

Abstract
The generation of new neurons in the adult mammalian hippocampus is thought to play a role in repairing the brain after injury. Here, we show that 7 d after focal cerebral ischemia, newly divided cells in the dentate gyrus of adult rats increased to approximately sevenfold, compared with sham controls. In the same area, this enhanced dentate neurogenesis was associated with activation of inducible nitric oxide synthase (iNOS). Inhibition of iNOS by aminoguanidine prevented ischemia-induced neurogenesis in the dentate gyrus. In null mutant mice lacking the iNOS gene, increased neurogenesis was not observed after focal cerebral ischemia. This study demonstrates that expression of iNOS is necessary for ischemia-stimulated cell birth in the dentate gyrus and indicates that activation of iNOS may provide a possible strategy for functional recovery from cerebral ischemic insult.
Keywords: neurogenesis, dentate granule neurons, focal cerebral ischemia, iNOS expression, iNOS activity, RT-PCR, green fluorescent protein
Introduction
Several thousand new neurons are produced each day in the adult hippocampal dentate gyrus (Kaplan and Hinds, 1977; Eriksson et al., 1998; Kornack and Rakic, 1999). Some of these new nerve cells are functionally recruited in the dentate gyrus circuitry and form appropriate synapses with already existing neurons (Paton and Nottebohm, 1984; Cameron and McKay, 2001; van Praag et al., 2002). It has been suggested that these new neurons play an important role in behavioral plasticity, including learning (Shors et al., 2001). These findings suggest that neurogenesis in the adult dentate gyrus occurs in response to diverse physiological stimuli, including stress (Gould et al., 1998). The number of proliferating cells is also increased in the adult rat dentate gyrus after both stroke (Liu et al., 1998; Takagi et al., 1999; Jin et al., 2001; Kee et al., 2001) and seizures (Bengzon et al., 1997). These findings suggest that neurogenesis in this area of the brain may be a critical element in brain repair. However, the molecular and cellular events underlying enhanced cell proliferation after cerebral ischemic stroke are unclear.
A recent study shows that nitric oxide (NO), a short-lived diffusible molecule produced from arginine by NO synthase (NOS), is involved in neurogenesis in the adult hippocampus (Zhang et al., 2001). Three distinct members of the NOS family in mammalian cells have been identified (Alderton et al., 2001). Both neuronal NOS (nNOS) and the endothelial NOS (eNOS) are constitutively expressed in many tissues (Thomas and Feron, 1997), whereas inducible NOS (iNOS) is usually not expressed in the CNS except in the inflammatory state (Heneka and Feinstein, 2001). Our previous studies in mice demonstrated that iNOS is induced in the core of focal cerebral ischemia (Zhu et al., 2002), but whether expression of iNOS mediates enhanced neurogenesis in the dentate gyrus of adult rats after a cerebral ischemic insult remains unexplored. In the present study, using a combined pharmacological and genetic approach, we determined that activation of iNOS in the dentate gyrus fulfills a necessary condition for it to be considered a mediator of ischemia-induced neurogenesis.
Materials and Methods
Focal cerebral ischemia. Sprague Dawley rats (120 ± 10 d old; ∼295 ± 20 gm) and 90 ± 5 d old homozygous iNOS-deficient mice (iNOS−/−mice; 34 ± 6 gm; C57/BL/6-NOS2tm1Lau; Jackson Laboratories, Bar Harbor, ME) and their wild-type littermates of similar genetic background (C57/BL/6 iNOS+/+ mice; 35 ± 5 gm; Jackson Laboratories) were used in this study. iNOS−/−mice showed undetectable levels of iNOS activity (see Fig.1D). Anesthesia was induced in animals with ketamine (100 mg/kg, i.p.) and xylazine (5 mg/kg, i.p.). Body temperature was maintained at 37 ± 0.5°C with heating pads until the animals had recovered from surgery. Focal cerebral ischemia was induced by intraluminal middle cerebral artery (MCA) occlusion, as described previously (Zhu et al., 2002). In brief, a 4/0 (for rats) or 8/0 (for mice) surgical nylon monofilament with rounded tip was introduced into the left internal carotid through the external carotid stump, advanced 20–21 mm (in rats) or 16–17 mm (in mice) past the carotid bifurcation. The filament was left in place for 90 min and then withdrawn. The sham-operated animals were treated identically, except that the MCAs were not occluded after the neck incision. The infarct volume was determined in both iNOS−/−and iNOS+/+ mice from the experiments with sham operation or ischemia, as described previously (Zhu et al., 2002). In brief, brains were removed rapidly and frozen at −20°C for 5 min. Coronal slices were made at 1–2 mm from the frontal tips, and sections were immersed in 2% 2,3,5-triphenyltetrazolium chloride (TTC) at 37°C for 20 min. The presence or absence of infarctions was determined by examining TTC-stained sections for the areas that did not stain with TTC. The infarct size was expressed as the percentage of the coronal section area.
Fig. 1.

Focal cerebral ischemia induces iNOS expression in the ipsilateral dentate gyrus. A, iNOS mRNA (a1) was examined by RT-PCR using RNA samples from the dentate gyrus of rats after focal cerebral ischemic insult at the indicated days. Sham-operated rats were used as control (Ctr). A PCR product 40 bp smaller than the band derived from the iNOS cDNA was used as iNOS internal standard (20 fg). Similar results were observed in each of three experiments and are summarized in a2. B, Immunoprecipitation (b1) of protein samples from the dentate gyrus of rats after focal cerebral ischemia at the indicated days with polyclonal mouse anti-iNOS. Blots were probed with monoclonal rabbit anti-iNOS. Similar results were observed in each of three experiments and are summarized in b2. C, Ischemia increases iNOS activity in the ipsilateral (filled bars) but not contralateral (open bars) dentate gyrus.D, NOS activity is not altered after ischemia in the dentate gyrus of iNOS null mutant mice (−/−). *p< 0.01, compared with controls (two-tail t test).
RNA extraction and reverse transcription-PCR. Total RNA was extracted from the dentate gyrus of rats that either suffered focal cerebral ischemic insult or sham operation (control) using Trizol reagent according to the manufacturer's instructions (Sigma, St. Louis, MO). The primers for iNOS reverse transcription (RT)-PCR were as follows: forward, 5′-CTGCATGGAACAGTATAAGGCAAAC-3′; reverse, 5′-CAGACAGTTTCTGGTCGATGTCATGA-3′. PCR conditions were 35 cycles of denaturation at 95°C for 30 sec, annealing at 63°C for 45 sec, and extension at 72°C for 45 sec. A 40 bp smaller internal iNOS standard was included in the PCR mixture as an iNOS internal standard. PCR products were separated by electrophoresis through 2% agarose containing 0.5 μg/ml ethidium bromide and imaged using a BioDoc-IT imaging system (Bio-Rad, Hercules, CA); band intensities were determined using GS-710 calibrated imaging Densitometer (Bio-Rad). The ratio of the iNOS mRNA and its internal standard was calculated.
Immunoprecipitation and iNOS activity assay. The dentate gyrus of rats that suffered either ischemic insult or the sham operation was microdissected as described previously (Lu et al., 1998) and homogenized in ice-cold lysis buffer containing 50 mm tris-HCl, pH 7.6, 150 mmNaCl, 1% NP-40, 2 mm EDTA, 1 mm sodium orthovanadate, and proteinase inhibitor mixture (Sigma; 5 μl/100 mg tissue). After debris was cleared by centrifuging at 14,000 × g at 4°C, protein concentration in the extracts was determined by Bradford assay (Bio-Rad). The extracts (∼500 μg protein) were incubated with polyclonal mouse anti-iNOS (2 μg; PharMingen, San Diego, CA) overnight at 4°C, followed by the addition of 40 μl of Protein G-Sepharose (Sigma) for 3 hr at 4°C. Immunoprecipitates were washed four times with lysis buffer and denatured with SDS sample buffer and separated by 12% SDS-PAGE. Proteins were transferred onto nitrocellulose membranes using a Bio-Rad mini-protein-III wet transfer unit overnight at 4°C. Transfer membranes were then incubated with blocking solution (5% nonfat dried milk dissolved in TBST buffer (pH 7.5, 10 mm Tris-HCl, 150 mmNaCl, and 0.1% Tween 20) for 1 hr at room temperature, washed three times, and incubated with polyclonal rabbit anti-iNOS (1: 2000; BD Transduction Laboratories, Lexington, KY) for 1 hr at room temperature. Membranes were washed three times with TBST buffer and incubated with the secondary antibodies (1:1000 dilution) for 1 hr followed by washing four times. Signal detection was performed with an enhanced chemiluminescence kit (Amersham Biosciences, Arlington Heights, IL). The band intensities were determined using a GS-710 calibrated imaging Densitometer (Bio-Rad). The ratio of the iNOS band intensities to the amount of loading proteins of the immunoprecipitates was calculated. The activity of iNOS was determined by a [14C]l-arginine conversion method (Zhu et al., 2002). The iNOS activity is defined as the difference between the EDTA-containing reaction system and the reaction system containing both EDTA and nonspecific NOS inhibitor NG-monomethyl-l-arginine (Sigma) at 50 nm. The protein content of the sample was determined using a total protein kit (Sigma). The iNOS activity (unit) was expressed as picomoles per milligram of protein per minute (1 U).
BrdU labeling. Animals were treated intraperitoneally with 50 mg/kg 5-bromodeoxyuridine (BrdU; Sigma) two consecutive times at 8 hr intervals 1, 7, 14, and 21 d after ischemia and killed the next day to estimate the number of proliferating cells. In the experiments of Figure 4, animals were killed 4 weeks after the BrdU injection to determine the phenotypes of BrdU-labeled cells. In the experiments with iNOS inhibitor, the rats were injected intraperitoneally with 100 mg/kg aminoguanidine (AG; Sigma) five times (5 consecutive days) immediately after ischemia. The rats were then injected with 50 mg/kg BrdU two consecutive times at 8 hr intervals immediately after the last AG injection. The rats were killed the next day for BrdU staining.
Fig. 4.
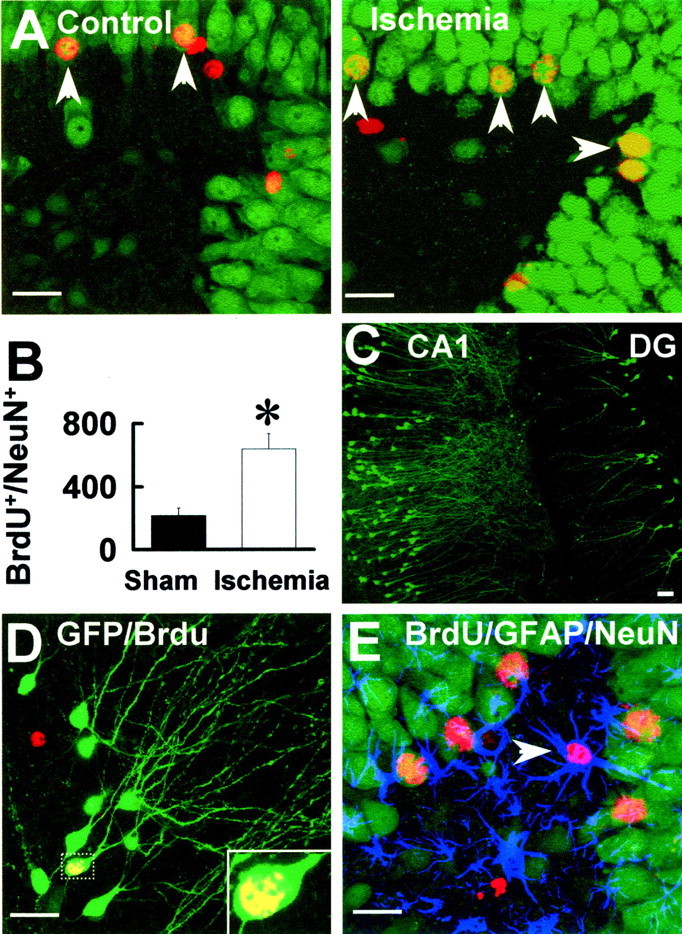
Neuronal identity of BrdU+ cells in the dentate gyrus of ischemic rats.A, Rats were treated with BrdU 7 d after sham operation or focal cerebral ischemia and killed 4 weeks after the BrdU injection. The hippocampal sections were double stained with BrdU (red) and NeuN (green).Arrows indicate the double-labeling cells.B, Summarized numbers of BrdU+/NeuN+ cells.C, Expression of GFP in the CA1 and dentate gyrus (DG) neurons with infusion of pSFV(pd)–GFP infectious vector into the rat hippocampus. D, The newly generated granule cells were identified by expressing GFP. Arrow indicates a GFP+(green)/BrdU+(red) double-labeling granule neuron.E, No colocalization of GFAP with BrdU+/NeuN+ cells in the granule cell layer. Arrow indicates a GFAP+(blue)/BrdU+(red) double-labeling cell in the hilus. Scale bars, 30 μm.
Immunocytochemistry. BrdU staining has been described previously (Liu et al., 2002). The sections were heated (85°C for 5 min) in antigen unmasking solution (Vector Laboratories, Burlingame, CA), incubated in 2 m HCl (room temperature for 30 min), rinsed in 0.1 m boric acid, pH 8.5, for 10 min, incubated in 1% H2O2 in PBS for 30 min, and blocked in 3% normal goat serum/0.3% Triton X-100/0.1% BSA in PBS (room temperature for 1 hr), followed by incubation with rat monoclonal anti-BrdU (1:200; Accurate Chemicals, Westbury, NY) at 4°C overnight. Subsequently, the sections were developed with an ABC kit (Vector). For double labeling, the sections were incubated with the affinity-purified second antibody, goat anti-rat Cy3 antibody (1:200; Chemicon, Temecula, CA). These sections were then incubated in mouse anti-NeuN (1:100; Chemicon) and reacted with goat anti-mouse fluorescein (1:50; Chemicon). For triple staining, the sections were further incubated with rabbit anti-GFAP (1:1000; Sigma). These sections were then incubated with goat anti-rabbit Cy5 (1:100; Chemicon). Sections were rinsed, dried, and coverslipped with Dako (Dako, Carpinteria, CA) fluorescence mounting medium. Control sections were processed with omission of the primary antisera. Double or triple labeling was imaged with a confocal laser scanning microscope (OlympusLSM-GB200) and analyzed with a three-dimensional (3D) constructor (Image-Pro Plus software). We produced 3D digital reconstructions from a series of confocal images taken at 0.5 μm intervals through the region of interest, and optical stacks of 6–12 images were produced for the figures.
Cell counting. An experimenter (D.Y.Z.) coded all slides from the experiments with sham operation or ischemia before quantitative analysis. Stereological analysis of total number of BrdU+ cells in the dentate gyrus was performed by another experimenter (H.S.S.) who was unaware of the experimental condition. The analysis was conducted using a modified version of the optical fractionator method on every sixth section of peroxide staining tissue. The number of BrdU+ nuclei in each section was divided by the area of the dentate gyrus that was determined using Stereoinvestigator. Double-stained numbers of BrdU+/NeuN+and BrdU+/GFAP+cells were analyzed by sampling every section from the experimental animals using a fluorescence microscope as described above.
Construction of Semliki Forest Virus-green fluorescent protein vectors and packaging of the recombinant virions. Construction of nontoxic virus vectors with a high infection rate in hippocampal neurons has been described previously (Liu et al., 2002). In brief, the Sacl–Xbal fragment from Semliki Forest Virus plasmid 1 (pSFV1) (Invitrogen, Gaithersburg, MD) was subcloned into the pGEM7Zf+ vector (Promega, Madison, WI). A PCR-based site-directed mutagenesis was used to change Ser259 to Pro (p) and Arg650 to Asp (d) in the nsP2 fragment (pd mutation). Subsequently, the original fragments in pSFV1 were replaced by the mutated fragments to obtain pSFV(pd) vectors. Mutations were confirmed by sequencing. A cDNA encoding enhanced green fluorescent (GFP) (Clontech, Cambridge, UK) was then inserted directly into pSFV(pd) vector to produce pSFV(pd)–GFP constructs. In vitrotranscribed RNA molecules from pSFV(pd)–GFP were cotransfected with pSFV-helper2 RNA (a gift from Markus U. Ehrengruber, University of Zurich) into baby hamster kidney cell line 21 (BHK-21) cells. All virus production was performed at 31°C. Twenty-four hours after electroporation, virus stocks were harvested, filter sterilized, and activated with Chymotrypsin A4 (Invitrogen). The reaction was terminated with the trypsin inhibitor aprotinin (Invitrogen) before use. Final virus titers (≥109 infectious units/ml) were determined by infection of BHK-21 cells with serial dilutions of virus stocks, followed by fluorescence microscopy examination at 3 d after infection.
Activated pSFV(pd)–GFP virus particles (2 μl at 0.2 μl/min) were infused (26 d after the last BrdU injection) into the left side of the hippocampus with an injection site 2 mm posterior to bregma, 1.5 mm lateral to the midline, and 2 mm below dura. Two days after infection, animals were fixed under Nembutal anesthesia with 4% paraformaldehyde in 0.1 m phosphate buffer for BrdU staining.
Results
Focal cerebral ischemia increases iNOS mRNA levels and enzymatic activity
In the adult hippocampal dentate gyrus, there was a low level of iNOS expression in control animals (Fig.1A). The iNOS mRNA level in the ipsilateral dentate gyrus began to increase 6 hr (data not shown) after focal cerebral ischemia and reached a peak 2 d later (Fig. 1A). Semiquantitative PCR analysis showed that the peak level of iNOS mRNA in the ipsilateral dentate gyrus of the ischemic rats was increased by ∼2.6-fold, compared with sham controls (Fig. 1A). Increased iNOS mRNA expression remained elevated for 3 d and then declined to control level 4 d later. In agreement with the mRNA results, increased iNOS protein content in the ipsilateral dentate gyrus of the ischemic rats was also observed (Fig. 1B). Consistent with this increase of iNOS expression, the enzymatic activity of iNOS was also stimulated. Thus, 2 d after ischemia, l-arginine conversion to NO in the ipsilateral dentate gyrus was 6.48 ± 0.43 U (Fig. 1C) (mean ± SEM; n = 5 rats), whereas in the sham control animals, the enzymatic activity of iNOS was only 1.68 ± 0.11 U (mean ± SEM; n = 4 rats). However, in the contralateral dentate gyrus of these ischemic rats, neither the iNOS mRNA level nor its enzymatic activity was changed at any time point, showing that focal cerebral ischemia upregulates iNOS activity only in the ipsilateral dentate gyrus. To confirm that enhanced l-arginine conversion was mediated by iNOS, we used mutant mice that lack the iNOS gene (iNOS−/−mice). In these animals, iNOS mRNA and iNOS protein were not detected, and 2 d after ischemia, l-arginine conversion in the ipsilateral dentate gyrus was only 1.46 ± 0.14 U (Fig. 1D) (mean ± SEM, n = 5 mice), compared with 1.32 ± 0.16 U (mean ± SEM;n = 5; p > 0.1) in sham-operation iNOS+/+ mice.
Focal cerebral ischemia stimulates cell proliferation
Next, in rats receiving the focal cerebral ischemia, we examined BrdU incorporation into dentate gyrus neurons. We treated separate cohorts of rats 1, 7, 14, and 21 d after ischemia with BrdU twice with an 8 hr interval between injections. The BrdU-treated rats were killed on the following day to estimate the number of BrdU-positive (BrdU+) cells. As can be seen (Fig.2A), the basal level of BrdU+ cells in the dentate gyrus was unchanged in the sham controls, or 1 d after focal cerebral ischemia (Fig. 2B), indicating that the stress of surgery did not alter cell proliferation. However, the number of BrdU+ cells increased in the ipsilateral but not in the contralateral dentate gyrus 3 d after focal cerebral ischemia and reached a peak 7 d later (Fig.2C). The number of BrdU+cells then decreased by day 21, but this number was still greater than sham control (Fig. 2D). Cell birth returned to the basal level in the dentate gyrus of rats 4 weeks after ischemia (Fig.3).
Fig. 2.
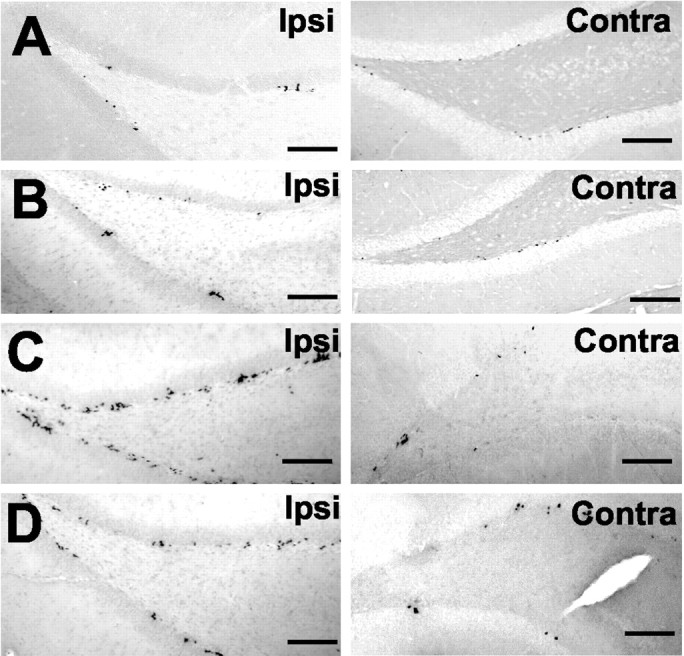
Ischemia increases cell proliferation in the ipsilateral dentate gyrus of adult rats. Rats were treated with BrdU after sham operation (A), or 1 d (B), 7 d (C), or 21 d (D) after focal cerebral ischemia. The next day after the BrdU injection, rats were killed, and the ipsilateral (Ipsi) and contralateral (Contra) hippocampi were processed for BrdU staining. Scale bars, 200 μm.
Fig. 3.

Time course of cell birth in the dentate gyrus after focal cerebral ischemia. In this series of the experiments, rats were treated with BrdU after sham operation (Control;n = 10), or 1 d (n = 10), 3 d (n = 9), 7 d (n = 11), or 21 d (n = 8) after focal cerebral ischemia. The next day after the BrdU injection, rats were killed, and the ipsilateral (filled bars) and contralateral (open bars) hippocampi were processed for BrdU staining. Data are mean ± SEM; *p < 0.01 compared with control (two-tail t test).
Focal cerebral ischemia increases the number of new neurons
To determine the fate of the BrdU+cells, we used antibodies against NeuN, a transcription factor that is expressed in mature neurons. By performing three-dimensional digital reconstruction, we found that 68% of the BrdU+ cells in the granule cell layer were colabeled with NeuN (Fig.4A,BrdU+/NeuN+) and therefore were differentiated neurons. In this series of experiments, animals received BrdU injection 7 d after ischemia and were killed 4 weeks later. At this time point, the number of BrdU+ cells in the ipsilateral dentate gyrus after focal cerebral ischemia was approximately threefold of the sham control (Fig. 4B).
To establish the neuronal identity of these BrdU+ cells, we developed mutant SFV (mSFV) vectors to express enhanced into neurons in the hippocampus (Fig. 4C). We found that a large number of the BrdU+ cells expressed GFP (BrdU+/GFP+) in the granule cell layer that had the morphological characteristics of a granule cell, i.e., ∼20 μm diameter round or oval cell body with long dendrites extending toward the molecular layer (Fig.4D). These data provide further evidence that most of the new cells in the dentate gyrus of rats that received focal cerebral ischemia matured into granule neurons.
Glia cell generation after focal cerebral ischemia
To exclude the possibility that the BrdU+ cells expressed in the dentate gyrus were glia cells, we triple stained sections with antibodies to GFAP. We found that none of the BrdU+/NeuN+cells in the granule cell layer of either control or ischemic animals were labeled with GFAP (Fig. 4E). Most of the GFAP-stained astrocytes were located in the hilus and not in the granule cell layer. Semiquantitative analysis showed that the number of BrdU+ cells in the hilus of the ischemic rats was not significantly different from that in the control (data not shown). This contrasts with the granule cell layer in which most post-ischemic BrdU+ cells survived and differentiated into mature granule cells.
Inhibition of iNOS prevents ischemia-induced cell birth
Next, we determined whether ischemia-activated iNOS is required for increased neurogenesis. We therefore treated rats with the iNOS inhibitor AG for 5 d at 24 hr intervals immediately after the ischemic insult. Our previous study in mice showed that this treatment significantly reduced the size of the infarct (Zhu et al., 2002). As can be seen (Fig. 5A), we observed enhanced iNOS expression in the ipsilateral dentate gyrus in these animals 3 d after ischemia. In contrast, there was no increased l-arginine conversion in AG-treated ischemic rats (Fig. 5B). iNOS activity in the sham control and AG-treated ischemic rats was 1.37 ± 0.12 U (mean ± SEM;n = 5) and 1.39 ± 0.16 U (mean ± SEM;n = 6), respectively. To characterize cell proliferation in the AG-treated rats, rats were injected with BrdU immediately after the last AG injection and killed the following day for BrdU staining. Representative BrdU+cell nuclei in the ipsilateral dentate gyrus of AG- and vehicle-treated ischemic rats are shown in Figure 6,A and B. The average number of BrdU+ cells in the ipsilateral dentate gyrus of AG-treated ischemic rats was not statistically different from that in AG-treated sham control rats (Fig. 6C) (p > 0.01; two-tailed t test).
Fig. 5.
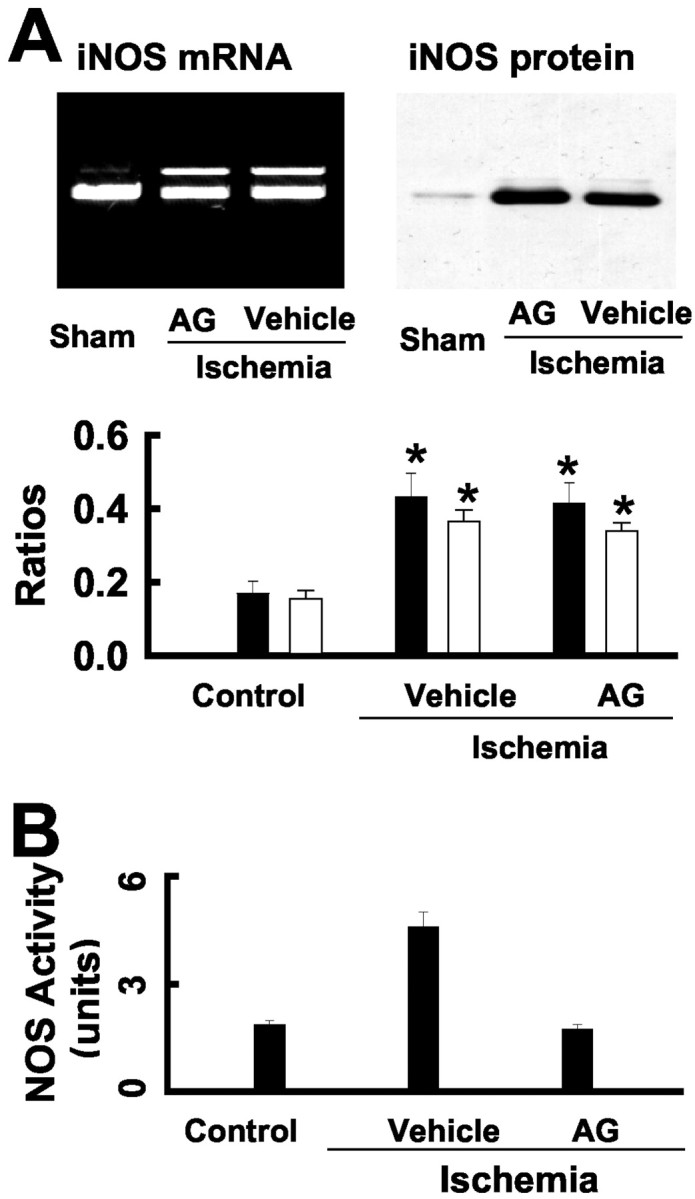
AG inhibits iNOS activity. A, AG did not alter iNOS expression. iNOS mRNA (top) was examined by RT-PCR using RNA samples, and blots (bottom) were probed with anti-iNOS. The samples of RNA and the protein were from the dentate gyrus of AG-treated ischemic rats. Similar results are observed in each of three experiments. Bar graph shows iNOS mRNA (filled bars) and iNOS protein (open bars) in the ipsilateral dentate gyrus of rats (n = 3 animals in each group). B, AG inhibits iNOS activity in the ipsilateral dentate gyrus. *p < 0.01, compared with control (two-tailt test).
Fig. 6.

Pharmacological inhibition of iNOS prevents ischemia-induced cell birth. Representatives of the BrdU labeling in the ipsilateral dentate gyrus of AG- (A), and vehicle-treated (B) rats that received sham-operation or focal cerebral ischemia (isc). Scale bars, 200 μm. C, Bar graph shows the summarized data of BrdU+ cells in the ipsilateral dentate gyrus of AG- (filled bars) and vehicle-treated (open bars) rats receiving sham operation or focal cerebral ischemia (n = 8 animals in each group). Data are mean ± SEM; *p < 0.01 compared with sham-operation control (two-tail t test).
We also investigated BrdU+ cells in the dentate gyrus of the iNOS−/−mice. A previous study shows that iNOS−/−mice are resistant to ischemic injury. Consistent with those data, we found that the infarct size in the iNOS−/−mice 2 d after focal cerebral ischemia was only 15 ± 4%, compared with 36 ± 8% in the iNOS+/+ mice (Fig.7A). iNOS−/− mice showed no increase of BrdU+ cells in the dentate gyrus after the ischemic insult (Fig.7B,C). The number of BrdU+ cells in the dentate gyrus of iNOS−/−and their wild-type littermates is summarized in Figure 7D. These data confirm that activation of iNOS induced by focal cerebral ischemia is necessary for enhanced cell proliferation in the dentate gyrus.
Fig. 7.
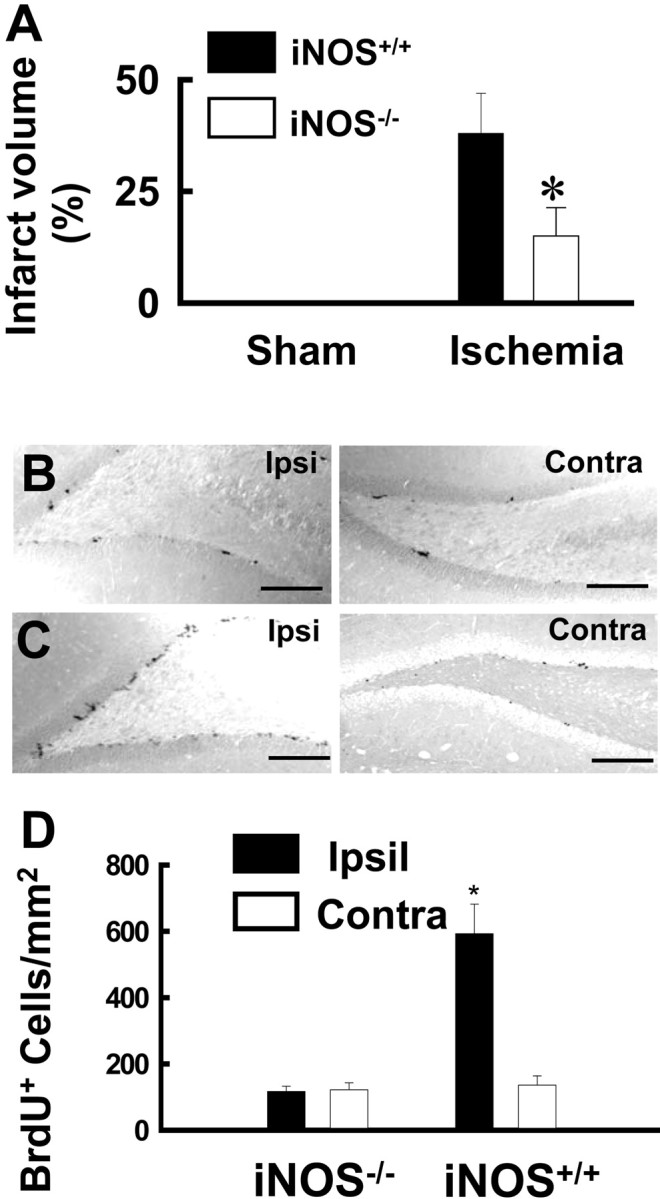
Genetic inhibition of iNOS prevents ischemia-induced neurogenesis. A, The infarct size in iNOS−/− mice (n = 8) and iNOS+/+ mice (n = 8) 2 d after focal cerebral ischemia. Data are mean ± SEM; *p < 0.01 compared with iNOS+/+ mice (two-tail t test).B, Representatives of BrdU labeling in the dentate gyrus of ischemic iNOS−/− mice are shown. C, Representatives of BrdU labeling in the dentate gyrus of ischemic iNOS+/+ mice are shown. Scale bars, 200 μm. D, The number of BrdU+ cells in the ipsilateral (filled bars) and contralateral (open bars) dentate gyrus of iNOS−/−(n = 10) and iNOS+/+ mice (n = 10) is summarized. Data are mean ± SEM; *p < 0.01 compared with control (two-tailt test).
Discussion
In the dentate gyrus of rats challenged with an ischemic insult, we found that there was a robust expression of iNOS coupled with an increase in its enzymatic activity. These changes were associated with increased cell proliferation in the dentate gyrus. We further showed that inhibition of endogenous iNOS by the iNOS inhibitor AG prevented the ischemia-induced increase in neurogenesis. Consistent with this finding are our data showing that ischemia in iNOS null mutant mice resulted in significantly fewer BrdU+cells in the dentate gyrus, compared with their wild-type ischemic littermates. Taken together, our data provide the first evidence showing that activation of iNOS is crucial for ischemia-induced neurogenesis.
Our findings of iNOS-dependent neurogenesis are in agreement with a recent study showing that there are increased BrdU+ cells in the dentate gyrus of rats that received a NO donor (Zhang et al., 2001). Although NO sometimes may be associated with deleterious effects in the CNS (Ye et al., 1996), there are many instances in which NO plays a beneficial role, including synaptic plasticity (Bon et al., 1992; Mori et al., 2001) and the formation of learning and memory (Papa et al., 1994). Our data show that activation of iNOS, which would lead to the production of NO, may contribute to the events that lead to brain repair. Thus, the increased expression of iNOS in the dentate gyrus (Fig. 1A) was associated with enhanced cell proliferation (Fig. 2). In keeping with the beneficial effects of iNOS, we found that activation of iNOS not only increased the number of proliferating cells but also increased the number of mature granule neurons (Fig. 4). Because the changes in iNOS activity in the dentate gyrus preceded cell proliferation induced by ischemia, we suggest that activation of iNOS is the primary event for initiating neurogenesis.
There are two constitutive forms of NOS (cNOS) in the CNS (Thomas and Feron, 1997). Because cNOS is still available to provide NO after inhibition of iNOS, reduced neurogenesis in iNOS-deficient mice may not be caused simply by a decreased production of NO. Interestingly, our earlier study showed that the enzymatic activity of cNOS in the cerebral cortex of mice is decreased after focal cerebral ischemia (Zhu et al., 2002). It is therefore possible that reduction of cNOS activity induces iNOS expression, thus providing the diffusible messenger, NO, to stimulate cell proliferating factors in the hippocampus. Consistent with this hypothesis, a recent study demonstrates that iNOS expression is suppressed by the basal activity of cNOS under normal circumstances and that inhibition of cNOS induces iNOS expression in the rat small intestine (Qu et al., 2001). Thus, it will be of importance to determine whether iNOS-dependent neurogenesis can be induced in the hippocampus by inhibition of cNOS.
Direct and indirect evidence suggests that brain injury stimulates the proliferation of endogenous progenitors in the hippocampus (Gould and Tanapat, 1997; Liu et al., 1998; Jin et al., 2001; Yoshimura et al., 2001). There is precedent for inhibition of iNOS reducing the ischemic infarct size in mice (Huang et al., 1994; Samdani et al., 1997; Ashwal et al., 1998; Zhu et al., 2002). Thus, it may be argued that reduced neurogenesis by inhibition of iNOS is caused simply by a reduced ischemic injury. As such, iNOS expression after focal cerebral ischemia may be involved directly in neuronal injury but indirectly in cell proliferation; however, our findings are not consistent with this interpretation. First, although inhibition of iNOS by the iNOS inhibitor AG reduced the size of the infarct after focal cerebral ischemia (Zhu et al., 2002), iNOS expression showed no difference between AG- and vehicle-treated ischemic animals (Fig.5A). Second, increased numbers of BrdU+ cells were observed before the hippocampal injury. For example, the damage to the CA1 neurons occurs 7 d after focal cerebral ischemia (Zhu and Auer, 1995; Nakatomi et al., 2002), whereas iNOS-dependent cell birth was observed 4 d before the injury (Fig. 3). Together these data suggest that iNOS expression in the dentate gyrus after ischemic insult is independent of infarct size. Thus, expression of iNOS is directly responsible for enhanced neurogenesis. A further study is required to investigate the underlying mechanisms for inducing iNOS expression.
The biochemical events underlying ischemia-induced neurogenesis in the dentate gyrus are thought to involve activation of NMDA receptors. In support of this notion are the data showing that rats receiving an NMDA receptor antagonist at the time of ischemia show no increased neurogenesis in the dentate gyrus (Nei et al., 1996;Arvidsson et al., 2001; Bernabeu and Sharp, 2000). There are also extensive data showing that NOS activity in the hippocampus is stimulated by activation of NMDA receptors (Cardenas et al., 2000;Jander et al., 2000). Thus, the most parsimonious explanation of our findings is that after focal cerebral ischemia, iNOS is rapidly activated through NMDA receptor-dependent events. We also stained hippocampal sections with an anti-iNOS antibody and observed that iNOS was expressed in both neurons and glia cells in the dentate gyrus after focal cerebral ischemia (data not shown). This finding is consistent with previous studies (Holta et al., 2001) (but seeIadecola et al., 1997). Thus, the expression of iNOS after focal cerebral ischemia would provide the diffusible messenger NO to stimulate neural cell proliferating factors.
Consistent with enhanced neurogenesis after ischemia (Liu et al., 1998;Takagi et al., 1999; Jin et al., 2001; Kee et al., 2001) is our demonstration that there is an approximately threefold increase in the number of newly generated granule neurons in the ipsilateral dentate gyrus of ischemic animals compared with sham controls. There could be some concerns that the BrdU+ cells that we observed were stained during the process of apoptosis. However, the BrdU+ neurons that we observed in the granule cell layer were labeled 4 weeks after the last BrdU injection, without any observable degeneration. Most of these BrdU+/GFP+neurons showed the normal neuronal morphology of mature granule cells (Fig. 4D). There are limitations using the expression of marker proteins to identify new neurons. For example, the most often used antibodies for mitogen-activated protein-2 and turned on after division/collapsing response-mediated protein/Unc-33-like phosphoprotein-4 also react with astrocytes and oligodendrocytes (Sensenbrenner et al., 1997) and are thus inappropriate as specific markers for neurons. To circumvent this limitation, we used mSFV vector expression of enhanced GFP in the dentate gyrus. In combination with BrdU staining, we showed in both ischemic rats and mice that the increased production of new granule neurons requires activation of iNOS in the dentate gyrus. The hippocampus plays a critical role in the formation of declarative memory, and declarative memory formation is impaired in ischemic rats and humans (Squire and Zola-Morgan, 1996). Because cell transplants reduce ischemia-caused memory deficits (Sinden et al., 1997), our study may provide a molecular strategy for improvement of learning and memory function via increased neurogenesis in patients suffering from cerebral ischemic stroke.
Footnotes
This work was supported by grants from the Canadian Institute for Health Research (Y.M.L), Heart and Stroke Foundation, Canada (Y.M.L), Alberta Heritage Foundation for Medical Research (Y.M.L), Canada Foundation for Innovation (Y.M.L), and Alberta Foundation for Innovation and Science (Y.M.L). We thank Drs. Ken Lukowiak and Wayne Giles for critical comments on this manuscript.
Correspondence should be addressed to Dr. You Ming Lu, Department of Physiology and Biophysics, Neuroscience Research Group, Faculty of Medicine, University of Calgary, Calgary, Canada, T2N 4N1. E-mail:luy@ucalgary.ca.
References
- 1.Alderton WK, Cooper CE, Knowles RG. Nitric oxide synthases: structure, function and inhibition. Biochem J. 2001;357:593–615. doi: 10.1042/0264-6021:3570593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Arvidsson A, Kokaia Z, Lindvall O. N-methyl-d-aspartate receptor-mediated increase of neurogenesis in adult rat dentate gyrus following stroke. Eur J Neurosci. 2001;14:10–18. doi: 10.1046/j.0953-816x.2001.01611.x. [DOI] [PubMed] [Google Scholar]
- 3.Bengzon J, Kokaia Z, Elmer E, Nanobashvili A, Kokaia M, Lindvall O. Apoptosis and proliferation of dentate gyrus neurons after single and intermittent limbic seizures. Proc Natl Acad Sci USA. 1997;94:10432–10437. doi: 10.1073/pnas.94.19.10432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bernabeu R, Sharp FR. NMDA and AMPA/kainite glutamate receptors modulate dentate gyrus neurogenesis and CA3 synapsin-I in normal and ischemic hippocampus. J Cereb Blood Flow Metab. 2000;20:1669–1680. doi: 10.1097/00004647-200012000-00006. [DOI] [PubMed] [Google Scholar]
- 5.Bon C, Bohme GA, Doble A, Stutzmann JM, Blanchard JC. A role for nitric oxide in long-term potentiation. Eur J Neurosci. 1992;4:420–424. doi: 10.1111/j.1460-9568.1992.tb00891.x. [DOI] [PubMed] [Google Scholar]
- 6.Cameron HA, McKay RD. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. J Comp Neurol. 2001;435:406–417. doi: 10.1002/cne.1040. [DOI] [PubMed] [Google Scholar]
- 7.Cardenas A, Moro MA, Hurtado O, Leza JC, Lorenzo P, Castrillo A, Bodelon OG, Bosca L, Lizasoain I. Implication of glutamate in the expression of inducible nitric oxide synthase after oxygen and glucose deprivation in rat forebrain slices. J Neurochem. 2000;74:2041–2048. doi: 10.1046/j.1471-4159.2000.0742041.x. [DOI] [PubMed] [Google Scholar]
- 8.Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313–1317. doi: 10.1038/3305. [DOI] [PubMed] [Google Scholar]
- 9.Gould E, Tanapat P. Lesion-induced proliferation of neuronal progenitors in the dentate gyrus of the adult rat. Neuroscience. 1997;80:427–436. doi: 10.1016/s0306-4522(97)00127-9. [DOI] [PubMed] [Google Scholar]
- 10.Gould E, Tanapat P, McEwen B, Flugge G, Fuchs E. Proliferation of granule cell precursors in the dentate gyrus of adult monkeys is diminished by stress. Proc Natl Acad Sci USA. 1998;95:3168–3171. doi: 10.1073/pnas.95.6.3168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Heneka MT, Feinstein DL. Expression and function of inducible nitric oxide synthase in neurons. J Neuroimmunol. 2001;114:8–18. doi: 10.1016/s0165-5728(01)00246-6. [DOI] [PubMed] [Google Scholar]
- 12.Holta ML, Craddock SD, Pettigrew C. Expression of neuronal and inducible nitric oxide synthases during post-ischemic reperfusion in rat brain. Brain Res. 2001;898:49–60. doi: 10.1016/s0006-8993(01)02140-0. [DOI] [PubMed] [Google Scholar]
- 13.Iadecola C, Zhang F, Cassey R, Nagayama M, Ross ME. Delayed reduction of ischemic brain injury and neurological deficits in mice lacking the inducible nitric oxide synthase gene. J Neurosci. 1997;17:9157–9164. doi: 10.1523/JNEUROSCI.17-23-09157.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jander S, Schroeter M, Stoll G. Role of NMDA receptor signaling in the regulation of inflammatory gene expression after focal brain ischemia. J Neuroimmunol. 2000;109:181–187. doi: 10.1016/s0165-5728(00)00317-9. [DOI] [PubMed] [Google Scholar]
- 15.Jin K, Minami M, Lan JQ, Mao XO, Batteur S, Simon RP, Greenberg DA. Neurogenesis in dentate subgranular zone and rostral subventricular zone after cerebral ischemia in the rat. Proc Natl Acad Sci USA. 2001;98:4710–4715. doi: 10.1073/pnas.081011098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kaplan MS, Hinds JW. Neurogenesis in the adult rat: electron microscopic analysis of light radioautographs. Science. 1977;197:1092–1094. doi: 10.1126/science.887941. [DOI] [PubMed] [Google Scholar]
- 17.Kee NJ, Preston E, Woitowicz JM. Enhanced neurogenesis after transient global ischemia in the dentate gyrus of the rat. Exp Brain Res. 2001;136:313–320. doi: 10.1007/s002210000591. [DOI] [PubMed] [Google Scholar]
- 18.Kornack DR, Rakic P. Continuation of neurogenesis in the hippocampus of the adult macaque monkey. Proc Natl Acad Sci USA. 1999;96:5768–5773. doi: 10.1073/pnas.96.10.5768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Liu J, Solway K, Messing RO, Sharp FR. Increased neurogenesis in the dentate gyrus after transient global ischemia in gerbils. J Neurosci. 1998;18:7768–7778. doi: 10.1523/JNEUROSCI.18-19-07768.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liu SH, Wang J, Zhu DY, Fu YP, Lu YM. Generation of functional inhibitory neurons in the adult rat hippocampus. Soc Neurosci Abstr. 2002;25:2. doi: 10.1523/JNEUROSCI.23-03-00732.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lu YM, Roder JC, Davidow J, Salter MW. Src activation in the induction of long-term potentiation in CA1 hippocampal neurons. Science. 1998;279:1363–1367. doi: 10.1126/science.279.5355.1363. [DOI] [PubMed] [Google Scholar]
- 22.Mori K, Togashi H, Ueno KI, Matsumoto M, Yoshioka M. Aminoguanidine prevented the impairment of learning behavior and hippocampal long-term potentiation following transient cerebral ischemia. Behav Brain Res. 2001;120:159–168. doi: 10.1016/s0166-4328(00)00371-5. [DOI] [PubMed] [Google Scholar]
- 23.Nakatomi H, Kuriu T, Okabe S, Yamamoto S, Hatano O, Kawahara N, Tamura A, Kirino T, Nakafuku M. Regeneration of hippocampal pyramidal neurons after ischemic brain injury by recruitment of endogenous neural progenitors. Cell. 2002;110:429–441. doi: 10.1016/s0092-8674(02)00862-0. [DOI] [PubMed] [Google Scholar]
- 24.Nei K, Matsuyama S, Shuntoh H, Tanaka C. NMDA receptor activation induces glutamate release through nitric oxide synthesis in guinea pig dentate gyrus. Brain Res. 1996;728:105–110. [PubMed] [Google Scholar]
- 25.Papa M, Pellicano MP, Sadile AG. Nitric oxide and long-term habituation to novelty in the rat. Ann NY Acad Sci. 1994;738:316–324. doi: 10.1111/j.1749-6632.1994.tb21817.x. [DOI] [PubMed] [Google Scholar]
- 26.Paton JA, Nottebohm F. Neurons generated in the adult brain are recruited into functional circuits. Science. 1984;225:1046–1048. doi: 10.1126/science.6474166. [DOI] [PubMed] [Google Scholar]
- 27.Qu XW, Wang H, De Plaen IG, Rozenfeld RA, Hsueh W. Neuronal nitric oxide synthase (NOS) regulates the expression of inducible NOS in rat small intestine via modulation of nuclear factor kappa B. FASEB J. 2001;15:439–446. doi: 10.1096/fj.99-0343com. [DOI] [PubMed] [Google Scholar]
- 28.Sensenbrenner M, Lucas M, Deloulme JC. Expression of two neuronal markers, growth-associated protein-43 and neuron-specific enolase, in rat glia cells. J Mol Med. 1997;75:653–663. doi: 10.1007/s001090050149. [DOI] [PubMed] [Google Scholar]
- 29.Squire LR, Zola-Morgan S. Ischemic brain damage and memory impairment: a commentary. Hippocampus. 1996;6:546–552. doi: 10.1002/(SICI)1098-1063(1996)6:5<546::AID-HIPO7>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 30.Sinden JD, Rashid-Doubell F, Kershaw TR, Nelson A, Chadwick A, Jat PS, Noble MD, Hodges H, Gray JA. Recovery of spatial learning by grafts of a conditionally immortalized hippocampal neuroepithelial cell line into the ischemia-lesioned hippocampus. Neuroscience. 1997;81:599–608. doi: 10.1016/s0306-4522(97)00330-8. [DOI] [PubMed] [Google Scholar]
- 31.Shors TJ, Miesegaes G, Beylin A, Zhao M, Rydel T, Gould E. Neurogenesis in the adult is involved in the formation of trace memories. Nature. 2001;410:372–376. doi: 10.1038/35066584. [DOI] [PubMed] [Google Scholar]
- 32.Takagi Y, Nozaki K, Takahashi J, Yodoi J, Ishikawa M, Hashimoto N. Proliferation of neuronal precursor cells in the dentate gyrus is accelerated after transient forebrain ischemia in mice. Brain Res. 1999;831:283–287. doi: 10.1016/s0006-8993(99)01411-0. [DOI] [PubMed] [Google Scholar]
- 33.Thomas M, Feron O. Nitric oxide synthases: which, where, and why? J Clin Invest. 1997;100:2146–2152. doi: 10.1172/JCI119750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.van Praag H, Schlinder AF, Christle BR, Toni N, Palmer TD, Gage FH. Functional neurogenesis in the adult hippocampus. Nature. 2002;415:1030–1034. doi: 10.1038/4151030a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ye YZ, Strong M, Huang ZQ, Beckman JS. Physiological and pathological processes. In: Packer L, editor. Nitric oxide. Academic; New York: 1996. pp. 201–209. [Google Scholar]
- 36.Yoshimura S, Takagi Y, Harada J, Teramoto T, Thomas SS, Waeber C, Bakowska JC, Breakefield XO, Moskowitz MA. FGF-2 regulation of neurogenesis in adult hippocampus after brain injury. Proc Natl Acad Sci USA. 2001;98:5874–5879. doi: 10.1073/pnas.101034998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zhang R, Zhang L, Zhang Z, Wang Y, Lu M, Lapointe M, Chopp M. A nitric oxide donor induces neurogenesis and reduces functional deficits after stroke in rats. Ann Neurol. 2001;50:602–611. doi: 10.1002/ana.1249. [DOI] [PubMed] [Google Scholar]
- 38.Zhu CZ, Auer RN. Graded hypotension and MCA occlusion duration: effect in transient focal ischemia. J Cereb Blood Flow Metab. 1995;15:980–988. doi: 10.1038/jcbfm.1995.124. [DOI] [PubMed] [Google Scholar]
- 39.Zhu DY, Deng Q, Yao HH, Wang DC, Deng Y, Liu GQ. Inducible nitric oxide synthase expression in the ischemic core and penumbra after transient focal cerebral ischemia in mice. Life Sci. 2002;17:1985–1997. doi: 10.1016/s0024-3205(02)01970-7. [DOI] [PubMed] [Google Scholar]