

Abstract
Intestinal microbiome and gut-brain axis have been receiving increasing attention for their role in the regulation of brain/behavior and possible biological basis of psychiatric disorders. Several recent clinical studies have linked the microbiome with neuropsychiatric conditions, although the literature on schizophrenia is quite limited. This study investigated gut microbiome composition in 50 individuals, including 25 persons with chronic schizophrenia and 25 demographically-matched non-psychiatric comparison subjects (NCs). Stool samples were collected and assayed using 16S rRNA sequencing of the V4 region. Examination of unweighted UniFrac and Bray-Curtis dissimilarities revealed significant community-level separation in microbiome composition between the two subject groups. At the phylum level, Proteobacteria were found to be relatively decreased in schizophrenia subjects compared to NCs. At the genus level, Anaerococcus was relatively increased in schizophrenia while Haemophilus, Sutterella, and Clostridium were decreased. Within individuals with schizophrenia, abundance of Ruminococcaceae was correlated with lower severity of negative symptoms; Bacteroides were associated with worse depressive symptoms; and Coprococcus was related to greater risk for developing coronary heart disease. Our findings provide evidence of altered gut microbial composition in persons with chronic schizophrenia and suggest a need for larger and longitudinal studies of microbiome in schizophrenia.
Keywords: psychosis, microbes, gut-brain axis, bacteria, probiotics, phylum
1. INTRODUCTION
Recent years have seen a rapid growth of interest in the human gut microbiome in the pathophysiology of human health and disease. The microbiome is a dynamic ecological community of microorganisms that inhabit the human body. The gut microbiome has a pivotal role in regulating inflammatory and metabolic pathways across a number of physical diseases, including gastrointestinal (GI) disorders, such as inflammatory bowel disease (Kostic et al., 2014), obesity and metabolic diseases (Bouter et al., 2017; Hartstra et al., 2015), cancer (Schwabe and Jobin, 2013), and chronic pulmonary diseases (Budden et al., 2017; O’Dwyer et al., 2016). There is also an emerging body of empirical support for the view that the gut microbiome can impact brain and behavior (Collins et al., 2012; Cryan and Dinan, 2012; Mayer, 2011). A number of animal studies suggest that the alterations in gut microbiota can induce physiological and behavioral features that resemble psychiatric disorders (e.g., Sudo et al., 2004; Zheng et al., 2016). However, human studies in this area have been few, particularly in schizophrenia (Nguyen et al., 2018).
Converging evidence suggests that schizophrenia and other serious mental illnesses are associated with chronic systemic and gastrointestinal inflammation, oxidative stress, and metabolic dysfunction (Nguyen et al., 2018). Some studies have found that schizophrenia is associated with elevated serological biomarkers of microbial translocation (Severance et al., 2012; Severance et al., 2016; Severance et al., 2013), suggesting increased permeability of the intestinal lumen, which may be a mechanism by which intestinal dysbiosis impacts systemic physiological functioning. Additionally, gut and digestive disturbances, including irritable bowel syndrome, colitis, and celiac disease, are highly prevalent comorbidities in persons with schizophrenia (Severance et al., 2015). Gastrointestinal diseases are the third leading cause of natural deaths in schizophrenia (Saha et al., 2007). Considering that microbial colonization of the gut is crucial for optimal function of the immune system (Kamada et al., 2013; Round and Mazmanian, 2009), inflammation, oxidative stress, and other physiological dysfunctions implicated in schizophrenia might be, at least in part, associated with changes in the microbiome.
The number of empirical studies of the gut microbiome in persons with schizophrenia is very limited (Nguyen et al., 2018). The four published studies of the gut microbiome in psychotic disorders include a study of individuals at high-risk and ultra-high-risk for schizophrenia (He et al., 2018), two investigations of patients with first-episode psychosis (Schwarz et al., 2018; Yuan et al., 2018), and a report on persons with schizophrenia of less than 10 years’ duration (Shen et al., 2018). These studies revealed that the species composition within the gut microbiomes of individual with or at risk for psychotic disorders is different from that of non-psychiatric comparison subjects (NCs), with varying bacterial taxa driving community separation in each study. Only one of these studies investigated the relationship between gut bacteria and clinical characteristics; first-episode patients with the greatest abnormalities in microbiota composition had more severe psychotic symptoms and worse global functioning at hospitalization and a lower rate of disease remission at one-year follow-up (Schwarz et al., 2018).
These studies focused on psychosis at earlier stages in the disease process and were conducted on European and Chinese subjects. Microbial composition varies across individuals with different countries of residence (McDonald et al., 2018). One study (Shen et al., 2018) excluded individuals with any chronic disease that may affect the stability of gut microbiota, including hypertension and diabetes, which are common in schizophrenia. To our knowledge, the present report is the first investigation of gut microbiome in US-based patients with chronic schizophrenia.
Our objective was to characterize gut microbiome in patients with schizophrenia, with duration of >10 years (range 12–56 years). We hypothesized that gut microbial composition would differ between schizophrenia subjects and NCs. We also sought to explore relationships of microbial composition differences with clinical and disease characteristics, specifically with psychiatric symptomatology, smoking, physical comorbidity, and antipsychotic medications.
2. METHODS
2.1. Participants
Fifty subjects, including 25 outpatients with schizophrenia or schizoaffective disorder (hereafter referred to collectively as schizophrenia) and 25 demographically matched non-psychiatric comparison subjects (NC), between the ages of 30 and 76 years, participated in this study. The diagnosis of schizophrenia was made based on the Structured Clinical Interview for the DSM-IV-TR (SCID) (First et al., 2002). Twenty-one patients were on antipsychotic medication at the time of the study. NC participants were recruited through multiple methods, including from an ongoing survey study of successful aging in healthy adults at UCSD, recruitment flyers in the community, www.ResearchMatch.org, and word-of-mouth. See Hong et al. (2017), Joseph et al. (2015), and Lee et al. (2016; 2017) for additional details regarding recruitment and subject selection criteria. NCs were screened using the Mini-International Neuropsychiatric Interview and excluded from the study if they had a past or present diagnosis of a major neuropsychiatric illnesses (Sheehan et al., 1998). Exclusion criteria were: other current major DSM-IV-TR Axis I diagnoses; alcohol or other substance (other than tobacco) abuse or dependence within 3 months prior to enrollment; diagnosis of dementia, intellectual disability disorder, or a major neurological disorder; or any medical disability that interfered with a subject’s ability to complete study procedures. We did not exclude persons with chronic diseases such as hypertension and diabetes. Although these conditions may influence the stability of the gut microbiota, patients with schizophrenia tend to have higher rates of these chronic medical conditions (De Hert et al., 2009; Hennekens et al., 2005). All procedures were approved by the UCSD Human Research Protections Program. All participants provided written informed consent.
Given that technical differences (e.g., DNA extraction methods, sequencing platforms, taxonomy databases) can produce systematic biases that may obscure biologically meaningful compositional differences (Lozupone et al., 2013; Walters et al., 2014), NC subjects were matched to schizophrenia subjects on sequencing plate after initial recruitment. We found the nearest matching neighbors on the sequencing plate based on age, sex, race, BMI category (obese vs. not obese), and history of antibiotic use (in the past year) to control for clinical factors and known major drivers of microbiome changes (McDonald et al., 2018) that could confound the results.
2.2. Sociodemographic and clinical assessment
Sociodemographic characteristics (i.e., age, gender, race/ethnicity, current smoking status) and illness-related factors (i.e., age of onset, duration of illness, antipsychotic medication) were ascertained through participant interview and review of available medical records. Daily dosages of antipsychotic medication were converted to total World Health Organization defined daily dose (Organization, 2009). BMI was based on assessment of height and weight (kg/m2). Positive and negative psychiatric symptoms were evaluated using interviewer-administered Scales for Assessment of Positive Symptoms and Negative Symptoms (SAPS and SANS, respectively; Andreasen, 1983, 1984). Depression was assessed using the Patient Health Questionnaire (PHQ-9; Kroenke and Spitzer, 2002). Health-related quality of life and functioning was evaluated using the physical and mental health component scores from the Medical Outcomes Study 36-item Short Form (SF-36; Ware Jr and Sherbourne, 1992). Medical comorbidity was measured with the total score and severity index from the Cumulative Illness Rating Scale (CIRS; Parmelee et al., 1995). The Framingham 10-year CHD relative risk score was calculated according to Wilson, et al. (1998), and Framingham 10-year CVD Risk according to D’Agostino, et al. (2008).
2.3. Stool collection and sequencing
Participants were provided with home stool collection kits (BD SWUBE Dual Swab Collection System; BD Worldwide) and given detailed instructions about how to collect stool samples. Participants returned samples via mail in a self-addressed envelope. Returned stool samples were frozen at −80°C until aliquoted into 96 well plates for DNA extraction.
DNA extraction, amplification, and sequencing were performed in the same way as for the American Gut Project samples (americangut.org; McDonald et al., 2018). Genomic DNA was extracted from samples using the Earth Microbiome 16S rRNA amplicon extraction protocol (Earth Microbiome Project, 2016). PCR amplification and library preparation were performed similarly to the protocol described by Caporaso et al. (2012). Illumina primers with unique reverse primer barcodes were used to target the V4 region of the 16S ribosomal RNA gene. V4 amplicons were sequenced on the Illumina HiSeq 2000 platform, yielding paired-end, 150-base-pair reads. Sequencing was performed at the UCSD IGM Genomics Center. Feature tables can be found in Qiita (qiita.ucsd.edu) as study ID 11710 and sequences can be found in EBI-ENA under accession number EBI: ERP107975.
2.4. Bioinformatics and statistical analyses
Sequence data were processed using QIIME2 (qiime2.org; Caporaso et al., 2010). Raw sequencing results were demultiplexed and microbial sOTUs (sub-Operational Taxonomic Units) were identified using Deblur algorithm (Amir et al., 2017). The output feature table was rarefied to 7,905 sequences per sample (the lowest value in the dataset). Microbial community structure was characterized using measures of alpha-diversity (within-sample) and beta-diversity (between-samples). Alpha-diversity represents the number (richness) and distribution (evenness) of taxa within a single sample. Metrics of alpha-diversity included number of observed features to represent species richness, Shannon diversity index to quantify species abundance and evenness (Shannon, 1948), and Faith’s Phylogenetic Diversity which measures the total length of branches in a reference phylogenetic tree for all species in a given sample (Faith, 1992). Beta-diversity indicates differences in taxa composition between samples based on either presence-absence or quantitative species abundance data, and may be presented in a distance matrix. Beta-diversities were calculated using Bray-Curtis dissimilarity (Sørensen, 1948) and unweighted UniFrac (Lozupone and Knight, 2005; Lozupone et al., 2007). Output matrices were ordinated using principal coordinate analysis (PCoA) and visualized using EMPeror (Vazquez-Baeza et al., 2013).
2.4.1. Data Analysis:
Statistical significance of beta-diversity distances between groups was assessed using PERMANOVA with 999 permutations. Alpha-diversity group significance was calculated using nonparametric Kruskal-Wallis H test in QIIME2. For taxonomic analyses and differential abundance testing, taxonomy information was provided for each sOTU sequence using UCLUST method implemented in QIIME 1 (Caporaso et al., 2010; Edgar, 2010). All differential abundance testing, at different taxonomic levels were performed using Calour (github.com/biocore/calour) with mean difference permutive test (equivalent to Kruskal-Wallis H test in the case of 2 groups) at 10,000 permutations with discrete False Discovery Rate (FDR) correction at α=0.1 (Jiang et al., 2017). To determine associations between specific taxa and clinical characteristics, data were pre-processed in the same way as for differential abundance testing (filtering features with less than 1,000 reads, normalizing data to 10,000 reads and clustering features with less than 100 reads together). Pearson’s correlations were then calculated between each sOTU (feature) and demographic and/or clinical characteristics within schizophrenia and NC groups separately; a t-statistic test using 1,000 permutations (where labels are randomly changed between samples) with discrete FDR correction was applied to determine the statistical significance of features (Jiang et al., 2017).
3. RESULTS
Demographic and clinical characteristics for groups are presented in Table 1. Schizophrenia and NC groups did not differ on age, gender, or race. As expected, persons with schizophrenia had worse psychiatric symptoms, higher depression and anxiety levels, lower levels of physical well-being, higher rates of smoking, and greater medical comorbidity. Comparing exact BMI values, persons with schizophrenia had higher BMI than NCs; however, in terms of BMI classification, there were no significant differences between the two groups.
Table 1.
Demographic and clinical characteristics of participants
| Schizophrenia (n = 25) |
Non-Psychiatric Comparison (n = 25) |
t or χ2 | p | |
|---|---|---|---|---|
| Age (years) | 52.9 (11.2) | 54.7 (10.7) | 0.58 | 0.56 |
| Gender [n (% female)] | 11 (44%) | 10 (40%) | 0.08 | 0.77 |
| Race [n (% Caucasian) | 16 (64%) | 19 (76%) | 1.07 | 0.79 |
| BMI | 31.8 (5.4) | 28.9 (4.0) | −2.17 | 0.04 |
| BMI classification1 [n (% obese)] | 16 (64%) | 10 (40%) | 2.89a | 0.09 |
| Antibiotic use [n (% in past year)] | 7 (28%) | 5 (20%) | 0.44a | 0.51 |
| Current smoking status [n (% smoker) | 14 (56%) | 1 (4%) | 16.1 | <0.001 |
| Smoking frequency | Daily: 12 Regularly3: 1 Rarely4: 1 Never: 11 |
Daily: 1 Never: 24 |
16.1 | 0.001 |
| Age of illness onset (years) | 21.5 (7.0) | -- | -- | -- |
| Illness duration (years) | 32.4 (11.1) | -- | -- | -- |
| Antipsychotic daily dosage2 | 2.01 (2.6) | -- | -- | -- |
| SAPS Total Score | 4.56 (3.3) | 0.67 (0.8) | −5.31 | <0.001 |
| SANS Total Score | 4.16 (5.1) | 0.50 (0.8) | −1.73 | 0.10 |
| PHQ-9 Severity Score | 7.21 (6.1) | 3.20 (2.3) | −2.96 | 0.006 |
| BSI Anxiety Score | 8.25 (6.1) | 2.00 (3.0) | −4.43 | <0.001 |
| SF-36 Mental Component | 43.5 (12.9) | 51.6 (8.3) | 2.40 | 0.2 |
| SF-36 Physical Component | 43.3 (11.6) | 52.8 (7.5) | 3.13 | 0.003 |
| CIRS Total Score | 6.6 (2.8) | 1.5 (2.0) | −4.24 | <0.001 |
| CIRS Severity Score | 1.57 (0.3) | 0.72 (0.9) | −2.39 | 0.06 |
| Framingham CHD risk score | 1.69 (1.0) | 0.94 (0.13) | −3.15 | 0.005 |
| Framingham CVD risk score | 19.1 (15.3) | 6.53 (2.28) | −3.48 | 0.002 |
| Medical diagnoses | ||||
| Diabetes [n (% with)] | 9 (36%) | 1 (4%) | 8.0 | 0.005 |
| Hypertension [n (% with)] | 19 (76%) | 2 (8%) | 4.0 | 0.05 |
Not applicable, groups were matched on these variables
χ2 value
World Health Organization classification; comparison of obese (BMI≥30) to non-obese (BMI<30) individuals
World Health Organization defined daily dose
Regularly = 3–5 times/week
Rarely = a few times/month
BMI = body mass index; BSI = Brief Symptom Inventory; CHD = coronary heart disease; CVD = cardiovascular disease; PHQ-9 = Patient Health Questionnaire; SANS = Scale for the Assessment of Negative Symptoms; SAPS = Scale for the Assessment of Positive Symptoms
3.1. Microbial community structure
There was no difference between schizophrenia and NC groups on any assessed measure of alpha-diversity (Supplemental Table 1). Analysis of beta-diversity indices revealed significant community-level separation between schizophrenia and NC groups using unweighted UniFrac (pseudo-F = 1.81, p = 0.005) and Bray-Curtis (pseudo-F = 2.20, p = 0.003). PCoA of unweighted UniFrac and Bray-Curtis distances showed that schizophrenia and NC groups formed distinct clusters (Figure 1). NCs also clustered more tightly than schizophrenia subjects, who were spread more widely across the PCoA space (within-group distance comparison with a two-sided t-test; unweighted UniFrac p < 0.001; Bray-Curtis p = 0.02). Across groups, significant beta-diversity differences were also observed for sex (unweighted UniFrac pseudo-F = 1.66, p = 0.007) and smoking status (unweighted UniFrac pseudo-F = 1.43, p = 0.027; Bray-Curtis pseudo-F = 1.55, p = 0.042), the latter was likely driven by diagnosis, as all but one smoker were schizophrenia subjects. Microbial community separation related to diagnostic group demonstrated the highest effect size (pseudo-F = 1.81).
Figure 1.
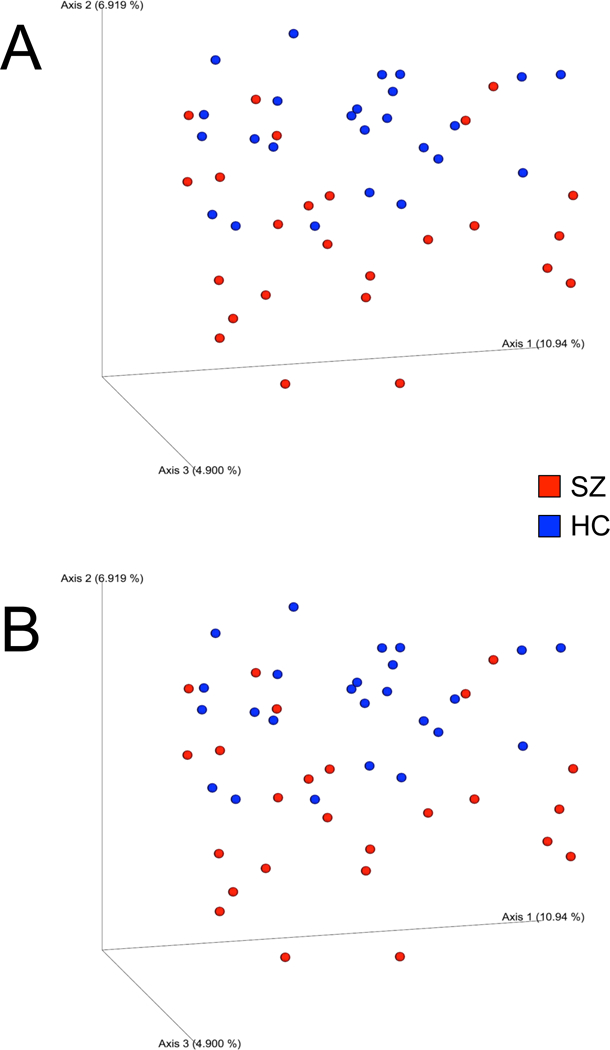
Principal coordinates analysis (PCoA) plot illustrating beta-diversity distance matrices of unweighted UniFrac distance (A) and Bray-Curtis distance (B) comparing sample distributions between schizophrenia and NC groups. Red dots represent schizophrenia patients, and blue dots represent NCs.
3.2. Bacterial taxonomic composition
Differential abundance testing at different taxonomic levels was performed to identify the drivers of community separation. At the phylum level, Proteobacteria was observed to be differentially abundant between groups, with schizophrenia subjects having decreased relative abundance of this phylum compared to NCs (Supplemental Figure 1). On the genus level, four differentially abundant genera were revealed. Anaerococcus was relatively increased in schizophrenia compared to NC (H = 8.32; p = 0.007), while Haemophilus (H = −11.3; p = 0.004), Sutterella (H = −12.0; p = 0.004), and Clostridium (H = −15.9; p = 0.0002) were decreased (Figure 2). When differential abundance testing was performed across taxonomic levels, 35 individual taxa were found to be differentially abundant between schizophrenia subjects and NCs. Thirty-three sOTUs belonged to the order Clostridiales, 1 sOTU to class Gammaproteobacteria (Haemophilus parainfluenzae) and 1 sOTU to class Erysipelotrichi (unknown genus) (Supplemental Table 2).
Figure 2.

Heat map illustrating differentially abundant genera in the two groups. Each vertical color bar represents an individual subject. The color of the bar indicates the relative abundance of each genus within a subject. The vertical color bar on the right side of the graph delineates relative abundance, and the horizontal bar at the top of the graph denotes schizophrenia and non-psychiatric comparison groups
3.3. Relationship with clinical characteristics
Age was not significantly associated with alpha-diversity (species richness, abundance, or evenness) across the entire sample (rs < 0.16; ps > 0.29) or within either schizophrenia (rs < 0.04; ps > 0.86) or NC (rs < 0.35; ps > 0.08) groups. Within schizophrenia subjects, age of disease onset was positively correlated with phylum Cyanobacteria (r = 0.53; p = 0.008), such that persons with earlier onset of illness had lower abundance of this taxa. No relationship was found between illness duration and specific taxa.
Specific microbial taxa were associated with level of psychopathology in schizophrenia. Greater severity of depressive symptoms was correlated with greater abundance of genus Bacteroides (r = 0.70; p = 0.0002), while increased negative symptoms were associated with decreased abundance of family Ruminococcaceae (r = −0.74; p = 0.0002). Overall self-reported mental well-being was positively correlated with phylum Verrucomicrobia (r = 0.63; p = 0.002). Higher level of the genus Coprococcus was associated with greater Framingham CHD risk (r = 0.77; p = 0.0003). Schizophrenia patients with and without diabetes (unweighted UniFrac pseudo-F = 1.21, p = 0.11; Bray-Curtis pseudo-F = 1.03, p = 0.39), hypertension (unweighted UniFrac pseudo-F = 0.82, p = 0.86; Bray-Curtis pseudo-F = 1.04, p = 0.38), or heart disease (unweighted UniFrac pseudo-F = 1.00, p = 0.41; Bray-Curtis pseudo-F = 1.15, p = 0.26) did not differ in gut microbial composition.
To disentangle the effects of smoking and diagnosis group, we compared smokers (n = 14) to non-smokers (n = 11) within the schizophrenia group and found no difference in beta-diversity (unweighted UniFrac pseudo-F = 1.26, p = 0.085; Bray-Curtis pseudo-F = 0.94, p = 0.485), suggesting that differences in microbial community composition between study groups are driven by disease rather than cigarette smoking. Within the schizophrenia group, greater number of years of smoking was associated with higher levels of phylum Actinobacteria (r = 0.48; p = 0.01). We found no significant associations between specific microbial taxa and antipsychotic daily dosage levels (rs < 0.09; ps > 0.66).
4. DISCUSSION
Consistent with our hypothesis, this study found several significant differences in the overall composition of and levels of specific bacterial taxa in the gut microbiome in persons with schizophrenia compared to NCs. To our knowledge, this investigation is the first to provide evidence for altered gut microbiota in a sample of chronically ill patients with schizophrenia living in the US. Notably, most of these differences remained significant after controlling for demographic and clinical factors that may influence microbial composition. A number of bacterial taxa were differentially abundant between groups, including decreased relative abundance of phylum Proteobacteria and genera Haemophilus, Sutterella and Clostridium and increased relative abundance of genus Anaerococcus in schizophrenia compared to NCs. The finding of abnormal microbial composition in patients with schizophrenia is consistent with previous reports among other schizophrenia-spectrum (He et al., 2018; Shen et al., 2018) and first-episode psychotic (Yuan et al., 2018) disorders, although the specifics of the observed differences vary.
When considering the relative abundance of specific taxa, findings have been mixed across studies. We found that abundance of Proteobacteria and Clostridium was relatively lower in schizophrenia patients than NCs, which was opposite to findings reported by Shen and colleagues (2018). Inconsistencies across investigations may be attributable partly to heterogeneity in sample characteristics across studies. Each of the prior four studies focused on schizophrenia and psychotic disorders and different developmental stages (He et al., 2018; Schwarz et al., 2018; Shen et al., 2018; Yuan et al., 2018). Furthermore, we did not exclude individuals with certain chronic diseases, as these bodily and physiological changes are inherent to the disorder and associated lifestyle. Similar to all previous studies, patients with schizophrenia and NCs did not differ in terms of within-sample (alpha) diversity, despite overall between-sample composition divergence between patients and NCs.
Finally, our study revealed some potentially interesting information about the relationship between the gut microbiome and clinical characteristics associated with schizophrenia. We found that the composition of the gut microbiome was associated with psychopathology. Increased Ruminococcaceae was correlated with decreased negative symptoms, and Bacteroides with worse depressive symptoms. Family Ruminococcaceae has been previously reported in the literature to be associated with major depressive disorder (Jiang et al., 2015; Zheng et al., 2016) and bipolar disorder (Evans et al., 2016). Studies have generally found this family to be decreased in serious mental illnesses, including bipolar and major depression disorders (Evans et al., 2016; Jiang et al., 2015), although it was reported to be overrepresented in one study (Zheng et al., 2016). Furthermore, higher levels of this taxon have been shown to be associated with decreased anxiety and depression and better sleep and physical well-being (Evans et al., 2016). The literature on the relationship between genus Bacteroides is mixed; it has been previously found to be relatively underrepresented in patients with active depression, yet overrepresented in patients with antidepressant-responsive depression (Jiang et al., 2015).
With regards to physical health, Coprococcus was associated with greater risk for developing coronary heart disease in patients with schizophrenia. This genus has been found to be enriched among individuals with a high lifetime cardiovascular risk profile (Kelly et al., 2016) and decreased following bariatric surgery in obese patients with diabetes (Graessler et al., 2013), suggesting that it may be a marker of vascular health. Co-morbid diagnoses of diabetes, hypertension, or heart disease did not contribute significantly to microbiome differences in schizophrenia patients.
As expected, cigarette smoking was a significant driver of global microbial composition. To disentangle the effect of smoking from the effect of diagnosis in schizophrenia and NC groups, we examined patients who were smokers vs. non-smokers and found no differences in beta-diversity, suggesting that the microbial abnormalities in schizophrenia may not be primarily attributable to cigarette smoking. Increased number of years of smoking was associated with higher levels of Actinobacteria, consistent with previous studies showing Actinobacteria is relatively enriched in current smokers compared to never smokers (Wu et al., 2016). The abundance of this phylum has also been found to increase after smoking cessation (Biedermann et al., 2013). Lastly, we did not detect any significant association between specific taxa and daily dosages of antipsychotics. Prior studies have evaluated the effects of antipsychotic medications on the gut microbiome in serious mental illnesses. In drug-naïve patients with first episode schizophrenia, 24-week risperidone treatment resulted in significant increases in the numbers of Bifidobacterium spp. and E. coli, and decreases in Clostridium coccoides group and Lactobacillus spp. (Yuan et al., 2018). Current atypical antipsychotic treatment was also associated with reduced gut alpha-diversity in women with bipolar disorder and significant beta-diversity separation (Flowers et al., 2016). Patients on atypical antipsychotic medications demonstrated relatively increased levels of Lachnospiraceae, while non-antipsychotic-treated individuals showed preferentially higher levels of Akkermansia.
4.1. Limitations
Several limitations to our study must be acknowledged. The sample size in this investigation was small, limiting the extent to which we could statistically explore potential confounds on gut microbial differences and the power of correlational analyses to assess the relationship of various clinical correlates and gut microbiome. Thus, these results should be considered preliminary. We were able to replicate community-level group differences found in other studies of the gut microbiome in populations with or at risk for serious mental illnesses (Evans et al., 2016; He et al., 2018; Schwarz et al., 2018; Shen et al., 2018), and we observed several intriguing relationships which should be further explored and replicated in future studies. Future investigations should aim to further assess the relationship of psychotropic medication use as well as disease severity, cognition, nutrition, and lifestyle behaviors to the gut microbiome. It is also a possibility that microbial differences between schizophrenia patients and NCs may be driven by the larger percentage of patients who have co-morbid medical illnesses (e.g., diabetes, hypertension). However, we did not find any beta-diversity differences between patients with and without these disorders, suggesting that these physical conditions did not significantly impact the differences observed between groups. Finally, the cross-sectional design restricts our ability to make causal inferences about the role of the gut microbiome in schizophrenia. Larger, prospective longitudinal studies of patients with varied duration of illness are required to determine whether intestinal dysbiosis is a key pathophysiological feature of schizophrenia and how it may impact aging in this disorder.
Despite these limitations, this study possesses several strengths. We included a NC sample that was matched to the schizophrenia sample on demographics, sequencing plate, and other clinical factors known to have major influences on the gut microbiome. Our study is also the first to examine schizophrenia patients living in the US and those with very long durations of illness. These results represent a first step in understanding the gut microbial ecosystem in clinical populations of persons with schizophrenia. These findings lay the groundwork for future investigations in elucidating the role of the gut microbiome in the development, presentation, and progression of schizophrenia and how it may be associated with (or contribute to) other physiological abnormalities (e.g., chronic inflammation, oxidative stress) in this population. Microbiome research holds promise for predicting clinical prognosis, assessing risk for medical morbidity and mortality, and informing intervention development (e.g., probiotics, prebiotics, fecal transplantation) to improve patients’ quality of life. Strategies for achieving and maintaining healthy microbiome may lead to numerous potential benefits, including improved physical and mental health, cognition, and consequently, everyday functioning.
Supplementary Material
Gut microbial composition at the phylum level for schizophrenia and NC groups. Compared to NCs, patients with schizophrenia exhibited increased relative abundance of Proteobacteria.
Summary of differentially abundant taxa between groups.
FUNDING
This work was supported, in part, by National Institute of Mental Health (grant numbers 2R01 MH094151-06 and 5T32 MH019934-24 to DVJ), UC San Diego Stein Institute for Research on Aging, and UC San Diego Center for Microbiome Innovation.
REFERENCES
- Amir A, McDonald D, Navas-Molina JA, Kopylova E, Morton JT, Zech Xu Z, Kightley EP, Thompson LR, Hyde ER, Gonzalez A, Knight R, 2017. Deblur Rapidly Resolves Single-Nucleotide Community Sequence Patterns. mSystems 2(2). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreasen NC, 1983. Scale for the assessment of Negative Symptoms (SANS). University of Iowa, Iowa City, IA. [Google Scholar]
- Andreasen NC, 1984. Scale for the Assessment of Positive Symptoms (SAPS). University of Iowa, Iowa City, IA. [Google Scholar]
- Biedermann L, Zeitz J, Mwinyi J, Sutter-Minder E, Rehman A, Ott SJ, Steurer-Stey C, Frei A, Frei P, Scharl M, Loessner MJ, Vavricka SR, Fried M, Schreiber S, Schuppler M, Rogler G, 2013. Smoking cessation induces profound changes in the composition of the intestinal microbiota in humans. PloS one 8(3), e59260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouter KE, van Raalte DH, Groen AK, Nieuwdorp M, 2017. Role of the Gut Microbiome in the Pathogenesis of Obesity and Obesity-Related Metabolic Dysfunction. Gastroenterology 152(7), 1671–1678. [DOI] [PubMed] [Google Scholar]
- Budden KF, Gellatly SL, Wood DLA, Cooper MA, Morrison M, Hugenholtz P, Hansbro PM, 2017. Emerging pathogenic links between microbiota and the gut-lung axis. Nat Rev Microbiol 15(1), 55–63. [DOI] [PubMed] [Google Scholar]
- Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Pena AG, Goodrich JK, Gordon JI, Huttley GA, Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, McDonald D, Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Turnbaugh PJ, Walters WA, Widmann J, Yatsunenko T, Zaneveld J, Knight R, 2010. QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7(5), 335–336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Huntley J, Fierer N, Owens SM, Betley J, Fraser L, Bauer M, Gormley N, Gilbert JA, Smith G, Knight R, 2012. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. The ISME journal 6(8), 1621–1624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins SM, Surette M, Bercik P, 2012. The interplay between the intestinal microbiota and the brain. Nat Rev Microbiol 10(11), 735–742. [DOI] [PubMed] [Google Scholar]
- Cryan JF, Dinan TG, 2012. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nature reviews. Neuroscience 13(10), 701–712. [DOI] [PubMed] [Google Scholar]
- D’Agostino RB Sr., Vasan RS, Pencina MJ, Wolf PA, Cobain M, Massaro JM, Kannel WB, 2008. General cardiovascular risk profile for use in primary care: the Framingham Heart Study. Circulation 117(6), 743–753. [DOI] [PubMed] [Google Scholar]
- De Hert M, Dekker JM, Wood D, Kahl KG, Holt RI, Moller HJ, 2009. Cardiovascular disease and diabetes in people with severe mental illness position statement from the European Psychiatric Association (EPA), supported by the European Association for the Study of Diabetes (EASD) and the European Society of Cardiology (ESC). European psychiatry : the journal of the Association of European Psychiatrists 24(6), 412–424. [DOI] [PubMed] [Google Scholar]
- Earth Microbiome Project, 2016. DNA Extraction Protocol. [Google Scholar]
- Edgar RC, 2010. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26(19), 2460–2461. [DOI] [PubMed] [Google Scholar]
- Evans SJ, Bassis CM, Hein R, Assari S, Flowers SA, Kelly MB, Young VB, Ellingrod VE, McInnis MG, 2016. The gut microbiome composition associates with bipolar disorder and illness severity. Journal of psychiatric research 87, 23–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faith DP, 1992. Conservation Evaluation and Phylogenetic Diversity. Biol Conserv 61(1), 1–10. [Google Scholar]
- First M, Spitzer RL, Gibbon M, Williams JB, 2002. Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version, Patient Edition With Psychotic Screen (SCID-I/PW/PSY SCREEN) New York: Biometrics Research, New York State Psychiatric Institute; 2002. Biometrics Research, New York State Psychiatric Institute, New York. [Google Scholar]
- Flowers SA, Evans SJ, Ward KM, McInnis MG, Ellingrod VL, 2016. Interaction between Atypical Antipsychotics and the Gut Microbiome in a Bipolar Disease Cohort. Pharmacotherapy [Epub 30 December 2016]. [DOI] [PubMed] [Google Scholar]
- Graessler J, Qin Y, Zhong H, Zhang J, Licinio J, Wong ML, Xu A, Chavakis T, Bornstein AB, Ehrhart-Bornstein M, Lamounier-Zepter V, Lohmann T, Wolf T, Bornstein SR, 2013. Metagenomic sequencing of the human gut microbiome before and after bariatric surgery in obese patients with type 2 diabetes: correlation with inflammatory and metabolic parameters. Pharmacogenomics J 13(6), 514–522. [DOI] [PubMed] [Google Scholar]
- Hartstra AV, Bouter KEC, Backhed F, Nieuwdorp M, 2015. Insights Into the Role of the Microbiome in Obesity and Type 2 Diabetes. Diabetes Care 38(1), 159–165. [DOI] [PubMed] [Google Scholar]
- He Y, Kosciolek T, Tang J, Zhou Y, Li Z, Ma X, Zhu Q, Yuan N, Yuan L, Li C, Jin K, Knight R, Tsuang MT, Chen X, 2018. Gut microbiome and magnetic resonance spectroscopy study of subjects at ultra-high risk for psychosis may support the membrane hypothesis. European psychiatry : the journal of the Association of European Psychiatrists 53, 37–45. [DOI] [PubMed] [Google Scholar]
- Hennekens CH, Hennekens AR, Hollar D, Casey DE, 2005. Schizophrenia and increased risks of cardiovascular disease. American heart journal 150(6), 1115–1121. [DOI] [PubMed] [Google Scholar]
- Hong S, Lee EE, Martin AS, Soontornniyomkij B, Soontornniyomkij V, Achim CL, Reuter C, Irwin MR, Eyler LT, Jeste DV, 2017. Abnormalities in chemokine levels in schizophrenia and their clinical correlates. Schizophrenia research 181, 63–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang H, Ling Z, Zhang Y, Mao H, Ma Z, Yin Y, Wang W, Tang W, Tan Z, Shi J, Li L, Ruan B, 2015. Altered fecal microbiota composition in patients with major depressive disorder. Brain, behavior, and immunity 48, 186–194. [DOI] [PubMed] [Google Scholar]
- Jiang L, Amir A, Morton JT, Heller R, Arias-Castro E, Knight R, 2017. Discrete False-Discovery Rate Improves Identification of Differentially Abundant Microbes. mSystems 2(6). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joseph J, Depp C, Martin AS, Daly RE, Glorioso DK, Palmer BW, Jeste DV, 2015. Associations of high sensitivity C-reactive protein levels in schizophrenia and comparison groups. Schizophrenia research 168(1–2), 456–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamada N, Seo SU, Chen GY, Nunez G, 2013. Role of the gut microbiota in immunity and inflammatory disease. Nature reviews. Immunology 13(5), 321–335. [DOI] [PubMed] [Google Scholar]
- Kelly TN, Bazzano LA, Ajami NJ, He H, Zhao J, Petrosino JF, Correa A, He J, 2016. Gut Microbiome Associates With Lifetime Cardiovascular Disease Risk Profile Among Bogalusa Heart Study Participants. Circulation research 119(8), 956–964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kostic AD, Xavier RJ, Gevers D, 2014. The Microbiome in Inflammatory Bowel Disease: Current Status and the Future Ahead. Gastroenterology 146(6), 1489–1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroenke K, Spitzer RL, 2002. The PHQ-9: a new depression diagnostic and severity measure. Psychiatric Annals 32(9), 509–515. [Google Scholar]
- Lee EE, Eyler LT, Wolkowitz OM, Martin AS, Reuter C, Kraemer H, Jeste DV, 2016. Elevated plasma F2-isoprostane levels in schizophrenia. Schizophrenia research 176(2–3), 320–326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee EE, Hong S, Martin AS, Eyler LT, Jeste DV, 2017. Inflammation in Schizophrenia: Cytokine Levels and Their Relationships to Demographic and Clinical Variables. The American journal of geriatric psychiatry : official journal of the American Association for Geriatric Psychiatry 25(1), 50–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lozupone C, Knight R, 2005. UniFrac: a new phylogenetic method for comparing microbial communities. Applied and environmental microbiology 71(12), 8228–8235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lozupone CA, Hamady M, Kelley ST, Knight R, 2007. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Applied and environmental microbiology 73(5), 1576–1585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lozupone CA, Stombaugh J, Gonzalez A, Ackermann G, Wendel D, Vazquez-Baeza Y, Jansson JK, Gordon JI, Knight R, 2013. Meta-analyses of studies of the human microbiota. Genome research 23(10), 1704–1714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer EA, 2011. Gut feelings: the emerging biology of gut-brain communication. Nature reviews. Neuroscience 12(8), 453–466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald D, Hyde E, Debelius JW, Morton JT, Gonzalez A, Ackermann G, Aksenov AA, Behsaz B, Brennan C, Chen Y, DeRight Goldasich L, Dorrestein PC, Dunn RR, Fahimipour AK, Gaffney J, Gilbert JA, Gogul G, Green JL, Hugenholtz P, Humphrey G, Huttenhower C, Jackson MA, Janssen S, Jeste DV, Jiang L, Kelley ST, Knights D, Kosciolek T, Ladau J, Leach J, Marotz C, Meleshko D, Melnik AV, Metcalf JL, Mohimani H, Montassier E, Navas-Molina J, Nguyen TT, Peddada S, Pevzner P, Pollard KS, Rahnavard G, Robbins-Pianka A, Sangwan N, Shorenstein J, Smarr L, Song SJ, Spector T, Swafford AD, Thackray VG, Thompson LR, Tripathi A, Vazquez-Baeza Y, Vrbanac A, Wischmeyer P, Wolfe E, Zhu Q, American Gut C, Knight R, 2018. American Gut: an Open Platform for Citizen Science Microbiome Research. mSystems 3(3). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montross LP, Depp C, Daly J, Reichstadt J, Golshan S, Moore D, Sitzer D, Jeste DV, 2006. Correlates of self-rated successful aging among community-dwelling older adults. The American journal of geriatric psychiatry : official journal of the American Association for Geriatric Psychiatry 14(1), 43–51. [DOI] [PubMed] [Google Scholar]
- Nguyen TT, Kosciolek T, Eyler LT, Knight R, Jeste DV, 2018. Overview and Systematic Review of Studies of Microbiome in Schizophrenia and Bipolar Disorder. Journal of psychiatric research 88, 50–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Dwyer DN, Dickson RP, Moore BB, 2016. The Lung Microbiome, Immunity, and the Pathogenesis of Chronic Lung Disease. J Immunol 196(12), 4839–4847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Organization WH, 2009. Guideline for ATC classification and DDD assignment, 2010, WHO Collaborating Centre for Drug Statistics Methodology, Oslo, Norway. [Google Scholar]
- Parmelee PA, Thuras PD, Katz IR, Lawton MP, 1995. Validation of the Cumulative Illness Rating Scale in a geriatric residential population. Journal of the American Geriatrics Society 43(2), 130–137. [DOI] [PubMed] [Google Scholar]
- Round JL, Mazmanian SK, 2009. The gut microbiome shapes intestinal immune responses during health and disease. Nature reviews. Immunology 9(5), 313–323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saha S, Chant D, McGrath J, 2007. A systematic review of mortality in schizophrenia: is the differential mortality gap worsening over time? Archives of general psychiatry 64(10), 1123–1131. [DOI] [PubMed] [Google Scholar]
- Schwabe RF, Jobin C, 2013. The microbiome and cancer. Nat Rev Cancer 13(11), 800–812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwarz E, Maukonen J, Hyytiainen T, Kieseppa T, Oresic M, Sabunciyan S, Mantere O, Saarela M, Yolken R, Suvisaari J, 2018. Analysis of microbiota in first episode psychosis identifies preliminary associations with symptom severity and treatment response. Schizophrenia research 192, 398–403. [DOI] [PubMed] [Google Scholar]
- Severance EG, Alaedini A, Yang S, Halling M, Gressitt KL, Stallings CR, Origoni AE, Vaughan C, Khushalani S, Leweke FM, Dickerson FB, Yolken RH, 2012. Gastrointestinal inflammation and associated immune activation in schizophrenia. Schizophrenia research 138(1), 48–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Severance EG, Gressitt KL, Stallings CR, Katsafanas E, Schweinfurth LA, Savage CL, Adamos MB, Sweeney KM, Origoni AE, Khushalani S, Leweke FM, Dickerson FB, Yolken RH, 2016. Candida albicans exposures, sex specificity and cognitive deficits in schizophrenia and bipolar disorder. NPJ schizophrenia 2, 16018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Severance EG, Gressitt KL, Stallings CR, Origoni AE, Khushalani S, Leweke FM, Dickerson FB, Yolken RH, 2013. Discordant patterns of bacterial translocation markers and implications for innate immune imbalances in schizophrenia. Schizophrenia research 148(1–3), 130–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Severance EG, Prandovszky E, Castiglione J, Yolken RH, 2015. Gastroenterology issues in schizophrenia: why the gut matters. Current psychiatry reports 17(5), 27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shannon CE, 1948. The mathematical theory of communication. The Bell System Technical Journal 27, 379–423, 623–656. [Google Scholar]
- Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, Hergueta T, Baker R, Dunbar GC, 1998. The Mini-International Neuropsychiatric Interview (M.I.N.I.): The development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. Journal of Clinical Psychiatry 59, 22–33. [PubMed] [Google Scholar]
- Shen Y, Xu J, Li Z, Huang Y, Yuan Y, Wang J, Zhang M, Hu S, Liang Y, 2018. Analysis of gut microbiota diversity and auxiliary diagnosis as a biomarker in patients with schizophrenia: A cross-sectional study. Schizophrenia research. [DOI] [PubMed] [Google Scholar]
- Sørensen T, 1948. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biol. Skr. 5, 1–34. [Google Scholar]
- Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu XN, Kubo C, Koga Y, 2004. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. The Journal of physiology 558(Pt 1), 263–275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vazquez-Baeza Y, Pirrung M, Gonzalez A, Knight R, 2013. EMPeror: a tool for visualizing high-throughput microbial community data. Gigascience 2(1), 16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walters WA, Xu Z, Knight R, 2014. Meta-analyses of human gut microbes associated with obesity and IBD. FEBS letters 588(22), 4223–4233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ware JE Jr, Sherbourne CD, 1992. The MOS 36-item short-form health survey (SF-36): I. Conceptual framework and item selection. Medical care, 473–483. [PubMed] [Google Scholar]
- Wilson PW, D’Agostino RB, Levy D, Belanger AM, Silbershatz H, Kannel WB, 1998. Prediction of coronary heart disease using risk factor categories. Circulation 97(18), 1837–1847. [DOI] [PubMed] [Google Scholar]
- Wu J, Peters BA, Dominianni C, Zhang Y, Pei Z, Yang L, Ma Y, Purdue MP, Jacobs EJ, Gapstur SM, Li H, Alekseyenko AV, Hayes RB, Ahn J, 2016. Cigarette smoking and the oral microbiome in a large study of American adults. The ISME journal 10(10), 2435–2446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan X, Zhang P, Wang Y, Liu Y, Li X, Kumar BU, Hei G, Lv L, Huang XF, Fan X, Song X, 2018. Changes in metabolism and microbiota after 24-week risperidone treatment in drug naive, normal weight patients with first episode schizophrenia. Schizophrenia research. [DOI] [PubMed] [Google Scholar]
- Zheng P, Zeng B, Zhou C, Liu M, Fang Z, Xu X, Zeng L, Chen J, Fan S, Du X, Zhang X, Yang D, Yang Y, Meng H, Li W, Melgiri ND, Licinio J, Wei H, Xie P, 2016. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Molecular psychiatry 21(6), 786–796. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Gut microbial composition at the phylum level for schizophrenia and NC groups. Compared to NCs, patients with schizophrenia exhibited increased relative abundance of Proteobacteria.
Summary of differentially abundant taxa between groups.