner M, Toro L. A neuronal β subunit (KCNMB4) makes the large conductance, voltage- and Ca2+-activated K+ channel resistant to charybdotoxin and iberiotoxin. Proc Natl Acad Sci USA. 2000;97:5562–5567. doi: 10.1073/pnas.100118597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Muller T, Möller T, Berger T, Schnitzer J, Kettenmann H. Calcium entry through kinate receptors and resulting potassium-channel blockade in Bergmann glial cells. Science. 1992;256:563–566. doi: 10.1126/science.1317969. [DOI] [PubMed] [Google Scholar]
- 34.Muller T, Möller T, Neuhaus J, Kettenmann H. Electrical coupling among Bergmann glia cells and its modulation by glutamate receptor activation. Glia. 1996;17:274–284. doi: 10.1002/(SICI)1098-1136(199608)17:4<274::AID-GLIA2>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 35.Nakanishi S. Molecular diversity of glutamate receptors and implications for brain function. Science. 1992;258:597–603. doi: 10.1126/science.1329206. [DOI] [PubMed] [Google Scholar]
- 36.Nelson MT, Quayle JM. Physiological roles and properties of potassium channels in arterial smooth muscle. Am J Physiol. 1995;268:C799–C822. doi: 10.1152/ajpcell.1995.268.4.C799. [DOI] [PubMed] [Google Scholar]
- 37.Nithipatikom K, Grall AJ, Holmes BB, Harder DR, Falck JR, Campbell WB. Liquid chromatographic-electrospray ionization-mass spectrometric analysis of cytochrome P450 metabolites of arachidonic acid. Anal Biochem. 2001;298:327–336. doi: 10.1006/abio.2001.5395. [DOI] [PubMed] [Google Scholar]
- 38.Paulson OB, Newman EA. Does the release of potassium from astrocyte endfeet regulate cerebral blood flow? Science. 1987;237:896–898. doi: 10.1126/science.3616619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Reinhart PH, Chung S, Levitan IB. A family of calcium-dependent potassium channels from rat brain. Neuron. 1989;2:1031–1041. doi: 10.1016/0896-6273(89)90227-4. [DOI] [PubMed] [Google Scholar]
- 40.Robert A, Magistretti PJ. AMPA/kinate receptor activation blocks K+ currents via internal Na+ increase in mouse cultured stellate astrocytes. Glia. 1997;20:38–50. [PubMed] [Google Scholar]
- 41.Rudy B. Diversity and ubiquity of K+ channels. Neuroscience. 1988;25:729–749. doi: 10.1016/0306-4522(88)90033-4. [DOI] [PubMed] [Google Scholar]
- 42.Rziglinski BA, Willoughby KA, Hoffman SW, Falck JR, Ellis EF. Calcium influx factor, further evidence it is 5,6-epoxyeicosatrienoic acid. J Biol Chem. 1999;274:175–182. doi: 10.1074/jbc.274.1.175. [DOI] [PubMed] [Google Scholar]
- 43.Schipke CG, Ohlemeyer C, Matyash M, Nolte C, Kettenmann H, Kirchhoff F. Astrocytes of the mouse neocortex express functional N-methyl-d-aspartate receptors. FASEB J. 2001;15:270–272. doi: 10.1096/fj.00-0439fje. [DOI] [PubMed] [Google Scholar]
- 44.Steinhauser C, Gallo V. News on glutamate receptors in glial cells. Trends Neurosci. 1996;19:339–345. doi: 10.1016/0166-2236(96)10043-6. [DOI] [PubMed] [Google Scholar]
- 45.Talvenheimo JA, Lam G, Gelband C. Charybdotoxin inhibits the 250 pS Ca2+-activated K+ channel in aorta and contracts aorta smooth muscle. Biophys J. 1988;53:258. [Google Scholar]