

Introduction
The majority of acute cardiovascular events (CVE) in patients are caused by occlusive thrombosis because of rupture or erosion of atherosclerotic plaques.1 Growth differentiation factor 15 (GDF‐15), a stress‐responsive member of the transforming growth factor‐β (TGF‐β) cytokine superfamily, has been shown to be a strong and independent predictor of mortality and disease progression in patients with atherosclerosis and coronary artery disease (CAD), such as acute coronary syndromes (ACS) and stable angina pectoris.2 The development of atherosclerosis is dependent upon a high‐inflammatory content, which has been shown to modulate lesion initiation, progression, and potentially devastating thrombotic complications.3 Angiogenesis plays an important role in the progression of atherosclerotic plaque and complications.4, 5, 6 Atherosclerosis and cancer arise from multiple factors and are consolidated from the very early stages of development up to the advanced forms in inflammatory processes. Uncontrolled cell proliferation and oxidative stress and angiogenesis appear to be unifying causal factors in both diseases.7 A local inflammatory state occurring in atherosclerotic lesions has been implicated in angiogenesis through activation of endothelial cells, release of chemokines, cytokines, growth factors, lipid mediators, proteases, and increase of endothelial metabolic rate. The angiogenesis allows extravasation of the plasma component, leading to future thromboembolic events.8, 9, 10, 11 GDF‐15 might be an acute phase modifier of TGF‐βRII‐dependent proinflammatory responses to atherosclerotic plaque rupture and thrombus formation12 (Figure). Although the exact biological functions of GDF‐15 are still poorly understood, it has been shown to regulate inflammatory and angiogenesis pathways (Figure). GDF‐15 exhibits differing and even opposing functions under various circumstances. For instance, GDF‐15 has proapoptosis, antiapoptosis, proangiogenesis, antiangiogenesis, proinflammatory, and anti‐inflammatory properties.12, 13 Therefore, GDF‐15 exhibits a complex pattern with beneficial and harmful functions. GDF‐15 promoter contains p53‐transcription factor binding sites that are required and sufficient for the induction of GDF‐15 expression.14 Activation of p53 is a fundamental cellular response to inflammation, oxidative stress, hypoxia, telomere erosion, and oncogene activation. The circulating levels of GDF‐15 reflect these acute and chronic inflammatory conditions linked with atherosclerosis and CAD.
Figure 1.
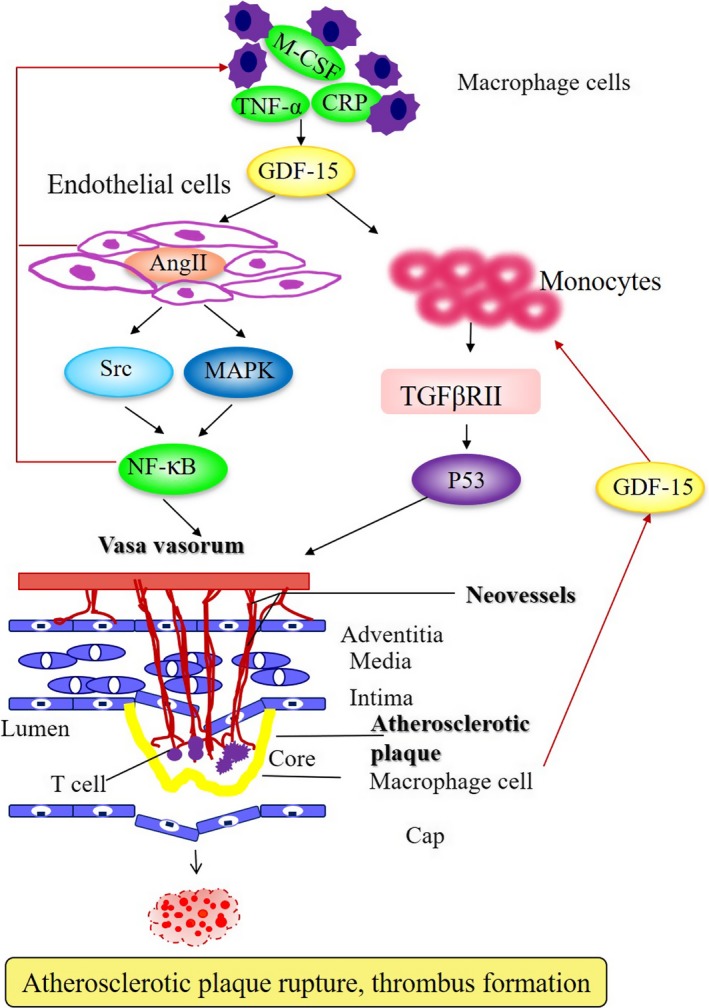
Schematic overview of a vulnerable plaque in advanced atherosclerosis. Plaque formation is initiated by endothelial cell dysfunction and subsequent angiogenesis and release of proinflammatory factors mediated by GDF‐15, contributing to the progression of atherosclerotic lesions and the development of plaque rupture and thrombus formation in atherosclerotic status. CRP indicates C‐reactive protein; GDF‐15, growth differentiation factor 15; MAPK, mitogen‐activated protein kinase; M‐CSF, macrophage colony‐stimulating factor; TGF‐βRII, transforming growth factor‐βRII; TNF‐α, tumor necrosis factor‐α.
Regulation and Roles of GDF‐15
Under normal physiological conditions, placenta is the only tissue expressing large quantities of GDF‐15.15 GDF‐15 levels are increased in various pathological conditions and diseases, including inflammation, cardiovascular disease, renal disease, pulmonary disease, and cancer.12 GDF‐15 is produced in activated macrophages,11 and in pathological conditions including proinflammatory status, vascular injury, pressure overload, and oxidative stress from human endothelial cells,16 vascular smooth muscle cells,17 and adipocytes.18 The expression of GDF‐15 in virtually all tissues suggests its importance in general and basic cellular functions. Although the exact biological functions of GDF‐15 remain largely unclear, it has been demonstrated to modulate inflammatory, apoptotic, and angiogenesis pathways.
GDF‐15 as a Novel Biomarker of CVE
GDF‐15 has been recognized as a consistent biomarker of CVE in patients with ACS or stable CAD.19 GDF‐15 levels are independently related to age, high‐sensitivity C‐reactive protein (hs‐CRP), natriuretic peptides, and renal dysfunction in patients with established CAD.2, 20, 21, 22, 23, 24, 25, 26 GDF‐15 concentrations are enhanced in patients with multivessel disease21, 27 and with a history of myocardial infarction (MI) or heart failure.27, 28, 29 The association of GDF‐15 with all‐cause mortality, cardiovascular mortality, MI, and stroke was further explored in our recently published research work, which included 3440 patients with established CAD independent of clinical predictors including age, diabetes mellitus, current smoking, hypertension, hyperlipidemia, and left ventricular ejection fraction.30 Our study simultaneously evaluated the incremental prognostic value of GDF‐15 and provided more information than other biomarkers (estimated glomerular filtration rate, fibrinogen, D‐dimer, sST2, pregnancy‐associated plasma protein A, and uric acid). Adding the information on GDF‐15 to the baseline clinical model improved the C‐index from 0.786 to 0.806. In addition, we examined whether there were heterogeneity in the hazard ratios based on presentation with stable CAD and ACS (unstable angina pectoris, non–ST‐segment–elevation myocardial infarction [NSTEMI], and ST‐segment–elevation myocardial infarction [STEMI]) beyond traditional risk factors. GDF‐15 was significantly associated with stable CAD and ACS. Recently, Gohar et al31 revealed that high circulating levels GDF‐15 are predictive of secondary CVE in women with carotid atherosclerosis, indicating contribution of high GDF‐15 levels to increased risk factors of CVE.
Roles of GDF‐15 in ACS
GDF‐15 is emerging as a prognostic biomarker in patients with ACS, including STEMI, NSTEMI, and unstable angina pectoris (Table 1), which result from the rupture or erosion of vulnerable atherosclerotic plaque leading to death and recurrent MI, which would be occurring at any time after the first attack episode.32, 33, 34, 35, 36, 37, 38, 39, 40, 41 The predictive value of GDF‐15 has been confirmed in the 2 large non–ST‐segment–elevation ACS (NSTE‐ACS) trials: the GUSTO‐IV (Global Utilisation of Strategies to Open Occluded Arteries IV) and FRISC II (Fast Revascularization during Instability in Coronary Artery Disease II) cohorts20, 21 (Table 1). As shown in patients from the GUSTO‐IV trial, GDF‐15 concentrations are closely related to all‐cause mortality in NSTE‐ACS20 (Table 1). In FRISC II, cumulative 1‐year mortality rates were 1.5, 5.0, and 14.1% in patients with low, moderately increased, and markedly increased concentrations of GDF‐15. GDF‐15 provided prognostic information beyond clinical predictors and other prognostic biomarkers, including cardiac troponin T, N‐terminal pro‐brain natriuretic peptide, hs‐CRP, and creatinine clearance.21 The independent association of GDF‐15 with mortality is confirmed in other patients with STEMI or NSTE‐ACS.22, 23 Lately, the prognostic value of GDF‐15 has been reevaluated in 16 876 patients with NSTE‐ACS or STEMI randomized to ticagrelor or clopidogrel in the PLATO (Platelet Inhibition and Patient Outcomes) study27(Table 1). Based on the large number of patients and outcome events, the PLATO biomarker study was able to explore the relation of GDF‐15 to specific outcome events during follow‐up. After adjustment for clinical predictors and other biomarkers, higher GDF‐15 concentrations were associated with an increased risk of all‐cause mortality, cardiovascular mortality, MI, and stroke. The results are confirmed by a secondary analysis of the PLATO study including 17 095 patients with ACS,42 demonstrating that GDF‐15 was a strong marker associated with all‐cause death, death caused by other vascular or nonvascular causes, and death caused by bleeding (Table 1). For GDF‐15, the possible signal of association with death caused by bleeding is in line with prior results indicating that GDF‐15 reflects nonoverlapping disease pathway contributing to the development of bleeding after ACS. Increased concentrations of GDF‐15 also identify patients at increased risk for adverse left ventricular remodeling and hospitalization for heart failure after ACS.23, 28, 43 In 3501 patients from the PROVE IT–TIMI‐22 (Pravastatin or Atorvastatin Evaluation and Infection Therapy‐Thrombolysis in Myocardial Infarction‐22) trial, GDF‐15 was associated with the risks of all‐cause mortality, recurrent MI, and hospitalization for new or worsening heart failure.28 The prognostic information provided by GDF‐15 was independent of clinical predictors and other biomarkers (hs‐CRP and brain natriuretic peptide). Notably, GDF‐15, in contrast to hs‐CRP,44 did not decline over time in response to more intensive statin therapy in PROVE IT–TIMI‐22,28 further indicating that GDF‐15 reflects a nonoverlapping atherosclerotic pathway contributing to the development of ACS.
Table 1.
GDF‐15 Related to Outcome Events in ACS
| Study | Participants | Outcomes | Follow‐Up (y) | Comparisons (ng/L) | RR (95% CI) |
|---|---|---|---|---|---|
| CAD patients, Kempf et al2 | ACS (n =877) | M | 6 (maximum) | <1200, 1200 to 1800, >1800 | 8.5 (3.81–18.99) |
| GUSTO‐IV, Wollert et al20 | NSTE‐ACS (n=2081) | M | 1 (maximum) | <1200, 1200 to 1800, >1800 | 2.08 (1.85–2.34) |
| FRISC‐II, Wollert et al21 | NSTE‐ACS (n=2079) | M, R | 2 (maximum) | <1200, 1200 to 1800, >1800 | 1.75 (1.48–2.07) |
| ASSENT‐2 and ASSENT‐plus trials, Kempf et al22 | STEMI (n =741) | M | 1 (maximum) | <1200, 1200 to 1800, >1800 | 6.6 (2.43–18.23) |
| AMI patients, Khan et al23 | AMI (n=1142) | M, HF | 1.4 (median) | 1470 (240–31 860) | 4.24 (3.21–5.62) |
| FRISC II, Eggers et al24 | NSTE‐ACS (n=950) | M, R | 0.5 (maximum) | <1200, 1200 to 1800, >1800 | 1.9 (1.2–3.0) |
| PLATO, Hagstrom et al27 | ACS (n=16 876) | M | 1 (maximum) | <1145, 1145 to 1550, 1550 to 2219, >2219 | 3.96 (2.91–5.39) |
| PROVE IT‐TIMI 22, Bonaca et al28 | ACS (n=3501) | M | 2 (maximum) | <1200, 1200 to 1800, >1800 | 4.76 (2.67–8.48) |
| STEMI patients, Eitel I et al32 | STEMI (n=238) | M,R | 0.5 (maximum) | <1319, ≥1319 | 19 (2.58, 139.66) |
| NSTE‐ACS patients, Widera et al33 | NSTE‐ACS (n=1122) | M, R | 0.5 (mean) | 1725 (1205–2797) | 2.4 (1.9–3.0) |
| NSTE‐ACS patients, Widera et al34 | NSTE‐ACS (n=1146) | M, R | 0.5 (mean) | 1770 (1262–2981) | 2.4 (2.0–3.0) |
| ICTUS, Damman et al35 | NSTE‐ACS | M | 5 (maximum) | <1200, 1200 to 1800, >1800 | 4.78 (3.71–6.18) |
| PLATO, Wallentin et al36 | NSTE‐ACS (n=9946) | M,R,S | 1 (maximum) | <1200, 1200 to 1800, >1800 | NA |
| NSTE‐ACS patients, Dominguez‐Rodriguez et al37 | NSTE‐ACS (n=255) | M,R,UA | 3 (maximum) | 1639 (median) | 52.3 (7–388.5) |
| Shock II, Fuernau et al38 | AMI (n=190) | M | 0.1 (maximum) | 7662 (median) | 1.88 (1.21–2.94) |
| FRISC‐II, Wallentin et al39 | NSTE‐ACS (n=2457) | M,R | 2 (maximum) | <1800, ≥1800 | NA |
| NSTE‐ACS patients, Dominguez‐Rodriguez et al40 | NSTE‐ACS (n=502) | M,R,UA | 2 (maximum) | 470 to 1765, 1766 to 2995, 2996 to 11 607 | 6.6 (4.28–10.2) |
| Västmanland Myocardial Infarction Study, Skau et al41 | AMI (n=847) | M | 6.9 (median) | NA | 2.57 (2.31–2.85) |
| PLATO, Lindholm et al42 | ACS (n=17 095) | M | 1 (maximum) | NA | 2.65 (2.17–3.24) |
ACS indicates acute coronary syndrome; AMI, acute myocardial infarction; ASSENT, assessment of the Safety and Efficacy of a New Thrombolytic; CAD, coronary artery disease; FRISC II, Fast Revascularization during Instability in Coronary artery disease II; GDF‐15, growth differentiation factor 15; GUSTO‐IV, Global Utilisation of Strategies to Open Occluded Arteries IV; HF, heart failure; ICTUS, Invasive versus Conservative Treatment in Unstable coronary Syndromes; M, mortality; NA, not applicable; NSTE‐ACS, non‐ST‐segment–elevation acute coronary syndrome; PLATO, Platelet Inhibition and Patient Outcomes; PROVE IT‐TIMI‐22, Pravastatin or Atorvastatin Evaluation and Infection Therapy‐Thrombolysis in Myocardial Infarction‐22 trial; R, recurrent myocardial infarction; RR, relative risk; S, stroke; STEMI, ST‐segment–elevation myocardial infarction; UA, unstable angina.
Roles of GDF‐15 in Stable CAD
GDF‐15 maintains its close association with an adverse prognosis in patients with ACS during the transition to the chronic stage of CAD.24, 25 In a serial analysis from FRISC‐II, GDF‐15 provided similar independent prognostic information on the composite end point of death or recurrent MI on admission and up to 6 months after an episode of NSTE‐ACS.24 Similarly, GDF‐15 was identified as an independent predictor of CAD mortality in patients with stable CAD2 (Table 2). In the AtheroGene (patients with stable CAD or ACS who had at least 1 stenosis >30% in a major coronary artery were enrolled in the AtheroGene registry) study, which included 1352 patients with stable angina pectoris undergoing coronary angiography, GDF‐15 was associated with CAD mortality independent of cardiovascular risk factors, clinical predictors, the number of diseased vessels, left ventricular ejection fraction, and other biomarkers (cTnI, N‐terminal pro‐brain natriuretic peptide, and hs‐CRP).2 Similarly, in a cohort of 984 patients with stable CAD, higher GDF‐15 levels were associated with lower left ventricular ejection fraction, worse diastolic function, and greater inducible ischemia. The association of GDF‐15 with MI, heart failure, and cardiovascular death persisted after extensive adjustment for traditional risk factors and the other biomarkers (NT‐proBNP, CRP, and hs‐cardiac troponin T)29 (Table 2). Recently, the prognostic value of GDF‐15 has been reevaluated in 14 577 patients with stable CAD in specific outcome events from STABILITY (The Stabilization of Atherosclerotic Plaque by Initiation of Darapladib Therapy) study 26 (Table 2). Our recent study further validated that GDF‐15 is associated with cardiovascular and noncardiovascular death (eg, cancer morbidity) in stable CAD patients with and without previous cancer diagnosis.30 Furthermore, our study also indicated the independent associations between the GDF‐15 and coronary thrombotic events (eg, MI), even after adjusting for other prognostic biomarkers (estimated glomerular filtration rate and left ventricular ejection fraction).
Table 2.
GDF‐15 related to outcome events in stable CAD
| Study | Participants | Outcomes | Follow‐Up (y) | Comparisons (ng/L) | RR (95% CI) |
|---|---|---|---|---|---|
| Kempf et al2 | Stable CAD (n=1352) | M | 3.6 (median) | 1128 (850–1553) | 2.7 (2.2–3.3) |
| Dallmeier et al25 | Stable CAD (n=1029) | M | 10 (median) | 1232 (916–1674) | 2.80 (1.98–3.37) |
| Hagstrom et al26 | Stable CAD (n=14 577) | M | 3.7 (median) | 1253 (915–1827) | 2.63 (1.91–3.63) |
| Schopfer et al29 | Stable CAD (n=948) | M | 8.9 (mean) | 2166 (1589–3057) | 2.97 (2.58–3.43) |
CAD indicates coronary artery disease; GDF‐15, growth differentiation factor 15; M, mortality; RR, relative risk.
GDF‐15 is a biomarker considered for introduction to the clinic. What questions remain to be answered to establish GDF‐15 as a clinically useful biomarker? Moreover, is GDF‐15 a risk biomarker or a causative risk factor, or more importantly, what are the circumstances under which GDF‐15 is just a marker of risk versus a causative factor? Its function as a protective or disease‐inducing factor remains largely unknown. The GDF‐15 puzzle is a good example of how epidemiological and mechanistic studies can interact successfully. The predictive value persists even a decade later, and the findings discussed above support the hypothesis that GDF‐15 is not a consequence of cardiovascular disease or a passive biomarker of the disease process, but in fact plays an active role in the pathophysiology of atherosclerosis and CAD.45, 46 The clinical significance of newly discovered mechanisms can be evaluated and conversely, the mechanisms behind epidemiologically proven associations can be elucidated.
GDF‐15 and Inflammation in Atherosclerosis
Potential mechanisms have been suggested for the association of GDF‐15 with adverse outcomes in atherosclerosis, including worse baseline cardiac disease severity, inflammation, ischemia, volume overload, and adipokines.12 Elevated GDF‐15 has been shown to promote inflammation and angiogenesis,47, 48, 49 implying that GDF‐15 may play an important role in the pathogenesis of atherosclerosis. While GDF‐15 is a cardiovascular risk factor, whether GDF‐15 contributes directly to atherosclerosis development has not been established and the precise relationships between GDF‐15 and atherosclerosis are incompletely understood. GDF‐15 was originally identified as a factor overexpressed in activated macrophages to regulate inflammation, which is involved in all stages of atherosclerosis, from its initiation and progression to its thrombotic complications. de Jager et al49 demonstrate that leukocyte deficiency of GDF‐15 improves atherosclerotic plaque stability by impairing macrophage migration and promoting collagen deposition. GDF‐15 deficiency in leukocytes is associated with reduced macrophage accumulation in an atherosclerosis model, suggesting a pro‐inflammatory role of GDF‐15 in atherosclerosis. Moreover, chromatin immunoprecipitation assays confirmed that p53 was recruited to both p53 binding sites 1 and 2 in the GDF‐15 promoter in response to CRP.50 Accordingly, CRP induces GDF‐15 expression through the regulation of p53 binding sites in the GDF‐15 promoter. Along this line, GDF‐15 is involved in orchestrating atherosclerotic lesion progression by regulating apoptotic cell death and IL‐6‐dependent inflammatory responses to vascular injury.51 These data suggest an involvement of GDF‐15 in the initiation and progression of atherosclerosis. GDF‐15 revealed a central role for this factor as a pro‐inflammatory cytokine that accelerates atherosclerosis.
GDF‐15 is in fact associated with subclinical atherosclerosis.52 GDF‐15 deficiency resulted in a reduction of early atherosclerotic lesion size after 4 weeks on a high cholesterol Western‐type diet. After 12 weeks, no differences in lesion size could be detected.49 It is known that lesions in mice become quite complex with increased duration of feeding.53 Moreover, GDF‐15 expression is significantly higher in acute stages of human plaque rupture (unstable angina pectoris) than in advanced stable lesions (stable angina pectoris). Paradoxically, overexpression of GDF‐15 in macrophages significantly attenuates atherosclerotic lesions in the ApoE−/− mouse model of atherosclerosis.54 GDF‐15 is thought to have anti‐inflammatory effects on cells, including cardiomyocytes.55 Preusch et al demonstrated a proinflammatory plaque phenotype in mice transplanted with bone marrow from GDF‐15−/− donors with enhanced macrophage accumulation, suggesting a protective effect of GDF‐15 on the atherosclerosis process.56 However, this effect may contribute to changes in lesion vulnerability such as thinning of fibrous caps and potential plaque rupture. It should, however, be noted that they did not focus on the onset of atherosclerotic changes within the vascular wall such as lipid accumulation in younger mice. It is known as a model of late‐stage disease in atherosclerosis and does not show much progress in early stages. To further elaborate on this, de Jager et al investigated the signal transduction cascades for GDF‐15. Blockade of TGFβRII, but not TGFβRI/ALK5, abrogated the GDF‐15‐elicited MCP‐1 response, suggesting the role of GDF‐15 in the underlying mechanism of atherosclerosis progress. Thus, GDF‐15 has a pleiotropic regulatory effect on the inflammatory process, in line with that of other TGF‐β family members such as activin‐A57 and TGF‐β1.58 Previous study pointed out that expression of GDF‐15 may be upregulated by a variety of proinflammatory stimuli in macrophages including interleukin (IL)‐1β, IL‐2, and tumor necrosis factor‐α.11 Recent study found a positive association between the IL‐1β and CVE,59 suggesting there is an interleukin‐1β/GDF‐15‐associated immunity pathway resulting in atherosclerosis. Accordingly, the high levels of GDF‐15 may result from high levels of monokines such as IL‐1β, tumor necrosis factor‐α, and CRP. GDF‐15 initiates pro‐ and anti‐inflammatory effects on atherosclerosis development and progression, depending on the pathophysiological context and progression stage. GDF‐15 functions as a proinflammatory factor in the process of atherosclerosis via TGFβRII signaling, especially in the early stage and acute inflammatory stage, leading to vulnerable plaque, which provides 1 of the possible mechanisms for the atherosclerosis process.
GDF‐15 and Angiogenesis
Plaques that are most at risk are characterized by large necrotic cores with a thin fibrous cap. Plaque angiogenesis and intraplaque hemorrhage are important contributors to unstable lesions.6, 60 Although commonly regarded as separate disease entities, there is a growing recognition that cardiovascular disease and cancer have various similarities with shared common biology and risk factors, including age, diabetes mellitus, hypertension, smoking, physical inactivity, and unhealthy diet. A novel function for GDF‐15 was identified as a potent angiogenic factor to be secreted from melanoma cells together with vascular endothelial growth factor to promote vascular development.61 During angiogenesis, endothelial cells emerge from the quiescent state and undergo progression in the cell cycle. GDF‐15 is causally involved in the pathological process of endothelial proliferation and angiogenesis.9, 62 Jin et al revealed the functional effect of GDF‐15 on the cell cycle progression of endothelial cells and demonstrated that GDF‐15 upregulates expression of cyclins D1 and E in human umbilical vein endothelial cells, leading to a rapid transition from G1 to S phase.9 GDF‐15 has been shown to promote cell viability, invasion, migration, and angiogenesis in HepG2 cells48 and hypoxic human umbilical vein endothelial cells possibly through inhibiting p53 signaling.62 Moreover, GDF‐15 induced the pro‐angiogenic effects through the phosphorylation of Src and its downstream pathways of AKT, MAPK, and NF‐κB signaling, implying regulatory roles of GDF‐15 in cell proliferation and angiogenesis in atherosclerosis.47 Intriguingly, proinflammatory factors such as IL‐1β, tumor necrosis factor‐α, and CRP induce GDF‐15 expression in macrophage cells through the regulation of p53 binding sites in the GDF‐15 promoter (Figure). GDF‐15 promoted macrophage chemotaxis in a strictly CCR2‐ and TGF‐β type II receptors (TGFβRII)–dependent manner in early and advanced atherosclerosis. Accordingly, the GDF‐15/TGFβRII/P53 and the GDF‐15/NF‐κB pathways are the critical mechanisms involved in the angiogenesis and acute inflammation in the unstable atherosclerotic plaque. The functional proatherogenic roles of GDF‐15 in lesion progression indicate that besides other TGF‐β superfamily members such as TGF‐β1 and BMP, interference with GDF‐15 may be a useful novel strategy for therapeutic intervention.63, 64
GDF‐15 and Stress in Atherosclerosis
GDF‐15 and brain natriuretic peptide are similarly induced by biomechanical stress in isolated rat cardiomyocytes and in the murine heart. GDF‐15 is upregulated in response to stressors including in macrophages exposed to oxidized low‐density lipoprotein in atherosclerotic carotid arteries.65 Specific to atherosclerosis, GDF‐15 has shown predictive abilities of CAD mortality and composite outcomes in stable CAD and ACS in patients with prevalent cardiovascular risk factors.2, 21, 24, 28 Recent findings support that GDF‐15 is associated with subclinical atherosclerosis as assessed by maximal internal carotid artery intima‐media thickness as well as the presence of carotid plaque. Whether GDF‐15 is a mediator of cardiovascular disease or upregulated in response to cardiovascular injury remains unclear. After further adjusting CRP and brain natriuretic peptide, the association of GDF‐15 with maximum internal carotid artery intima‐media thickness and carotid plaque was more robust. This suggests that GDF‐15 may reflect an orthogonal pathway associated with cardiovascular disease, the mechanism of which remains unclear.
Roles of GDF‐15 in Cancer and Other Diseases
GDF‐15 is characterized by a wide tissue distribution pattern with high expression in the prostate and placenta, heart, intestine, liver, kidney, pancreas, colon, lung, brain, and skeletal muscle.66 It acts as a multifunctional cytokine by controlling numerous physiological and pathological processes. Acting on the hypothalamus and hindbrain, GDF‐15 is a key inducer of cancer‐related anorexia and weight loss.67 Moreover, GDF‐15 plays an important role in the physiological regulation of energy intake and expenditure, with a more pronounced effect in women than in men.68 Although several studies suggest antitumoral activity, the protumoral effects of GDF‐15 appear to prevail.12, 47, 69
Like the other members of the TGFβ‐superfamily, GDF‐15 has opposite effects depending on cellular context, disease stage, or microenvironment. GDF‐15 has both antitumorigenic and protumoral properties. In fact, these apparently paradoxical data could be explained by a dual role of GDF‐15 in cancer progression: inhibition of carcinogenesis in normal tissue at early stages of tumor development and promotion of tumor at late stages of the disease.70 GDF‐15 induces pleiotropic effects in cancer by modulating cancer cell proliferation and chemoprotection but also the tumoral microenvironment (angiogenesis, invasion and metastasis processes, and immunomodulation), as well as more unexpected processes (cancer‐induced anorexia). GDF‐15 has been implicated in chronic disease, such as rheumatoid arthritis, end‐stage renal failure, or diabetes mellitus.71, 72, 73 As for cancer or cardiovascular diseases, GDF‐15 plasma concentration was an independent predictor of disease worsening and/or death. The biological processes that could explain such a link are obscure and often not known. A recent study emphasizes the positive effects of GDF‐15 on peripheral nerve regeneration.74 In this case, GDF‐15 seems to reduce the number of regenerated axons but it increases the maturation of newly formed ones. This leads to better recovery of sensorimotor function.
Potential Implications of GDF‐15 in Atherosclerosis
GDF‐15 functions as a direct participant in the atherosclerotic process. Plaque angiogenesis is a physiological response to the increased oxygen demand in the plaque but has adverse effects by facilitating intraplaque hemorrhage and influx of inflammatory mediators. The angiogenesis inhibitor angiostatin reduces plaque angiogenesis, and the secondary reduction of macrophages may have beneficial effects on plaque stability.75 GDF‐15 deficiency contributes to angiogenesis and improves atherosclerotic plaque stability by impairing macrophage migration and promoting collagen deposition. A high level of serum GDF‐15 is detected in human atherosclerotic lesions,49, 65 which are broadly proportional to the disease burden. Thus, we speculate that GDF‐15 is located in arteriosclerotic lesions or in circulation and promotes atherosclerotic plaque vulnerability by increasing angiogenesis and inflammation. Intriguingly, GDF‐15 promotes indirect proinflammatory effects in atherosclerosis49, 51 but mediates anti‐inflammatory effects in acute MI by directly inhibiting myeloid cell recruitment.76 Although it is possible that GDF‐15 itself could be causative in the development of ACS,54 GDF‐15 has anti‐apoptotic and antihypertrophic properties of in cardiomyocytes subjected to simulated ischemia/reperfusion injury.77 Notably, the biological effects of GDF‐15 are context dependent and may vary with the stage of the disease.34, 36, 55, 56, 76, 78, 79, 80 In line with these investigations, GDF‐15 has been shown to be associated with subclinical atherosclerosis involved in macrophage accumulation in atherosclerosis.52 Correspondingly, GDF‐15 is responsible for early‐stage atherosclerotic lesions. Inflammatory factors (IL‐1β or CRP) secreted from macrophages induce GDF‐15 expression through the regulation of p53 binding sites in the GDF‐15 promoter further activates its downstream NF‐κB signaling, accelerating the progression of atherosclerosis in the early stage, and promoting the formation of vulnerable plaque. GDF‐15 is also linked with endothelial dysfunction and more advanced coronary atherosclerosis, suggesting the regulatory roles of GDF‐15 in chronic myocardial and vascular damage in the late stage of the atherosclerosis process.19 More basic research into the pathobiological features of GDF‐15 is needed to explore the mechanism related to the risk of new atherosclerosis and recurrent ischemic events after ACS.
Potential Implications for GDF‐15 in ACS and CAD
GDF‐15 appears to be a very consistent marker of adverse long‐term outcome in ACS. However, may GDF‐15 be used to identify groups of patients who will or will not benefit from various interventions or treatments? Are there any treatments for which monitoring of GDF‐15 concentrations might be useful to guide the treatment (dose and/or duration)? Several studies illustrate the potential of the marker to risk stratify unselected contemporary patient populations treated outside clinical trials. In a recent investigation that compared the incremental prognostic value of 9 biomarkers on top of the GRACE (Global Registry of Acute Coronary Events) score in unselected patients with NSTE‐ACS, GDF‐15 emerged as the most promising biomarker.34 Underscoring its potential to add information to what is clinically available, GDF‐15 also added discriminatory information to GRACE when hs‐cardiac troponin T was considered as an additional continuous variable. In accordance with previous observations, we noted in our population‐based cohort that addition of GDF‐15 to standard cardiovascular risk factors resulted in modest but significant improvements in the C‐statistic (discrimination) as well as reclassification, as measured by the integrated discrimination improvement and net reclassification improvement for all‐cause death and cardiovascular death.30 In addition, GDF‐15 predicted all‐cause mortality more accurately independently of hs‐cardiac troponin T and N‐terminal pro‐brain natriuretic peptide in patients with acute pain.81 The PLATO trial showed that GDF‐15 contributes information on the magnitude of benefit by a successful intervention such as ticagrelor regardless of invasive or noninvasive management.36 The FRISC‐II trial supported that a high concentration of the biomarker GDF‐15 implies heightened risk and functions as a useful identification of patients who might expect the longest postponement of death or MI with an early invasive strategy.39 Thresholds offer a convenient way to classify patients into risk categories that may be linked to treatment decisions. However, the use of thresholds may reduce statistical power given the continuous association of GDF‐15 with cardiovascular risk. Alternatively, GDF‐15 might be incorporated as a continuous variable into established or novel risk scores that can be presented as nomograms or applications on (handheld) electronic devices. Therefore, in a setting with a need for prioritization among different patients with NSTE‐ACS for early invasive procedures, direct access to treatment for patients with elevated troponin, in addition to a fast track for those with high GDF‐15, might be a useful strategy. New algorithms for decision support in ACS are currently under evaluation, including variables such as troponin and GDF‐15 showing significant interactions with the effects of an early invasive treatment strategy. More importantly, there are medical therapies that reduce the risk of cardiovascular disease and cancer. For example, use of daily aspirin for the primary prevention of major CVE reduces the incidence of cancer and cancer mortality, although more research is required to identify which individuals are likely to benefit most.82 In previous studies of patients with NSTE‐ACS, GDF‐15 has been found to predict future events and contribute to the identification of high‐risk patients with a benefit of an early invasive strategy. Thus, GDF‐15 might be of clinical value in refining risk stratification and tailoring treatment of patients with ACS. GDF‐15 provides unique information on underlying disease processes leading to a raised risk of severe events, eg, fatal CVE and death.27 Moreover, the previous studies published on the functional proatherogenic role of GDF‐15 in lesion progression indicate that besides other TGF‐β superfamily members such as TGF‐β1 and BMP,83 interference with GDF‐15 may be a useful novel strategy for therapeutic intervention.48, 51 Therefore, circulating levels of GDF‐15 are suggested as a prognostic marker to improve risk stratification of patients with ACS, along with benefit from treatment with a high‐dose, highly efficient statin. Increased GDF‐15 plasma concentrations at the time of PCI and stent implantation might classify high‐risk patients with ACS who benefit from high‐dose, highly efficient statins, implicating that high‐dose statins are more effective in high‐risk patients and obtaining GDF‐15 may help identify these patients.45, 46
Conclusions and Future Directions
GDF‐15 functions as a cardiovascular risk and outcome marker and appears to be a direct participant in the atherosclerotic process. GDF‐15 is responsible for vulnerable atherosclerotic lesions by proinflammation and angiogenesis, accelerating the progression of atherosclerosis especially in the early stage, subsequently contributing to promotion of the vulnerable plaque formation. Moreover, GDF‐15 has been found to predict CVE and identify high‐risk patients with a benefit of an early invasive strategy. The importance of the GDF‐15/TGFβRII/P53 and the GDF‐15/NF‐κB signaling pathways in the cardiovascular system sparked hopes that manipulating its pathophysiological activity could provide novel therapeutic agents for atherosclerosis and CAD. In fact, the clinical and experimental studies clearly support a physiological and pathophysiological role for the GDF‐15 system in atherosclerosis and CAD. Targeting the GDF‐15 pathway represents a novel therapeutic approach against atherosclerosis and CAD that will increase our understanding of the pathophysiology of these diseases.
Sources of Funding
This research program was supported by the National Major Research Plan Training Program of China (91849111) and the National Natural Science Foundation of China (81770253; 81670214; 81370362), Natural Cultivation Foundation of the Capital Medical University (PYZ2018106) and Talent project of Beijing Chaoyang Hospital Affiliated to Capital Medical University.
Disclosures
None.
J Am Heart Assoc. 2019;8:e012826 DOI: 10.1161/JAHA.119.012826.
Contributor Information
Xinchun Yang, Email: yxccyxn@163.com.
Jiuchang Zhong, Email: jiuchangzhong@aliyun.com.
References
- 1. Bentzon JF, Otsuka F, Virmani R, Falk E. Mechanisms of plaque formation and rupture. Circ Res. 2014;114:1852–1866. [DOI] [PubMed] [Google Scholar]
- 2. Kempf T, Sinning JM, Quint A, Bickel C, Sinning C, Wild PS, Schnabel R, Lubos E, Rupprecht HJ, Munzel T, Drexler H, Blankenberg S, Wollert KC. Growth‐differentiation factor‐15 for risk stratification in patients with stable and unstable coronary heart disease: results from the atherogene study. Circ Cardiovasc Genet. 2009;2:286–292. [DOI] [PubMed] [Google Scholar]
- 3. Couchie D, Vaisman B, Abderrazak A, Mahmood DFD, Hamza MM, Canesi F, Diderot V, El Hadri K, Negre‐Salvayre A, Le Page A, Fulop T, Remaley AT, Rouis M. Human plasma thioredoxin‐80 increases with age and in apoE(‐/‐) mice induces inflammation, angiogenesis, and atherosclerosis. Circulation. 2017;136:464–475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Maracle CX, Agca R, Helder B, Meeuwsen JAL, Niessen HWM, Biessen EAL, de Winther MPJ, de Jager SCA, Nurmohamed MT, Tas SW. Noncanonical NF‐kappaB signaling in microvessels of atherosclerotic lesions is associated with inflammation, atheromatous plaque morphology and myocardial infarction. Atherosclerosis. 2018;270:33–41. [DOI] [PubMed] [Google Scholar]
- 5. Mujaj B, Bos D, Muka T, Lugt AV, Ikram MA, Vernooij MW, Stricker BH, Franco OH. Antithrombotic treatment is associated with intraplaque haemorrhage in the atherosclerotic carotid artery: a cross‐sectional analysis of the rotterdam study. Eur Heart J. 2018;39:3369–3376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Wang J, Liu H, Sun J, Xue H, Xie L, Yu S, Liang C, Han X, Guan Z, Wei L, Yuan C, Zhao X, Chen H. Varying correlation between 18F‐fluorodeoxyglucose positron emission tomography and dynamic contrast‐enhanced MRI in carotid atherosclerosis: Implications for plaque inflammation. Stroke. 2014;45:1842–1845. [DOI] [PubMed] [Google Scholar]
- 7. Koene RJ, Prizment AE, Blaes A, Konety SH. Shared risk factors in cardiovascular disease and cancer. Circulation. 2016;133:1104–1114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. de Vries MR, Quax PH. Plaque angiogenesis and its relation to inflammation and atherosclerotic plaque destabilization. Curr Opin Lipidol. 2016;27:499–506. [DOI] [PubMed] [Google Scholar]
- 9. Jin YJ, Lee JH, Kim YM, Oh GT, Lee H. Macrophage inhibitory cytokine‐1 stimulates proliferation of human umbilical vein endothelial cells by up‐regulating cyclins d1 and e through the PI3k/Akt‐, ERK‐, and JNK‐dependent AP‐1 and E2F activation signaling pathways. Cell Signal. 2012;24:1485–1495. [DOI] [PubMed] [Google Scholar]
- 10. Raggi P, Genest J, Giles JT, Rayner KJ, Dwivedi G, Beanlands RS, Gupta M. Role of inflammation in the pathogenesis of atherosclerosis and therapeutic interventions. Atherosclerosis. 2018;276:98–108. [DOI] [PubMed] [Google Scholar]
- 11. Bootcov MR, Bauskin AR, Valenzuela SM, Moore AG, Bansal M, He XY, Zhang HP, Donnellan M, Mahler S, Pryor K, Walsh BJ, Nicholson RC, Fairlie WD, Por SB, Robbins JM, Breit SN. MIC‐1, a novel macrophage inhibitory cytokine, is a divergent member of the TGF‐beta superfamily. Proc Natl Acad Sci USA. 1997;94:11514–11519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Unsicker K, Spittau B, Krieglstein K. The multiple facets of the TGF‐β family cytokine growth/differentiation factor‐15/macrophage inhibitory cytokine‐1. Cytokine Growth Factor Rev. 2013;24:373–384. [DOI] [PubMed] [Google Scholar]
- 13. Xu X, Li Z, Gao W. Growth differentiation factor 15 in cardiovascular diseases: from bench to bedside. Biomarkers. 2011;16:466–475. [DOI] [PubMed] [Google Scholar]
- 14. Yang H, Filipovic Z, Brown D, Breit SN, Vassilev LT. Macrophage inhibitory cytokine‐1: a novel biomarker for p53 pathway activation. Mol Cancer Ther. 2003;2:1023–1029. [PubMed] [Google Scholar]
- 15. Fairlie WD, Moore AG, Bauskin AR, Russell PK, Zhang HP, Breit SN. MIC‐1 is a novel TGF‐beta superfamily cytokine associated with macrophage activation. J Leukoc Biol. 1999;65:2–5. [DOI] [PubMed] [Google Scholar]
- 16. Secchiero P, Corallini F, Gonelli A, Dell'Eva R, Vitale M, Capitani S, Albini A, Zauli G. Antiangiogenic activity of the MDM2 antagonist nutlin‐3. Circ Res. 2007;100:61–69. [DOI] [PubMed] [Google Scholar]
- 17. Bermudez B, Lopez S, Pacheco YM, Villar J, Muriana FJ, Hoheisel JD, Bauer A, Abia R. Influence of postprandial triglyceride‐rich lipoproteins on lipid‐mediated gene expression in smooth muscle cells of the human coronary artery. Circ Res. 2008;79:294–303. [DOI] [PubMed] [Google Scholar]
- 18. Ding Q, Mracek T, Gonzalez‐Muniesa P, Kos K, Wilding J, Trayhurn P, Bing C. Identification of macrophage inhibitory cytokine‐1 in adipose tissue and its secretion as an adipokine by human adipocytes. Endocrinology. 2009;150:1688–1696. [DOI] [PubMed] [Google Scholar]
- 19. Farhan S, Freynhofer MK, Brozovic I, Bruno V, Vogel B, Tentzeris I, Baumgartner‐Parzer S, Huber K, Kautzky‐Willer A. Determinants of growth differentiation factor 15 in patients with stable and acute coronary artery disease. A prospective observational study. Cardiovasc Diabetol. 2016;15:60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wollert KC, Kempf T, Peter T, Olofsson S, James S, Johnston N, Lindahl B, Horn‐Wichmann R, Brabant G, Simoons ML, Armstrong PW, Califf RM, Drexler H, Wallentin L. Prognostic value of growth‐differentiation factor‐15 in patients with non‐ST‐elevation acute coronary syndrome. Circulation. 2007;115:962–971. [DOI] [PubMed] [Google Scholar]
- 21. Wollert KC, Kempf T, Lagerqvist B, Lindahl B, Olofsson S, Allhoff T, Peter T, Siegbahn A, Venge P, Drexler H, Wallentin L. Growth differentiation factor 15 for risk stratification and selection of an invasive treatment strategy in non ST‐elevation acute coronary syndrome. Circulation. 2007;116:1540–1548. [DOI] [PubMed] [Google Scholar]
- 22. Kempf T, Bjorklund E, Olofsson S, Lindahl B, Allhoff T, Peter T, Tongers J, Wollert KC, Wallentin L. Growth‐differentiation factor‐15 improves risk stratification in ST‐segment elevation myocardial infarction. Eur Heart J. 2007;28:2858–2865. [DOI] [PubMed] [Google Scholar]
- 23. Khan SQ, Ng K, Dhillon O, Kelly D, Quinn P, Squire IB, Davies JE, Ng LL. Growth differentiation factor‐15 as a prognostic marker in patients with acute myocardial infarction. Eur Heart J. 2009;30:1057–1065. [DOI] [PubMed] [Google Scholar]
- 24. Eggers KM, Kempf T, Lagerqvist B, Lindahl B, Olofsson S, Jantzen F, Peter T, Allhoff T, Siegbahn A, Venge P, Wollert KC, Wallentin L. Growth‐differentiation factor‐15 for long‐term risk prediction in patients stabilized after an episode of non‐ST‐segment‐elevation acute coronary syndrome. Circ Cardiovasc Genet. 2010;3:88–96. [DOI] [PubMed] [Google Scholar]
- 25. Dallmeier D, Brenner H, Mons U, Rottbauer W, Koenig W, Rothenbacher D. Growth differentiation factor 15, its 12‐month relative change, and risk of cardiovascular events and total mortality in patients with stable coronary heart disease: 10‐year follow‐up of the KAROLA study. Clin Chem. 2016;62:982–992. [DOI] [PubMed] [Google Scholar]
- 26. Hagstrom E, Held C, Stewart RA, Aylward PE, Budaj A, Cannon CP, Koenig W, Krug‐Gourley S, Mohler ER III, Steg PG, Tarka E, Ostlund O, White HD, Siegbahn A, Wallentin L. Growth differentiation factor 15 predicts all‐cause morbidity and mortality in stable coronary heart disease. Clin Chem. 2017;63:325–333. [DOI] [PubMed] [Google Scholar]
- 27. Hagstrom E, James SK, Bertilsson M, Becker RC, Himmelmann A, Husted S, Katus HA, Steg PG, Storey RF, Siegbahn A, Wallentin L. Growth differentiation factor‐15 level predicts major bleeding and cardiovascular events in patients with acute coronary syndromes: results from the PLATO study. Eur Heart J. 2016;37:1325–1333. [DOI] [PubMed] [Google Scholar]
- 28. Bonaca MP, Morrow DA, Braunwald E, Cannon CP, Jiang S, Breher S, Sabatine MS, Kempf T, Wallentin L, Wollert KC. Growth differentiation factor‐15 and risk of recurrent events in patients stabilized after acute coronary syndrome: observations from PROVE IT‐TIMI 22. Arterioscler Thromb Vasc Biol. 2011;31:203–210. [DOI] [PubMed] [Google Scholar]
- 29. Schopfer DW, Ku IA, Regan M, Whooley MA. Growth differentiation factor 15 and cardiovascular events in patients with stable ischemic heart disease (the heart and soul study). Am Heart J. 2014;167:186–192.e181. [DOI] [PubMed] [Google Scholar]
- 30. Wang J, Liu J, Guo W, Bai Y, Li H, Chen H, Han L, Lyu L, Xu C, Liu H. Multiple biomarkers in the context of conventional risk factors in patients with coronary artery disease. J Am Coll Cardiol. 2017;69:2769–2770. [DOI] [PubMed] [Google Scholar]
- 31. Gohar A, Goncalves I, Vrijenhoek J, Haitjema S, van Koeverden I, Nilsson J, de Borst GJ, de Vries JP, Pasterkamp G, den Ruijter HM, Bjorkbacka H, de Jager SCA. Circulating GDF‐15 levels predict future secondary manifestations of cardiovascular disease explicitly in women but not men with atherosclerosis. Int J Cardiol. 2017;241:430–436. [DOI] [PubMed] [Google Scholar]
- 32. Eitel I, Blase P, Adams V, Hildebrand L, Desch S, Schuler G, Thiele H. Growth‐differentiation factor 15 as predictor of mortality in acute reperfused ST‐elevation myocardial infarction: insights from cardiovascular magnetic resonance. Heart. 2011;97:632–640. [DOI] [PubMed] [Google Scholar]
- 33. Widera C, Pencina MJ, Meisner A, Kempf T, Bethmann K, Marquardt I, Katus HA, Giannitsis E, Wollert KC. Adjustment of the grace score by growth differentiation factor 15 enables a more accurate appreciation of risk in non‐ST‐elevation acute coronary syndrome. Eur Heart J. 2012;33:1095–1104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Widera C, Pencina MJ, Bobadilla M, Reimann I, Guba‐Quint A, Marquardt I, Bethmann K, Korf‐Klingebiel M, Kempf T, Lichtinghagen R, Katus HA, Giannitsis E, Wollert KC. Incremental prognostic value of biomarkers beyond the GRACE (Global Registry of Acute Coronary Events) score and high‐sensitivity cardiac troponin T in non‐ST‐elevation acute coronary syndrome. Clin Chem. 2013;59:1497–1505. [DOI] [PubMed] [Google Scholar]
- 35. Damman P, Kempf T, Windhausen F, van Straalen JP, Guba‐Quint A, Fischer J, Tijssen JG, Wollert KC, de Winter RJ, Hirsch A. Growth‐differentiation factor 15 for long‐term prognostication in patients with non‐ST‐elevation acute coronary syndrome: an invasive versus conservative treatment in unstable coronary syndromes (ictus) substudy. Int J Cardiol. 2014;172:356–363. [DOI] [PubMed] [Google Scholar]
- 36. Wallentin L, Lindholm D, Siegbahn A, Wernroth L, Becker RC, Cannon CP, Cornel JH, Himmelmann A, Giannitsis E, Harrington RA, Held C, Husted S, Katus HA, Mahaffey KW, Steg PG, Storey RF, James SK. Biomarkers in relation to the effects of ticagrelor in comparison with clopidogrel in non‐ST‐elevation acute coronary syndrome patients managed with or without in‐hospital revascularization: a substudy from the Prospective Randomized Platelet Inhibition and Patient Outcomes (PLATO) trial. Circulation. 2014;129:293–303. [DOI] [PubMed] [Google Scholar]
- 37. Dominguez‐Rodriguez A, Abreu‐Gonzalez P, Hernandez‐Baldomero IF, Avanzas P, Bosa‐Ojeda F. Change in growth differentiation factor 15, but not c‐reactive protein, independently predicts major cardiac events in patients with non‐ST elevation acute coronary syndrome. Mediators Inflamm. 2014;2014:929536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Fuernau G, Poenisch C, Eitel I, de Waha S, Desch S, Schuler G, Adams V, Werdan K, Zeymer U, Thiele H. Growth‐differentiation factor 15 and osteoprotegerin in acute myocardial infarction complicated by cardiogenic shock: a biomarker substudy of the IABP‐SHOCK II‐trial. Eur J Heart Fail. 2014;16:880–887. [DOI] [PubMed] [Google Scholar]
- 39. Wallentin L, Lindhagen L, Arnstrom E, Husted S, Janzon M, Johnsen SP, Kontny F, Kempf T, Levin LA, Lindahl B, Stridsberg M, Stahle E, Venge P, Wollert KC, Swahn E, Lagerqvist B. Early invasive versus non‐invasive treatment in patients with non‐ST‐elevation acute coronary syndrome (FRISC‐II): 15 year follow‐up of a prospective, randomised, multicentre study. Lancet. 2016;388:1903–1911. [DOI] [PubMed] [Google Scholar]
- 40. Dominguez‐Rodriguez A, Abreu‐Gonzalez P, Avanzas P, Consuegra‐Sanchez L. Effect of serum growth differentiation factor‐15 and the syntax score on 2‐year outcomes in patients with acute coronary syndrome. Am J Cardiol. 2016;117:1569–1574. [DOI] [PubMed] [Google Scholar]
- 41. Skau E, Henriksen E, Wagner P, Hedberg P, Siegbahn A, Leppert J. GDF‐15 and TRAIL‐R2 are powerful predictors of long‐term mortality in patients with acute myocardial infarction. Eur J Prev Cardiol. 2017;24:1576–1583. [DOI] [PubMed] [Google Scholar]
- 42. Lindholm D, James SK, Gabrysch K, Storey RF, Himmelmann A, Cannon CP, Mahaffey KW, Steg PG, Held C, Siegbahn A, Wallentin L. Association of multiple biomarkers with risk of all‐cause and cause‐specific mortality after acute coronary syndromes: a secondary analysis of the plato biomarker study. JAMA Cardiol. 2018;3:1160–1166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Dominguez‐Rodriguez A, Abreu‐Gonzalez P, Avanzas P. Relation of growth‐differentiation factor 15 to left ventricular remodeling in ST‐segment elevation myocardial infarction. Am J Cardiol. 2011;108:955–958. [DOI] [PubMed] [Google Scholar]
- 44. Ridker PM, Cannon CP, Morrow D, Rifai N, Rose LM, McCabe CH, Pfeffer MA, Braunwald E. C‐reactive protein levels and outcomes after statin therapy. N Engl J Med. 2005;352:20–28. [DOI] [PubMed] [Google Scholar]
- 45. Daniels LB, Clopton P, Laughlin GA, Maisel AS, Barrett‐Connor E. Growth‐differentiation factor‐15 is a robust, independent predictor of 11‐year mortality risk in community‐dwelling older adults: the rancho bernardo study. Circulation. 2011;123:2101–2110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Tentzeris I, Farhan S, Freynhofer MK, Rohla M, Jarai R, Vogel B, Baumgartner‐Parzer S, Nurnberg M, Geppert A, Wessely E, Wojta J, Huber K, Kautzky‐Willer A. Usefulness of elevated levels of growth differentiation factor‐15 to classify patients with acute coronary syndrome having percutaneous coronary intervention who would benefit from high‐dose statin therapy. Am J Cardiol. 2017;120:747–752. [DOI] [PubMed] [Google Scholar]
- 47. Dong G, Zheng QD, Ma M, Wu SF, Zhang R, Yao RR, Dong YY, Ma H, Gao DM, Ye SL, Cui JF, Ren ZG, Chen RX. Angiogenesis enhanced by treatment damage to hepatocellular carcinoma through the release of GDF15. Cancer Med. 2018;7:820–830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Wang L, Liu Y, Li W, Song Z. Growth differentiation factor 15 promotes cell viability, invasion, migration, and angiogenesis in human liver carcinoma cell line HepG2. Clin Res Hepatol Gastroenterol. 2017;41:408–414. [DOI] [PubMed] [Google Scholar]
- 49. de Jager SC, Bermudez B, Bot I, Koenen RR, Bot M, Kavelaars A, de Waard V, Heijnen CJ, Muriana FJ, Weber C, van Berkel TJ, Kuiper J, Lee SJ, Abia R, Biessen EA. Growth differentiation factor 15 deficiency protects against atherosclerosis by attenuating CCR2‐mediated macrophage chemotaxis. J Exp Med. 2011;208:217–225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Kim Y, Noren Hooten N, Evans MK. CRP stimulates GDF‐15 expression in endothelial cells through p53. Mediators Inflamm. 2018;2018:8278039 DOI: 10.1155/2018/8278039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Bonaterra GA, Zugel S, Thogersen J, Walter SA, Haberkorn U, Strelau J, Kinscherf R. Growth differentiation factor‐15 deficiency inhibits atherosclerosis progression by regulating interleukin‐6‐dependent inflammatory response to vascular injury. J Am Heart Assoc. 2012;1:e002550 DOI: 10.1161/JAHA.112.002550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Gopal DM, Larson MG, Januzzi JL, Cheng S, Ghorbani A, Wollert KC, Kempf T, D'Agostino RB, Polak JF, Ramachandran VS, Wang TJ, Ho JE. Biomarkers of cardiovascular stress and subclinical atherosclerosis in the community. Clin Chem. 2014;60:1402–1408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Silvola JM, Saraste A, Laitinen I, Savisto N, Laine VJ, Heinonen SE, Yla‐Herttuala S, Saukko P, Nuutila P, Roivainen A, Knuuti J. Effects of age, diet, and type 2 diabetes on the development and FDG uptake of atherosclerotic plaques. JACC Cardiovasc Imaging. 2011;4:1294–1301. [DOI] [PubMed] [Google Scholar]
- 54. Johnen H, Kuffner T, Brown DA, Wu BJ, Stocker R, Breit SN. Increased expression of the TGF‐b superfamily cytokine MIC‐1/GDF15 protects ApoE(‐/‐) mice from the development of atherosclerosis. Cardiovasc Pathol. 2012;21:499–505. [DOI] [PubMed] [Google Scholar]
- 55. Ago T, Sadoshima J. GDF15, a cardioprotective TGF‐beta superfamily protein. Circ Res. 2006;98:294–297. [DOI] [PubMed] [Google Scholar]
- 56. Preusch MR, Baeuerle M, Albrecht C, Blessing E, Bischof M, Katus HA, Bea F. GDF‐15 protects from macrophage accumulation in a mouse model of advanced atherosclerosis. Eur J Med Res. 2013;18:19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Engelse MA, Neele JM, van Achterberg TA, van Aken BE, van Schaik RH, Pannekoek H, de Vries CJ. Human activin‐A is expressed in the atherosclerotic lesion and promotes the contractile phenotype of smooth muscle cells. Circ Res. 1999;85:931–939. [DOI] [PubMed] [Google Scholar]
- 58. Li MO, Wan YY, Sanjabi S, Robertson AK, Flavell RA. Transforming growth factor‐beta regulation of immune responses. Annu Rev Immunol. 2006;24:99–146. [DOI] [PubMed] [Google Scholar]
- 59. Ridker PM, Everett BM, Thuren T, MacFadyen JG, Chang WH, Ballantyne C, Fonseca F, Nicolau J, Koenig W, Anker SD, Kastelein JJP, Cornel JH, Pais P, Pella D, Genest J, Cifkova R, Lorenzatti A, Forster T, Kobalava Z, Vida‐Simiti L, Flather M, Shimokawa H, Ogawa H, Dellborg M, Rossi PRF, Troquay RPT, Libby P, Glynn RJ. Antiinflammatory therapy with canakinumab for atherosclerotic disease. N Engl J Med. 2017;377:1119–1131. [DOI] [PubMed] [Google Scholar]
- 60. Wang J, Chen H, Sun J, Hippe DS, Zhang H, Yu S, Cai J, Xie L, Cui B, Yuan C, Zhao X, Yuan W, Liu H. Dynamic contrast‐enhanced mr imaging of carotid vasa vasorum in relation to coronary and cerebrovascular events. Atherosclerosis. 2017;263:420–426. [DOI] [PubMed] [Google Scholar]
- 61. Huh SJ, Chung CY, Sharma A, Robertson GP. Macrophage inhibitory cytokine‐1 regulates melanoma vascular development. Am J Pathol. 2010;176:2948–2957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Song H, Yin D, Liu Z. GDF‐15 promotes angiogenesis through modulating p53/HIF‐1α signaling pathway in hypoxic human umbilical vein endothelial cells. Mol Biol Rep. 2012;39:4017–4022. [DOI] [PubMed] [Google Scholar]
- 63. Yao Y, Bennett BJ, Wang X, Rosenfeld ME, Giachelli C, Lusis AJ, Bostrom KI. Inhibition of bone morphogenetic proteins protects against atherosclerosis and vascular calcification. Circ Res. 2010;107:485–494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Khaidakov M, Szwedo J, Mitra S, Mehta JL. Angiostatic effects of aspirin in hypoxia‐reoxygenation are linked to modulation of TGFβ1 signaling. J Cardiovasc Pharmacol Ther. 2011;16:105–110. [DOI] [PubMed] [Google Scholar]
- 65. Schlittenhardt D, Schober A, Strelau J, Bonaterra GA, Schmiedt W, Unsicker K, Metz J, Kinscherf R. Involvement of growth differentiation factor‐15/macrophage inhibitory cytokine‐1 (GDF‐15/MIC‐1) in oxLDL‐induced apoptosis of human macrophages in vitro and in arteriosclerotic lesions. Cell Tissue Res. 2004;318:325–333. [DOI] [PubMed] [Google Scholar]
- 66. Desmedt S, Desmedt V, De Vos L, Delanghe JR, Speeckaert R, Speeckaert MM. Growth differentiation factor 15: a novel biomarker with high clinical potential. Crit Rev Clin Lab Sci. 2019;56:333–350. [DOI] [PubMed] [Google Scholar]
- 67. Johnen H, Lin S, Kuffner T, Brown DA, Tsai VW, Bauskin AR, Wu L, Pankhurst G, Jiang L, Junankar S, Hunter M, Fairlie WD, Lee NJ, Enriquez RF, Baldock PA, Corey E, Apple FS, Murakami MM, Lin EJ, Wang C, During MJ, Sainsbury A, Herzog H, Breit SN. Tumor‐induced anorexia and weight loss are mediated by the TGF‐beta superfamily cytokine MIC‐1. Nat Med. 2007;13:1333–1340. [DOI] [PubMed] [Google Scholar]
- 68. Tsai VW, Macia L, Johnen H, Kuffner T, Manadhar R, Jorgensen SB, Lee‐Ng KK, Zhang HP, Wu L, Marquis CP, Jiang L, Husaini Y, Lin S, Herzog H, Brown DA, Sainsbury A, Breit SN. TGF‐b superfamily cytokine MIC‐1/GDF15 is a physiological appetite and body weight regulator. PLoS One. 2013;8:e55174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Unal B, Alan S, Bassorgun CI, Karakas AA, Elpek GO, Ciftcioglu MA. The divergent roles of growth differentiation factor‐15 (GDF‐15) in benign and malignant skin pathologies. Arch Dermatol Res. 2015;307:551–557. [DOI] [PubMed] [Google Scholar]
- 70. Eling TE, Baek SJ, Shim M, Lee CH. NSAID activated gene (NAG‐1), a modulator of tumorigenesis. J Biochem Mol Biol. 2006;39:649–655. [DOI] [PubMed] [Google Scholar]
- 71. Brown DA, Moore J, Johnen H, Smeets TJ, Bauskin AR, Kuffner T, Weedon H, Milliken ST, Tak PP, Smith MD, Breit SN. Serum macrophage inhibitory cytokine 1 in rheumatoid arthritis: a potential marker of erosive joint destruction. Arthritis Rheum. 2007;56:753–764. [DOI] [PubMed] [Google Scholar]
- 72. Breit SN, Carrero JJ, Tsai VW, Yagoutifam N, Luo W, Kuffner T, Bauskin AR, Wu L, Jiang L, Barany P, Heimburger O, Murikami MA, Apple FS, Marquis CP, Macia L, Lin S, Sainsbury A, Herzog H, Law M, Stenvinkel P, Brown DA. Macrophage inhibitory cytokine‐1 (MIC‐1/GDF15) and mortality in end‐stage renal disease. Nephrol Dial Transplant. 2012;27:70–75. [DOI] [PubMed] [Google Scholar]
- 73. Hellemons ME, Mazagova M, Gansevoort RT, Henning RH, de Zeeuw D, Bakker SJ, Lambers‐Heerspink HJ, Deelman LE. Growth‐differentiation factor 15 predicts worsening of albuminuria in patients with type 2 diabetes. Diabetes Care. 2012;35:2340–2346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Mensching L, Borger AK, Wang X, Charalambous P, Unsicker K, Haastert‐Talini K. Local substitution of GDF‐15 improves axonal and sensory recovery after peripheral nerve injury. Cell Tissue Res. 2012;350:225–238. [DOI] [PubMed] [Google Scholar]
- 75. Van der Veken B, De Meyer GRY, Martinet W. Axitinib attenuates intraplaque angiogenesis, haemorrhages and plaque destabilization in mice. Vascul Pharmacol. 2018;100:34–40. [DOI] [PubMed] [Google Scholar]
- 76. Kempf T, Zarbock A, Widera C, Butz S, Stadtmann A, Rossaint J, Bolomini‐Vittori M, Korf‐Klingebiel M, Napp LC, Hansen B, Kanwischer A, Bavendiek U, Beutel G, Hapke M, Sauer MG, Laudanna C, Hogg N, Vestweber D, Wollert KC. GDF‐15 is an inhibitor of leukocyte integrin activation required for survival after myocardial infarction in mice. Nat Med. 2011;17:581–588. [DOI] [PubMed] [Google Scholar]
- 77. Kempf T, Eden M, Strelau J, Naguib M, Willenbockel C, Tongers J, Heineke J, Kotlarz D, Xu J, Molkentin JD, Niessen HW, Drexler H, Wollert KC. The transforming growth factor‐beta superfamily member growth‐differentiation factor‐15 protects the heart from ischemia/reperfusion injury. Circ Res. 2006;98:351–360. [DOI] [PubMed] [Google Scholar]
- 78. Xu J, Kimball TR, Lorenz JN, Brown DA, Bauskin AR, Klevitsky R, Hewett TE, Breit SN, Molkentin JD. GDF15/MIC‐1 functions as a protective and antihypertrophic factor released from the myocardium in association with smad protein activation. Circ Res. 2006;98:342–350. [DOI] [PubMed] [Google Scholar]
- 79. Mimeault M, Batra SK. Divergent molecular mechanisms underlying the pleiotropic functions of macrophage inhibitory cytokine‐1 in cancer. J Cell Physiol. 2010;224:626–635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Tsai VW, Lin S, Brown DA, Salis A, Breit SN. Anorexia‐cachexia and obesity treatment may be two sides of the same coin: role of the TGF‐b superfamily cytokine GDF15/MIC‐1. Int J Obes. 2016;40:193–197. [DOI] [PubMed] [Google Scholar]
- 81. Schaub N, Reichlin T, Twerenbold R, Reiter M, Steuer S, Bassetti S, Stelzig C, Wolf C, Winkler K, Haaf P, Meissner J, Drexler B, Mueller C. Growth differentiation factor‐15 in the early diagnosis and risk stratification of patients with acute chest pain. Clin Chem. 2012;58:441–449. [DOI] [PubMed] [Google Scholar]
- 82. Mehta RS, Song M, Bezawada N, Wu K, Garcia‐Albeniz X, Morikawa T, Fuchs CS, Ogino S, Giovannucci EL, Chan AT. A prospective study of macrophage inhibitory cytokine‐1 (MIC‐1/GDF‐15) and risk of colorectal cancer. J Natl Cancer Inst. 2014;106:dju016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Muppala S, Xiao R, Krukovets I, Verbovetsky D, Yendamuri R, Habib N, Raman P, Plow E, Stenina‐Adognravi O. Thrombospondin‐4 mediates TGF‐beta‐induced angiogenesis. Oncogene. 2017;36:5189–5198. [DOI] [PMC free article] [PubMed] [Google Scholar]