

ABSTRACT
Multiple strategies have been developed to facilitate the efficient production of bispecific IgG (BsIgG) in single host cells. For example, we previously demonstrated near quantitative (≥90%) formation of BsIgG of different species and isotypes by combining ‘knob-into-hole’ mutations for heavy chain heterodimerization with engineered antigen-binding fragments (Fabs) for preferential cognate heavy/light chain pairing. Surprisingly, in this study we found high yield (>65%) of BsIgG1 without Fab engineering to be a common occurrence, i.e., observed for 33 of the 99 different antibody pairs evaluated. Installing charge mutations at both CH1/CL interfaces was sufficient for near quantitative yield (>90%) of BsIgG1 for most (9 of 11) antibody pairs tested with this inherent cognate chain pairing preference. Mechanistically, we demonstrate that a strong cognate pairing preference in one Fab arm can be sufficient for high BsIgG1 yield. These observed chain pairing preferences are apparently driven by variable domain sequences and can result from a few specific residues in the complementarity-determining region (CDR) L3 and H3. Transfer of these CDR residues into other antibodies increased BsIgG1 yield in most cases. Mutational analysis revealed that the disulfide bond between heavy and light chains did not affect the yield of BsIgG1. This study provides some mechanistic understanding of factors contributing to antibody heavy/light chain pairing preference and subsequently contributes to the efficient production of BsIgG in single host cells.
KEYWORDS: Bispecific antibody, bispecific IgG, single-cell production
Introduction
Bispecific antibodies (BsAb) can target two epitopes on the same or different antigens. BsAb can support novel mechanisms of action that are not available to monospecific antibodies, and thereby offer a promising approach to next-generation antibody drugs.1,2 Novel mechanisms of action for bispecifics are exemplified by the two currently marketed BsAb, namely, blinatumomab (anti-CD19/CD3) and emicizumab (anti-factor IXa/factor X).2 Blinatumomab directs T cells to kill tumor cells expressing CD19 and is approved for the treatment of B cell precursor acute lymphoblastic leukemia.3 Emicizumab mimics the function of factor VIII in promoting the formation of a factor IXa/factor X complex and is approved for the treatment of hemophilia A.4,5 Over 100 different BsAb formats have been described, including at least 10 different formats that have reached clinical trials.1
BsIgG is a widely used tetrameric BsAb format commonly consisting of two different light chains (LC) and two different heavy chains (HC). Production of BsIgG in single host cells can be difficult due to low yield of the desired bispecific and the formidable purification challenge of removing closely related IgG contaminants. These challenges result from co-expression of the four component chains (two LC and two HC), which may assemble into nine different mispaired IgG species in addition to the correctly paired BsIgG.6 Undesired chain mispairing events associated with BsIgG production in single cells include formation of HC homodimers and non-cognate HC/LC pairs.1,7
Several different strategies have been developed to overcome or avoid chain mispairing to allow efficient production of BsIgG for drug development.1,7 For example, efficient assembly of HC heterodimers with minimal homodimers was first achieved in the 1990s using ‘knob-into-hole’ (KIH) mutations.8–10 Subsequently, several other methods for efficient HC heterodimerization were developed.1 Alternatively, the HC pairing problem has been entirely avoided by using a common HC, as demonstrated with κλ-bodies.11
LC mispairing was initially circumvented using a common LC,9 and more recently by expression of engineered half-IgG in separate host cells followed by in vitro assembly into IgG.12–14 Efficient assembly of cognate HC/LC pairs has been accomplished by domain swapping with the CrossMab format.15 Subsequently, orthogonal antigen-binding fragment (Fab) interfaces were developed16 and further improved17 to favor cognate over non-cognate HC/LC pairing. Additional successful strategies developed for engineering Fab domain interfaces include replacement of a native inter-chain disulfide bond with an engineered one18,19 and installation of charge pairs either alone20,21 or in combination with KIH mutations.21
Previously we transiently expressed BsIgG in single mammalian cells (Expi293F) by co-transfection of DNAs encoding the constituent HC and LC.21,22 The resultant IgG species was purified from the cell culture supernatant by protein A chromatography. The BsIgG yield, defined here as the percentage of correctly assembled BsIgG in the purified pool of different IgG species, was estimated by high resolution and accurate mass liquid chromatography-mass spectrometry (LCMS) using an extended mass range (EMR) Orbitrap mass spectrometer.21,22 The HC in the BsIgG included KIH mutations for preferential HC heterodimerization, and the Fab was engineered to favor cognate over non-cognate HC/LC pairing.21 Control BsIgG were constructed with KIH HC mutations but without Fab engineering. For some control BsIgG such as anti-HER2/CD3 and anti-VEGFA/ANG2, the yield of BsIgG was 22–24%,21 consistent with random pairing of HC and LC as previously observed by others.23 In contrast, high BsIgG yield, arbitrarily defined here as >65%, was observed for other control bispecifics such as anti-EGFR/MET and anti-IL-4/IL-13 BsIgG, consistent with a cognate HC/LC pairing preference.21 Examples of elevated BsIgG yield without Fab mutations have also been observed by others.16,24
Prior to this study, the prevalence of cognate HC/LC pairing preferences and the molecular basis for this phenomenon were only poorly understood. Here, high throughput production and high-resolution LCMS analysis21,22 were utilized to survey 99 different antibody pairs with KIH HC, but without Fab mutations, for the yield of BsIgG1. One-third of antibody pairs showed high (>65%) BsIgG1 yield, consistent with a strong inherent cognate HC/LC chain pairing preference. Installation of previously identified charge mutations at the two CH1/CL domain interfaces21 for such antibody pairs was used to enhance the production of BsIgG1. Next, we investigated whether a cognate chain pairing preference in one or both arms was needed for high yield of BsIgG1. Mutational analysis was used to identify specific residues in CDR L3 and H3 contributing to high BsIgG1 yield. The CDR L3 and H3 and specific residues identified were then inserted into other available antibodies that show random HC/LC chain pairing to determine their effect upon BsIgG1 yield. Finally, mutational analysis was used to investigate the effect of the interchain disulfide bond upon the yield of BsIgG1.
Results
High yield of BsIgG1 without Fab mutations is a common phenomenon that depends on the constituent antibody pairs
Previously, we observed high yield of BsIgG (>65%) with KIH HC but without Fab arm mutations for two bispecifics, namely, anti-EGFR/MET and anti-IL-13/IL-4.21 Here we investigated the strength and frequency of occurrence of cognate HC/LC pairing preference. A large panel of antibody pairs (n = 99) was used to generate bispecific IgGs. For simplicity, all bispecifics in this study were constructed with human IgG1 HC constant domains. Six antibodies binding to either IL-13, IL-4, MET, EGFR, HER2 or CD321 were used to construct a matrix of all 15 possible BsIgG1. Next, these six antibodies were permuted with 14 additional antibodies that were mainly κ LC isotype with three λ LC isotype (anti-DR5, anti-α5β1, anti-RSPO2) (Table S1). All but three of the antibodies used in this study were humanized; the 3 exceptions were fully human antibodies (anti-CD33, anti-PDGF-C, anti-FluB). Antibody pairs were co-expressed at optimized chain ratios, and the yield of BsIgG1 was determined with an improved version of our previously described method.21,22 Briefly, following co-expression of antibody pairs and protein A chromatography, the purified IgG1 pools were further purified by size exclusion chromatography (SEC) to remove any small quantities of aggregates and half IgG1 present prior to quantitation by high-resolution LCMS. The yield of correctly assembled BsIgG1 in isobaric mixtures that also contained LC-scrambled IgG1 was estimated using a previously developed algebraic formula.22
The yield of BsIgG1 for the 99 unique antibody pairs varied over a very wide range: 22–95% (Table 1). Strikingly, non-random HC/LC pairing (>30% yield of BsIgG1) was observed for the majority (>80%) of antibody pairs, with high (>65%) and intermediate (30–65%) yield of BsIgG1 seen for 33 and 48 antibody pairs, respectively. Near quantitative (>90%) formation of BsIgG1 was measured for two antibody pairs (anti-MET/DR5 and anti-IL-13/DR5). High-resolution LCMS data for representative examples of low (anti-LGR5/IL-4, Figure 1(a,d)) intermediate (anti-SIRPα/IL-4, Figure 1(b,e)) and high (anti-MET/DR5, Figure 1(c,f)) yield of BsIgG1 are shown. The BsIgG1 yield for each antibody studied varied over a wide range depending upon its partner antibody. For example, the BsIgG1 yield for the anti-MET antibody varied from as little as ~21% when paired with anti-IL-33 to as much as ~95% when paired with anti-DR5 (Table 1). To investigate any influence of ‘knob’ and ‘hole’ mutations on the cognate HC/LC pairing preference, BsIgG1 were produced with the HC containing the ‘knob’ mutation in mAb1 and ‘hole’ mutations in mAb2 or vice versa (Table 1). The yield of BsIgG1 was minimally influenced by which HC contained the ‘knob’ and ‘hole’ mutations in all cases (n = 15) tested (Table 1). The recovery of IgG species from 30 mL cultures by protein A chromatography varied over ~5-fold (1.5 to 8.0 mg, Figure S1).
Table 1.
Survey of antibody pairs to investigate BsIgG1 yields without Fab mutations but with KIH HC mutations for HC heterodimerization. Different combinations of IgG1 HC and LC pairs were co-transfected into Expi293F cells and the yield of BsIgG1 determined by high-resolution LCMS following protein A chromatography and SEC. The yield of correctly assembled BsIgG1 in isobaric mixtures that also contained LC-scrambled IgG1 (see Figure 1 for representative examples) was estimated using a previously developed algebraic formula.22 Data shown are the yield of BsIgG1 from optimized LC DNA ratios. BsIgG1 yields >65% are indicated in bold. The HC of mAb-1 contained the ‘hole’ mutations (T366S:S368A:Y407V)8 and the HC for mAb-2 contained a ‘knob’ mutation (T366W).8 NA, not applicable: monospecific antibodies.
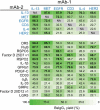 |
Figure 1.
High-resolution LCMS analysis profiles highlighting representative examples from Table 1 for (a and d) low (<30%), (b and e) intermediate (30–65%) or (c and f) high (>65%) yield of BsIgG1. Corresponding antibody pairs were transiently co-transfected into Expi293F cells. The IgG1 species were purified by protein A chromatography and SEC before quantification of the BsIgG1 yield by high-resolution LCMS.21,22 (a-c) Data shown are mass envelopes for charge states 38+ and 39+ along with (d-f) corresponding deconvoluted data. Also shown are cartoons representing the different IgG1 species present. See Table 1 for additional experimental details.
Enhanced yield of BsIgG1 using CH1/CL interface charge mutations for antibody pairs with a cognate HC/LC chain pairing preference
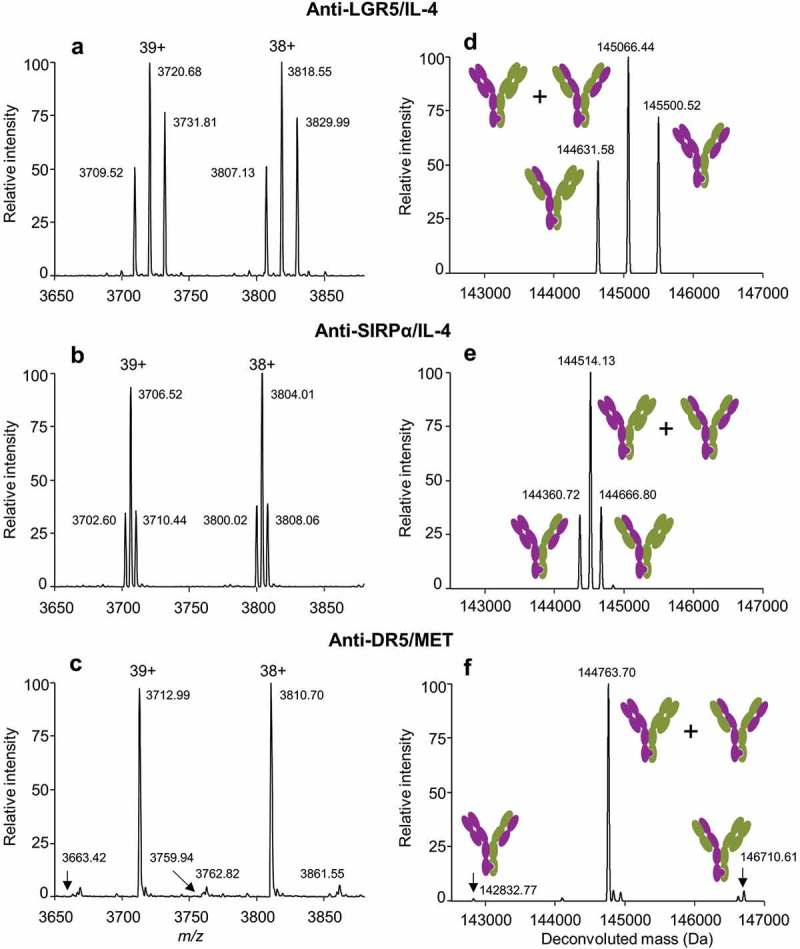
Previously, we used a combination of mutations at all four domain/domain interfaces (i.e., both VH/VL and both CH1/CL) in conjunction with KIH HC mutations for near quantitative assembly of BsIgG of different isotypes in single mammalian host cells.21 Here, we readily identified antibody pairs that give a high yield of BsIgG1 without any Fab mutations (Table 1). These high yields of BsIgG1 are presumably due to differences in variable domain sequences rather than constant domains (IgG1 CH1 and κ CL) that were identical in most cases. We hypothesized that for such antibody pairs mutations at the two CH1/CL interfaces alone might be sufficient to enhance the bispecific yield close to 100%. Eleven different antibody pairs were selected, and the yield of BsIgG1 was compared in the presence or absence of previously reported CH1/CL domain interface charge mutations.21 Specifically, the ‘knob’ arms were engineered with CL V133E and CH1 S183K mutations and the ‘hole’ arm with CL V133K and CH1 S183E mutations.21 The charge mutations at the two CH1/CL interfaces increased the BsIgG1 yield for all antibody pairs by ~12–34% to ≥90% BsIgG1 yield in the majority (9/11) of cases (Figure 2). The charge mutations at the two CH1/CL interfaces, however, have little to no impact on antigen binding (Table S2).
Figure 2.
Incorporation of CH1/CL charge pair variants increases BsIgG1 yields for antibody pairs that have a strong intrinsic HC/LC pairing preference. For the charge pair variants, the first listed antibody contains the CL V133E and CH1 S183K mutations and the second listed antibody contains the CL V133K and CH1 S183E mutations.21 See Table 1 for additional experimental details. 90% yield of BsIgG1 is indicated by a dotted horizontal line.
Cognate heavy/light chain pairing preference in one arm of a bispecific IgG can be sufficient for high bispecific yield
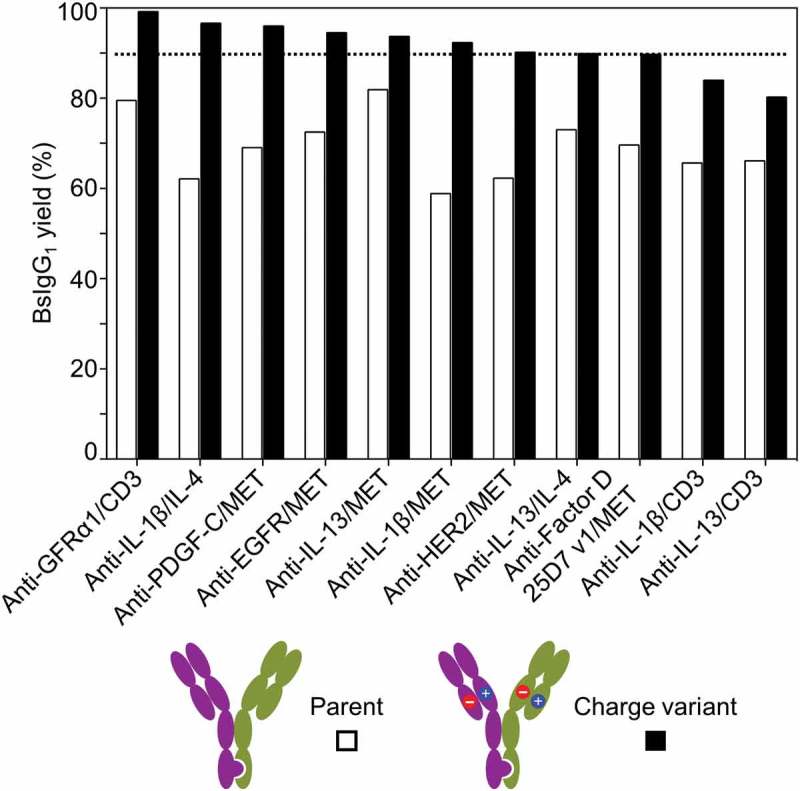
The mechanistic basis for high yield of BsIgG1 observed for some antibody pairs was investigated. Two antibody pairs, namely anti-EGFR/MET and anti-IL-13/IL-4, were selected for this study based on their high yield of BsIgG1 without Fab mutations (Table 1).21 A priori, either one or both Fab may exhibit a cognate HC/LC pairing preference contributing to the high yield of BsIgG1. Three chain co-expression experiments were undertaken to distinguish between these possibilities. A single HC (HC1) with either ‘knob’ or ‘hole’ mutations was transiently co-expressed in Expi293F cells with its cognate LC (LC1) and a competing non-cognate LC (LC2) (Figure 3(a)). The resultant half IgG species were purified from the corresponding cell culture supernatant by protein A affinity chromatography, and the extent of cognate and non-cognate HC/LC pairing was assessed by high-resolution LCMS21 (Figure 3(b)).
Figure 3.
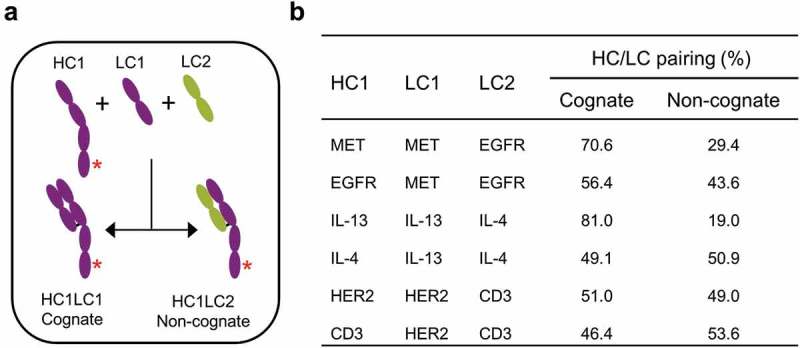
Investigation of cognate chain pairing preferences by co-expression of three chains. (a) Co-expression of an antibody heavy chain (HC1) with its cognate light chain (LC1) and a non-cognate light chain (LC2) in Expi293F cells. (b) Quantification of antibody cognate chain preferences following co-expression. The IgG1 and half IgG1 species were purified from the cell culture supernatant by protein A chromatography and analyzed by LCMS.21,22 The percentage of cognate HC/LC pairing was calculated by quantifying the half IgG1 species. The HC of anti-EGFR, anti-IL-13 and anti-HER2 contain a ‘knob’ mutation (T366W)8 whereas the HC of anti-MET, anti-IL-4 and anti-CD3 contain ‘hole’ mutations (T366S:S368A:Y407V).8 The asterisk (*) denotes the presence of either ‘knob’ or ‘hole’ mutations.
The anti-MET HC shows a strong preference for its cognate LC (~71%) over the non-cognate anti-EGFR LC, whereas the anti-EGFR HC shows only a slight preference for its cognate LC (~56%) over the non-cognate anti-MET LC (Figure 3(b)). The anti-IL-13 HC shows a strong preference for its cognate LC (81%) over the non-cognate anti-IL-4 LC, whereas the anti-IL-4 HC shows no preference (49%) for its cognate LC (Figure 3(b)). These data are consistent with the notion that the high BsIgG1 yield for anti-EGFR/MET results from the strong and weak cognate HC/LC pairing preference for the anti-MET and anti-EGFR antibodies, respectively. In contrast, the high BsIgG1 yield for anti-IL-13/IL-4 apparently reflects a strong cognate HC/LC pairing preference for the anti-IL-13 antibody alone. Thus, a cognate HC/LC pairing preference in one or both arms can apparently be sufficient for high yield of BsIgG1 in a single cell without the need for Fab mutations.
Anti-HER2/CD3 was selected as a control for this study based on its low yield of BsIgG1 (Table 1).21 The anti-HER2 HC shows no pairing preference for its cognate LC over the non-cognate anti-CD3 LC. Similarly, the anti-CD3 HC shows no pairing preference for its cognate LC over the non-cognate anti-HER2 LC (Figure 3(b)). However, in the absence of competition, HC can assemble efficiently with a cognate as well as with a non-cognate LC, as judged by all different matched and mismatched HC/LC pairs tested (Table S3).
CDR L3 and H3 of the anti-MET antibody contribute to high bispecific yield of anti-EGFR/MET BsIgG1
We investigated the sequence determinants in the anti-MET antibody contributing to high bispecific yield of the anti-EGFR/MET BsIgG1. The amino acid sequence differences between the anti-EGFR and anti-MET antibodies are located entirely within the CDRs plus one additional framework region (FR) residue, VH 94, immediately adjacent to CDR H3 (Figure S2). The remaining FR, plus Cκ and CH1 constant domain sequences of these antibodies are identical. CDR L3 and H3 are the CDRs that are most extensively involved at the VH/VL domain interface of the anti-MET antibody as evidenced by the X-ray crystallographic structure of the anti-MET Fab complexed with its antigen (Protein Data Bank (PDB) identification code 4K3J).25 These observations led to the hypothesis that CDR L3 and H3 of the anti-MET antibody may contribute to high bispecific yield for the anti-EGFR/MET BsIgG1. To test this hypothesis, we set out to perform CDR replacement experiments. We chose to replace CDR L3 and H3 from an anti-CD3 antibody that showed random pairing when combined with anti-HER2. The amino acid sequence of anti-CD3 is different from anti-MET or anti-EGFR within CDR L3 and H3 (Figure S2). The FR region (except VL 71 and VH 49, 71, 73, 94) plus Cκ and CH1 constant domain sequence of anti-CD3 is identical to anti-MET or anti-EGFR antibodies. Consistent with our hypothesis, replacement of both CDR L3 and H3 of the anti-MET antibody with corresponding sequences from an anti-CD3 antibody led to substantial loss of bispecific yield (~85% to 33%, Figure 4(a)). In contrast, replacement of both CDR L3 and H3 of the anti-EGFR arm of the anti-EGFR/MET bispecific resulted in only a small reduction in BsIgG yield (~85% to 75% Figure 4(a)). Replacement of CDR L3 and H3 for both anti-EGFR and anti-MET arms resulted in random HC/LC pairing. These data support the notion that CDR L3 and H3 of anti-MET and anti-EGFR antibodies make major and minor contributions, respectively, to the high bispecific yield observed for the anti-EGFR/MET BsIgG1. Replacement of CDR L1 and H1 or CDR L2 and H2 from the anti-MET antibody with corresponding anti-CD3 antibody sequences had little to no effect upon bispecific yield for the anti-EGFR/MET BsIgG1 (Figure S4).
Figure 4.
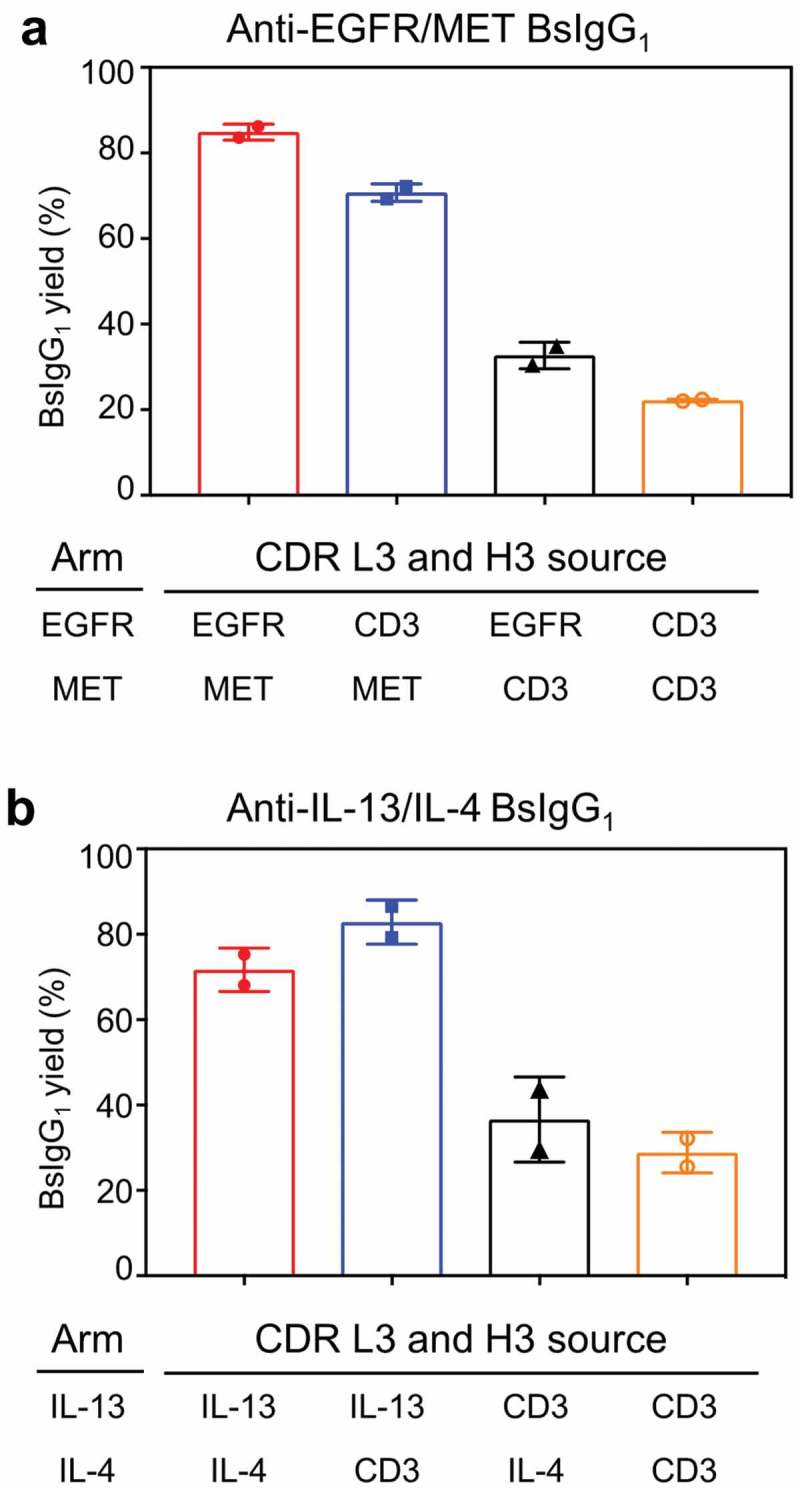
Contributions of CDR L3 and H3 to bispecific yield for (a) anti-EGFR/MET and (b) anti-IL-13/IL-4 BsIgG1. CDR L3 and H3 for one of both arms of each BsIgG1 were replaced with corresponding sequences from an anti-CD3 antibody to investigate their contribution to BsIgG1 yield. Data plotted are the mean ± SD of two independent experiments from optimized LC DNA ratios. See Table 1 for additional experimental details.
Residues within CDRs L3 and H3 of anti-MET antibody contributing to high bispecific yield of anti-EGFR/MET BsIgG1
The X-ray crystallographic structure of the anti-MET Fab (PDB accession code 4K3J)25 revealed contact residues between CDR L3 and H3 (Figure 5(a)) and was used to guide the selection of residues for mutational analysis. Alanine-scanning mutagenesis26 of anti-MET CDR L3 and H3 was used to map residues contributing to the high bispecific yield of anti-EGFR/MET BsIgG1 (Figure 5(b)). The VL Y91A mutation in CDR L3 gave the largest reduction in bispecific yield (84% to 57%) of any of the 12 single alanine mutants tested. As few as two alanine replacements in CDR L3, namely VL Y91A:Y94A, abolished the high bispecific yield (84% to 23%). Thus, CDR L3 residues VL Y91 and Y94 appear to make critical contributions to high bispecific yield for the anti-EGFR/MET BsIgG1. The expression titers of all the mutants were comparable to the parent BsIgG1 as estimated by the recovered yield from protein A chromatography (data not shown).
Figure 5.
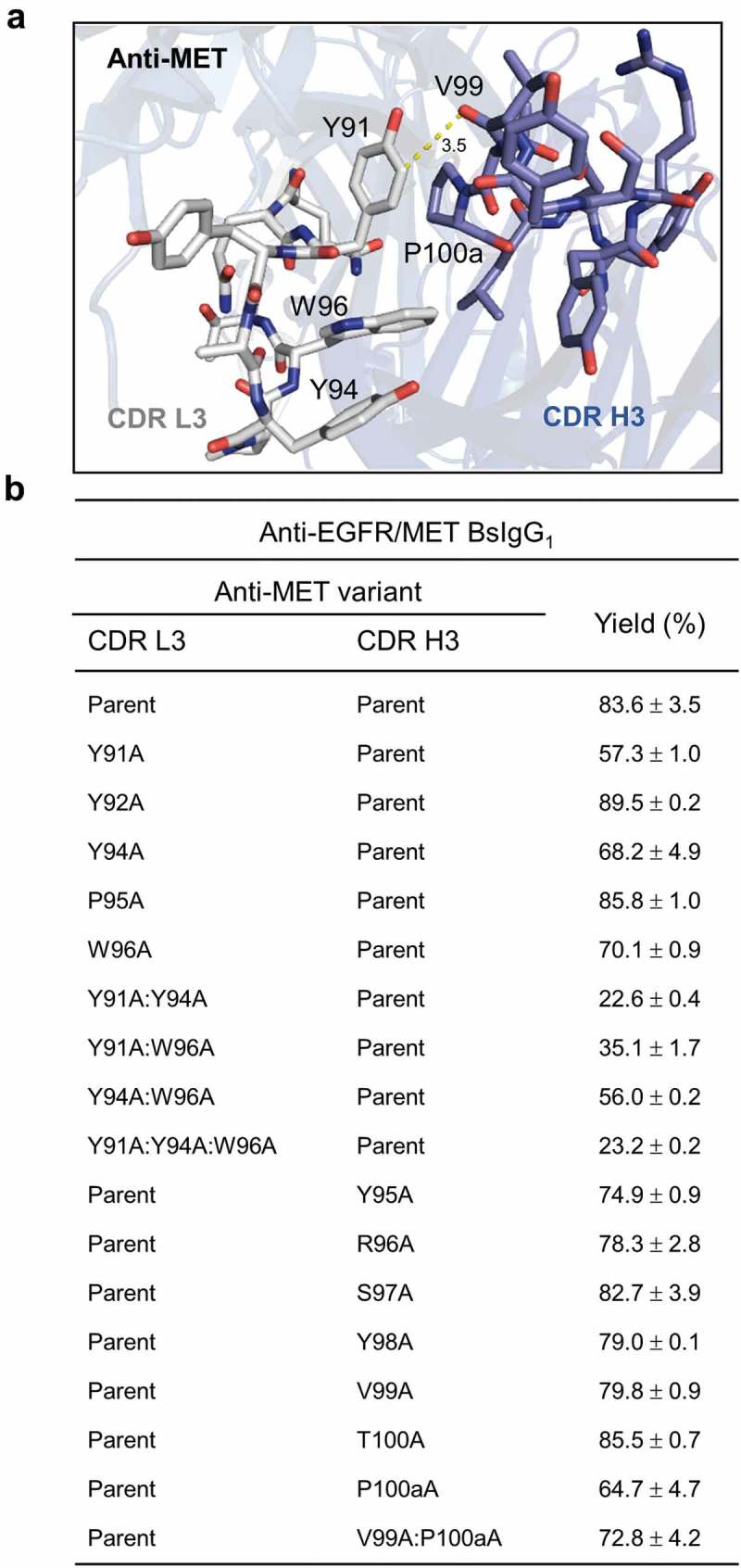
Structure-guided mutational analysis of CDR L3 and H3 contact residues for an anti-MET antibody. (a) X-ray structure of the anti-MET Fab (PDB 4K3J)25 highlighting CDR L3 and H3 contact residues. (b) Alanine-scanning mutagenesis of CDR L3 and H3 of the anti-MET antibody to identify residues contributing to high bispecific yield of the anti-EGFR/MET BsIgG1. Data shown represent mean ± SD for two independent experiments using optimized LC DNA ratios. See Table 1 for additional experimental details.
CDR L3 and H3 of anti-IL-13 antibody contribute to high bispecific yield for anti-IL-13/IL-4 BsIgG1
Specific residues in CDR L3 of the anti-MET antibody are important for high bispecific yield for the anti-EGFR/MET BsIgG1 (see above). We postulated that similar principles may apply to the anti-IL-13 antibody in contributing to high bispecific yield of the anti-IL-13/IL-4 BsIgG1. An analogous experimental strategy was used to investigate this possibility. One notable difference between these two antibody pairs is that the anti-IL-13 and anti-IL-4 antibodies differ in both their CDR and FR sequences (Figure S3) whereas the anti-MET and anti-EGFR antibodies have identical FR sequences (except for VH 94) and differ in their CDR sequences (Figure S2).
Replacement of CDR L3 and H3 of the anti-IL-13 antibody with corresponding sequences from an anti-CD3 antibody led to substantial loss of bispecific yield of the anti-IL-13/IL-4 BsIgG1 (~72% to 37%, Figure 4(b) and Figure S3). In contrast, a slight increase was observed when CDR L3 and H3 of the anti-IL-4 antibody were replaced in a similar manner (Figure 4(b)). These results suggest that CDR L3 and H3 of the anti-IL-13 antibody contribute to high bispecific yield of the anti-IL-13/IL-4 BsIgG1.
Alanine-scanning mutational analysis26 of anti-IL-13 CDR L3 and H3 was used to map residues contributing to the high bispecific yield of anti-IL-13/IL-4 BsIgG1. The X-ray crystallographic structure of the anti-IL-13 Fab in complex with IL-13 (PDB accession code 4I77)27 revealed the contact residues between CDR L3 and H3 (Figure 6(a)) and was used to select residues for mutational analysis (Figure 6(b)). The CDR L3 mutation VL R96A gave the largest reduction in bispecific yield of any of the nine single alanine mutants tested for CDRs L3 and H3 and abolished the high bispecific yield (72% to 29%). As few as two alanine replacements in CDR H3, namely VH D95A:P99A, also abolished the high bispecific yield (72% to 26%). The expression titers of all the mutants were comparable to the parent BsIgG1 as estimated by the recovered yield from protein A chromatography (data not shown).
Figure 6.
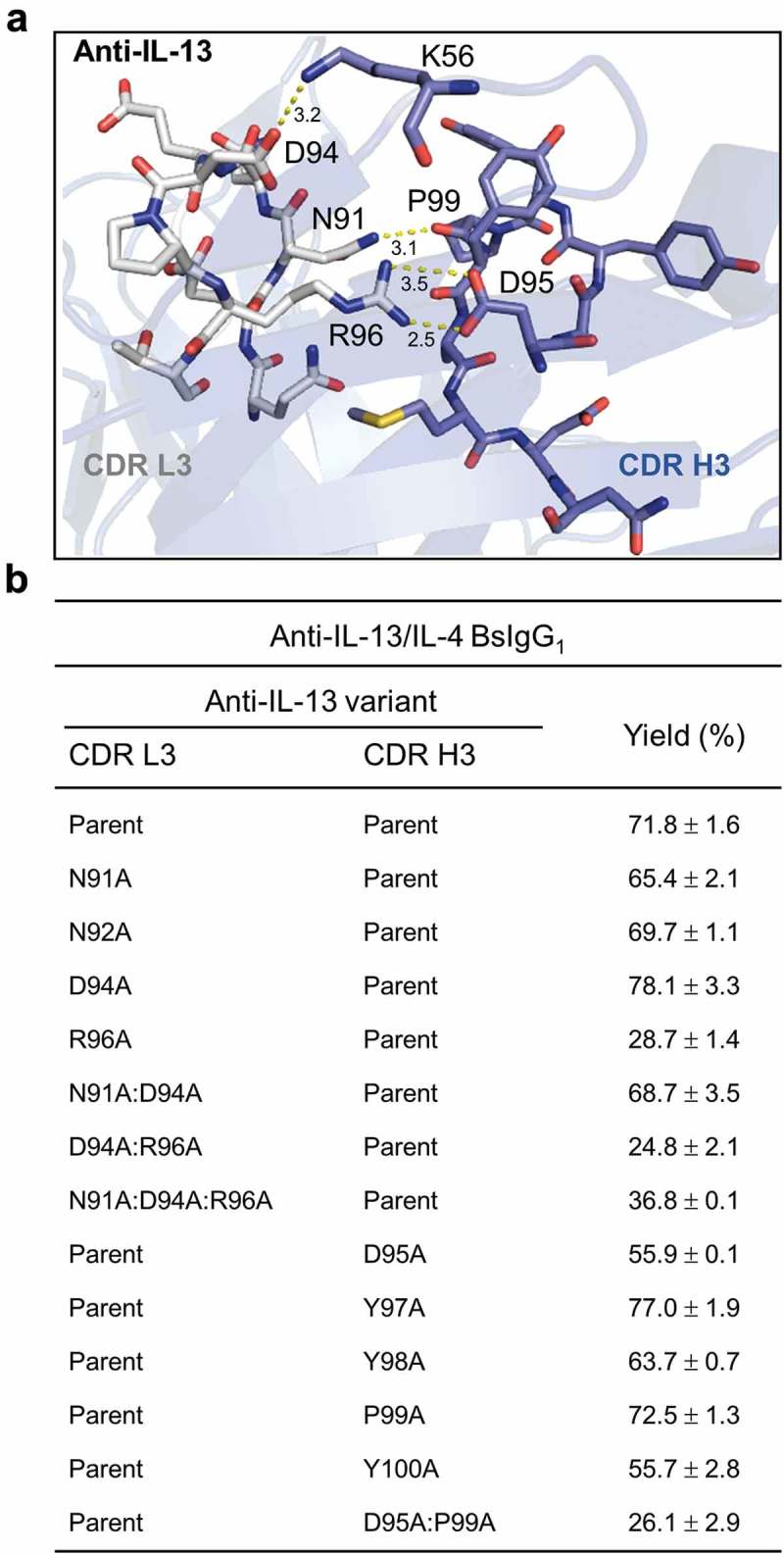
Structure-guided mutational analysis of CDR L3 and H3 contact residues for an anti-IL-13 antibody. (a) X-ray structure of the anti-IL-13 Fab (PDB 4I77)27 highlighting CDR L3 and H3 contact residues. (b) Alanine-scanning mutagenesis of CDR L3 and H3 of anti-IL-13 to identify residues contributing to high bispecific yield of anti-IL-13/IL-4 BsIgG1. Data shown represent mean ± SD for two independent experiments using optimized LC DNA ratios. See Table 1 for additional experimental details.
CDR L3 and H3 can increase bispecific yield for BsIgG1
Critical contributions to high bispecific yield can be made by CDR L3 and H3 as judged by both the anti-EGFR/MET and anti-IL-13/IL-4 BsIgG1 studied here. Next, we investigated whether CDR L3 and H3 from these antibodies could be sufficient for providing high bispecific yield for other antibody pairs. We selected two antibody pairs that have low bispecific yield, namely anti-HER2/CD3 (22–24%) (Table 1)21 and anti-VEGFA/ANG2 (24%).21 CDR L3 and H3 for one arm each of these two BsIgG1 were replaced with corresponding CDR sequences from either the anti-MET or anti-IL-13 antibodies. A substantial increase in yield of BsIgG1 (from ~24% up to 40–65%) was observed in three of four CDR L3 and H3 recruitment cases for both anti-HER2/CD3 (Figure 7(a)) and anti-VEGFA/ANG2 (Figure 7(b)). Thus, recruitment of CDR L3 and H3 from antibodies with a cognate HC/LC pairing preference can enhance the yield of BsIgG1 with no pairing preference but does not invariably do so.
Figure 7.
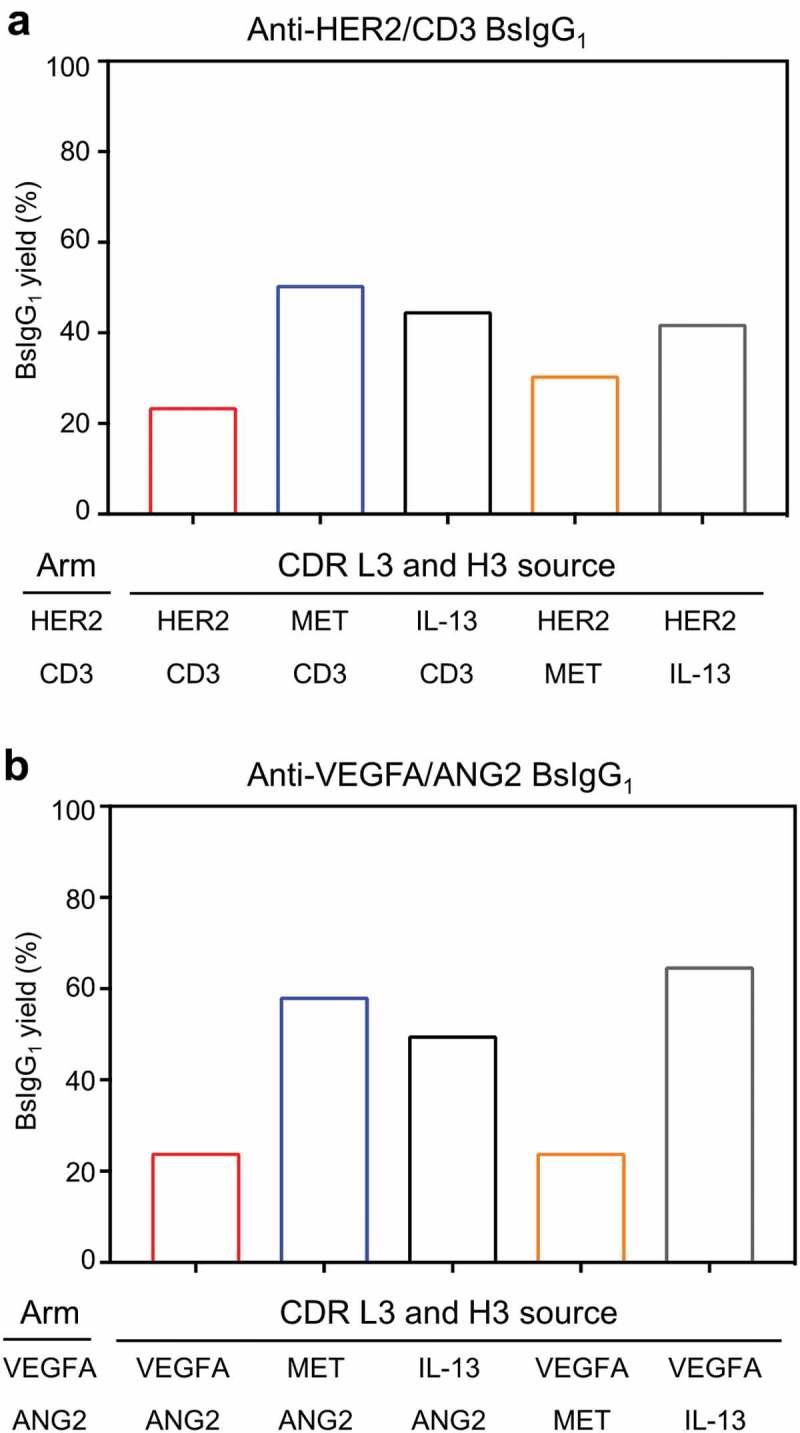
Recruitment of CDR L3 and H3 from either anti-MET or anti-IL-13 antibodies into other antibody pairs to determine the effect upon BsIgG1 yield. CDR L3 and H3 of (a) anti-HER2 and anti-CD3 antibodies and (b) anti-VEGFA and anti-ANG2 antibodies were replaced with CDR L3 and H3 of either the anti-MET or anti-IL-13 antibodies. The data presented are from optimized LC DNA ratios. The BsIgG1 yield was determined as for Table 1.
When critical residues for pairing preference for anti-IL-13 were transplanted to the corresponding position in anti-HER2, anti-VEGFA or anti-VEGFC antibodies, some increase in bispecific yield was observed, albeit less than for the parental anti-IL-13/IL-4 BsIgG1 (Table S4). Together, these results suggested that CDR L3 and H3 can impart pairing preference for some, but not all, antibody pairs.
HC-LC inter-chain disulfide bond does not affect the yield of BsIgG1
Previously, we hypothesized that formation of the interchain disulfide bond between the HC and LC acts as a kinetic trap that prevents chain exchange.21 Here we investigated if the disulfide bond between HC and LC affects the bispecific yield for two BsIgG1 with a pronounced cognate chain preference (anti-EGFR/MET and anti-IL-13/IL-4) and two controls with random HC/LC pairing (anti-HER2/CD3 and anti-VEGFA/VEGFC). BsIgG1 variants lacking the inter-chain disulfide bond were generated using cysteine to serine mutations: LC C214S and HC C220S. Removal of the inter-chain disulfide bond in the engineered variants was verified by SDS PAGE (Figure S5). No clear evidence was found that the inter-chain disulfide bond affects BsIgG1 yield for any of the four antibody pairs tested as judged by native mass spectrometry (Table 2). The yields of BsIgG1 of the parental and the inter-chain disulfide bond lacking variants were similar.
Table 2.
Mutational analysis suggests that the disulfide bond between HC and LC does not affect BsIgG1 yield. Experimental conditions were similar to those described in Table 1 legend except that the quantification of BsIgG1 utilized native LCMS (see Materials and Methods).28 Variants lacking the interchain disulfide bond were constructed using CL C214S and CH1 C220S mutations. Data shown are the mean ± SD for three biological replicates using optimized DNA LC ratios.
| BsIgG1 yield (%) |
||
|---|---|---|
| BsIgG1 | Parent with HC/LC disulfide bond | Variant without HC/LC disulfide bond |
| Anti-EGFR/MET | 81.1 ± 1.4 | 82.8 ± 2.6 |
| Anti-IL-13/IL-4 | 73.3 ± 4.5 | 75.1 ± 0.8 |
| Anti-HER2/CD3 | 24.5 ± 0.8 | 27.0 ± 2.4 |
| Anti-VEGFA/VEGFC | 28.8 ± 5.9 | 38.0 ± 6.0 |
Discussion
BsAb, including BsIgG, holds significant promise as next-generation antibody drugs prompting much interest in the development of robust and efficient production methods for these complex molecules. BsIgG have been efficiently produced in single host cells by multiple strategies, including a combination of Fab and Fc engineering to favor cognate over non-cognate HC/LC pairing and HC heterodimerization over homodimerization, respectively.16–21,24 Assessing the success of such Fab engineering requires comparison of variants with and without Fab mutations. For example, we transiently co-expressed two different HC and LC in mammalian cells, purified the mixture of different IgG species, and then determined the percentage of BsIgG present (defined here as BsIgG yield) by high-resolution LCMS.21,22 Two antibody pairs gave >65% BsIgG1 with KIH mutations for HC heterodimerization, but without Fab mutations to favor cognate HC/LC pairing.21 This observation suggests the existence of a strong intrinsic preference for cognate HC/LC pairing for some antibody pairs that has also been reported by other groups.16,24 We sought to better understand cognate HC/LC pairing preference, including strength and frequency of occurrence of this phenomenon. Additionally, this chain pairing preference information was used to reduce the number of Fab arm mutations needed for the efficient production of BsIgG1 in single host cells for some antibody pairs.
In this study, we extended prior work on understanding cognate HC/LC preferences by expanding upon the small number of previously reported BsIgG examples.16,21,24 We produced 99 different BsIgG1 with KIH HC, but without Fab mutations. Strikingly, non-random HC/LC pairing (>30% yield BsIgG1) was observed for most (>80%) antibody pairs, with high (>65%) BsIgG1 yield seen for one-third of antibody pairs (Table 1). The observed differences in BsIgG1 yield for different antibody pairs are presumably driven by VH and VL sequence differences because most antibodies evaluated here share identical human CH1 and κ CL sequences. A priori, either one or both Fab arms of a bispecific may exhibit a cognate HC/LC pairing preference contributing to the high yield of BsIgG1. Evidence of each of these possibilities was found from co-expression of different HC in the presence of competing cognate and non-cognate LC (Figure 3).
One caveat to the interpretation of CDR replacement experiments discussed here is that the observed effect upon BsIgG1 yield is a combination of both removal of the original CDRs as well as the newly installed CDR. This inherent methodological limitation notwithstanding, the diverse data types present in this study (three chain co-expression, CDR replacements and CDR point mutations) are consistent with the notion that CDR L3 and H3, and specific residues within these CDRs can contribute to cognate HC/LC pairing preference. This hypothesis appears plausible since CDR L3 and H3 interactions can contribute to VH/VL domain interfaces as evidenced by X-ray crystallographic studies.16,17,20 Replacements of CDR L3 and H3 and mutations therein are likely to affect antigen binding. Since the major focus of this study was to understand the molecular basis of HC/LC chain pairing preferences, we did not measure the affinities for antigen binding. Optimization and utilization of the molecular understanding of HC/LC chain pairing preference for antigen binding are beyond the scope of this investigation. Removal of the inter-chain disulfide bond by mutagenesis had no clear effect upon BsIgG1 yield (Table 2), suggesting HC/LC pairs are not readily prone to exchange.
BsIgG designed for efficient production in single cells has in several cases utilized mutations at all four Fab domain interfaces, i.e., VH/VL and CH1/CL in both Fab arms.16,17,20,21 Extensively engineered proteins can be successfully developed as drugs, as exemplified by the BsIgG1, emicizumab.4,5,29–31 This achievement notwithstanding, reducing the number of Fab arm mutations for assisting BsIgG production in single cells is possible for many antibody pairs and may be desirable as discussed below. The many potential benefits of introducing mutations to enhance antibody and other protein drug candidates2,32 need to be weighed against possible untoward effects that can occasionally occur. Undesirable attributes occasionally encountered with engineered proteins include off-target binding and faster pharmacokinetic clearance,33 reduced thermostability,8,20,34 reduced affinity for binding partners,34,35 introduction of chemical degradation ‘hotspots’,36 and higher incidence of anti-drug antibodies (ADA) elicited in patients.37 Many such risks can be identified and mitigated through preclinical experimentation as elegantly demonstrated by the elimination of untoward off-target binding of an affinity-matured anti-RSV antibody.33 Other developability risks with engineered proteins are more challenging to assess preclinically, such as the likelihood for eliciting neutralizing ADAs in patients.38,39
Ideally, BsIgG would be produced in single cells without the need for Fab mutations as recently demonstrated to be possible for one antibody pair.24 Here we identified two additional examples of near quantitative (>90%) formation of BsIgG1 using KIH HC but without Fab mutations (Table 1). However, such a high stringency, inherent HC/LC pairing preference was rare and observed here in only ~2% of the antibody pairs we surveyed. The development of methods to identify or screen for such antibody pairs may be helpful in constructing BsIgG and other BsAb. The use of engineered disulfide bonds to enhance the assembly of BsIgG1 was first shown in the context of promoting HC heterodimerization.9 More recently, the native disulfide bond between CH1 and CL domains was replaced with an engineered disulfide between these domains for enhancing the production of BsIgG1 in single cells.18 In this case, efficient assembly of BsIgG1 was achieved with as few as four amino-acid replacements.18 The native disulfide bond between HC and LC has also been replaced with an engineered disulfide between VH and VL for robust one cell production of BsIgG.19 Comparison of different sites suggests that installing the engineered disulfide at the VH/VL rather than CH1/CL interface is the preferred site to prevent non-cognate VH/VL pairing.
Initial studies to assist BsIgG production in single cells by engineering the CH1/CL interfaces alone met with limited success and led to the idea that variable domains can dominate HC/LC pairing preference.16 We later demonstrated that a different set of CH1/CL mutations increased the BsIgG1 yield somewhat for an anti-HER2/CD3 BsIgG1 with random HC/LC pairing.21 More recently, Bonisch et al. demonstrated that alternative CH1/CL mutations alone could promote near quantitative formation of BsIgG1 for two antibody pairs with a strong cognate chain pairing preference.24 Here, we further developed this concept by showing that antibodies with a strong cognate pairing preference occur commonly, comprising one-third of the antibody pairs surveyed here (Table 1). Additionally, we demonstrate that for most antibody pairs (9 of 11) with a strong (>65%) HC/LC pairing preference, CH1/CL mutations (same as previously used)21 in conjunction with KIH HC are often sufficient to support near quantitative yield of BsIgG1 (Figure 2). Thus, the number of Fab arm mutations to facilitate efficient BsIgG1 assembly was reduced from a minimum of eight or more in our previous study21 to just four in some cases (Figure 2.) or even zero in rare (~2%) instances (Table 1).
In summary, this study demonstrates that a cognate HC/LC pairing preference in producing BsIgG in single cells is a common phenomenon that is highly dependent upon the specific antibody pair. Mechanistically, this chain pairing preference can be strongly influenced by residues in CDR L3 and H3. Practically, this cognate HC/LC pairing preference was used to reduce the number of Fab mutations needed to drive the production of BsIgG1, and potentially BsIgG of other isotypes, in single cells.
Materials and methods
Antibody construct design and synthesis
All antibodies in this work are numbered using the Kabat40 and EU41 numbering systems for variable and constant domains, respectively. Antibody constructs were generated by gene synthesis (GeneWiz), and wherever applicable, sub-cloned into the expression plasmid (pRK5) as described previously.21 All antibody HC in this study were aglycosylated (N297G mutation) and with the carboxy-terminal lysine deleted (∆K447) to reduce product heterogeneity, and thereby facilitate accurate quantification of BsIgG1 by LCMS.21,22 The two-component HC of all BsIgG1 in this study were engineered to contain either a ‘knob’ mutation (T366W)8 in the first listed antibody or ‘hole’ mutations (T366S:L368A:Y407V)8 in the second listed antibody to facilitate HC heterodimerization.8
For a few of the BsIgG1 in this study, FR mutations were judiciously made to provide sufficient mass difference between correctly paired and mispaired BsIgG1 species for more accurate quantitation by LCMS analysis. The mass difference needed for accurate quantification of bispecific IgG yield is ≤118 Da.22 Specifically, the antibodies and mutations were anti-HER2 VL R66G when combined with anti-CD3 or variants (Figure 7(a)), anti-IL-1β or anti-GFRα (Table 1); anti-VEGFA VL F83A when combined with anti-ANG2 or variants (in Figure 7(b)); anti-CD3 VL N34A:F83A when combined with anti-Factor D 25D7 v1 or anti-IL-33; anti-RSPO3 VL F83A, when combined with anti-CD3; anti-EGFR VL F83A when combined with anti-SIRPα or anti-Factor D 20D12 v1; plus anti-IL-4 VL N31A:F83A when combined with anti-GFRα1 (Table 1 or Figure 1). The chosen residues had no detectable impact on BsIgG1 yield based upon comparison with parental antibodies.
Antibody expression and purification
All BsIgG1 were transiently expressed in Expi293F cells as described previously.21 Four plasmids corresponding to the two LC and two HC were co-transfected into Expi293F cells. The LC DNA was varied for each experiment and the highest bispecific yield with the optimal LC:HC ratio was reported as described previously.21 Briefly, the LC:HC ratios used in this study were 1:3, 1:2, 1:1, 2:1 or 3:1. The ratio of the two HC was not optimized and fixed at 1:1. The optimized LC:HC DNA ratios from the four chain co-expression experiments were used for experiments in Figure 3. The LC1:LC2 DNA ratios used were 1:1 for all except 1:2 for EGFR:MET. The transfected cell culture (30 mL) was grown for 7 days at 37°C with shaking. BsIgG1 from the filtered cell culture supernatants were purified in a high throughput fashion by Toyopearl AF-rProtein A affinity chromatography (Tosoh Bioscience). Impurities such as aggregates and half IgG1 were removed by SEC using a Zenix-C SEC-300 column (10 mm × 300 mm, 3 μm particle size, Sepax Technology). The IgG1 concentration was calculated using an extinction coefficient A0.1%280nm of 1.4. Purification yield was estimated after protein A chromatography by multiplying the protein concentration with elution volume.
Analytical characterization of BsIgG1 by SEC HPLC
BsIgG1 samples (20 μL) were chromatographed under isocratic conditions on a TSKgel SuperSW3000 column (4.6 × 150 mm, 4 μm) (Tosoh Bioscience) connected to a Dionex UltiMate 3000 HPLC (Thermo Scientific). The mobile phase was 200 mM potassium phosphate and 250 mM potassium chloride at pH 7.2 with a flow rate of 0.3 mL/min with absorbance measurement at a wavelength of 280 nm.
BsIgG1 yield determination by high-resolution LCMS
Quantification of BsIgG1 yield (intensity of correctly paired LC species over all three mispaired IgG1 species) was performed using a Thermo Exactive Plus Extended Mass Range Orbitrap mass spectrometer as described previously and assumes no response bias amongst the different mass peaks.22
For denaturing mass spectrometry, samples (3 μg) were injected onto a MAbPac reversed-phase liquid chromatography column (Thermo Fisher Scientific, 2.1 mm × 50 mm) heated to 80°C using a Dionex Ultimate 3000 RSLC system. A binary gradient pump was used to deliver solvent A (99.88% water containing 0.1% formic acid and 0.02% trifluoroacetic acid) and solvent B (90% acetonitrile containing 9.88% water plus 0.1% formic acid and 0.02% trifluoroacetic acid) as a gradient of 20% to 65% solvent B over 4.5 min at 300 μL/min. The solvent was step-changed to 90% solvent B over 0.1 min and held at 90% for 6.4 min to clean the column. Finally, the solvent was step-changed to 20% solvent B over 0.1 min and held for 3.9 min for re-equilibration. Samples were analyzed online via electrospray ionization into the mass spectrometer using the following parameters for data acquisition: 3.90 kV spray voltage; 325°C capillary temperature; 200 S-lens RF level; 15 sheath gas flow rate and 4 AUX gas flow rate in ESI source; 1,500 to 6,000 m/z scan range; desolvation, in-source CID 100 eV, CE 0; resolution of 17,500 at m/z 200; positive polarity; 10 microscans; 3E6 AGC target; fixed AGC mode; 0 averaging; 25 V source DC offset; 8 V injection flatapole DC; 7 V inter-flatapole lens; 6 V bent flatapole DC; 0 V transfer multipole DC tune offset; 0 V C-trap entrance lens tune offset; and trapping gas pressure setting of 2.
For native mass spectrometry, samples (10 μg) were injected onto an Acquity UPLC BEH SEC column (Waters, 4.6 mm × 150 mm) heated to 30°C using a Dionex Ultimate 3000 RSLC system. Isocratic chromatography runs (10 min) utilized an aqueous mobile phase containing 50 mM ammonium acetate at pH 7.0 with a flow rate of 300 μL/min.
Samples were analyzed online via electrospray ionization into the mass spectrometer using the following parameters for data acquisition: 4.0 kV spray voltage; 320°C capillary temperature; 200 S-lens RF level; 40 sheath gas flow rate and 20 AUX gas flow rate in ESI source; 300 to 20,000 m/z scan range; desolvation, in-source CID 100 eV, CE 0; resolution of 17,500 at m/z 200; positive polarity; 10 microscans; 1E6 AGC target; fixed AGC mode; 0 averaging; 25 V source DC offset; 8 V injection flatapole DC; 7 V inter-flatapole lens; 6 V bent flatapole DC; 0 V transfer multipole DC tune offset; 0 V C-trap entrance lens tune offset; and trapping gas pressure setting of 2.
Acquired mass spectral data were analyzed using Protein Metrics Intact Mass and Thermo BioPharma Finder 3.0 software. The signal intensity of the correctly paired LC species from the deconvolved spectrum of each sample was used for quantification relative to the three mispaired IgG1 species. HC homodimers and half IgG were either undetectable or present in trace amounts and excluded from the calculations. The correctly LC paired BsIgG1 were estimated from the isobaric mixture of BsIgG1 and the double LC mispaired IgG1 by using the algebraic formula described previously.22
Abbreviations
| BsAb | bispecific antibodies |
| BsIgG | bispecific IgG |
| CDR | complementarity-determining region |
| EMR | extended mass range |
| FR | framework region |
| HC | heavy chain |
| KIH | knob-into-hole |
| LC | light chain |
| LCMS | liquid chromatography-mass spectrometry |
| NA | not applicable |
| ND | not detected |
| PDB | Protein Data Bank |
| SD | standard deviation |
| SEC | size-exclusion chromatography |
Acknowledgments
We thank the Research Materials Group in the Early Stage Cell Culture Department at Genentech for mammalian expression and the Antibody Production Group in the Antibody Engineering Department for protein purification. We thank Kobe Yuen, for help in generating Table 1 using the R software tool.
Disclosure of potential conflicts of interest
All authors are current or former employees of Genentech, Inc., which develops and commercializes therapeutics including bispecific antibodies. This work was funded by Genentech, Inc.
Supplementary material
Supplemental data for this article can be accessed on the publisher’s website.
References
- 1.Brinkmann U, Kontermann RE.. The making of bispecific antibodies. mAbs. 2017;9:182–212. doi: 10.1080/19420862.2016.1268307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Carter PJ, Lazar GA. Next generation antibody drugs: pursuit of the ‘high-hanging fruit’. Nat Rev Drug Discov. 2018;17:197–223. doi: 10.1038/nrd.2017.227. [DOI] [PubMed] [Google Scholar]
- 3.Sanford M. Blinatumomab: first global approval. Drugs. 2015;75:321–27. doi: 10.1007/s40265-015-0356-3. [DOI] [PubMed] [Google Scholar]
- 4.Oldenburg J, Mahlangu JN, Kim B, Schmitt C, Callaghan MU, Young G, Santagostino E, Kruse-Jarres R, Negrier C, Kessler C, et al. Emicizumab prophylaxis in hemophilia A with inhibitors. N Engl J Med. 2017;377:809–18. doi: 10.1056/NEJMoa1703068. [DOI] [PubMed] [Google Scholar]
- 5.Scott LJ, Kim ES. Emicizumab-kxwh: first global approval. Drugs. 2018;78:269–74. doi: 10.1007/s40265-018-0861-2. [DOI] [PubMed] [Google Scholar]
- 6.Suresh MR, Cuello AC, Milstein C. Bispecific monoclonal antibodies from hybrid hybridomas. Methods Enzymol. 1986;121:210–28. [DOI] [PubMed] [Google Scholar]
- 7.Spiess C, Zhai Q, Carter PJ. Alternative molecular formats and therapeutic applications for bispecific antibodies. Mol Immunol. 2015;67:95–106. doi: 10.1016/j.molimm.2015.01.003. [DOI] [PubMed] [Google Scholar]
- 8.Atwell S, Ridgway JB, Wells JA, Carter P. Stable heterodimers from remodeling the domain interface of a homodimer using a phage display library. J Mol Biol. 1997;270:26–35. doi: 10.1006/jmbi.1997.1116. [DOI] [PubMed] [Google Scholar]
- 9.Merchant AM, Zhu Z, Yuan JQ, Goddard A, Adams CW, Presta LG, Carter P. An efficient route to human bispecific IgG. Nat Biotechnol. 1998;16:677–81. doi: 10.1038/nbt0798-677. [DOI] [PubMed] [Google Scholar]
- 10.Ridgway JB, Presta LG, Carter P. ‘Knobs-into-holes’ engineering of antibody CH3 domains for heavy chain heterodimerization. Protein Eng. 1996;9:617–21. [DOI] [PubMed] [Google Scholar]
- 11.Fischer N, Elson G, Magistrelli G, Dheilly E, Fouque N, Laurendon A, Gueneau F, Ravn U, Depoisier JF, Moine V, et al. Exploiting light chains for the scalable generation and platform purification of native human bispecific IgG. Nat Commun. 2015;6:6113. doi: 10.1038/ncomms7113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Labrijn AF, Meesters JI, de Goeij BE, van Den Bremer ET, Neijssen J, van Kampen MD, Strumane K, Verploegen S, Kundu A, Gramer MJ, et al. Efficient generation of stable bispecific IgG1 by controlled Fab-arm exchange. Proc Natl Acad Sci U S A. 2013;110:5145–50. doi: 10.1073/pnas.1220145110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Spiess C, Merchant M, Huang A, Zheng Z, Yang NY, Peng J, Ellerman D, Shatz W, Reilly D, Yansura DG, et al. Bispecific antibodies with natural architecture produced by co-culture of bacteria expressing two distinct half-antibodies. Nat Biotechnol. 2013;31:753–58. doi: 10.1038/nbt.2621. [DOI] [PubMed] [Google Scholar]
- 14.Strop P, Ho WH, Boustany LM, Abdiche YN, Lindquist KC, Farias SE, Rickert M, Appah CT, Pascua E, Radcliffe T, et al. Generating bispecific human IgG1 and IgG2 antibodies from any antibody pair. J Mol Biol. 2012;420:204–19. doi: 10.1016/j.jmb.2012.04.020. [DOI] [PubMed] [Google Scholar]
- 15.Schaefer W, Regula JT, Bahner M, Schanzer J, Croasdale R, Durr H, Gassner C, Georges G, Kettenberger H, Imhof-Jung S, et al. Immunoglobulin domain crossover as a generic approach for the production of bispecific IgG antibodies. Proc Natl Acad Sci U S A. 2011;108:11187–92. doi: 10.1073/pnas.1019002108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lewis SM, Wu X, Pustilnik A, Sereno A, Huang F, Rick HL, Guntas G, Leaver-Fay A, Smith EM, Ho C, et al. Generation of bispecific IgG antibodies by structure-based design of an orthogonal Fab interface. Nat Biotechnol. 2014;32:191–98. doi: 10.1038/nbt.2797. [DOI] [PubMed] [Google Scholar]
- 17.Froning KJ, Leaver-Fay A, Wu X, Phan S, Gao L, Huang F, Pustilnik A, Bacica M, Houlihan K, Chai Q, et al. Computational design of a specific heavy chain/kappa light chain interface for expressing fully IgG bispecific antibodies. Protein Sci. 2017;26:2021–38. doi: 10.1002/pro.3240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mazor Y, Oganesyan V, Yang C, Hansen A, Wang J, Liu H, Sachsenmeier K, Carlson M, Gadre DV, Borrok MJ, et al. Improving target cell specificity using a novel monovalent bispecific IgG design. MAbs. 2015;7:377–89. doi: 10.1080/19420862.2015.1007816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Vaks L, Litvak-Greenfeld D, Dror S, Matatov G, Nahary L, Shapira S, Hakim R, Alroy I, Benhar I. Design principles for bispecific IgGs, opportunities and pitfalls of artificial disulfide bonds. Antibodies. 2018;7. doi: 10.3390/antib7030027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liu Z, Leng EC, Gunasekaran K, Pentony M, Shen M, Howard M, Stoops J, Manchulenko K, Razinkov V, Liu H, et al. A novel antibody engineering strategy for making monovalent bispecific heterodimeric IgG antibodies by electrostatic steering mechanism. J Biol Chem. 2015;290:7535–62. doi: 10.1074/jbc.M114.620260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Dillon M, Yin Y, Zhou J, McCarty L, Ellerman D, Slaga D, Junttila TT, Han G, Sandoval W, Ovacik MA, et al. Efficient production of bispecific IgG of different isotypes and species of origin in single mammalian cells. MAbs. 2017;9:213–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yin Y, Han G, Zhou J, Dillon M, McCarty L, Gavino L, Ellerman D, Spiess C, Sandoval W, Carter PJ. Precise quantification of mixtures of bispecific IgG produced in single host cells by liquid chromatography-Orbitrap high-resolution mass spectrometry. MAbs. 2016;8:1467–76. doi: 10.1080/19420862.2016.1232217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Schaefer W, Völger HR, Lorenz S, Imhof-Jung S, Regula JT, Klein C, Mølhøj M. Heavy and light chain pairing of bivalent quadroma and knobs-into-holes antibodies analyzed by UHR-ESI-QTOF mass spectrometry. MAbs. 2016;8:49–55. doi: 10.1080/19420862.2015.1111498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bönisch M, Sellmann C, Maresch D, Halbig C, Becker S, Toleikis L, Hock B, Ruker F. Novel CH1: cLinterfaces that enhance correct light chain pairing in heterodimeric bispecific antibodies. Protein Eng Des Sel. 2017;30:685–96. doi: 10.1093/protein/gzx044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Merchant M, Ma X, Maun HR, Zheng Z, Peng J, Romero M, Huang A, Yang NY, Nishimura M, Greve J, et al. Monovalent antibody design and mechanism of action of onartuzumab, a MET antagonist with anti-tumor activity as a therapeutic agent. Proc Natl Acad Sci U S A. 2013;110:E2987–96. doi: 10.1073/pnas.1302725110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cunningham BC, Wells JA. High-resolution epitope mapping of hGH-receptor interactions by alanine-scanning mutagenesis. Science. 1989;244:1081–85. doi: 10.1126/science.2471267. [DOI] [PubMed] [Google Scholar]
- 27.Ultsch M, Bevers J, Nakamura G, Vandlen R, Kelley RF, Wu LC, Eigenbrot C. Structural basis of signaling blockade by anti-IL-13 antibody lebrikizumab. J Mol Biol. 2013;425:1330–39. doi: 10.1016/j.jmb.2013.01.024. [DOI] [PubMed] [Google Scholar]
- 28.Schachner L, Han G, Dillon M, Zhou J, McCarty L, Ellerman D, Yin Y, Spiess C, Lill JR, Carter PJ, et al. Characterization of chain pairing variants of bispecific IgG expressed in a single host cell by high-resolution native and denaturing mass spectrometry. Anal Chem. 2016;88:12122–27. doi: 10.1021/acs.analchem.6b02866. [DOI] [PubMed] [Google Scholar]
- 29.Kitazawa T, Igawa T, Sampei Z, Muto A, Kojima T, Soeda T, Yoshihashi K, Okuyama-Nishida Y, Saito H, Tsunoda H, et al. A bispecific antibody to factors IXa and X restores factor VIII hemostatic activity in a hemophilia A model. Nature Medicine. 2012;18:1570–74. doi: 10.1038/nm.2942. [DOI] [PubMed] [Google Scholar]
- 30.Sampei Z, Igawa T, Soeda T, Funaki M, Yoshihashi K, Kitazawa T, Muto A, Kojima T, Nakamura S, Hattori K. Non-antigen-contacting region of an asymmetric bispecific antibody to factors IXa/X significantly affects factor VIII-mimetic activity. MAbs. 2015;7:120–28. doi: 10.4161/19420862.2015.989028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sampei Z, Igawa T, Soeda T, Okuyama-Nishida Y, Moriyama C, Wakabayashi T, Tanaka E, Muto A, Kojima T, Kitazawa T, et al. Identification and multidimensional optimization of an asymmetric bispecific IgG antibody mimicking the function of factor VIII cofactor activity. PLoS One. 2013;8:e57479. doi: 10.1371/journal.pone.0057479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Carter PJ. Introduction to current and future protein therapeutics: a protein engineering perspective. Exp Cell Res. 2011. doi: 10.1016/j.yexcr.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 33.Wu H, Pfarr DS, Johnson S, Brewah YA, Woods RM, Patel NK, White WI, Young JF, Kiener PA. Development of motavizumab, an ultra-potent antibody for the prevention of respiratory syncytial virus infection in the upper and lower respiratory tract. J Mol Biol. 2007;368:652–65. doi: 10.1016/j.jmb.2007.02.024. [DOI] [PubMed] [Google Scholar]
- 34.Cooke HA, Arndt J, Quan C, Shapiro RI, Wen D, Foley S, Vecchi MM, Preyer M. EFab domain substitution as a solution to the light-chain pairing problem of bispecific antibodies. MAbs. 2018;10:1248–59. doi: 10.1080/19420862.2018.1519631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tiller KE, Li L, Kumar S, Julian MC, Garde S, Tessier PM. Arginine mutations in antibody complementarity-determining regions display context-dependent affinity/specificity trade-offs. J Biol Chem. 2017;292:16638–52. doi: 10.1074/jbc.M117.783837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Dashivets T, Stracke J, Dengl S, Knaupp A, Pollmann J, Buchner J, Schlothauer T. Oxidation in the complementarity-determining regions differentially influences the properties of therapeutic antibodies. MAbs. 2016;8:1525–35. doi: 10.1080/19420862.2016.1231277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lamberth K, Reedtz-Runge SL, Simon J, Klementyeva K, Pandey GS, Padkjaer SB, Pascal V, Leon IR, Gudme CN, Buus S, et al. Post hoc assessment of the immunogenicity of bioengineered factor VIIa demonstrates the use of preclinical tools. Sci Transl Med. 2017;9:1–11. eaag1286. doi: 10.1126/scitranslmed.aag1286. [DOI] [PubMed] [Google Scholar]
- 38.Harding FA, Stickler MM, Razo J, DuBridge R. The immunogenicity of humanized and fully human antibodies. mAbs. 2014;2:256–65. doi: 10.4161/mabs.2.3.11641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sekiguchi N, Kubo C, Takahashi A, Muraoka K, Takeiri A, Ito S, Yano M, Mimoto F, Maeda A, Iwayanagi Y, et al. MHC-associated peptide proteomics enabling highly sensitive detection of immunogenic sequences for the development of therapeutic antibodies with low immunogenicity. MAbs. 2018;10:1168–81. doi: 10.1080/19420862.2018.1518888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kabat EA, Wu TT, Perry HM, Gottesman KS, Foeller C. Sequences of proteins of immunological interest. Bethesda (MD): NIH; 1991. [Google Scholar]
- 41.Edelman GM, Cunningham BA, Gall WE, Gottlieb PD, Rutishauser U, Waxdal MJ. The covalent structure of an entire gammaG immunoglobulin molecule. Proc Natl Acad Sci U S A. 1969;63:78–85. doi: 10.1073/pnas.63.1.78. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.