

Abstract
Numerous observations suggest diverse and modulatory roles for serotonin (5-HT) in cortex. Because of the diversity of cell types and multiple receptor subtypes and actions of 5-HT, it has proven difficult to determine the overall role of 5-HT in cortical function. To provide a broader perspective of cellular actions, we studied the effects of 5-HT on morphologically and physiologically identified pyramidal and nonpyramidal neurons from layers I–III of primary somatosensory and motor cortex. We found cell type-specific differences in response to 5-HT. Four cell types were observed in layer I: Cajal Retzius, pia surface, vertical axon, and horizontal axon cells. The physiology of these cells ranged from fast spiking (FS) to regular spiking (RS). In layers II–III, we observed interneurons with FS, RS, and late spiking physiology. Morphologically, these cells varied from bipolar to multipolar and included basket-like and chandelier cells. 5-HT depolarized or hyperpolarized pyramidal neurons and reduced the slow afterhyperpolarization and spike frequency. Consistent with a role in facilitating tonic inhibition, 5-HT2 receptor activation increased the frequency of spontaneous IPSCs in pyramidal neurons. In layers II–III, 70% of interneurons were depolarized by 5-HT. In layer I, 57% of cells with axonal projections to layers II–III (vertical axon) were depolarized by 5-HT, whereas 63% of cells whose axons remain in layer I (horizontal axon) were hyperpolarized by 5-HT. We propose a functional segregation of 5-HT effects on cortical information processing, based on the pattern of axonal arborization.
Keywords: 5-HT, interneuron, potassium, cortex, pyramidal cell, biocytin
Normal fluctuations in cortical states, mood, attention, and information processing rely on activation of neocortical serotonin (5-HT) receptors (Waterhouse et al. 1986b;Jacobs et al., 1990; Reuter and Jacobs, 1996; Marek and Aghajanian, 1998). Altered serotonergic transmission is implicated in anxiety and depression, seizures, action of psychotropic hallucinogens, and schizophrenia (Statnick et al., 1996; Marek and Aghajanian, 1998). Frontoparietal cortex receives serotonergic projections from the dorsal and median raphe nuclei (Waterhouse et al. 1986a; Tork, 1990). More than one dozen 5-HT receptor types have been described (Hoyer et al., 1994), several of which are expressed in frontoparietal cortex (Bruinvels et al., 1994; Pompeiano et al., 1994; Burnet et al., 1995;Wright et al., 1995; Morales and Bloom, 1997; Vysokanov et al., 1998). With the exception of the 5-HT3 receptor (ligand-gated ion channel), 5-HT receptors couple to G-proteins to exert their effects (Hoyer et al., 1994). These observations suggest diverse modulatory roles for 5-HT in cortex.
In vivo, firing rates of pyramidal neurons are generally decreased by iontophoresis of 5-HT (Krnjevic and Phillis, 1963; Roberts and Straughan, 1967; Jordan et al., 1972; Reader et al., 1979; Lakoski and Aghajanian, 1985). 5-HT also reduced the responses of somatosensory cortical neurons to afferent input (Waterhouse et al. 1986b) and altered memory fields in prefrontal cortex (Williams et al., 2002).
Previous in vitro studies of the actions of 5-HT in cortex concentrated on layer V pyramidal neurons. In rat cortex, these cells were hyperpolarized or depolarized by 5-HT, with most cells exhibiting both responses (Davies et al., 1987; Araneda and Andrade, 1991; Tanaka and North, 1993; Spain, 1994; Marek and Aghajanian, 1998). The hyperpolarizations involved activation of 5-HT1Areceptors and increased potassium conductance (GK). Depolarizations were reported to involve 5-HT2A receptors and a decrease inGK. 5-HT also reduced the slow afterhyperpolarization (sAHP) (Araneda and Andrade, 1991; Tanaka and North, 1993) and induced an afterdepolarization (ADP) (Araneda and Andrade, 1991). In cat sensorimotor cortex, the 5-HT response correlated with pyramidal cell firing type (Spain, 1994).
Understanding serotonergic effects on cortical function requires knowledge of the actions of 5-HT on other cell types in cortical local circuits. GABAergic interneurons in the neocortex are morphologically and physiologically diverse (White, 1989; Kawaguchi, 1993, 1995; Cauli et al., 1997; Kawaguchi and Kubota, 1997, 1998), and GABAergic inhibition has been proposed to be a target of serotonergic modulation (DeFelipe et al., 1991; Smiley and Goldman-Rakic, 1996; Abi-Saab et al., 1999). Little is known, however, about the responses of specific interneuron types to 5-HT. A few layer V nonpyramidal cells were included in the Tanaka and North (1993) study, and a preliminary report suggests 5-HT3 responses in neocortical layer V interneurons (Xiang et al., 1999). Altered tuning of memory fields was reported for thin spiking neurons (putative interneurons) in primate prefrontal cortex (Williams et al., 2002). 5-HT2A receptor activation increased the frequency of spontaneous EPSCs (sEPSCs) and sIPSCs on both pyramidal cells and layer I interneurons (Zhou and Hablitz, 1999b). Effects of other transmitters on cortical interneurons are cell type specific (Kawaguchi and Shindou, 1998; Para et al., 1998; Xiang et al., 1999; Zhou and Hablitz, 1999a).
We examined serotonergic effects on identified interneurons and pyramidal cells from rat sensorimotor cortex (layers I–III). Our major finding was that 5-HT had different effects on interneurons, and these differences corresponded to laminar differences in axonal projections.
Some of these data have been presented previously in abstract form (Foehring et al., 1996).
MATERIALS AND METHODS
Sprague Dawley rats [postnatal day (P) 6–24; most were P18–23] were deeply anesthetized with an intraperitoneal injection (1.8 ml/kg) of a mixture of 1:3 xylazine/ketamine dissolved in an equal volume of saline. These ages correspond to the peak expression of 5-HT2A receptors in cortex (approximately P13) (Roth et al., 1991). The density of serotonergic fibers (Foote and Morrison, 1984; Nakazawa et al., 1992), 5-HT1A receptors (Zifa et al., 1988), and 5-HT levels (Edagawa et al., 2001) continue to increase with age, at least through 5 weeks postnatally. 5-HT7receptors are expressed in low abundance in this cortical region (Gustafson et al., 1996), with developmental regulation of receptor abundance (Vizuete et al., 1997). It is possible that changes in relative abundance of receptor types could alter the effects of 5-HT at other ages. We therefore restrict our conclusions to ∼2–3 weeks postnatally, an especially important time for generation of new synapses in neocortex (Armstrong-James and Johnson, 1970; Wise and Jones, 1976).
Once the animals were areflexic, a shallow ventral incision severed the carotid arteries. The skull was opened dorsally, and the brain was exposed. The dura was reflected, and cold artificial CSF (ACSF) (see below) was applied to the brain surface while four scalpel cuts isolated the left somatosensory and motor cortex (sensorimotor cortex) and underlying forebrain. The isolated block of tissue was then lifted clear of the brain with cottonoid and glued to the stage of a modifiedVibratome (Pelco 101 series 1000; Ted Pella, Inc., Redding, CA). The tissue was immersed in cutting solution (see below) at 4°C, and coronal sections were cut at 300 μm. The slices were maintained in ACSF at 35°C for 1 hr and thereafter stored at room temperature.
Solutions. All chemicals were obtained from Sigma (St. Louis, MO), unless noted otherwise. The cutting solution contained (in mm): KCl 5, NaH2PO4 1.25, NaHCO3 26, MgCl2 5, TEA-Cl 20, Choline-Cl 105, sucrose 20, and dextrose 10 [320 mOsm/l, pH maintained at 7.4 by bubbling with carbogen (95% O2/5% CO2)]. ACSF contained (in mm): NaCl 125, KCl, 3, NaH2PO4 1.25, NaHCO3 26, dextrose 20, MgCl2 2, and CaCl2 2 (310 mOsm/l). The pH was maintained at 7.4 by bubbling with carbogen. For whole-cell recordings, the internal recording solution contained (in mm): KCH3SO4 135, MgCl2 2, KCl 5, HEPES 10, ATP 2, Na3GTP 0.5, biocytin (0.5%), and EGTA 0.1, pH 7.2 (270 mOsm/l). For recording spontaneous IPSCs, CsCH3SO4 replaced KMeSO4 in the internal solution 5-HT and serotonergic agonists and antagonists were obtained from RBI. They were prepared in d,d H2O and frozen in aliquots until use. Agonists were only tested in cells with stable membrane potential for >2 min in control solution. Na-metabisulfite was included (final concentration 50–100 μm) with 5-HT and agonists to prevent oxidation (Sutor and ten Bruggencate, 1990). Na-metabisulfite caused no effects on its own (n = 4). Solutions were held in separate bottles at 35°C. A manifold and valves were used to control which solution flowed into the common line to the recording chamber.
Recordings. A single slice was transferred to a recording chamber on the fixed stage of a Zeiss Axioscope. Slices were submerged and perfused continuously in ACSF (∼2 ml/min). Cortical layers were located under low power, and then a 40× water immersion lens with differential interference contrast (DIC) optics, and near infrared (bandpass 750–800 nm) illumination was used to identify individual neurons. The image was projected onto a video camera sensitive to infrared light and displayed on a video monitor (Dodt and Zieglgansberger, 1994). Solutions were bubbled with carbogen and preheated to 37°C in a water bath. The solutions were fed by gravity to the recording chamber. An in-line heater and temperature probe were used to regulate bath temperature at 32 ± 2°C. In most cases, 5-HT and agonists were applied in the bath solution (10–60 μm). In a few cases, drugs were applied directly to visualized neurons using pressure (Picospritzer; General Valve, Fairfield, NJ). Antagonists were added in the bath for 5–10 min before application of agonist plus antagonist.
Whole-cell electrodes were fabricated from VWR (VWR Scientific Products, Westchester, PA) hematocrit glass (4–8 MΩ) with a two-stage pull on a List vertical puller. Electrodes were not polished or coated for current-clamp recordings. We coated the electrodes with Sylgard (Dow Corning Corporation, Midland, MI) for recording of IPSCs. Recordings were made in current clamp (Axoclamp 2A, bridge mode; Axon Instruments, Union City, CA) or voltage clamp (Axopatch 1C or 200B). The electrodes were visually guided onto the cell bodies while positive pressure was being applied. On cell contact, negative pressure was applied until a tight seal (>1 GΩ) was formed. Further negative pressure was used to attain the whole-cell configuration. Whole-cell current and membrane potential were filtered (<3 kHz), amplified, and recorded on a video recorder with pulse code modulation (Neurocorder, 44 kHz sampling rate; Neurodata Instrument Corp., New York, NY) for later analysis or digitized on-line with pClamp 6 (Axon Instruments). A chart recorder was used to monitor slow changes in membrane currents and potentials. Data were analyzed off-line using Axograph (Axon Instruments). Voltage records for current clamp were corrected for the liquid junction potential [determined using the method of Neher (1992)], which was 10 mV. Series resistance was typically 10–20 MΩ. Cells were eliminated if series resistance increased greatly during the recording.
Spontaneous synaptic currents were detected and analyzed as described previously (van Brederode et al., 2001). Current segments (1–2 min) were scanned for synaptic events using a threshold detection method (Mini Analysis, Synaptosoft, Decatur, GA).
Morphology. Neurons were filled with Biocytin during the recording and identified as interneurons or pyramidal cells on the basis of morphology. After recording, the electrode was slowly retracted from the cell. The slice was then placed in 4% paraformaldehyde for 24 hr. We typically recorded one cell per slice, or if a second cell was recorded from, it was chosen to be in a different layer and location in the slice. The fixed slices were then washed in PBS and then processed according to the avidin–biotin method (Vectastain) of Horikawa and Armstrong (1988). Biocytin was visualized using DAB/peroxidase. Filled cells were drawn under high power (40 or 100× oil immersion lens) with the aid of a drawing tube on a microscope. These drawings were later traced in ink, transferred to acetate, and photographed. Selected cells were drawn in detail using the Eutectics system (Eutectics Corp.). The cell dimensions were not corrected for tissue shrinkage or two-dimensional projection errors.
RESULTS
We recorded from 106 cells (52 pyramidal and 54 nonpyramidal) that were tested with bath-applied 5-HT and that met our criteria of overshooting action potentials (APs) and stable resting potential (negative to −50 mV) and input resistance (current-clamp recordings). Additional cells were examined with direct pressure application of 5-HT or 5-HT3 agonist (see below). All of the recordings were obtained with whole-cell patch clamp of cells visualized with infrared illumination and DIC optics (Stuart et al., 1993; Dodt and Zieglgansberger, 1994) in the supragranular layers of sensorimotor cortex. Data are presented as mean ± SEM, unless noted otherwise. Table 1 lists the nonpyramidal neurons included in the analyses, including cortical layer, age of animal, and morphological and physiological classification (see below). Table 2contains representative data from 42 interneurons (21 from layer I, 21 from layer II) for which complete physiological data were obtained.
Table 1.
List of interneurons (bath-applied 5-HT)
| Cell | Age | Layer | Morph type | Phys type | 5-HT effect |
|---|---|---|---|---|---|
| 121096h11 | 14 | I | Horizontal | FS | −5 |
| 92795u1 | 17 | I | Horizontal | FS | −9 |
| 92795u2 | 17 | I | Horizontal | FS | −5 |
| 92795u3 | 17 | I | Horizontal | FS | −5 |
| 63098u2 | 17 | I | Horizontal | FS | 1 |
| 7298u2 | 19 | I | Horizontal | FS | 2 |
| 72597u2 | 18 | I | Horizontal | LS | 0 |
| 92895u1 | 16 | I | Horizontal | RS | −4 |
| 1211h11 | 14 | I | Horizontal | −2 | |
| 121196u3 | 14 | I | Horizontal | 2 | |
| 63098u1 | 17 | I | Horizontal | −12 | |
| 72397u2 | 17 | I | Horizontal | −5 | |
| 72397u1 | 17 | I | Horizontal | 0 | |
| 72397h11 | 17 | I | Vertical | FS | 0 |
| 72597h3 | 18 | I | Vertical | FS | 10 |
| 62698u3 | 18 | I | Vertical | FS | 5 |
| 93095u5 | 13 | I | Vertical | RS | 10 |
| 62598u1 | 17 | I | Vertical | RS | 121-a |
| 61096u1 | 17 | I | Vertical | 0 | |
| 72597u1 | 18 | I | Vertical | −2 | |
| 92895u2 | 16 | I | Neuroglia | FS | −5 |
| 62498u1 | 16 | I | Neuroglia | FS | 3 |
| 725975H | 18 | I | Neuroglia | FS | −4 |
| 71398u3 | 14 | I | Pia surface | FS | 9 |
| 62698u2 | 18 | I | Pia surface | FS | 2 |
| 7298u1 | 19 | I | Pia surface | FS | 3 |
| 92395u1 | 10 | I | Pia surface | RS | 5 |
| 92395u2 | 8 | I | CR | 0 | |
| 72297u1 | 16 | I | CR | −1 | |
| 72297u2 | 16 | I | CR | −2 | |
| 72497u2 | 18 | I | CR | −2 | |
| 92795u4 | 17 | II | Multipolar | FS | 4 |
| 61296u2 | 18 | II | Multipolar | FS | 1 |
| 61296u4 | 18 | II | Multipolar | FS | 4 |
| 7298u3 | 19 | II | Multipolar | FS | 3 |
| 61296u3 | 18 | II | Multipolar | RS | 141-a |
| 12196h11 | 15 | II | Basket | FS | −3 |
| 121296h21 | 17 | II | Basket | FS | −3 |
| 7198u2 | 18 | II | Basket | FS | 3 |
| 7198u3 | 18 | II | Basket | FS | 0 |
| 71498u2 | 15 | II | Basket | LS | −1 |
| 121096h31 | 14 | II | Bipolar | FS | 5 |
| 61496u2 | 20 | II | Bipolar | FS | 5 |
| 121196u2 | 14 | II | Bipolar | RS | 5 |
| 625982h | 18 | II | Chandelier | FS | 0 |
| 61396u2 | 20 | II | FS | 111-a | |
| 61496u3 | 20 | III | Multipolar | FS | 151-a |
| 121396u2 | 18 | III | Basket | LS | 10 |
| 61196u2 | 17 | III | Bipolar | RS | 101-a |
| 121096u3 | 13 | III | FS | 0 | |
| 121096u5 | 13 | III | FS | 101-a | |
| 121296u3 | 15 | III | FS | 5 | |
| 121096u3 | 15 | III | FS | 0 | |
| 61396u3 | 20 | III | FS | 9 |
Layer, Cortical layer; Morph type, classification by morphological cell type; Phys type, classification by physiological cell type; 5-HT effect, change in membrane potential (mV; minus sign indicates hyperpolarization); Age, age of animal in postnatal days (birth is P0).
Depolarized to firing.
Table 2.
Physiological properties
| Type | RMP | AP | HW | RN |
|---|---|---|---|---|
| Interneurons | ||||
| Layer I | ||||
| Horizontal | −68 ± 1 (7) | 69 ± 3 (8) | 0.8 ± 0.1 (8) | 237 ± 42 (8) |
| Vertical | −70 ± 6 (6) | 65 ± 5 (6) | 0.9 ± 0.2 (6) | 435 ± 77 (6) |
| All | −69 ± 1 (21) | 69 ± 3 (22) | 0.9 ± 0.1 (23) | 347 ± 34 (21) |
| Layers II–III | ||||
| FS/LS | −74 ± 2 (15) | 75 ± 4 (15) | 0.8 ± 0.1 (14) | 327 ± 62 (15) |
| All | −74 ± 2 (21) | 75 ± 4 (21) | 0.8 ± 0.1 (21) | 330 ± 58 (21) |
| Pyramidal | ||||
| Layer II | −71 ± 2 (22) | 90 ± 3 (22) | 1.2 ± 0.1 (22) | 254 ± 23 (22) |
| Layer III | −71 ± 2 (6) | 92 ± 4 (6) | 1.2 ± 0.1 (6) | 195 ± 53 (6) |
| All | −71 ± 2 (28) | 90 ± 2 (28) | 1.2 ± 0.1 (28) | 241 ± 21 (28) |
RMP, Resting membrane potential (mV); AP, action potential amplitude (from RMP; mV); HW, action potential width at one-half of maximum amplitude (msec); RN, input resistance (MΩ). This was calculated from Ohm's law for peak voltage deflection caused by hyperpolarizing current. Currents were chosen for final voltage changes of 10–20 mV.
Our goal was to determine the overall, dominant effect of 5-HT on different cell types. Unless stated otherwise, all data presented below for pyramidal and nonpyramidal neurons reflect bath application of transmitter or agonists. Both hyperpolarizations and depolarizations were fully reversible if the agonists were applied for more than ∼1 min (see Fig. 2C2,E) and recovery often required long (>10 min) washes in control solutions (n = 25 cells).
Fig. 2.
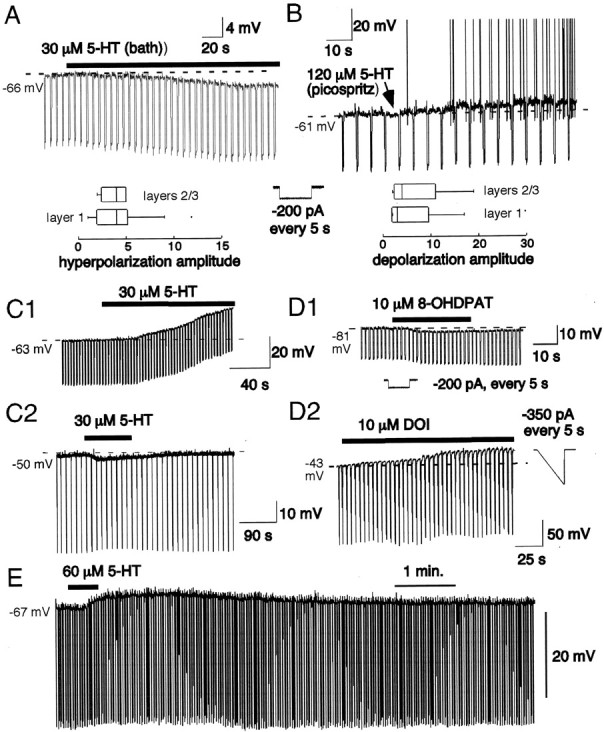
Membrane potential responses of interneurons to 5-HT. Time of agonist application is indicated bysolid bar above traces. A, Bath application of 5-HT hyperpolarized a subpopulation of supragranular interneurons. The dotted line indicates the original membrane potential (−66 mV). This cell (layer I horizontal axon cell) was hyperpolarized 2.7 mV by 5-HT. The negative deflections are responses to a 500 msec input conductance stimulus (−200 pA; inset). The box plots (Tukey, 1977) indicate the median values (vertical line withinbox) and inner quartile (box margin) and outer quartile (horizontal lines extending frombox) of the responses for layers II (n = 14 cells) and II–III (n = 3). B, Other interneurons were depolarized by 5-HT. This cell (layer I, vertical axon) was depolarized to spiking. More typically, depolarizations did not reach spike threshold. The box plots summarize the data for response amplitude by layer (layer I:n = 12 cells; layers II–III: n= 16 cells). C, The membrane potential responses to 5-HT were retained in the presence of 1 μm TTX.C1, 5-HT was added to the bath. This layer II nonpyramidal cell was depolarized by 15 mV. Downward deflections represent response to current ramps (−350 pA, 500 msec).C2, This basket-like cell (layer II) was hyperpolarized ∼3 mV by 5-HT in the presence of TTX. Downward deflections represent response to current ramps (−400 pA, 500 msec) repeated at 5 sec intervals. Note reversal of effect in control wash. D, Receptor pharmacology. D1, The hyperpolarization was mimicked by bath application of the 5-HT1A receptor agonist 10 μm 8-OHDPAT in this layer II FS cell. Dotted line indicates original resting potential. Downward deflections represent response to current steps (−300 pA; seeinset). D2, The application of the 5-HT2A/2C receptor agonist 30 μm DOI led to a 15 mV depolarization in this layer I FS cell (pia surface cell; 1 μm TTX present). Downward deflections represent response to current ramps (−350 pA, 500 msec). E, Depolarizing effects of 5-HT were fully reversible. In this layers II–III interneuron, 5-HT was applied in the bath for 30 sec. The cell depolarized by 7 mV, and this depolarization was reversed after wash in control solution.
Our bath application data are biased against rapidly desensitizing responses (cf. 5-HT3 receptor mediated). We also used direct pressure application (Picospritzer) of 30 μm5-HT to 13 cells (7 pyramidal, 6 interneurons). Of these cells, three were depolarized, four hyperpolarized, and six showed no response to 5-HT. Multiple 5-HT receptor types are likely colocalized on individual cells (Martin-Ruiz et al., 2001), and dominance of a particular response does not rule out expression of other receptor subtypes (Araneda and Andrade, 1991).
Pyramidal neurons
To determine whether responses of supragranular pyramidal neurons to 5-HT were similar to those reported for layer V pyramidal neurons (Davies et al., 1987; Araneda and Andrade, 1991; Tanaka and North, 1993), we examined the effects of 5-HT on layers II–III pyramidal neurons (n = 38 cells) (Fig.1). In superficial layers II–III (near I–II border; layer II), 14 of 28 (50%) cells were depolarized by 5-HT (Fig. 1A). The other 14 cells (50%) were hyperpolarized (Fig. 1B). In deep layers II–III (layer III), 9 of 10 cells (90%) were depolarized by 5-HT, and 1 cell was hyperpolarized (Table 3). Many (18 of 28) of the layer II pyramidal cells had atypical orientation of the apical dendrite: the dendrite was horizontal, oblique, or even inverted (van Brederode et al., 2000). Of these 18 cells, 9 were depolarized and 9 were hyperpolarized by 5-HT.
Fig. 1.

Response of layers II–III pyramidal neurons to 5-HT. Time of 5-HT application is indicated by solid barabove traces. A, Twenty-three of 38 pyramidal cells were depolarized by bath application of 50 μm 5-HT. Downward deflections represent response to current ramps repeated at 5 sec intervals (see inset, below). In this cell, input conductance decreased in 5-HT. This cell was from layer III and was depolarized to firing. B, Other pyramidal cells were hyperpolarized by 5-HT (15 of 38). This cell was from layer II. Downward deflections are response to 500 msec, 200 pA current injections. In this cell, the peak hyperpolarization was 2 mV. C, Firing of layer II inverted pyramidal neuron (van Brederode et al., 2000) in response to 1 sec, 1.3 nA current injection. Note prominent spike frequency adaptation. D, Firing of the same cell as in B, except in the presence of 60 μm 5-HT (bath application). The membrane potential was manually returned to the original resting potential by DC injection. In 5-HT, the cell fired more action potentials, although the current stimulus was reduced (1.2 nA). In addition, 5-HT reduced the sAHP.E, The top trace is the response of a horizontally oriented pyramidal neuron from layer II (van Brederode et al., 2000) to a 500 msec, 145 pA current injection. The bottom trace shows the response of the same cell in the presence of bath-applied 30 μm 5-HT to a 500 msec, 92 pA current injection. The membrane potential was manually returned to the original resting potential by DC injection. The current stimuli were chosen to elicit the same numbers of spikes (10) in both control and 5-HT solutions. Note the different patterns of firing, with marked spike frequency adaptation in control solution and reduced adaptation in 5-HT. F, Same cell and traces as inE, with expanded time base and superimposition oftraces to show the reduction in the sAHP by 5-HT.
Table 3.
Membrane potential responses (bath-applied 5-HT)
| Type | Number depolarized | Number hyperpolarized | Number no change | n |
|---|---|---|---|---|
| All interneurons3-a | 28 (7 ± 2) | 18 (4 ± 1) | 8 | 54 |
| Layer I interneurons | ||||
| Cajal Retzius | 0 | 3 (2 ± 1) | 1 | 4 |
| Pia surface | 4 (5 ± 3) | 0 | 0 | 4 |
| Horizontal axon | 4 (3 ± 1)3-150 | 10 (6 ± 1)3-159 | 2 | 16 |
| Vertical axon | 4 (14 ± 6)3-150 | 1 (2 mV)3-159 | 2 | 7 |
| All | 12 (6 ± 1) | 14 (5 ± 1) | 5 | 31 |
| Layers II–III interneurons | ||||
| FS/LS3-b | 14 (9 ± 2) | 4 (2 ± 1) | 3 | 21 |
| RS | 2 (10 ± 7) | 0 | 0 | 2 |
| All | 16 (9 ± 2) | 4 (2 ± 1) | 3 | 23 |
| Pyramidal | ||||
| Layer II | 14 (7 ± 1) | 14 (5 ± 2) | 0 | 28 |
| Layer III | 9 (4 ± 1) | 1 (3 mV) | 0 | 10 |
Data in parentheses are mean ± SEM (in mV).
F3-150: ,
F3-159: , These two means are significantly different at p < 0.05 (t test).
All interneurons includes nonpyramidal cells from layers I–III.
Includes cells with FS physiology but no morphology.
Although not studied in detail, the sAHP was significantly reduced (from 4.5 ± 2 to 1.6 ± 2 mV; 10–100% reduction) by 5-HT in 12 of 12 layers II–III cells tested (Figs.1E,F), with associated reduction in spike frequency adaptation (Fig. 1, compare C,D) (n = 12) (Araneda and Andrade, 1991;Spain, 1994). The sAHPs were elicited by repetitive firing in response to 500 msec constant current injections. Currents were adjusted to match numbers of spikes (3–21) and DC current injection was used to match resting potential, between 5-HT and control solutions. The sAHPs were measured at 500 msec after firing (after the mAHP has fully decayed) (Lorenzon and Foehring, 1993). In two cells, an ADP was induced by bath-applied 30 μm 5-HT (data not shown) (Araneda and Andrade, 1991; Spain, 1994).
Nonpyramidal neurons
Stable recordings and 5-HT application were obtained for 54 cells from layers I–III that were later identified by morphology as nonpyramidal. Overall, 28 of 54 nonpyramidal cells (52%) were depolarized by 5-HT (Fig. 2, Table 3). Eight cells (15%) showed no response to 5-HT, and 18 of 54 cells (33%) were hyperpolarized (Fig. 2, Table 3).
Neocortical nonpyramidal cells are highly diverse with respect to morphology, firing behavior, and expression of peptides and calcium binding proteins (White, 1989; Kawaguchi and Kubota 1996, 1997, 1998;Cauli et al., 1997). We therefore separated the data by layer and cell type. For each layer, we classified cells by axonal and dendritic morphology. Secondary classification was on the basis of the combination of firing pattern (amount of spike frequency adaptation, regularity of firing to near rheobasic currents), AP width, and input resistance.
Layer I receives a dense serotonergic projection (Mulligan and Tork, 1988; DeFelipe et al., 1991; Hornung and Celio, 1992; Smiley and Goldman-Rakic, 1996). We recorded from 31 morphologically identified interneurons in layer I (see Fig. 4, Table 1). Twelve of these cells (39%) were depolarized by 5-HT. Five cells (16%) showed no response to 5-HT, and 14 cells (45%) were hyperpolarized (Table 2). We recorded from 23 cells in layers II–III (Table 3) that were tested with 5-HT and (1) could either be classified on the basis of morphology into a particular cell type or (2) were clearly not pyramidal in morphology and showed fast spiking (FS) or late spiking (LS) physiology (McCormick et al., 1985; Cauli et al., 1997; Kawaguchi and Kubota 1997, 1998) (see Figs. 4, 5). Of these 23 cells, 16 (70%) were depolarized by 5-HT. Three cells did not respond to 5-HT (13%), and four were hyperpolarized (17%) (Fig. 2, Table 3).
Fig. 4.

Camera lucida reconstruction of biocytin-filled (Horikawa and Armstrong, 1988) layer I interneurons. A, Vertical axon cell (soma, axon, dendrites). The soma and dendrites (no axon) of this cell are shown separately on the right. The pia (solid line) and layers I–II border (broken line) are indicated on the left. The dendritic tree branches throughout the entire thickness of layer I, whereas most axonal branches are confined to layers II–III and do not enter layer I. Although this cell was located at the I–II border, vertical axon cells were located at all depths of layer I.B, Horizontal axon cell. The axons originated from two opposite poles of the cell body from a short trunk. The precise transition between dendrites and axons was difficult to discern. Note that the axonal and dendritic branches are primarily confined to thetop half of layer I. C, Cajal Retzius neuron (P9 animal). The soma and dendrites of this cell are shown at higher magnification (2×) on the right. Note the thick dendrites and that the axonal branches were mostly confined to the top half of layer I. The cells inB and C were drawn with a 100× objective, whereas the cell in A was drawn with a 40× objective. Scale bars, 100 μm (and apply to the reconstruction of axons). The drawings of the dendritic trees on the rightwere enlarged 2× from the drawings on the left.
Fig. 5.

Morphological types of interneurons in layers II and III. All cells were filled with biocytin during recording and processed with modifications of Horikawa and Armstrong (1988). The cells were then drawn with the aid of a camera lucida, copied onto acetate, and digitized with a scanner. A, Layer II basket-like cell. Note long vertical axonal projection as well as the horizontal one. Multipolar soma and dendrites are shown without the axon at right. B, Neurogliaform cell in layer II. Note extensive local processes. C, A chandelier cell in layer II. This cell did not respond to 5-HT. Note extensive vertical cassettes. Soma and dendrites shown in isolation below. All of the cells were drawn with a 100× objective. Scale bars:A, left, 100 μm; A,dendritic inset, 25 μm; B, 50 μm;C, 100 μm; C, dendritic inset, 25 μm.
Pharmacology
Both hyperpolarizations (n = 6; 4 pyramidal, 2 nonpyramidal) and depolarizations (n = 8 pyramidal, 4 nonpyramidal) were observed in the presence of TTX, suggesting that the effects were not dependent on presynaptic or postsynaptic action potentials (Fig. 2C). We also tested the response to 30 μm 5-HT in several cells in the presence of 100 μm picrotoxin, 50 μmAP5, and 10 μm CNQX to block transmission via GABAA, NMDA, and AMPA receptors, respectively (data not shown). We observed depolarizations (two of four pyramidal and four of six nonpyramidal cells) and hyperpolarizations (two of four pyramidal and two of six nonpyramidal cells) in the presence of these blockers, suggesting that the membrane potential effects were direct on the cells recorded from rather than caused by altered synaptic input.
Although we did not quantify dose–response relationships for the responses to 5-HT, we tested two or more doses in seven cells. For depolarizations (n = 3) and hyperpolarizations (n = 4), the higher dose always caused a greater response (data not shown). Repeated application of the same dose of agonist resulted in similar response to 5-HT (five depolarization, three hyperpolarization), 2,5-dimethoxy-4-iodoamphetamine hydrochloride (DOI) (see below; four cells), and 8-hydroxy-2-(di-n-propylamino)tetralin (8-OHDPAT) (two cells).
Receptors
Hyperpolarizations
To gain insight into which receptor types were responsible for the membrane potential changes in response to 5-HT, we tested responses to specific 5-HT receptor agonists and antagonists. On the basis of previous work in neocortex (Davies et al., 1987; Araneda and Andrade, 1991; Tanaka and North 1993; Spain, 1994) and other cell types (Andrade and Nicoll, 1987; Collino and Halliwell, 1987), we first examined responses to the bath application of 10 μm of the 5-HT1A/5-HT7 agonist 8-OHDPAT (Fig. 2D1). In layer I cells, 8-OHDPAT (10 μm) hyperpolarized six of eight cells tested (3 ± 2 mV; n = 6). Two horizontal axon cells showed no response. In layer II, five interneurons were tested with 8-OHDPAT; two cells were hyperpolarized, and three cells did not respond. 8-OHDPAT also hyperpolarized four of six layers II–III pyramidal cells (4 ± 1 mV; data not shown; two cells did not respond).
5-HT elicited hyperpolarizations in three of three interneurons and three of three pyramidal neurons tested in the presence of the 5-HT2/5-HT7 antagonists ritanserin or ketanserin (2 μm; data not shown). Combined with the 8-OHDPAT data, these data suggest that the hyperpolarizations are mediated by 5-HT1A receptors, although further study with more specific agonists/antagonists will be required.
Depolarizations
On the basis of receptor localization studies (Pompeiano et al., 1994; Wright et al., 1995; Morales et al., 1996; Morales and Bloom, 1997; Jakab and Goldman-Rakic, 1998) and physiology in various cell types (Lakoski and Aghajanian, 1985; Davies et al., 1987; Jackson and White, 1990; Araneda and Andrade, 1991; Bobker, 1994; Gellman and Aghajanian, 1994; Kawa, 1994; Spain, 1994; McMahon and Kauer, 1997;Roerig et al., 1997; Marek and Aghajanian, 1998), we expected the depolarizations to be mediated by 5-HT2 or 5-HT3 receptors [5-HT7receptors are another possibility (Gustafson et al., 1996)]. Because 5-HT3 receptor-mediated events are reported to rapidly desensitize (Jackson and Yakel, 1995), we thought it unlikely that the depolarizations that we observed after bath application of 5-HT would be 5-HT3 mediated. Consistent with this hypothesis, bath application of the 5-HT3agonist 1-(m-chlorophenyl) methyl biguanide (biguanide; 10–30 μm) caused no response in both interneurons tested (data not shown). Pressure application of 30 μm biguanide to 3 interneurons and 10 pyramidal cells elicited no membrane potential response in 12 cells; 1 interneuron was depolarized (data not shown).Zhou and Hablitz (1999b) reported inward currents in response to local application of 5-HT3 agonists in just 4 of 43 layer I cells. In contrast, bath application of the specific 5-HT2A/5-HT2C agonist DOI (10–30 μm) (Hoyer et al., 1994) caused a depolarization in four of four layer I interneurons tested (5 ± 2 mV) and two of four layer II cells (two cells showed no response) (Fig.2D2). The specific 5-HT2A/2C/7antagonist ketanserin prevented the depolarization by 5-HT in three of four interneurons tested (data not shown). Ketanserin alone had no effect on membrane potential (n = 4). Zhou and Hablitz (1999b) did not observe inward currents in layer I (n = 20) or layer II (n = 3) cells in response to 5-HT2 agonists.
DOI (10–30 μm) depolarized four of seven layers II–III pyramidal cells (5 ± 2 mV). Two cells did not respond, and one cell was hyperpolarized (data not shown). Ketanserin prevented the depolarization in three of four pyramidal cells (data not shown). Ketanserin alone had no effect on membrane potential (n= 4). Together, these data suggest that at least some of the depolarizations in both pyramidal and nonpyramidal cells were mediated by 5-HT2A or 5-HT2Creceptors (Davies et al., 1987; cf. Araneda and Andrade, 1991). Further study with more specific agonists and antagonists is required to test for potential roles of other receptor subtypes (e.g., 5-HT3, 5-HT4, 5-HT7) (Beique and Andrade, 2001).
Ionic basis
Hyperpolarization
In layer V pyramidal neurons, the 5-HT1A-mediated hyperpolarization was found to be caused by an increase in K+ conductance (Spain 1994). On average, we found that the 5-HT-mediated hyperpolarization in interneurons (measured at the original resting potential) was associated with no significant change in input resistance (control: 231 ± 38 MΩ; 5-HT: 277 ± 40 MΩ;n = 10). Input resistance was calculated from Ohm's law (RN =V/I). The current injection (500 msec step) was adjusted to elicit a 10–20 mV voltage response.
We tested the reversal potential of the hyperpolarization in six interneurons (Fig. 3) (in the presence of 1 μm TTX). With 3 mm extracellular K+, the reversal potential was −86 ± 4 mV (Fig. 3A). At 32°C and with 140 mm internal K+, the Nernst potential for 3 mm extracellular K+ is approximately −100 mV. For 9 mm extracellular K+, the Nernst potential is −71.5 mV (predicted shift with change from 3 to 9 mm extracellular K+ = 28.5 mV). The 5-HT-mediated hyperpolarization reversed at −62 ± 4 mV in 9 mm K+ (average shift = 24 ± 3 mV) (Fig. 3B). In pyramidal cells with 3 mm extracellular K+, the hyperpolarization induced by 5-HT reversed at −86 ± 2 mV (n = 8; data not shown). This reversal potential shifted to −65 ± 4 mV (n= 5) in 9 mm extracellular K+ (average shift = 21 ± 3 mV; data not shown). There was also no significant change in pyramidal cell input resistance with 5-HT (control: 224 ± 66 MΩ; 5-HT: 227 ± 67 MΩ; n = 4). These data suggest that the 5-HT1A-mediated hyperpolarization in both interneurons and pyramidal neurons involved a small increase in K+ conductance (as evidenced by the decrease in slope of the voltage response to current ramps) (Fig.3).
Fig. 3.

Ionic basis for 5-HT-induced hyperpolarizations.A, Response of a layer II nonpyramidal neuron to bath application of 30 μm 5-HT (3 mm extracellular K+). The recording was done in the presence (extracellular) of 1 μm TTX. The control (Ctl) and 5-HT traces intersect at approximately −91 mV (calculated Nernst potential for K+ = −100 mV). Calibration also applies to B. Current ramp used as stimulus shown above. Bottom, Box plot illustrating population data (n = 10 cells) for reversal potential in 3 mm extracellular K+. B, Same cell showing response to 5-HT in 9 mm extracellular K+. The reversal potential shifted positive (by 28 mV) to −63 mV (expected shift if Nernstian = 28.5 mV). Bottom, Box plot illustrating population data (n = 4 cells) for reversal potential in 9 mm extracellular K+. Current ramp used as stimulus shown above.
Depolarization
For the depolarization of nonpyramidal cells, there was also no significant change in average input resistance (control: 289 ± 40 MΩ; 5-HT: 339 ± 54 MΩ; n = 16;p = 0.06). We tested the reversal potential of the depolarization in six nonpyramidal cells, but no consistent pattern emerged. On average, the reversal potential was −73 ± 6 mV with 3 mm extracellular K+ (data not shown). In two cells the reversal potential approached EK(−95, −86 mV), but in the remaining four cells the reversal potential was more positive (−73, −65, −61, −58 mV). Similarly, in pyramidal neurons, the reversals were −95 and −65 mV (in 3 mm K+). The 5-HT2A-mediated depolarization in pyramidal neurons has been attributed to a decrease in K+ conductance (Davies et al., 1987;Araneda and Andrade, 1991). Depolarization in cat pyramidal neurons was found to be caused by an increasedIH (Spain, 1994). It has been suggested that the 5-HT2-mediated depolarization in layer V pyramidal cells from medial prefrontal cortex is caused by activation of a nonspecific cation current (Haj-Dahmane and Andrade, 1998). Our data do not differentiate between these possibilities. In pyramidal cells, there was no significant change in input resistance with hyperpolarization by 5-HT (control: 232 ± 27 MΩ; 5-HT: 216 ± 25 MΩ; n = 8).
Cell types
We next tested whether the variability in interneuronal responses to 5-HT was caused by cell type-specific effects.
Layer I
Our morphological findings for layer I neurons were similar to previous investigations (Hestrin and Armstrong, 1996; Zhou and Hablitz, 1996a). We identified four groups of cells in layer I (Fig.4): Cajal Retzius cells (CR), small cells at the pia surface, vertical axon cells, and horizontal axon cells. We identified seven vertical axon cells (Fig. 4A). These cells varied in soma size and sent an axonal projection to layers deeper than layer I. The horizontal extent of both the axonal and dendritic arbors was restricted (typically <200 μm) (Fig.4A). Our vertical axon cells may include both the “vertical cell” and the “cells sending axon collaterals to deeper layers” groups of Zhou and Hablitz (1996a).
Sixteen horizontal axon cells (Fig. 4B) were recorded from, two of which were clearly neurogliaform (cf. Hestrin and Armstrong, 1996). The axonal and dendritic arbors of the horizontal axon cells extended horizontally in layer I for up to 300–400 μm (Fig. 4B). These cells did not send axons deeper than the border between layers I–II. Zhou and Hablitz (1996a) found this cell type to be the most common in layer I. Layer I neurogliaform cells (Hestrin and Armstrong, 1996) had a very dense local axonal projection, and we considered them to be a subpopulation of horizontal axon cells.
We recorded from four CR cells in slices from animals aged P9–13 (Fig.4C). These cells were characterized by their large size, horizontal orientation, and thick dendrites (Fairen et al., 1984;Hestrin and Armstrong, 1996; Zhou and Hablitz, 1996a). The pia surface cells (n = 4) were small, with fine, sparse dendrites that remained local (data not shown). A subpial cell type was described by Zhou and Hablitz (1996a, their Fig. 14).
Interneurons could also be classified on the basis of their physiology (Kawaguchi and Kubota 1996, 1997, 1998; Cauli et al., 1997). FS interneurons have brief spikes followed by a large fast afterhyperpolarization (fAHP), show little spike frequency adaptation or sAHP, and exhibit irregular, interrupted firing in response to near threshold current injections. A variant of the FS pattern is exhibited by LS cells, which fire after a prolonged voltage ramp (Kawaguchi and Kubota, 1996, 1997, 1998). In contrast, RS interneurons exhibit broad spikes, repetitive firing with spike frequency adaptation, and sAHPs (Kawaguchi and Kubota 1996, 1997, 1998; Cauli et al., 1997). Zhou and Hablitz (1996b) reported that layer I neurons had FS physiology.Hestrin and Armstrong (1996) found layer I cells to be more heterogeneous. We found that the firing pattern of layer I interneurons (other than CR cells) varied from regular spiking (RS) to fast spiking. The distribution of AP half-widths formed a continuum in this layer (data not shown), and there was no clear relationship between firing patterns and cell morphology or response to 5-HT. Some (but not all) horizontal axon, vertical axon, and pia surface interneuron types had properties resembling those of FS cells (Hestrin and Armstrong, 1996;Zhou and Hablitz, 1996b), i.e., brief spikes, large fAHPs, and interrupted firing to near threshold stimuli.
Layer II
Seven of the layer II cells could be classified on the basis of axonal morphology as basket-like (extended plexus) cells (Feldman and Peters, 1978; White 1989; Somogyi et al., 1998) (Fig.5A). These cells had smooth dendrites with few or no spines. Pericellular varicosities were observed throughout the axonal arbor. In some cases, one or more collaterals descended to deeper layers (Fig. 5A). All of the basket-like cells exhibited physiological characteristics of LS or FS cells: brief action potentials, lack of an sAHP or spike frequency adaptation, and interruptions in the firing pattern in response to near rheobasic current injections (McCormick et al., 1985; Kawaguchi and Kubota, 1996, 1997, 1998). We also sampled cells that were bipolar and multipolar (data not shown), and we recorded from one chandelier cell with characteristic vertical axonal cassettes (Fig. 5C). This cell had FS physiology and did not respond to 5-HT. One cell was classified as neurogliaform (Fig. 5B). Two layer II interneurons (both bipolar) had RS physiology, and 21 had FS or LS physiology.
5-HT effects by cell type/layer
There was no clear relationship between the firing type or cell morphology of layer II interneurons and their response to 5-HT (Tables1, 3) (Para et al., 1998). A few cells were inadequately filled for classification by the axonal arbor, but were clearly not pyramidal (no prominent apical dendrite) and showed physiological characteristics of FS or LS cells. The basket-like and chandelier cells also had FS physiology, so we combined them with the morphologically unclassified cells as an FS/LS group. 5-HT depolarized 14 of these 21 cells (67%), hyperpolarized 3 (14%); 1 cell showed both responses (5%), and 4 cells did not respond to 5-HT (19%) (Fig. 1, Table 3).
In layer I, three of four CR cells were hyperpolarized, and one did not respond (Tables 1, 3). All four pia surface cells that we recorded from were depolarized by 5-HT (Table 1). In response to 5-HT, four or seven vertical axon cells (57%) were strongly depolarized (Table 3). One cell did not respond to 5-HT, and two of seven cells were hyperpolarized (29%; both 2 mV) (Tables 1, 3). Bath application of 5-HT led to depolarization in 4 of 16 (25%) of horizontal axon cells (including neurogliaform) (Table 3). Two cells (12.5%) were not affected by 5-HT, and 10 (62.5%) were hyperpolarized (Tables 1,3).
Statistics
We used Fisher's exact test to test the hypothesis that responses to 5-HT differed by layer and cell type. We found that the distribution of cells (Table 3) responding with depolarization versus hyperpolarization (vs no change) was statistically different (p = 0.03) for layer I versus II. When the amplitude and sign of 5-HT responses were compared, Student'st test also revealed a statistical difference between layers I and II (p < 0.001; two-tailed test). On average, layer I cell membrane potential depolarized by 0.5 ± 1 mV (n = 31) and layers II–III interneurons were depolarized by 4.3 ± 1 mV (n = 23). Because horizontal and vertical axon cells in layer I appeared to respond differently to 5-HT, we also tested whether these two populations of cells were different. The distributions (Table 3) were not statistically different (Fisher's exact test; p = 0.1), but amplitude of the response did differ between the two cell types (t test; p < 0.001). On average, vertical axon cells were depolarized (5 ± 2 mV; n= 7), and horizontal axon cells were hyperpolarized (−3 ± 1 mV;n = 16) by 5-HT.
The responses of layer I cells whose axons projected to layers II–III (vertical cell) and those of interneurons whose somas (and axons) were found in layers II–III were similar, suggesting that they form a functional group. We tested this hypothesis by comparing the combination of layer I vertical cells plus layers II–III interneurons with layer I horizontal cells (Table 3). CR cells were not included in this analysis because they may not be GABAergic. Pia surface cells were also not included because their axonal projections were not well filled. The distribution of responses was statistically different (p < 0.002; Fisher's exact test), and response amplitude also differed significantly (t test). Cells whose projection remained in layer I were on average hyperpolarized by 3 ± 1 mV (n = 16), and cells projecting to layers II–II were depolarized (5 ± 1 mV; n = 30). Responses of layer I projecting (horizontal axon cells) versus layers II–III projecting cells (layer I vertical axon cells plus layers II–III interneurons) are summarized graphically in Fig.6. These data suggest that interneuronal responses to 5-HT can be functionally differentiated by axonal targets. That is, axonal projection pattern is a better predictor of response to 5-HT than soma location or physiological type.
Fig. 6.

Box plot (Tukey, 1977) illustrating the distribution of membrane potential responses of interneurons whose axonal projection was restricted to layer I (layer I horizontal axon cells) to the responses of interneurons with axonal projections within layers II–III (layer I vertical axon cells plus layers II–III interneurons). The horizontal dashed line indicates no response to 5-HT. Hyperpolarizing responses fall below theline; depolarizations are above theline.
5-HT effects on sIPSCs
We have described spontaneous synaptic currents in whole-cell recordings of pyramidal cells (van Brederode et al., 2001). Most of these currents are GABAA-mediated IPSCs (van Brederode et al., 2001).
To test whether 5-HT2-mediated depolarization of supragranular interneurons results in altered tonic inhibition of pyramidal cells, we examined the effects of the selective 5-HT2 agonist DOI (10 μm, bath applied) on sIPSCs in pyramidal cells from layers II and III (Fig.7). We used whole-cell recordings with a CsCH3SO4-based internal solution (to block effects of postsynaptic K+ conductances) and applied CNQX (10 μm) and AP5 (50 μm) to block AMPA receptor- and NMDA receptor-mediated responses, respectively. This selects for GABAA receptor-mediated events (van Brederode et al., 2001).
Fig. 7.

5-HT2A receptor activation increased the frequency of spontaneous IPSCs recorded in layer II pyramidal neurons. A, Histogram of the inter-IPSC interval for a single pyramidal cell as a function of time before, during, and after bath application of 30 μm of the 5-HT2agonist DOI (means ± SEM). DOI was present for the time indicated by the black bar. The interevent interval reversibly decreased in DOI (increased frequency of IPSCs). B, Representative traces showing IPSCs from the initial control period, in the presence of DOI, and control wash for the same cell as in A. C, Cumulative frequency plots for IPSC amplitude (left) or interval (right) showing that in DOI the intervals decrease (frequency increase) with no change in IPSC amplitude.
We tested 14 cells with DOI (Fig. 7) at a holding potential of −60 mV. Mean sIPSC amplitude did not change in DOI (Fig. 7C). In 9 of 14 cells, we found an increase in mean frequency of at least 5% (range, 6–40%); there was a decrease in 1 cell, and no change in 4 cells. The initial control sIPSC frequency was 36 ± 4 Hz (17–73 Hz). In DOI, the mean frequency was significantly increased to 39 ± 4 Hz (19–73 Hz; p < 0.05; paired ttest). After a 10 min wash in control media, the mean frequency was 35 ± 11 Hz (13–75 Hz). We also calculated the percentage change in frequency for each cell. For all 14 cells combined, the average response was an increase in frequency of 11 ± 3%. In the nine cells showing an increase, the average increase was 16 ± 4%. Thus 5-HT2 receptor activation of interneurons results in an increased frequency of tonic inhibition in pyramidal neurons [see also Zhou and Hablitz (1999b)].
DISCUSSION
We determined the dominant effect of 5-HT on membrane potential in different types of neocortical neurons. Previous work on cellular effects of 5-HT concentrated on layer V pyramidal cells, which were depolarized, hyperpolarized, or showed mixed responses to bath-applied 5-HT (Davies et al., 1987; Araneda and Andrade, 1991; Spain, 1994;Marek and Aghajanian, 1998). Responses of supragranular pyramidal cells to 5-HT had not been studied previously. We found that approximately half of the superficial layers II–III pyramidal cells were depolarized and approximately half were hyperpolarized by bath-applied 5-HT. Mixed responses were not observed. In deep layers II–III, nearly all pyramidal cells (90%) were depolarized. Additionally, the sAHP was reduced and firing behavior was altered. Overall, these data indicate that 5-HT variably affects pyramidal cell membrane potential and facilitates responses to large suprathreshold depolarizations.
To obtain a broader perspective of the action of 5-HT on cortical circuitry, we applied 5-HT to nonpyramidal cells. As with pyramidal neurons, interneuronal responses to 5-HT were variable. Our main finding was that the direction and amplitude of interneuron responses were related to the laminar pattern of their axonal projections.
Hyperpolarizations
Our data suggest that interneuronal hyperpolarizations were caused by activation of 5-HT1A receptors, on the basis of mimicry of the hyperpolarization by 8-OHDPAT (specific 5-HT1A and 5-HT7 agonist) (Middlemiss et al., 1986). Hyperpolarizations were not prevented by 5-HT2/5-HT7 antagonists (Hoyer et al., 1994). This is consistent with findings in other cell types (Andrade and Nicoll, 1987; Collino and Halliwell, 1987; Araneda and Andrade, 1991; Schmitz et al., 1998). Immunocytochemistry andin situ hybridization indicate that 5-HT1A receptors are expressed by interneurons and pyramidal cells in supragranular layers of sensorimotor cortex (Pazos and Palacios, 1985; Morilak et al., 1993; Wright et al., 1995).
Reversal potentials and ion substitution experiments clearly implicate K+ channels in the 5-HT-induced hyperpolarizations of both interneurons and pyramidal neurons, consistent with findings on layer V pyramidal neurons (Spain, 1994) and other cell types (Collino and Halliwell, 1987). The lack of measurable change in input conductance suggests that conductance changes are small, obscured by other effects, or occur in a location remote from the soma. In CA1 pyramidal neurons, 5-HT1Areceptors activate G-proteins (Gi/Go subclass), which interact with inwardly rectifying K+channels (Andrade and Nicoll, 1987).
Depolarizations
With bath-applied 5-HT, depolarizations are at least partly 5-HT2 receptor mediated. This conclusion is based on mimicry of the depolarization by a specific 5-HT2 agonist (DOI) and block of 5-HT-induced depolarizations by specific 5-HT2 antagonists (ketanserin, ritanserin). The agents that we used do not allow us to distinguish between 5-HT2A and 5-HT2C receptors. Both receptor types are present in cortex (Pompeiano et al., 1994; Wright et al., 1995). In prefrontal cortex, layer III was intensely stained for 5-HT2A receptors (Jakab and Goldman-Rakic, 1998). 5-HT2C receptors are sparsely distributed in cortex (Molineaux et al., 1989). A role for other receptors (cf. Gustafson et al., 1996; Beique and Andrade, 2001) requires further study.
In primate (Jakab and Goldman-Rakic, 1998) and rat (Willins et al., 1997; Hamada et al., 1998) prefrontal cortex, 5-HT2A receptors are found on proximal apical dendrites of pyramidal neurons. This region is hypothesized to be a “hot spot” for 5-HT2A-mediated physiological actions relevant to normal and “psychotic” states (Jakab and Goldman-Rakic, 1998). 5-HT2A receptors are also present on terminals and in a subset of interneurons (Jakab and Goldman-Rakic, 1998; Vysokanov et al., 1998). In piriform cortex, 5-HT2A receptors are preferentially located on nonpyramidal interneurons, which are depolarized by 5-HT2 receptor activation (Sheldon and Aghajanian, 1991; Gellman and Aghajanian, 1994).
In hippocampal interneurons, fast depolarizations were induced by activation of 5-HT3 receptors (Kawa, 1994;McMahon and Kauer, 1997). In neocortex, 5-HT3receptors are reportedly primarily expressed on a subset of nonpyramidal cells [but see Edwards et al. (1990) and Morales and Bloom (1997)]. 5-HT3 induced fast inward currents in a small percentage of layer I interneurons (Zhou and Hablitz, 1999b) and cultured neocortical neurons (Matsuoka et al., 1997). 5-HT3 receptor activation may depolarize a subset of interneurons in layer V (Williams et al., 1999; Xiang et al., 1999). We observed one positive response to a limited number of direct (n = 1 of 13) or bath applications (n = 3) of a 5-HT3 agonist. Because we did not systematically test direct applications of higher doses of 5-HT3 agonists, and bath application is likely to cause significant desensitization of 5-HT3receptors [Jackson and Yakel (1995); but see Roerig et al. (1997)], we do not consider the role of 5-HT3 receptors to be adequately tested.
Laminar and cell type comparisons
In layer I, 5-HT depolarized or hyperpolarized cells with similar frequency. An interesting pattern emerged from the most commonly recorded neuron types: horizontal axon cells and vertical axon cells.
The principal effect of 5-HT on horizontal axon cells was hyperpolarization. This could reduce tonic inhibition and selectively disinhibit responses of pyramidal cells to distal inputs. The horizontal extent of the axonal arbors suggests that this effect would influence a large area (e.g., multiple columns). Such disinhibition could also reduce the independence of Ca2+dynamics in distal dendrites (e.g., apical tuft) of layer V pyramidal cells versus the soma/proximal dendrite (Yuste et al., 1994; Callaway and Ross, 1995; Schiller et al., 1997). Alternatively, the distal dendritic responses to GABAA activation may be depolarizing (Owens et al., 1996; Cerne and Spain, 1997; Mienville, 1998). Serotonergic reduction in such inputs would then decrease pyramidal cell excitability.
Most layer I vertical axon cells were depolarized by 5-HT. Axonal anatomy suggests that these cells could inhibit pyramidal cell dendrites proximally, within a more local area than horizontal axon cells (e.g., within column), suggesting possible differences between serotonergic effects on inter-column and intra-column inhibition. 5-HT would thus increase tonic inhibition of nearby pyramidal cells.
Most layers II–III nonpyramidal cells, including RS and FS cells, are depolarized by 5-HT. Our major finding is that interneurons whose axons terminate in layers II–III (layers II–III interneurons, layer I vertical axon) form a functional group. Depolarization of these cells would increase tonic inhibition of pyramidal cells, thereby reducing repetitive firing (Kim et al., 1995; van Brederode and Spain, 1995), responses to excitatory inputs, and backpropagation of APs (Tsubokawa and Ross, 1996). This could bias responsiveness of pyramidal neurons to strong versus weak stimuli. Somatic and proximal dendritic inputs have also been proposed to control rhythmicity of local circuitry in cortex (Somogyi et al., 1998; Whittington et al., 2000). Serotonergic depolarization of interneurons providing proximal inputs to pyramidal cells (e.g., basket and chandelier cells) (Somogyi et al., 1998) may therefore play a more complicated role than direct control of membrane excitability.
Synaptic transmission
DOI application increases GABA release in cortex, suggesting excitation of GABAergic interneurons via 5-HT2receptors (Abi-Saab et al., 1999). We confirmed that the frequency of sIPSCs on pyramidal neurons was increased by 5HT2 activation (Zhou and Hablitz, 1999b). Zhou and Hablitz (1999b) further suggest that the effects of 5-HT are dependent on AP-induced GABA release, because miniature IPSCs (in the presence of TTX) were not modulated by 5-HT. The 5-HT2-induced increase in spontaneous IPSCs on pyramidal neurons should result in increased tonic inhibition, reducing overall network excitability. Williams et al. (2002) suggested that increased feedforward inhibition in response to activation of 5HT2 receptors on FS interneurons might enhance spatial tuning of working memory in primate prefrontal cortex.
5-HT increased the frequency of sEPSCs recorded in neocortical pyramidal cells (layer V >> II/) (Zhou and Hablitz, 1999b;Aghajanian and Marek, 2000; Lambe et al., 2000; Beique and Andrade, 2001) [but see Newberry et al. (1999)] and hippocampus (Ropert and Guy, 1991; Shen and Andrade, 1998). Zhou and Hablitz (1999b)also suggest a temporal variable to 5-HT modulation of cortical circuitry: 5-HT2A activation initially causes increase in the frequency of sIPSCs, which then desensitizes. The enhancement of sEPSCs is weaker but longer lasting. Thus, short epochs of 5-HT release would lead to increased inhibition of cortical circuitry, but prolonged 5-HT presence would facilitate this circuitry. We did not observe biphasic responses, but we did not systematically examine responses to long applications of DOI.
Function
5-HT2A receptors on apical dendrites of pyramidal neurons may act to gate incoming signals (Jakab and Goldman-Rakic, 1998). A proper balance of 5-HT activation would be required in prefrontal cortex to facilitate working memory and latent inhibition. Too much activation of these receptors would prevent pyramidal cells from shutting down, possibly leading to the schizophrenic state (Abi-Dargham et al., 1997). The present results suggest that this balance must also include the differential laminar modulation of GABAergic inhibition by 5-HT. Interestingly, increased levels of GABAA receptors are reported in superficial layers of cingulate cortex in schizophrenic patients (Benes et al., 1992).
5-HT2A receptors may be the site of action of hallucinogens and antipsychotic drugs (Jakab and Goldman-Rakic, 1998;Aghajanian and Marek, 1999). The preponderance of serotonergic excitation of interneurons and consequent tonic inhibition of pyramidal cells would also suppress seizure generation [genetically epilepsy-prone rats have a low density of serotonergic projections to forebrain (Jobe, 1973; Statnick et al., 1996)].
Footnotes
This work was supported by a Veterans Affairs Merit Award (W.J.S), National Institutes of Health Grant NS34769 (J.F.M.v.B.), and National Institute of Neurological Disorders and Stroke Grant NS 33579 (R.C.F.). We thank Richard Lee for excellent technical assistance and Dr. W. Armstrong for reading an earlier version of this manuscript.
Correspondence should be addressed to Dr. Robert C. Foehring, Department of Anatomy and Neurobiology, University of Tennessee, Memphis, 855 Monroe Avenue, Memphis, TN 38163. E-mail:foehring@nb.utmem.edu.
REFERENCES
- 1.Abi-Dargham A, Laruelle M, Aghajanian GK, Charney D, Krystal J. The role of serotonin in the pathophysiology and treatment of schizophrenia. J Neuropsychiatry Clin Neurosci. 1997;9:1–17. doi: 10.1176/jnp.9.1.1. [DOI] [PubMed] [Google Scholar]
- 2.Abi-Saab, Bubser M, Roth RH, Deutch AY. 5-HT2 receptor regulation of extracellular GABA levels in the prefrontal cortex. Neuropsychopharmacology. 1999;20:92–96. doi: 10.1016/S0893-133X(98)00046-3. [DOI] [PubMed] [Google Scholar]
- 3.Aghajanian GK, Marek GJ. Serotonin and hallucinogens. Neuropsychopharmacology. 1999;21:16S–23S. doi: 10.1016/S0893-133X(98)00135-3. [DOI] [PubMed] [Google Scholar]
- 4.Aghajanian GK, Marek GJ. Serotonin model of schizophrenia: emerging role of glutamate mechanisms. Brain Res Rev. 2000;31:302–312. doi: 10.1016/s0165-0173(99)00046-6. [DOI] [PubMed] [Google Scholar]
- 5.Andrade R, Nicoll RA. Pharmacologically distinct actions of serotonin on single pyramidal neurons of the rat hippocampus recorded in vitro. J Physiol (Lond) 1987;394:99–124. doi: 10.1113/jphysiol.1987.sp016862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Araneda R, Andrade R. 5-Hydroxytryptamine2 and 5-hydroxytryptamine1A receptors mediate opposing responses on membrane excitability in rat association cortex. Neuroscience. 1991;40:399–412. doi: 10.1016/0306-4522(91)90128-b. [DOI] [PubMed] [Google Scholar]
- 7.Armstrong-James MA, Johnson FR. Quantitative studies of post-natal changes in synapses in rat superficial motor cerebral cortex. An electron microscopical study. Z Zellforsch Mikrosk Anat. 1970;110:559–568. doi: 10.1007/BF00330104. [DOI] [PubMed] [Google Scholar]
- 8.Beique JC, Andrade R. Multiple serotonin receptor subtypes increase glutamatergic synaptic activity in rat prefrontal cortex. Soc Neurosci Abstr. 2001;27:380.3. [Google Scholar]
- 9.Benes FM, Vincent SL, Alsterberg G, Bird ED, SanGiovani JP. Increased GABAA receptor binding in superficial layers of cingulate cortex in schizophrenics. J Neurosci. 1992;12:924–929. doi: 10.1523/JNEUROSCI.12-03-00924.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bobker DH. A slow excitatory potential mediated by 5-HT2 receptors in nucleus prepositus hypoglossi. J Neurosci. 1994;14:2428–2434. doi: 10.1523/JNEUROSCI.14-04-02428.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bruinvels AT, Landwehrmeyer B, Gustafson EL, Durkin MM, Mengod G, Branchek TA, Hoyer D, Palacios JM. Localization of the 5-HT1B, 5-HT1Da, 5-HT1E and 5-HT1F receptor messenger RNA in rodent and primate brain. Neuropharmacology. 1994;33:367–386. doi: 10.1016/0028-3908(94)90067-1. [DOI] [PubMed] [Google Scholar]
- 12.Burnet PWJ, Eastwod SL, Lacey K, Harrison PJ. The distribution of 5-HT1A and 5-HT2 receptor mRNA in human brain. Brain Res. 1995;676:157–168. doi: 10.1016/0006-8993(95)00104-x. [DOI] [PubMed] [Google Scholar]
- 13.Callaway JC, Ross WN. Frequency-dependent propagation of sodium action potentials in dendrites of hippocampal CA1 pyramidal neurons. J Neurophysiol. 1995;74:1395–1403. doi: 10.1152/jn.1995.74.4.1395. [DOI] [PubMed] [Google Scholar]
- 14.Cauli B, Lambolez B, Angulo MC, Ropert N, Tsuzuki K, Hestrin S, Rossier J. Molecular and physiological diversity of cortical nonpyramidal cells. J Neurosci. 1997;17:3894–3906. doi: 10.1523/JNEUROSCI.17-10-03894.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cerne R, Spain WJ. GABAA mediated afterdepolarization in pyramidal neurons from rat neocortex. J Neurophysiol. 1997;77:1039–1045. doi: 10.1152/jn.1997.77.2.1039. [DOI] [PubMed] [Google Scholar]
- 16.Collino A, Halliwell JV. Differential modulation of three separate K-conductances in hippocampal CA1 neurons by serotonin. Nature. 1987;328:73–77. doi: 10.1038/328073a0. [DOI] [PubMed] [Google Scholar]
- 17.Davies FM, Diesz RA, Prince DA, Peroutka SJ. Two distinct effects of 5-hydroxytryptamine on single cortical neurons. Brain Res. 1987;423:347–352. doi: 10.1016/0006-8993(87)90861-4. [DOI] [PubMed] [Google Scholar]
- 18.DeFelipe J, Hendry SHC, Hashikawa T, Jones EG. Synaptic relationships of serotonin-immunoreactive terminal baskets on GABA neurons in the cat auditory cortex. Cereb Cortex. 1991;1:177–183. doi: 10.1093/cercor/1.2.117. [DOI] [PubMed] [Google Scholar]
- 19.Dodt H-U, Zieglgansberger W. Infrared videomicroscopy: a new look at neuronal structure and function. Trends Neurosci. 1994;17:453–458. doi: 10.1016/0166-2236(94)90130-9. [DOI] [PubMed] [Google Scholar]
- 20.Edagawa Y, Saito H, Abe K. Endogenous serotonin contributes to a developmental decrease in long-term potentiation in the rat visual cortex. J Neurosci. 2001;21:1532–1537. doi: 10.1523/JNEUROSCI.21-05-01532.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Edwards E, Hampton E, Ashby CR, Zhang J, Wang RY. 5-HT3-like receptors in the rat medial prefrontal cortex: further pharmacological characterization. Brain Res. 1990;733:21–30. doi: 10.1016/0006-8993(96)00529-x. [DOI] [PubMed] [Google Scholar]
- 22.Fairen A, DeFelipe J, Martinez-Ruiz R. Nonpyramidal neurons: general account. In: Peters A, Jones E, editors. The cerebral cortex, V1. Plenum; New York: 1984. pp. 201–253. [Google Scholar]
- 23.Feldman ML, Peters A. The forms of non-pyramidal neurons in the visual cortex of the rat. J Comp Neurol. 1978;179:761–794. doi: 10.1002/cne.901790406. [DOI] [PubMed] [Google Scholar]
- 24.Foehring RC, van Brederode JFM, Spain WJ. Serotonergic and cholinergic modulation of visually identified interneurons in slices of immature rat sensorimotor cortex. Soc Neurosci Abstr. 1996;22:606. [Google Scholar]
- 25.Foote SL, Morrison JH. Postnatal development of laminar innervation patterns by monoaminergic fibers in monkey (Macaca fascicularis) primary visual cortex. J Neurosci. 1984;11:2667–2680. doi: 10.1523/JNEUROSCI.04-11-02667.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gellman RL, Aghajanian GK. Serotonin2 receptor-mediated excitation of interneurons in piriform cortex: antagonism by atypical antipsychotic drugs. Neuroscience. 1994;58:515–525. doi: 10.1016/0306-4522(94)90077-9. [DOI] [PubMed] [Google Scholar]
- 27.Gustafson EL, Durkin MM, Bard JA, Zgombick J, Branchek TA. A receptor autoradiographic and in situ hybridization analysis of the distribution of the 5-HT7 receptor in rat brain. Br J Pharmacol. 1996;117:657–666. doi: 10.1111/j.1476-5381.1996.tb15241.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Haj-Dahmane S, Andrade R. Ionic mechanism of the slow afterdepolarization induced by muscarinic receptor activation in rat prefrontal cortex. J Neurophysiol. 1998;80:1197–1210. doi: 10.1152/jn.1998.80.3.1197. [DOI] [PubMed] [Google Scholar]
- 29.Hamada S, Senzaki K, Hamaguchi-Hamada K, Tabuchi K, Yamamoto H, Yamamoto T, Okano H, Okado N. Localization of 5-HT2A receptor in rat cerebral cortex and olfactory system revealed by immunocytochemistry using two antibodies raised in rabbit and chicken. Mol Brain Res. 1998;54:199–211. doi: 10.1016/s0169-328x(97)00322-7. [DOI] [PubMed] [Google Scholar]
- 30.Hestrin S, Armstrong WE. Morphology and physiology of cortical neurons in layer I. J Neurosci. 1996;16:5290–5300. doi: 10.1523/JNEUROSCI.16-17-05290.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Horikawa K, Armstrong WE. A versatile means of intracellular labeling: injection of biocytin and its detection with avidin conjugates. J Neurosci Methods. 1988;25:1–12. doi: 10.1016/0165-0270(88)90114-8. [DOI] [PubMed] [Google Scholar]
- 32.Hornung J, Celio MR. The selective innervation by serotonergic axons of calbindin-containing interneurons in the neocortex and hippocampus of the marmoset. J Comp Neurol. 1992;297:165–181. doi: 10.1002/cne.903200404. [DOI] [PubMed] [Google Scholar]
- 33.Hoyer D, Clarke DE, Fozard JR, Hartig PR, Martin GR, Mylecharane EJ, Saxena PR, Humphrey PA. VII. International union of pharmacology classification of receptors for 5-hydroxytryptamine (serotonin). Pharmacol Rev. 1994;46:157–203. [PubMed] [Google Scholar]
- 34.Jackson DA, White SR. Receptor subtypes mediating facilitation by serotonin of excitability of spinal motoneurons. Neuropharmacology. 1990;29:787–797. doi: 10.1016/0028-3908(90)90151-g. [DOI] [PubMed] [Google Scholar]
- 35.Jackson MB, Yakel JL. The 5-HT3 receptor channel. Annu Rev Physiol. 1995;57:447–468. doi: 10.1146/annurev.ph.57.030195.002311. [DOI] [PubMed] [Google Scholar]
- 36.Jacobs BL, Wilkinson LO, Fornal CA. The role of brain serotonin: a neurophysiologic perspective. Neuropsychopharmacology. 1990;3:473–479. [PubMed] [Google Scholar]
- 37.Jakab RL, Goldman-Rakic PS. 5-Hydroxytryptamine2A serotonin receptors in the primate cerebral cortex: possible site of action of hallucinogenic and antipsychotic drugs in pyramidal cell apical dendrites. Proc Natl Acad Sci USA. 1998;95:735–740. doi: 10.1073/pnas.95.2.735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jobe PC. Role of brain 5-hydroxytryptamine in audiogenic seizures in the rat. Life Sci. 1973;13:1–13. [PubMed] [Google Scholar]
- 39.Jordan LM, Fredrickson RCA, Phillis JM, Lake N. Microelectrophoresis of 5-hydroxytryptamine: a clarification of its action on cerebral cortical neurones. Brain Res. 1972;40:552–558. doi: 10.1016/0006-8993(72)90161-8. [DOI] [PubMed] [Google Scholar]
- 40.Kawa K. Distribution and functional properties of 5-HT3 receptors in the rat hippocampal dentate gyrus: a patch-clamp study. J Neurophysiol. 1994;71:1935–1947. doi: 10.1152/jn.1994.71.5.1935. [DOI] [PubMed] [Google Scholar]
- 41.Kawaguchi Y. Groupings of nonpyramidal and pyramidal cells with specific physiological and morphological characteristics in rat frontal cortex. J Neurophysiol. 1993;69:416–431. doi: 10.1152/jn.1993.69.2.416. [DOI] [PubMed] [Google Scholar]
- 42.Kawaguchi Y. Physiological subgroups of nonpyramidal cells with specific morphological characteristics in layer II/III of rat frontal cortex. J Neurosci. 1995;15:2638–2655. doi: 10.1523/JNEUROSCI.15-04-02638.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kawaguchi Y, Kubota Y. Physiological and morphological identification of somatostatin- or vasoactive intestinal polypeptide-containing cells among GABAergic cell subtypes in rat frontal cortex. J Neurosci. 1996;16:2701–2715. doi: 10.1523/JNEUROSCI.16-08-02701.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kawaguchi Y, Kubota Y. GABAergic cell subtypes and their synaptic connections in rat frontal cortex. Cereb Cortex. 1997;7:476–486. doi: 10.1093/cercor/7.6.476. [DOI] [PubMed] [Google Scholar]
- 45.Kawaguchi Y, Kubota Y. Neurochemical features and synaptic connections of large physiologically-identified GABAergic cells in the rat frontal cortex. Neuroscience. 1998;85:677–701. doi: 10.1016/s0306-4522(97)00685-4. [DOI] [PubMed] [Google Scholar]
- 46.Kawaguchi Y, Shindou T. Noradrenergic excitation and inhibition of GABAergic cell types in rat frontal cortex. J Neurosci. 1998;18:6963–6976. doi: 10.1523/JNEUROSCI.18-17-06963.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kim HG, Beierlein M, Connors BW. Inhibitory control of excitable dendrites in neocortex. J Neurophysiol. 1995;74:1810–1814. doi: 10.1152/jn.1995.74.4.1810. [DOI] [PubMed] [Google Scholar]
- 48.Krnjevic K, Phillis JW. Ionophoretic studies of neurons in the mammalian cerebral cortex. J Physiol (Lond) 1963;165:274–304. doi: 10.1113/jphysiol.1963.sp007057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lakoski JM, Aghajanian GK. Effects of ketanserin on neuronal responses to serotonin in the prefrontal cortex, lateral geniculate, and dorsal raphe nucleus. Neuropharmacology. 1985;24:265–273. doi: 10.1016/0028-3908(85)90130-3. [DOI] [PubMed] [Google Scholar]
- 50.Lambe EK, Goldman-Rakic PS, Aghajanian GK. Serotonin induces EPSCs preferentially in layer V pyramidal neurons of the frontal cortex in the rat. Cereb Cortex. 2000;10:974–980. doi: 10.1093/cercor/10.10.974. [DOI] [PubMed] [Google Scholar]
- 51.Lorenzon NM, Foehring RC. The ontogeny of repetitive firing and its modulation by norepinephrine in rat neocortical neurons. Dev Brain Res. 1993;73:213–223. doi: 10.1016/0165-3806(93)90141-v. [DOI] [PubMed] [Google Scholar]
- 52.Marek GJ, Aghajanian GK. 5-hydroxytryptamine-induced excitatory postsynaptic currents in neocortical layer V pyramidal cells: suppression by μ-opiate receptor activation. Neuroscience. 1998;86:485–497. doi: 10.1016/s0306-4522(98)00043-8. [DOI] [PubMed] [Google Scholar]
- 53.Martin-Ruiz R, Puig MV, Celada P, Shapiro DA, Roth BL, Mengod G, Artigas F. Control of serotonergic function in medial prefrontal cortex by serotonin-2A receptors through a glutamate-dependent mechanism. J Neurosci. 2001;21:9856–9866. doi: 10.1523/JNEUROSCI.21-24-09856.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Matsuoka T, Nishizaka T, Ikeuchi Y, Okada Y, Sumino K. Differential effects of the serotonin receptors on cultured rat cerebral cortical neurons. Cell Mol Life Sci. 1997;53:233–236. doi: 10.1007/PL00000596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.McCormick DA, Connors BW, Lighthall JW, Prince DA. Comparative electrophysiology of pyramidal and sparsely spiny stellate neurons of the neocortex. J Neurophysiol. 1985;54:782–806. doi: 10.1152/jn.1985.54.4.782. [DOI] [PubMed] [Google Scholar]
- 56.McMahon LL, Kauer JA. Hippocampal interneurons are excited via serotonin-gated ion channels. J Neurophysiol. 1997;78:2493–2502. doi: 10.1152/jn.1997.78.5.2493. [DOI] [PubMed] [Google Scholar]
- 57.Middlemiss DN, Palmer AM, Edel N, Bowen DM. Binding of the novel serotonin agonist 8-hydroxy-2-(Di-n-propylamino) tertalin in normal and alzheimer brain. J Neurochem. 1986;46:993–996. doi: 10.1111/j.1471-4159.1986.tb13069.x. [DOI] [PubMed] [Google Scholar]
- 58.Mienville J-M. Persistent depolarizing action of GABA in rat Cajal-Retzius cells. J Physiol (Lond) 1998;512:809–817. doi: 10.1111/j.1469-7793.1998.809bd.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Molineaux SM, Jessel TM, Axel R, Julius D. 5-HT1c is a prominent serotonin receptor subtype in the central nervous system. Proc Natl Acad Sci USA. 1989;86:6793–6897. doi: 10.1073/pnas.86.17.6793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Morales M, Bloom FE. The 5-HT3 receptor is present in different subpopulations of GABAergic neurons in the rat telencephalon. J Neurosci. 1997;17:3157–3167. doi: 10.1523/JNEUROSCI.17-09-03157.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Morales M, Battenberg E, de Lecea L, Bloom FE. The type 3 serotonin receptor is expressed in a subpopulation of GABAergic neurons in the rat neocortex and hippocampus. Brain Res. 1996;731:199–202. doi: 10.1016/0006-8993(96)00557-4. [DOI] [PubMed] [Google Scholar]
- 62.Morilak DA, Garlow SJ, Ciaranello RD. Immunocytochemical localization and description of neurons expressing serotonin receptors in the rat brain. Neuroscience. 1993;54:701–717. doi: 10.1016/0306-4522(93)90241-7. [DOI] [PubMed] [Google Scholar]
- 63.Mulligan KA, Tork I. Serotoninergic innervation of the cat cerebral cortex. J Comp Neurol. 1988;270:86–110. doi: 10.1002/cne.902700108. [DOI] [PubMed] [Google Scholar]
- 64.Nakazawa M, Koh T, Kani K, Maeda T. Transient patterns of serotonergic innervation in the rat visual cortex: normal development and effect of neonatal enucleation. Dev Brain Res. 1992;66:77–90. doi: 10.1016/0165-3806(92)90143-k. [DOI] [PubMed] [Google Scholar]
- 65.Neher E. Correction for liquid junction potentials in patch clamp experiments. Methods Enzymol. 1992;207:123–131. doi: 10.1016/0076-6879(92)07008-c. [DOI] [PubMed] [Google Scholar]
- 66.Newberry NR, Footitt DR, Papanastassiou V, Reynolds DJ. Actions of 5-HT on human neocortical neurones in vitro. Brain Res. 1999;833:93–100. doi: 10.1016/s0006-8993(99)01540-1. [DOI] [PubMed] [Google Scholar]
- 67.Owens DF, Boyce LH, Davis MBE, Kriegstein AR. Excitatory GABA responses in embryonic and neonatal cortical slices demonstrated by gramicidin perforated patch recordings and calcium imaging. J Neurosci. 1996;16:6414–6423. doi: 10.1523/JNEUROSCI.16-20-06414.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Para P, Gulyas AI, Miles R. How many subtypes of inhibitory cells in the hippocampus? Neuron. 1998;20:983–993. doi: 10.1016/s0896-6273(00)80479-1. [DOI] [PubMed] [Google Scholar]
- 69.Pazos A, Palacios JM. Quantitative autoradiographic mapping of serotonin receptors in the rat brain. I. Serotonin-1 receptors. Brain Res. 1985;346:205–230. doi: 10.1016/0006-8993(85)90856-x. [DOI] [PubMed] [Google Scholar]
- 70.Pompeiano M, Palacios JM, Mengod G. Distribution of the serotonin 5-HT2 receptor family mRNAs: comparison between 5-HT2A and 5-HT2C receptors. Mol Brain Res. 1994;23:163–178. doi: 10.1016/0169-328x(94)90223-2. [DOI] [PubMed] [Google Scholar]
- 71.Reader TA, Ferron A, Descarries L, Jasper HH. Modulatory role for biogenic amines in the cerebral cortex. Microiontophoretic studies. Brain Res. 1979;160:217–229. doi: 10.1016/0006-8993(79)90420-7. [DOI] [PubMed] [Google Scholar]
- 72.Reuter LE, Jacobs BL. A microdialysis examination of serotonin release in the rat forebrain induced by behavioral/environmental manipulations. Brain Res. 1996;739:57–69. doi: 10.1016/s0006-8993(96)00809-8. [DOI] [PubMed] [Google Scholar]
- 73.Roberts MHT, Straughan DW. Excitation and depression of cortical neurones by 5-hydroxytryptamine. J Physiol (Lond) 1967;193:269–294. doi: 10.1113/jphysiol.1967.sp008357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Roerig B, Nelson DA, Katz LC. Fast synaptic signaling by nicotinic acetylcholine and serotonin 5-HT3 receptors in developing visual cortex. J Neurosci. 1997;17:8353–8362. doi: 10.1523/JNEUROSCI.17-21-08353.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Ropert N, Guy N. Serotonin facilitates GABAergic transmission in the CA1 region of rat hippocampus in vitro. J Physiol (Lond) 1991;441:121–136. doi: 10.1113/jphysiol.1991.sp018742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Roth BL, Hamblin MW, Ciaranello RD. Developmental regulation of 5-HT2 and 5-HT1C mRNA and receptor levels. Dev Brain Res. 1991;58:51–58. doi: 10.1016/0165-3806(91)90236-c. [DOI] [PubMed] [Google Scholar]
- 77.Schiller J, Schiller Y, Stuart G, Sakmann B. Calcium action potentials restricted to distal apical dendrites of rat neocortical pyramidal neurons. J Physiol (Lond) 1997;505:605–616. doi: 10.1111/j.1469-7793.1997.605ba.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Schmitz D, Gloveli T, Empson RM, Heineman U. Serotonin reduces polysynaptic inhibition via 5-HT1A receptors in the superficial entorhinal cortex. J Neurophysiol. 1998;80:1116–1121. doi: 10.1152/jn.1998.80.3.1116. [DOI] [PubMed] [Google Scholar]
- 79.Sheldon PW, Aghajanian GK. Excitatory responses to serotonin (5-HT) in neurons in rat pyriform cortex: evidence for mediation by 5-HT1c in pyramidal cells and 5-HT2 receptors in interneurons. Synapse. 1991;9:208–218. doi: 10.1002/syn.890090307. [DOI] [PubMed] [Google Scholar]
- 80.Shen RY, Andrade R. 5-Hydroxytryptamine2 receptor facilitates GABAergic neurotransmission in rat hippocampus. J Pharmacol Exp Ther. 1998;285:805–812. [PubMed] [Google Scholar]
- 81.Smiley JF, Goldman-Rakic PS. Serotonergic axons in monkey prefrontal cortex synapse preferentially on interneurons as demonstrated by serial section electron microscopy. J Comp Neurol. 1996;367:431–443. doi: 10.1002/(SICI)1096-9861(19960408)367:3<431::AID-CNE8>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 82.Somogyi P, Tamas G, Lujan R, Buhl EH. Salient features of synaptic organisation in the cerebral cortex. Brain Res Rev. 1998;26:113–135. doi: 10.1016/s0165-0173(97)00061-1. [DOI] [PubMed] [Google Scholar]
- 83.Spain WJ. Serotonin has different effects on two classes of Betz cells from the cat. J Neurophysiol. 1994;72:1925–1937. doi: 10.1152/jn.1994.72.4.1925. [DOI] [PubMed] [Google Scholar]
- 84.Statnick MA, Dailey JW, Jobe PC, Browning RA. Abnormalities in 5-HT1A and 5-HT1B receptor binding in severe-seizure genetically epilepsy-prone rats. Neuropharmacology. 1996;35:111–118. doi: 10.1016/0028-3908(95)00141-7. [DOI] [PubMed] [Google Scholar]
- 85.Stuart GJ, Dodt H-U, Sakmann B. Patch-clamp recordings from the soma and dendrites of neurons in brain slices using infrared videomicroscopy. Pflügers Arch. 1993;423:511–518. doi: 10.1007/BF00374949. [DOI] [PubMed] [Google Scholar]
- 86.Sutor B, ten Bruggencate G. Ascorbic acid: a useful reductant to avoid oxidation of catecholamines in electrophysiological experiments in vitro? Neurosci Lett. 1990;116:287–292. doi: 10.1016/0304-3940(90)90088-q. [DOI] [PubMed] [Google Scholar]
- 87.Tanaka E, North RA. Actions of 5-hydroxytryptamine on neurons of the rat cingulate cortex. J Neurophysiol. 1993;69:1749–1757. doi: 10.1152/jn.1993.69.5.1749. [DOI] [PubMed] [Google Scholar]
- 88.Tsubokawa H, Ross WN. IPSPs modulate back propagation and associated [Ca2+]i changes in dendrites of hippocampal CA1 pyramidal neurons. J Neurophysiol. 1996;76:2896–2906. doi: 10.1152/jn.1996.76.5.2896. [DOI] [PubMed] [Google Scholar]
- 89.Tork I. Anatomy of the serotonergic system. Ann NY Acad Sci. 1990;600:9–34. doi: 10.1111/j.1749-6632.1990.tb16870.x. [DOI] [PubMed] [Google Scholar]
- 90.Tukey JW. Exploratory data analysis. Addison-Wesley; Menlo Park, CA: 1977. [Google Scholar]
- 91.van Brederode JFM, Spain WJ. Differences in inhibitory synaptic input between layer II–III and layer V neurons of the cat neocortex. J Neurophysiol. 1995;74:1149–1166. doi: 10.1152/jn.1995.74.3.1149. [DOI] [PubMed] [Google Scholar]
- 92.van Brederode JFM, Foehring RC, Spain WJ. Morphological and electrophysiological properties of atypically oriented layer 2 pyramidal cells of the juvenile rat neocortex. Neuroscience. 2000;101:851–861. doi: 10.1016/s0306-4522(00)00430-9. [DOI] [PubMed] [Google Scholar]
- 93.van Brederode JFM, Rho JM, Cerne R, Tempel BL, Spain WJ. Evidence of altered inhibition in layer V pyramidal neurons from neocortex of KCNA1-null mice. Neuroscience. 2001;103:921–929. doi: 10.1016/s0306-4522(01)00041-0. [DOI] [PubMed] [Google Scholar]
- 94.Vizuete ML, Venero JL, Traiffort E, Vargas C, Machado A, Cano J. Expression of 5-HT7 receptor mRNA in rat brain during postnatal development. Neurosci Lett. 1997;227:53–56. doi: 10.1016/s0304-3940(97)00302-9. [DOI] [PubMed] [Google Scholar]
- 95.Vysokanov A, Flores-Hernandez J, Surmeier DJ. mRNAs for clozapine-sensitive receptors co-localize in rat prefrontal cortex neurons. Neurosci Lett. 1998;258:179–182. doi: 10.1016/s0304-3940(98)00882-9. [DOI] [PubMed] [Google Scholar]
- 96.Waterhouse BD, Mihailoff GA, Baack JC, Woodward DJ. Topographical distribution of dorsal and median raphe neurons projecting to motor, sensorimotor, and visual cortical areas in the rat. J Comp Neurol. 1986a;249:460–476. doi: 10.1002/cne.902490403. [DOI] [PubMed] [Google Scholar]
- 97.Waterhouse BD, Moises HC, Woodward DJ. Interaction of serotonin with somatosensory cortical neuronal responses to afferent synaptic inputs and putative neurotransmitters. Brain Res Bull. 1986b;17:507–518. doi: 10.1016/0361-9230(86)90218-2. [DOI] [PubMed] [Google Scholar]
- 98.White EL. Cortical circuits. Synaptic organization of the cerebral cortex. Structure, function and theory. Birkhauser; Boston: 1989. [Google Scholar]
- 99.Whittington MA, Traub RD, Kopell N, Ermentrout B, Buhl EH. Inhibition-based rhythms: experimental and mathematical observations on network dynamics. Int J Psychophysiol. 2000;38:315–336. doi: 10.1016/s0167-8760(00)00173-2. [DOI] [PubMed] [Google Scholar]
- 100.Williams GV, Rao SG, Goldman-Rakic P. The influence of 5-HT3 receptors on local circuit function in primate prefrontal cortex. Soc Neurosci Abstr. 1999;25:885. [Google Scholar]
- 101.Williams GV, Rao SG, Goldman-Rakic P. The physiological role of 5-HT2A receptors in working memory. J Neurosci. 2002;22:2843–2854. doi: 10.1523/JNEUROSCI.22-07-02843.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Willins DL, Deutsch AY, Roth BL. Serotonin 5-HT2A receptors are expressed on pyramidal neurons and interneurons in the rat cortex. Synapse. 1997;27:79–82. doi: 10.1002/(SICI)1098-2396(199709)27:1<79::AID-SYN8>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- 103.Wise SP, Jones EG. The organization and postnatal development of the commissural projection of the rat sometosensory cortex. J Comp Neurol. 1976;168:313–343. doi: 10.1002/cne.901680302. [DOI] [PubMed] [Google Scholar]
- 104.Wright DE, Seroogy KB, Lundgren KH, Davis BM, Jennes L. Comparative localization of serotonin 1A, 1C, and 2 receptor subtype mRNAs in rat brain. J Comp Neurol. 1995;351:357–373. doi: 10.1002/cne.903510304. [DOI] [PubMed] [Google Scholar]
- 105.Xiang Z, Huguenard JA, Prince DA. Serotonergic modulation of interneuronal functions in rat visual cortex. Soc Neurosci Abstr. 1999;25:439. [Google Scholar]
- 106.Yuste R, Gutnick MJ, Saar D, Delaney KR, Tank DW. Ca2+ accumulations in dendrites of neocortical pyramidal neurons: an apical band and evidence for two functional compartments. Neuron. 1994;13:23–43. doi: 10.1016/0896-6273(94)90457-x. [DOI] [PubMed] [Google Scholar]
- 107.Zhou F-M, Hablitz JJ. Layer I neurons of rat neocortex. I. Action potential and repetitive firing properties. J Neurophysiol. 1996a;76:651–667. doi: 10.1152/jn.1996.76.2.651. [DOI] [PubMed] [Google Scholar]
- 108.Zhou F-M, Hablitz JJ. Morphological properties of intracellularly labeled layer I neurons in rat neocortex. J Comp Neurol. 1996b;376:198–213. doi: 10.1002/(SICI)1096-9861(19961209)376:2<198::AID-CNE3>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- 109.Zhou FM, Hablitz JJ. Dopamine modulation of membrane and synaptic properties of interneurons in rat cerebral cortex. J Neurophysiol. 1999a;81:967–976. doi: 10.1152/jn.1999.81.3.967. [DOI] [PubMed] [Google Scholar]
- 110.Zhou FM, Hablitz JJ. Activation of serotonin receptors modulates synaptic transmission in rat cerebral cortex. J Neurophysiol. 1999b;82:2989–2999. doi: 10.1152/jn.1999.82.6.2989. [DOI] [PubMed] [Google Scholar]
- 111.Zifa E, Hernandez J, Fayolle C, Fillion G. Postnatal development of 5-HT1 receptors: [3H]5-HT binding sites and 5-HT induced ademylate cyclase activations in rat brain cortex. Brain Res Dev Brain Res. 1988;44:133–140. doi: 10.1016/0165-3806(88)90124-1. [DOI] [PubMed] [Google Scholar]