

Abstract
AIMS
B-cell acute lymphoblastic leukemia (B-ALL) is amongst the most prevalent cancers of children in Pakistan. Genetic variations in FLT3 are associated with auto-phosphorylation of kinase domain that leads to increased proliferation of blast cells. Paired box family of transcription factor (PAX5) plays a critical role in commitment and differentiation of B-cells. Variations in PAX5 are associated with the risk of B-ALL. We aimed to analyze the association of FLT3 and PAX5 polymorphisms with B cell leukemia in Pakistani cohort.
METHODS
We collected 155 B-ALL subject and 155 control blood samples. For analysis, genotyping was done by tetra ARMS-PCR. SPSS was used to check the association of demographic factors of SNPs present in the population with the risk of B-ALL.
RESULTS
Risk allele frequency A at locus 13q12.2 (rs35958982, FLT3) was conspicuous and showed positive association (OR = 2.30, CI [1.20–4.50], P = 0.005) but genotype frequency (OR = 3.67, CI [0.75–18.10], P = 0.088) failed to show any association with the disease. At locus 9p13.2 (rs3780135, PAX5), the risk allele frequency was significantly higher in B-ALL subjects than ancestral allele frequency (OR = 2.17, CI [1.37–3.43], P = 0.000). Genotype frequency analysis of rs3780135 polymorphism exhibited the protective effect (OR = 0.55, CI [0.72–1.83], P = 0.029). At locus 13q12.2 (rs12430881, FLT3), the minor allele frequency G (OR = 1.15, CI [1.37–3.43], P = 0.043) and genotype frequency (OR = 2.52, P = 0.006) reached significance as showed p < 0.05.
CONCLUSION
In the present study, a strong risk of B-cell acute lymphoblastic leukemia was associated with rs35958982 and rs12430881 polymorphisms. However, rs3780135 polymorphism showed the protective effect. Additionally, other demographic factors like family history, smoking and consanguinity were also found to be important in risk assessment. We anticipate that the information from genetic variations in this study can aid in therapeutic approach in the future.
Keywords: Acute lymphoblastic leukemia, Single nucleotide polymorphism, B-cell ALL, PAX5 gene, FLT3 variant
Introduction
According to the Punjab cancer registry report, acute lymphoblastic leukemia (ALL) is a predominant malignancy among children and it makes up most prevalent cancer in Punjab, Pakistan. The worldwide incidence rate is 1–4.75 per 100,000 people. In Pakistan ALL contributes to 17.9% of all cancers. It is characterized by mutation in blast cells in hematopoietic stem cells, spleen, neurons, gonads, lymph nodes, and hepatic cells (Portell, Wenzell & Advani, 2013). Although, B-ALL is very common in children but it may also occur in the adult populace (Forero, Hernández & Rivas, 2013). Several demographic parameters like gender, age, family history and biological factors also play an important role in the prevalence of disease. Other factors like exposure to UV, radiations, lifestyle may also act as risk factors (Levine et al., 2016; Acharya et al., 2018). Mutation in certain genes involved in different processes like apoptosis, proliferation and differentiation of B-cells may also cause B-ALL. These genetic alterations largely affect the prediction and therapeutic approach used for medication and therapy of ALL (Tasian & Hunger, 2017).
FMS-like tyrosine kinase (FLT3) belongs to class III receptor tyrosine kinase (RTK) family. Structurally, FLT3 consists of an extracellular domain at the amino terminus. This domain comprises of immunoglobulin-like transmembrane region and intracellular juxta-membrane domain (JMD). At the carboxyl terminus, there are two kinase domains, separated by a kinase insert region (Gilliland & Griffin, 2002). FLT3 is expressed in normal human bone marrow especially in CD34+ hematopoietic stem, brain (Çakmak Görür et al., 2019) and gonads (Matthews et al., 1991; Small et al., 1994) and encodes 1,000 amino acid protein in humans. In the hematopoietic tissues, binding of FL with its receptor causes auto-phosphorylation of tyrosine residues present in the kinase domain and stimulates growth of progenitor cells in the marrow and blood (Marhäll et al., 2018). This results in downstream activation of signaling pathways that are involved in regulation of cell cycle or apoptosis, including (PI3K), caspase-9 and Ras/Raf pathways and causes multiplied proliferation of cells, reduced cell apoptosis, and inhibition of B-cell differentiation (Zhang & Broxmeyer, 2000).
In hematologic malignancy, 70% to 100% increased expression of FLT3 in acute myeloid leukemia (AML) and acute lymphoblastic leukemia (ALL) is reported previously (Brown et al., 2005; Griffith et al., 2016). Rosnet and colleagues reported that three out of five ALL subjects with increased expression of FLT3 in leukemia blasts (Rosnet et al., 1996). Another study showed that up regulation of FLT3 gene is a potential risk factor of leukemia (Cheng et al., 2018).
B-cell-specific activator protein (PAX5) encodes transcription factors that are the member of a paired box domain. PAX5 plays imperative role in the commitment of B-cell lineage from blast cells as it controls the differentiation of a pro-B cell to pre-B cells (Fuxa & Skok, 2007; Lang et al., 2007). In pre-pro-B cells the immunoglobin gene rearrangement starts and matures into pro-B cells. Expression of PAX5 gene initiates from pro-B stage and terminates at pre-B stage. In late B-lymphoposis, PAX5 maintains the function of mature B-cells (Shahjahani et al., 2015).
In B-cell malignancies, PAX5 act as an oncogene. Down-regulation of PAX5 halts B-cells and reverts B-cell precursors (BCPs) to progenitors (pro B-cell stage) (Schebesta et al., 2007; Carotta & Nutt, 2008). Conversely, uncontrolled proliferation of the B-cells leads to the abnormal expression of PAX5 in precursor cells and inhibit T-cell proliferation (Souabni, Jochum & Busslinger, 2007). It is reported that in childhood ALL, translocations and mutation in PAX5 are more prevalent (Bousquet et al., 2007; Nebral et al., 2009; Santoro et al., 2009; Iacobucci & Mullighan, 2017). Alternative splicing of PAX5 in exon 7 to exon 9 results into five isoforms. These isoforms are more expressed in primary B-cell lymphoma tissues and cancerous cell lines (Zwollo et al., 1997; Arseneau et al., 2009).
Previous studies showed that the presence of single nucleotide polymorphisms (SNPs) in genome maybe risk causing or protective for the disease and it may also alters the pharmacokinetic and pharmacodynamics properties of drugs (Kumanayake, 2013; Pui, 2015; Tasian & Hunger, 2017). We selected two non-synonomous SNPs including 13q12.2 (rs35958982, FLT3), 557 (Val > Ile) at position Chr13:28034336 (GRCh38.p12) and 9p13.2 (rs3780135, PAX5), 293 (Thr > Ile) at position Chr9:36840626 (GRCh38.p12). A synonomous SNP 13q12.2 (rs12430881, FLT3), (A > G) at position Chr13:28020665 (GRCh38.p12) was also selected. The change in amino acid sequence due to non-synonomous SNP alters the protein structure implicating its expression and function. Current study is designed to evaluate the role of FLT3 and PAX5 genes in B-cell lymphoblastic leukemia. For this purpose, a case control analysis was conducted to evaluate the polymorphic association of rs35958982, rs3780135 and rs12430881 with B-cell acute lymphoblastic leukemia (B-ALL) incidence.
Materials & Methods
Study subjects
The present study was conducted at the University of Punjab, Pakistan and granted ethical approval to carry out the study within its facilities (Ethical Application Ref: sbs/222/18). Blood samples were collected during the period of January 2017 to February 2017 from Children’s Hospital, Lahore, Pakistan. Study population comprised of 155 cases and 155 controls younger than 15 years of age. The diagnostic criteria for B-ALL cases include B-cell positive markers (CD19, CD10, CD22, and CD20) confirmed by flow cytometry analysis. Cases with relapsed and newly diagnosed B-ALL were also included. All 310 subjects recruited were consented to participate in this study after filling the questionnaire. The subjects with any other type of leukemia, blood infectious disease, and B-ALL subjects older than 15 years of age were excluded from the study. Family history with cancer, parental consanguinity (first and second degree relatives) and smoking status (>100 cigarettes in lifetime) were gathered by questionnaire interviewed.
Genotyping
Venous blood samples of cases and controls were collected in EDTA vials. DNA extraction was done using Sam brook 2001 organic protocol. The genes and SNPs associated with B-ALL were screened using DisGeNET platform (Queralt-Rosinach et al., 2016) and were verified by dbSNP database (Sherry et al., 2001). Presence of the selected SNPs in Pakistani population was confirmed by Ensembl genome browser (Frankish et al., 2017). In order to identify the SNPs, tetra arm primers were designed using Primer1 software (Ye et al., 2001) as shown in Table 1. Tetra arms PCR was done using advanced primus 96 (PeqLab) thermal cycler (Table 2). PCR products were further analyzed by gel electrophoresis (Figs. 1–3).
Table 1. Tetra-ARMS primers.
| Primer | Sequence (5′–3′) | Tm (°C) |
|---|---|---|
| rs35958982 | TGTGACAAATTAGCAGGGTTAACAC | 57.3 |
| CACAGAAGAGATCACAGAAGGAGTCT | 60.7 | |
| GAAACTCCCATTTGAGATCATATTCA | 56.0 | |
| AGACAGAGACAAGCAGACATTCG | 58.4 | |
| rs3780135 | CTCTTCCAGGCTCCCCCGAC | 59.2 |
| GGGCGGCAGCGCTATAAGAA | 59.5 | |
| ACCCCAGCTCTAGATGGCGAAG | 56.6 | |
| ATAGGTGCCATCAGTGTTTGGTGC | 58.4 | |
| rs12430881 | GTTTGTCTCCTCTTCATTGGCA | 56.0 |
| GCCTCAGTGTCATCTTCGAATT | 56.3 | |
| CCTTTTATCTTCACATCAGGCCT | 56.6 | |
| CTTAGTAGAGATGGGGTTTTGCC | 58.4 |
Table 2. PCR program for SNPs.
| PCR steps | Temperature (°C) | Duration of steps | No. of cycles |
|---|---|---|---|
| Initial duration | 92 | 5 min | 30–35 |
| Denaturation | 94 | 30 sec | |
| Annealing rs35958982 | 58.4 | 1 min | |
| rs3780135 | 56.6 | ||
| rs12430881 | 58.8 | ||
| Extension | 72 | 1 min | |
| Final extension | 72 | 5 min |
Figure 1. SNP rs35958982.
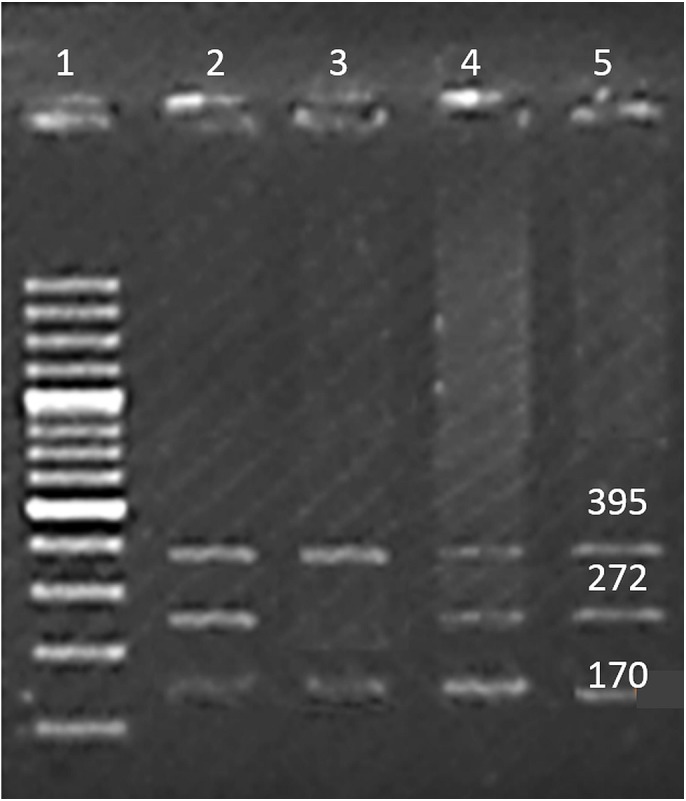
Well 1 indicates the DNA ladder (100 bp), an amplicon (395 bp) is outer band. Amplicon 272 bp: allele ‘A’ and amplicon of 170 bp allele ‘G’.
Figure 3. SNP rs12430881.
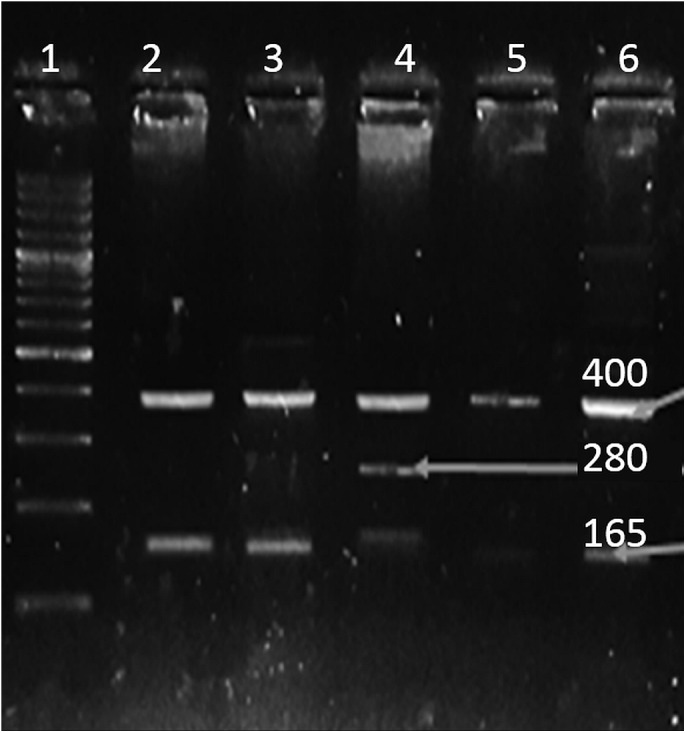
Well 1 indicates the DNA ladder (100 bp), an amplicon (400 bp) is outer band. Amplicon 165 bp: allele G and amplicon of 280 bp: allele A.
Figure 2. SNP rs3780135.
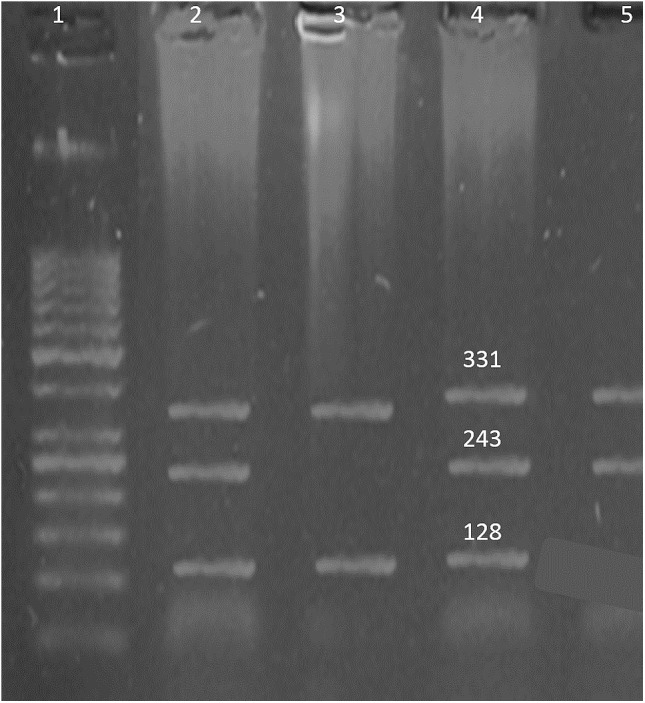
Well 1 indicates the DNA ladder (50 bp), an amplicon (331 bp) is outer band. Amplicon 243 bp: allele ‘A’ and amplicon of 128 bp: allele ‘G’.
Statistical analysis
Statistical studies were performed using IBM SPSS 23. Chi-square test was conducted to compare categorical data. Allele and genotype association between SNPs and B-ALL were calculated by computing odds ratio (OR) and 95% confidence interval (CI). The Bonferroni corrections were applied for all multiple tests. A logistic regression model was used to adjust different B-ALL risk factors. The probability level accepted for significance was P < 0.05.
Results
Family history of cancer and parental consanguinity showed significant association with B-ALL while, there was no association with the smoker parents. Subjects with a family history of any type of cancer showed a high risk of having B-ALL (OR = 15.42, P = 0.000). Previous studies showed smoking as a risk factor for cancer but our cohort displayed a contradictory results as no significant association was found in B-ALL subjects (OR = 0.85, P = 0.580). In the present study, more B-ALL subjects were product of parental consanguinity and showed highly significant association with the risk of B-ALL (OR = 1.87, P = 0.050) (Table 3).
Table 3. Association of demographic factors with risk of B-ALL.
| Parameters | Patients (%) | Control (%) | Odd ratio | Chi-square | P value |
|---|---|---|---|---|---|
| Age (mean) | 7.30 | 11.70 | |||
| A positive family history | 16.77 | 1.29 | 15.42 | 14.59 | 0.000* |
| A negative family history | 83.22 | 98.70 | |||
| Smoking by parent | 38.06 | 41.94 | 0.85 | 0.31 | 0.580 |
| No smoking parent | 61.93 | 58.06 | |||
| Parental cousin marriage | 33.55 | 21.29 | 1.87 | 3.78 | 0.050* |
| No cousin marriage | 66.45 | 78.70 | |||
| Females | 56 | 72 | 0.65 | 3.41 | 0.070 |
| Males | 99 | 83 |
Notes.
Significant values are shown in (*).
Our data showed that none of the subjects and their parents was exposed to radiations. Furthermore, 18 patients had liver hepatomegaly sized 11.7 ± 3.3 mm, nine cases had nephropathy of right kidney 9.47 ± 2.9 mm and left kidney 10.01 ± 2.3 mm. Symptoms like night sweating, dizziness, abdominal pain, vomiting, bruises, pallor, enlarged lymph nodes, cough with blood, loose stools, jaundice, pedal edema, pain, dehydration, hepatosplenomegaly, atypical blast cells mild abdominal ascites, low leukocytes and thrombocytopenia were also recorded.
In our cohort, rs35958982 encoding isloleucine form of codon frequency in subjects was 13.4% and 5.6% in controls. Moreover, statistical analysis showed positive association of allele frequency (OR = 2.30, CI [1.20–4.50], P = 0.005) and no association of genotype frequency (OR = 3.67, CI [0.75–18.10], P = 0.088) with the disease. Another polymorphism rs3780135, a minor allele frequency in subjects was 47.1% and 32.58% in controls showing positive association with B-ALL (OR = 2.17, CI [1.37–3.43], P = 0.000). The genotype frequency showed protective effect with B-ALL (OR = 0.55, CI [0.72–1.83], P = 0.029). The SNP rs12430881 allele frequency (OR = 1.15, CI [1.37–3.43], P = 0.043) and genotype frequency (OR = 2.52, CI [1.28–4.95], P = 0.006) showed strong association with the disease as shown in Table 4. After applying Bonferroni correction, SNPs rs35958982, rs3780135 and rs12430881 remained statistically significant and showed P-value 0.030, 0.010 and 0.002 respectively.
Table 4. Allele and genotype frequency.
Adjusted ORs were obtained from logistic regression model with adjustment for family history, smoking and consanguinity.
| Gene/SNP | Allele/ Genotype | Controls (%) | Cases (%) | Crude OR (95% CI) | Adjusted OR (95% CI) | χ2 | aP-value | bP-value |
|---|---|---|---|---|---|---|---|---|
| rs35958982 | Allele | |||||||
| A | 5.60 | 13.40 | 2.30 (1.20-4.50) | – | 7.79 | 0.005* | 0.002* | |
| G | 94.40 | 86.60 | – | |||||
| Genotype | ||||||||
| GG | 90.70 | 79.60 | 3.67 (0.75-18.10) | 1.00 | 2.90 | 0.088 | 0.030* | |
| GA | 7.60 | 13.80 | 1.13 (0.41–3.08) | |||||
| AA | 1.85 | 6.50 | 1.30 (0.37–5.08) | |||||
| rs3780135 | Allele | |||||||
| A | 32.58 | 47.10 | 2.17 (1.37-3.43) | – | 13.63 | 0.000* | 0.000* | |
| G | 67.42 | 52.90 | – | |||||
| Genotype | ||||||||
| GG | 52.26 | 33.55 | 0.55 (0.39-0.95) | 1.00 | 4.72 | 0.029* | 0.010* | |
| GA | 30.32 | 38.70 | 1.19 (0.52–2.73) | |||||
| AA | 17.42 | 27.74 | 0.97 (0.26–1.42) | |||||
| rs12430881 | Allele | |||||||
| G | 22 | 29 | 1.15 (0.72-1.83) | – | 4.11 | 0.043* | 0.014* | |
| A | 78 | 71 | – | |||||
| Genotype | ||||||||
| AA | 65.16 | 61.93 | 2.52 (1.28 -4.95) | 1.00 | 7.51 | 0.006* | 0.002* | |
| GA | 25.80 | 18.06 | 1.09 (0.48–2.69) | |||||
| GG | 9.03 | 20 | 1.03 (0.50–2.68) |
Notes.
Critical P value.
Bonferroni corrected P value.
Significant values are shown in (*).
Multivariate regression analysis was performed after adjusting the baseline for conventional B-ALL risk factors such as family history, smoking and parental consanguinity. As shown in Table 4, the multivariate analysis indicated that outcome of heterozygous genotype GA in SNPs rs35958982, rs3780135 and rs12430881 had significant association with B-ALL and showed odds ratio (OR = 1.13, CI [0.41–3.08]), (OR = 1.19, CI [0.52–2.73]) and (OR = 1.09, CI [0.48–2.69]) respectively. Additionally, risk genotypes in SNPs rs35958982 (AA) and rs12430881 (GG) showed positive association with disease having an odds ratio (OD = 1.30, CI [0.37–5.08]) and (OD = 1.03, CI [0.50–2.68]), respectively. However, the risk genotype (AA) of SNP rs3780135 displayed no association with B-ALL after adjusting for environmental factors. Stratification analysis of environmental factors showed smoking as major risk factor in both heterozygous and risk genotype of SNP rs3780135 and rs12430881 whereas, parental consanguinity act as risk factor only in heterozygous genotype of SNP rs3780135 as shown in Table 5.
Table 5. Stratification analysis for association between genotypes and risk of B-ALL.
ORs were obtained from logistic regression model with adjustment for family history, smoking and consanguinity.
| OR (95% CI) | |||
|---|---|---|---|
| rs35958982 | GG | GA | AA |
| Family history status | |||
| Yes | 1 | 0.92(0.18–4.56) | 0.50(0.06–4.15) |
| No | |||
| Smoking status | |||
| Yes | 1 | 0.30(0.08–1.09) | 0.46(0.10–2.12) |
| No | |||
| Consanguinity status | |||
| Yes | 1 | 0.86(0.24–3.10) | 0.94(0.20–4.37) |
| No | |||
| rs3780135 | GG | GA | AA |
| Family history status | |||
| Yes | 1 | 0.60(1.70–0.21) | 0.44(0.15–1.30) |
| No | |||
| Smoking status | |||
| Yes | 1 | 1.08(0.46–2.50) | 1.3(0.59–2.90) |
| No | |||
| Consanguinity status | |||
| Yes | 1 | 0.62(0.26–1.44) | 0.60(0.26–1.35) |
| No | |||
| rs12430881 | AA | AG | GG |
| Family history status | |||
| Yes | 1 | 0.58(0.16–2.15) | 1.80(0.65–4.98) |
| No | |||
| Smoking status | |||
| Yes | 1 | 1.05(0.45–2.42) | 1.08(0.45–2.60) |
| No | |||
| Consanguinity status | |||
| Yes | 1 | 1.15(0.50–2.65) | 0.49(0.18–1.35) |
| No | |||
Discussion
According to previous studies, association of first and second degree family history of cancer signifies genetic and environmental risk factor for causing acute lymphoblastic leukemia. Our study also showed positive association of family history with B-ALL (OR = 15.42, P = 0.000). Earlier, parental smoking has also been associated with the prevalence of ALL but our study showed contrary results (OR = 0.85, P = 0.580) (Belson, Kingsley & Holmes, 2007). Parental consanguinity is still practiced in Pakistan, which results in minor allele pool and contributes to the occurrence of disease. Our results are in accordance with (Steinberg & Steinfeld, 1960; Urtishak et al., 2016) which states that familial occurrence of leukemia exists (OR = 1.87, P = 0.050). Some studies found a correlation between parental exposure to radiation before conception, that may be due to their working environment (Shu et al., 2002). In our analysis, neither patients nor parents were ever exposed to radiations. Hepatomegaly and nephropathy are often seen in B-ALL subjects having chemotherapy. Malfunctioned leucocytes in the liver and kidney leads to enlargement of these organs (Rasool et al., 2015). Another study suggests that hepatomegaly and nephropathy may be the consequence of chemotherapeutic toxicity (Giamanco et al., 2016).
It is well established fact that cancer risk is influenced by numerous genetic variants having any risk or protective effect. The degree of penetrance of a certain genotype in the population and environmental factors is a major cause of cancer (Fletcher & Houlston, 2010). The information given by allelic and genotypic data of single nucleotide polymorphism in a population propose the possible genetic markers for cancer risk and predict possible targeted therapies (Griffith et al., 2016; Wu & Li, 2018).
In this study, SNP rs35958982 is a germline polymorphism present in transmembrane region of FLT3 gene. It is a non-synonymous variant which leads to the change in structure of the protein. High throughput DNA sequence analysis has been done to check the frequency of rs35958982 with leukemiogenesis in drivers and passengers which showed no association with AML (Fröhling et al., 2007). Present study in contrast displayed the association of SNP rs35958982 with the disease (OR = 2.30, CI [1.20–4.50], P = 0.005). Detailed analysis of genotype frequency in the population showed no association with B-ALL (OR = 3.67, CI [0.75–18.10], P = 0.088). This might be due to the fact that SNP rs35958982 is rare in acute lymphoblastic leukemia with low penetrance. Current study also depicts that individuals with risk allele A at locus 13q12.2 (rs12430881, FLT3) (OR = 1.15, CI [1.37–3.43], P = 0.0426) and genotype GG were more prone to B-ALL (OR = 2.52, CI [1.28–4.95], P = 0.006). It has been found that disruption of FLT3 gene due to the presence of mutation or SNP leads to deficiency of B-lymphoid progenitors suggesting its critical role in survival and proliferation of blast cells (Zriwil et al., 2018).
Bodian et al. (2014) studied allele frequency of paired box domain (PAX5) polymorphism rs3780135 in different populations, i.e., African 34%, African European 49%, Central Asian 85%, East Asian 94%, European 95% and Hispanic 88%. Pakistan lies in South East Asia having frequency of rs3780135 (47.1%) which is lower than previously reported in East Asian population. Firtina et al. (2012) found polymorphism rs3780135 in B-ALL subjects with increased mRNA expression of PAX5 suggesting the possible role of SNP with increased proliferation of blast cells. In Pakistani population, minor allele frequency was significantly identified in B-ALL subjects (OR = 2.17, CI [1.37–3.43], P = 0.000). Heterozygous genotype GA (38.7%) was more frequently identified in our cohort than homozygous risk genotype AA (27.74%) which manifested significant difference in frequency (CI [0.72–1.83], P = 0.029) and also showed protective effect (OR = 0.55). PAX5 is involved in repression of T-cells, activation of B-cell proliferation from blast cell therefore, presence of any variant in this gene affects its pathway which may leads to increased expression of PAX5 and results into B-ALL (Firtina et al., 2012).
Conclusions
The findings of the present study significantly demonstrate that SNPs rs35958982 and rs12430881 correlate with the increase risk of B-ALL, however, SNP rs3780135 has a protective effect. The environmental risk factors of B-ALL, including family history, parental consanguinity and smoking, are found to have an imperative role in progression of disease. Although the data is balanced, but not robust, the small cohort of subjects limits the conclusion of article. To eliminate this limitation and validate the results of present study, larger prospective studies need to be conducted in the same ethnic group. Furthermore, various other demographic and environmental factors should also be considered and appraised for their association with B-ALL.
Supplemental Information
Raw data showing genotype of 3 SNPs and environmental factors associated with the disease. Sheet 1 displays patients data and sheet 2 display controls data.
Acknowledgments
Authors would like to acknowledge The Children’s Hospital and Institute of Child Health, Lahore, Pakistan for their assistance and support during blood sample collection.
Funding Statement
The authors received no funding for this work.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Ammara Khalid conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Sara Aslam performed the experiments, prepared figures and/or tables.
Mehboob Ahmed analyzed the data, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper.
Shahida Hasnain contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper.
Aimen Aslam analyzed the data, prepared figures and/or tables.
Ethics
The following information was supplied relating to ethical approvals (i.e., approving body and any reference numbers):
The University of the Punjab, Lahore, Pakistan granted Ethical approval to carry out the study within its facilities (Ethical Application Ref: sbs/222/18).
Data Availability
The following information was supplied regarding data availability:
Data is available as Supplemental File.
References
- Acharya et al. (2018).Acharya UH, Halpern AB, Wu Q, Voutsinas JM, Walter RB, Yun S, Kanaan M, Estey EH. Impact of region of diagnosis, ethnicity, age, and gender on survival in acute myeloid leukemia (AML) Journal of Drug Assessment. 2018;7:51–53. doi: 10.1080/21556660.2018.1492925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arseneau et al. (2009).Arseneau JR, Laflamme M, Lewis SM, Maïcas E, Ouellette RJ. Multiple isoforms of PAX5 are expressed in both lymphomas and normal B-cells. British Journal of Haematology. 2009;147:328–338. doi: 10.1111/j.1365-2141.2009.07859.x. [DOI] [PubMed] [Google Scholar]
- Belson, Kingsley & Holmes (2007).Belson M, Kingsley B, Holmes A. Risk factors for acute leukemia in children: a review. Environmental Health Perspectives. 2007;115:138–145. doi: 10.1289/ehp.9023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bodian et al. (2014).Bodian DL, McCutcheon JN, Kothiyal P, Huddleston KC, Iyer RK, Vockley JG, Niederhuber JE. Germline variation in cancer-susceptibility genes in a healthy, ancestrally diverse cohort: implications for individual genome sequencing. PLOS ONE. 2014;9:e94554. doi: 10.1371/journal.pone.0094554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bousquet et al. (2007).Bousquet M, Broccardo C, Quelen C, Meggetto F, Kuhlein E, Delsol G, Dastugue N, Brousset P. A novel PAX5-ELN fusion protein identified in B-cell acute lymphoblastic leukemia acts as a dominant negative on wild-type PAX5. Blood. 2007;109:3417–3423. doi: 10.1182/blood-2006-05-025221. [DOI] [PubMed] [Google Scholar]
- Brown et al. (2005).Brown P, Levis M, Shurtleff S, Campana D, Downing J, Small D. FLT3 inhibition selectively kills childhood acute lymphoblastic leukemia cells with high levels of FLT3 expression. Blood. 2005;105:812–820. doi: 10.1182/blood-2004-06-2498. [DOI] [PubMed] [Google Scholar]
- Çakmak Görür et al. (2019).Çakmak Görür N, Radke J, Rhein S, Schumann E, Willimsky G, Heppner FL, Blankenstein T, Pezzutto A. Intracellular expression of FLT3 in Purkinje cells: implications for adoptive T-cell therapies. Leukemia. 2019;33:1039–1043. doi: 10.1038/s41375-018-0330-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carotta & Nutt (2008).Carotta S, Nutt SL. Losing B cell identity. Bioessays. 2008;30:203–207. doi: 10.1002/bies.20725. [DOI] [PubMed] [Google Scholar]
- Cheng et al. (2018).Cheng J, Qu L, Wang J, Cheng L, Wang Y. High expression of FLT3 is a risk factor in leukemia. Molecular Medicine Reports. 2018;17:2885–2892. doi: 10.3892/mmr.2017.8232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Firtina et al. (2012).Firtina S, Sayitoglu M, Hatirnaz O, Erbilgin Y, Oztunc C, Cinar S, Yildiz I, Celkan T, Anak S, Unuvar A. Evaluation of PAX5 gene in the early stages of leukemic B cells in the childhood B cell acute lymphoblastic leukemia. Leukemia Research. 2012;36:87–92. doi: 10.1016/j.leukres.2011.07.017. [DOI] [PubMed] [Google Scholar]
- Fletcher & Houlston (2010).Fletcher O, Houlston RS. Architecture of inherited susceptibility to common cancer. Nature Reviews Cancer. 2010;10:353–361. doi: 10.1038/nrc2840. [DOI] [PubMed] [Google Scholar]
- Forero, Hernández & Rivas (2013).Forero RM, Hernández M, Rivas JMH. Chapter 01: genetics of acute lymphoblastic leukemia. In: Guenova M, Balatzenko G, editors. Leukemia. InTech; Rijeka: 2013. [Google Scholar]
- Fröhling et al. (2007).Fröhling S, Scholl C, Levine RL, Loriaux M, Boggon TJ, Bernard OA, Berger R, Döhner H, Döhner K, Ebert BL. Identification of driver and passenger mutations of FLT3 by high-throughput DNA sequence analysis and functional assessment of candidate alleles. Cancer Cell. 2007;12:501–513. doi: 10.1016/j.ccr.2007.11.005. [DOI] [PubMed] [Google Scholar]
- Fuxa & Skok (2007).Fuxa M, Skok JA. Transcriptional regulation in early B cell development. Current Opinion in Immunology. 2007;19:129–136. doi: 10.1016/j.coi.2007.02.002. [DOI] [PubMed] [Google Scholar]
- Giamanco et al. (2016).Giamanco NM, Cunningham BS, Klein LS, Parekh DS, Warwick AB, Lieuw K. Allopurinol Use During Maintenance Therapy for Acute Lymphoblastic Leukemia Avoids Mercaptopurine-related Hepatotoxicity. Journal of Pediatric Hematology/Oncology. 2016;38:147–151. doi: 10.1097/MPH.0000000000000499. [DOI] [PubMed] [Google Scholar]
- Gilliland & Griffin (2002).Gilliland DG, Griffin JD. The roles of FLT3 in hematopoiesis and leukemia. Blood. 2002;100:1532–1542. doi: 10.1182/blood-2002-02-0492. [DOI] [PubMed] [Google Scholar]
- Griffith et al. (2016).Griffith M, Griffith OL, Krysiak K, Skidmore ZL, Christopher MJ, Klco JM, Ramu A, Lamprecht TL, Wagner AH, Campbell KM. Comprehensive genomic analysis reveals FLT3 activation and a therapeutic strategy for a patient with relapsed adult B-lymphoblastic leukemia. Experimental Hematology. 2016;44:603–613. doi: 10.1016/j.exphem.2016.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iacobucci & Mullighan (2017).Iacobucci I, Mullighan CG. Genetic basis of acute lymphoblastic leukemia. Journal of Clinical Oncology. 2017;35:975–983. doi: 10.1200/JCO.2016.70.7836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumanayake (2013).Kumanayake P. Genome-wide SNP discovery in associating with human diseases phenotypes. Sri Lanka Journal of Bio-Medical Informatics. 2013;3:25–31. [Google Scholar]
- Lang et al. (2007).Lang D, Powell SK, Plummer RS, Young KP, Ruggeri BA. PAX genes: roles in development, pathophysiology, and cancer. Biochemical Pharmacology. 2007;73:1–14. doi: 10.1016/j.bcp.2006.06.024. [DOI] [PubMed] [Google Scholar]
- Levine et al. (2016).Levine PH, Ajmera K, O’Neill B, Venkatesh V, Garcia-Gonzalez P, Hoffman HJ. Demographic factors related to young age at diagnosis of chronic myeloid leukemia in India. Clinical Epidemiology and Global Health. 2016;4:188–192. doi: 10.1016/j.cegh.2016.06.001. [DOI] [Google Scholar]
- Marhäll et al. (2018).Marhäll A, Heidel F, Fischer T, Rönnstrand L. Internal tandem duplication mutations in the tyrosine kinase domain of FLT3 display a higher oncogenic potential than the activation loop D835Y mutation. Annals of Hematology. 2018;97:773–780. doi: 10.1007/s00277-018-3245-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews et al. (1991).Matthews W, Jordan CT, Wiegand GW, Pardoll D, Lemischka IR. A receptor tyrosine kinase specific to hematopoietic stem and progenitor cell-enriched populations. Cell. 1991;65:1143–1152. doi: 10.1016/0092-8674(91)90010-V. [DOI] [PubMed] [Google Scholar]
- Nebral et al. (2009).Nebral K, Denk D, Attarbaschi A, König M, Mann G, Haas OA, Strehl S. Incidence and diversity of PAX5 fusion genes in childhood acute lymphoblastic leukemia. Leukemia. 2009;23:134–143. doi: 10.1038/leu.2008.306. [DOI] [PubMed] [Google Scholar]
- Portell, Wenzell & Advani (2013).Portell CA, Wenzell CM, Advani AS. Clinical and pharmacologic aspects of blinatumomab in the treatment of B-cell acute lymphoblastic leukemia. Clinical Pharmacology: Advances and Applications. 2013;5:5–11. doi: 10.2147/CPAA.S42689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pui (2015).Pui C-H. Genomic and pharmacogenetic studies of childhood acute lymphoblastic leukemia. Frontiers of Medicine. 2015;9:1–9. doi: 10.1007/s11684-015-0381-3. [DOI] [PubMed] [Google Scholar]
- Queralt-Rosinach et al. (2016).Queralt-Rosinach N, Pinero J, Bravo À, Sanz F, Furlong LI. DisGeNET-RDF: harnessing the innovative power of the Semantic Web to explore the genetic basis of diseases. Bioinformatics. 2016;32:2236–2238. doi: 10.1093/bioinformatics/btw214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasool et al. (2015).Rasool M, Farooq S, Malik A, Shaukat A, Manan A, Asif M, Sani S, Qazi MH, Kamal MA, Iqbal Z. Assessment of circulating biochemical markers and antioxidative status in acute lymphoblastic leukemia (ALL) and acute myeloid leukemia (AML) patients. Saudi Journal of Biological Sciences. 2015;22:106–111. doi: 10.1016/j.sjbs.2014.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosnet et al. (1996).Rosnet O, Bühring H, Marchetto S, Rappold I, Lavagna C, Sainty D, Arnoulet C, Chabannon C, Kanz L, Hannum C. Human FLT3/FLK2 receptor tyrosine kinase is expressed at the surface of normal and malignant hematopoietic cells. Leukemia. 1996;10:238–248. [PubMed] [Google Scholar]
- Santoro et al. (2009).Santoro A, Bica MG, Dagnino L, Agueli C, Salemi D, Cannella S, Veltroni M, Cetica V, Giarin E, Fabbiano F. Altered mRNA expression of PAX5 is a common event in acute lymphoblastic leukaemia. British Journal of Haematology. 2009;146:686–689. doi: 10.1111/j.1365-2141.2009.07815.x. [DOI] [PubMed] [Google Scholar]
- Schebesta et al. (2007).Schebesta A, McManus S, Salvagiotto G, Delogu A, Busslinger GA, Busslinger M. Transcription factor Pax5 activates the chromatin of key genes involved in B cell signaling, adhesion, migration, and immune function. Immunity. 2007;27:49–63. doi: 10.1016/j.immuni.2007.05.019. [DOI] [PubMed] [Google Scholar]
- Shahjahani et al. (2015).Shahjahani M, Norozi F, Ahmadzadeh A, Shahrabi S, Tavakoli F, Asnafi AA, Saki N. The role of Pax5 in leukemia: diagnosis and prognosis significance. Medical Oncology. 2015;32:360. doi: 10.1007/s12032-014-0360-6. [DOI] [PubMed] [Google Scholar]
- Sherry et al. (2001).Sherry ST, Ward M-H, Kholodov M, Baker J, Phan L, Smigielski EM, Sirotkin K. dbSNP: the NCBI database of genetic variation. Nucleic Acids Research. 2001;29:308–311. doi: 10.1093/nar/29.1.308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shu et al. (2002).Shu XO, Potter JD, Linet MS, Severson RK, Han D, Kersey JH, Neglia JP, Trigg ME, Robison LL. Diagnostic X-rays and ultrasound exposure and risk of childhood acute lymphoblastic leukemia by immunophenotype. Cancer Epidemiology and Prevention Biomarkers. 2002;11:177–185. [PubMed] [Google Scholar]
- Small et al. (1994).Small D, Levenstein M, Kim E, Carow C, Amin S, Rockwell P, Witte L, Burrow C, Ratajczak MZ, Gewirtz AM. STK-1, the human homolog of Flk-2/Flt-3, is selectively expressed in CD34+ human bone marrow cells and is involved in the proliferation of early progenitor/stem cells. Proceedings of the National Academy of Sciences of the United States of America. 1994;91:459–463. doi: 10.1073/pnas.91.2.459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Souabni, Jochum & Busslinger (2007).Souabni A, Jochum W, Busslinger M. Oncogenic role of Pax5 in the T-lymphoid lineage upon ectopic expression from the immunoglobulin heavy-chain locus. Blood. 2007;109:281–289. doi: 10.1182/blood-2006-03-009670. [DOI] [PubMed] [Google Scholar]
- Steinberg & Steinfeld (1960).Steinberg AG, Steinfeld JL. The genetics of acute leukemia in children. Cancer. 1960;13:985–999. doi: 10.1002/1097-0142(196009/10)13:5<985::AID-CNCR2820130520>3.0.CO;2-J. [DOI] [Google Scholar]
- Tasian & Hunger (2017).Tasian SK, Hunger S. Genomic characterization of paediatric acute lymphoblastic leukaemia: an opportunity for precision medicine therapeutics. British Journal of Haematology. 2017;176:867–882. doi: 10.1111/bjh.14474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urtishak et al. (2016).Urtishak KA, Robinson BW, Rappaport EF, Sarezky MD, Biegel JA, Nichols KE, Wilmoth DM, Wang LS, Stern JW, Felix CA. Unique familial MLL (KMT2A)-rearranged precursor B-Cell infant acute lymphoblastic leukemia in non-twin siblings. Pediatric Blood & Cancer. 2016;63:1175–1180. doi: 10.1002/pbc.25957. [DOI] [PubMed] [Google Scholar]
- Wu & Li (2018).Wu C, Li W. Genomics and pharmacogenomics of pediatric acute lymphoblastic leukemia. Critical Reviews in Oncology/Hematology. 2018;126:100–111. doi: 10.1016/j.critrevonc.2018.04.002. [DOI] [PubMed] [Google Scholar]
- Ye et al. (2001).Ye S, Dhillon S, Ke X, Collins AR, Day IN. An efficient procedure for genotyping single nucleotide polymorphisms. Nucleic Acids Research. 2001;29:e88–e88. doi: 10.1093/nar/29.17.e88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zerbino et al. (2017).Zerbino DR, Achuthan P, Akanni W, Amode MR, Barrell D, Bhai J, Billis K, Cummins C, Gall A, Girón CG, Gil L, Gordon L, Haggerty L, Haskell E, Hourlier T, Izuogu OG, Janacek SH, Juettemann T, To JK, Laird MR, Lavidas I, Liu Z, Loveland JE, Maurel T, McLaren W, Moore B, Mudge J, Murphy DN, Newman V, Nuhn M, Ogeh D, Ong CK, Parker A, Patricio M, Riat HS, Schuilenburg H, Sheppard D, Sparrow H, Taylor K, Thormann A, Vullo A, Walts B, Zadissa A, Frankish A, Hunt SE, Kostadima M, Langridge N, Martin FJ, Muffato M, Perry E, Ruffier M, Staines DM, Trevanion SJ, Aken BL, Cunningham F, Yates A, Flicek P. Ensembl 2018. Nucleic Acids Research. 2017;46:D754–D761. doi: 10.1093/nar/gkx1098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang & Broxmeyer (2000).Zhang S, Broxmeyer HE. Flt3 ligand induces tyrosine phosphorylation of gab1 and gab2 and their association with shp-2, grb2, and PI3 kinase. Biochemical and Biophysical Research Communications. 2000;277:195–199. doi: 10.1006/bbrc.2000.3662. [DOI] [PubMed] [Google Scholar]
- Zriwil et al. (2018).Zriwil A, Böiers C, Kristiansen TA, Wittmann L, Yuan J, Nerlov C, Sitnicka E, Jacobsen SE. Direct role of FLT 3 in regulation of early lymphoid progenitors. British Journal of Haematology. 2018;183:588–600. doi: 10.1111/bjh.15578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zwollo et al. (1997).Zwollo P, Arrieta H, Ede K, Molinder K, Desiderio S, Pollock R. The Pax-5 gene is alternatively spliced during B-cell development. Journal of Biological Chemistry. 1997;272:10160–10168. doi: 10.1074/jbc.272.15.10160. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Raw data showing genotype of 3 SNPs and environmental factors associated with the disease. Sheet 1 displays patients data and sheet 2 display controls data.
Data Availability Statement
The following information was supplied regarding data availability:
Data is available as Supplemental File.