

Abstract
The most common type of epilepsy in adults is temporal lobe epilepsy. After epileptogenic injuries, dentate granule cell axons (mossy fibers) sprout and form new synaptic connections. Whether this synaptic reorganization strengthens recurrent inhibitory circuits or forms a novel recurrent excitatory circuit is unresolved. We labeled individual granule cells in vivo, reconstructed sprouted mossy fibers at the EM level, and identified postsynaptic targets with GABA immunocytochemistry in the pilocarpine model of temporal lobe epilepsy. Granule cells projected an average of 1.0 and 1.1 mm of axon into the granule cell and molecular layers, respectively. Axons formed an average of one synapse every 7 μm in the granule cell layer and every 3 μm in the molecular layer. Most synapses were with spines (76 and 98% in the granule cell and molecular layers, respectively). Almost all of the synapses were with GABA-negative structures (93 and 96% in the granule cell and molecular layers, respectively). By integrating light microscopic and EM data, we estimate that sprouted mossy fibers form an average of over 500 new synapses per granule cell, but <25 of the new synapses are with GABAergic interneurons. These findings suggest that almost all of the synapses formed by mossy fibers in the granule cell and molecular layers are with other granule cells. Therefore, after epileptogenic treatments that kill hilar mossy cells, mossy fiber sprouting does not simply replace one recurrent excitatory circuit with another. Rather, it replaces a distally distributed and disynaptic excitatory feedback circuit with one that is local and monosynaptic.
Keywords: temporal lobe epilepsy, mossy fibers, axon sprouting, electron microscopy, granule cell, GABA
Temporal lobe epilepsy is the most common type of epilepsy in adults (Engel et al., 1997). The recurrent excitation hypothesis of temporal lobe epilepsy proposes that, after epileptogenic injuries, granule cell axons (mossy fibers) reorganize and establish an abnormal recurrent excitatory circuit that generates seizure activity through positive feedback between granule cells (Nadler et al., 1980; Tauck and Nadler, 1985). Data supporting the recurrent excitation hypothesis include evidence that mossy fibers invade the granule cell layer and molecular layer of the dentate gyrus (regions mossy fibers normally avoid) in tissue from patients (de Lanerolle et al., 1989; Sutula et al., 1989; Houser et al., 1990; Babb et al., 1991; Isokawa et al., 1993; Franck et al., 1995; Masukawa et al., 1995; Zhang and Houser, 1999) and models of temporal lobe epilepsy (Cronin and Dudek, 1988; Sutula et al., 1988, 1998; Mello et al., 1993;Represa et al., 1993; Okazaki et al., 1995; Buckmaster and Dudek, 1997b; Kotti et al., 1997; Wenzel et al., 2000). Electron microscopic analyses suggest that at least some of the new synaptic contacts formed by sprouted mossy fibers are with granule cell dendrites (Frotscher and Zimmer, 1983; Babb et al., 1991; Represa et al., 1993; Franck et al., 1995; Okazaki et al., 1995; Zhang and Houser, 1999; Wenzel et al., 2000).
In contrast to the recurrent excitation hypothesis, the recurrent inhibition hypothesis proposes that sprouted mossy fibers preferentially synapse with inhibitory interneurons rather than with granule cells. The cell bodies and dendrites of inhibitory interneurons appear to be contacted by more mossy fiber terminals after axon reorganization (Sloviter, 1992; Kotti et al., 1997). If this hypothesis is correct, mossy fiber sprouting may be a homeostatic mechanism to control hyperexcitability by enhancing recurrent inhibition in the dentate gyrus.
Thus, currently available data are consistent with both the recurrent excitation and recurrent inhibition hypotheses. The net effect of mossy fiber sprouting will depend on the number of synapses made with each cell type: granule cells versus GABAergic interneurons. To address this issue, we measured the axon length and synaptic density and examined the ultrastructure and neurochemistry of the postsynaptic targets of sprouted mossy fibers in a model of temporal lobe epilepsy.
MATERIALS AND METHODS
Animals. All experiments were approved by the Stanford University Institutional Animal Care and Use Committee and performed in accordance with the National Institute of HealthGuide for the Care and Use of Laboratory Animals. Sprague Dawley male rats (2 months old) were treated with pilocarpine (380 mg/kg, i.p.) 20 min after atropine methylbromide (5 mg/kg, i.p.). Diazepam (10 mg/kg, i.p.) was administered 2–3 hr after the onset of status epilepticus and repeated as needed. Rats were video monitored for seizure activity 40 hr/week. Their first observed spontaneous seizure occurred 45 ± 7 d (mean ± SEM) after status epilepticus, and they were used in an experiment 45 ± 19 d after their first observed seizure.
Intracellular labeling. We chose to intracellularly label granule cells in vivo rather than in hippocampal slices, because the in vivo technique permits labeling of the entire axon arbor of the cell, and it provides superior tissue preservation for ultrastructural analyses and immunocytochemical labeling. The methods used for in vivo intracellular labeling of granule cells have been described previously (Buckmaster and Dudek, 1999). Cells were labeled with biocytin by passing 300 msec pulses of 0.1–0.3 nA hyperpolarizing current, 50% duty cycle, for an average of 16 min. The rat was then killed by urethane overdose (2.5 gm/kg, i.p.) and perfused through the ascending aorta at 30 ml/min with 0.9% NaCl for 1 min and 2.5% paraformaldehyde and 1% glutaraldehyde in 0.1 m phosphate buffer (PB), pH 7.4, for 30 min. The brain was removed, hemisected, postfixed overnight, and then cryoprotected in 30% sucrose in 0.1 m PB.
Tissue processing. The hippocampus was isolated, straightened, frozen, and sectioned perpendicular to the septotemporal axis with a microtome set at 40 μm. The methods used for the Timm's stain were similar to those described previously (Buckmaster and Dudek, 1997b). For biocytin processing, serial sections were collected in 0.1m Tris buffer (TB), pH 7.4, and treated with 1% sodium borohydride for 30 min and 1% hydrogen peroxide for 2 hr. After rinsing, sections were placed in blocking solution consisting of 2% bovine serum albumin (BSA), 0.25% DMSO, and 0.05m Tris-buffered saline (TBS), pH 7.4, for 1 hr. Sections incubated in avidin–biotin–horseradish peroxidase complex (1:250; Vector Laboratories, Burlingame, CA) in 0.5% BSA, 0.25% DMSO, and 0.05 m TBS at 4°C for 86 hr. After thorough washing in TB, sections were exposed to 0.04% diaminobenzidine and 0.05% NiCl for 15 min. Hydrogen peroxide was added to result in a 0.0025% solution, and sections were reacted for 1 hr. The reaction was stopped in washes of TB. Sections were postfixed with 1% OsO4 in sodium cacodylate buffer for 1 hr, dehydrated in a series of ethanols, placed in propylene oxide, gradually transferred to pure Araldite/Eponate-12 (Ted Pella, Redding, CA), and flat embedded between sheets of ACLAR at 60°C for 24 hr.
The axon arbors of nine granule cells were drawn with a camera lucida, and two-dimensional axon lengths were measured from the drawings with respect to position within strata of the dentate gyrus. Axon arbors of two of the nine cells were also reconstructed three-dimensionally with a Neurolucida system (MicroBrightField, Colchester, VT). Axon lengths measured two-dimensionally were adjusted for three-dimensionality by a correction factor (1.25), which was determined using values from the cells analyzed both ways. Axon lengths were adjusted for tissue shrinkage using previously determined shrinkage factors (1.06× in the transverse plane and 1.96× in the z-axis) (Buckmaster and Dudek, 1999).
Electron microscopy. After light microscopic analysis, biocytin-labeled axon segments in the granule cell layer and molecular layer were selected for ultrastructural analysis. Selected regions were remounted on a blank Araldite/Eponate-12 block. From each thick section, ∼500 serial ultrathin sections (80 nm) were made (Reichert Ultracut S; Leica, Vienna, Austria) and collected on single-slot nickel grids coated with support film. For postembedding GABA immunocytochemistry, sections were etched with 0.5% periodic acid for 30 min and 7% sodium metaperiodate for 30 min. Sections were exposed to blocking solution consisting of 0.8% ovalbumin and 5% fetal calf serum in 0.05 m TBS, pH 7.6, for 1 hr. After sections were incubated overnight in rabbit anti-GABA serum (1:80) in blocking solution, they were gently rinsed and then incubated in anti-rabbit colloidal gold (10 nm diameter, 1:80; Ted Pella) in 0.1% Triton X-100 and 0.05 m TB, pH 8.2, for 90 min. After rinsing, sections were stained with 2% uranyl acetate for 6 min and Sato's lead stain for 4 min. All chemicals and reagents were from Sigma (St. Louis, MO) unless specified otherwise.
Using a transmission electron microscope (Jeol 100CX; Jeol, Peabody, MA), the biocytin-labeled axon collateral and surrounding structures were photographed. Sections processed for postembedding GABA immunocytochemistry were checked for positive controls on a batch-by-batch basis. A batch of sections was included for immunocytochemical analysis only if specifically GABA-labeled aspiny dendrites were found in the molecular layer.
Axon segments were three-dimensionally reconstructed from electron micrographs using Neurolucida software (MicroBrightField) and a data tablet (Summagraphics, Seymour, CT). Low-magnification (1800–18,500×) prints at intervals of <50 sections were used to align biocytin-labeled profiles with surrounding landmarks, such as granule cell nuclei. Serial high-magnification (39,600–58,300×) prints were used to reconstruct axon segments. The number of high-magnification prints greatly exceeded the number of sections, because axon segments snaked up and down and sometimes extended longitudinally within a section so that montages of multiple prints had to be assembled. Axon length was measured only from reconstructed segments; gaps in the reconstruction were not included.
The criteria used to identify ultrastructural profiles have been described previously (Gray, 1959; Peters et al., 1991; Buckmaster et al., 1996, 2002). Briefly, synapses were identified by cleft material between parallel membranes of a vesicle-filled biocytin-labeled presynaptic element and a postsynaptic element with a postsynaptic density. Sometimes it was difficult to determine whether or not a synaptic contact was present, because the plane of section was not always perpendicular to the plane of the apposed cell membranes. In those cases, the specimen holder of the electron microscope was tilted to check suspected synapses, and ∼30% of them were synaptic contacts. Dendritic shafts were identified by morphological features of dendrites, including microtubules and mitochondria. Dendritic spines were identified by morphological features of spines, including a spine apparatus and continuity with a dendritic shaft.
RESULTS
Sprouted mossy fiber length
In control tissue, mossy fibers are confined to the hilus and CA3 region, but, in epileptic tissue, mossy fibers project into the granule cell layer and inner one-third of the molecular layer. This axon reorganization is revealed by the Timm's stain, which labels black the zinc-rich mossy fiber terminals (Fig. 1). Previous studies have shown that, in rat models of temporal lobe epilepsy, individual granule cells have sprouted mossy fiber collaterals that extend an average summed length of ∼1 mm in the granule cell layer and ∼1 mm in the molecular layer (Sutula et al., 1998; Buckmaster and Dudek, 1999). Our findings confirm these results. In six epileptic rats, nine granule cells were labeled: two in the superior blade, two at the apex, and five in the inferior blade of the granule cell layer. In the granule cell layer, the average axon length per cell was 1.01 ± 0.41 mm (mean ± SEM), and, in the molecular layer, it was 1.12 ± 0.59 mm. All of the labeled granule cells had only one primary axon arising from the soma. Therefore, all of the sprouted axons that projected into the granule cell layer and molecular layer were collaterals and not new primary axons. We cannot exclude the possibility that axon arbors were incompletely labeled with biocytin, but this seems unlikely. All of the cells had darkly labeled dendrites, dendritic spines, and axons, including fine collaterals and a primary branch that projected to the distal CA3 region.
Fig. 1.

Timm's-stained sections of the hippocampus in a control (a, c) and a pilocarpine-induced epileptic rat (b, d). Boxed regions in a and b are shown at higher magnification in c and d, respectively. Black mossy fiber terminals are evident in the granule cell layer (gcl) and inner one-third of the molecular layer (ml) in the epileptic but not the control rat.
Sprouted mossy fibers preferentially synapse with dendritic spines
Granule cells have spiny dendrites unlike many inhibitory interneurons in the dentate gyrus (Ramón y Cajal, 1995). If sprouted mossy fibers preferentially synapse with granule cells, one would expect to find a high proportion of synapses with dendritic spines. On the other hand, if sprouted mossy fibers preferentially synapse with inhibitory interneurons, one would expect to find a high proportion of synapses with dendritic shafts. To identify synapses and postsynaptic ultrastructure, segments of sprouted mossy fibers were reconstructed at the EM level (Fig. 2). Many synaptic contacts were found along the reconstructed axon segments. Although there was variability in the size of the postsynaptic densities, the vast majority of synapses formed by sprouted mossy fibers were clearly asymmetric (type 1) synaptic contacts (Fig. 3a). From six epileptic rats, eight sprouted mossy fibers were reconstructed for a summed length of 312 μm in the granule cell layer and 973 μm in the molecular layer (Fig. 4). In addition, three axon segments were examined for synaptic contacts but were not reconstructed. The proportion of synapses formed with dendritic spines versus dendritic shafts was measured (Table1). In the granule cell layer, 45 synaptic contacts were identified: 76% with dendritic spines and 24% with dendritic shafts. In the molecular layer, 471 synaptic contacts were identified: 93% with dendritic spines and 7% with dendritic shafts. The proportion of synapses formed with dendritic spines was significantly higher in the molecular layer than in the granule cell layer (p < 0.005; χ2 test). These findings suggest that sprouted mossy fibers preferentially synapse with spiny granule cells. However, some classes of inhibitory interneurons in the dentate gyrus do have spines (Halasy and Somogyi, 1993; Sik et al., 1996;Acsády et al., 1998; Buckmaster et al., 2002). Therefore, some of the synapses with spines might have been with inhibitory interneurons and not granule cells. In addition, granule cells could receive excitatory synaptic contacts on their dendritic shafts. Therefore, some of the synapses with shafts (Fig. 3b) might have been with spiny granule cells and not interneurons. To address these issues, we used postembedding GABA immunocytochemistry to evaluate the neurochemical identity of the synaptic targets of sprouted mossy fibers.
Fig. 2.
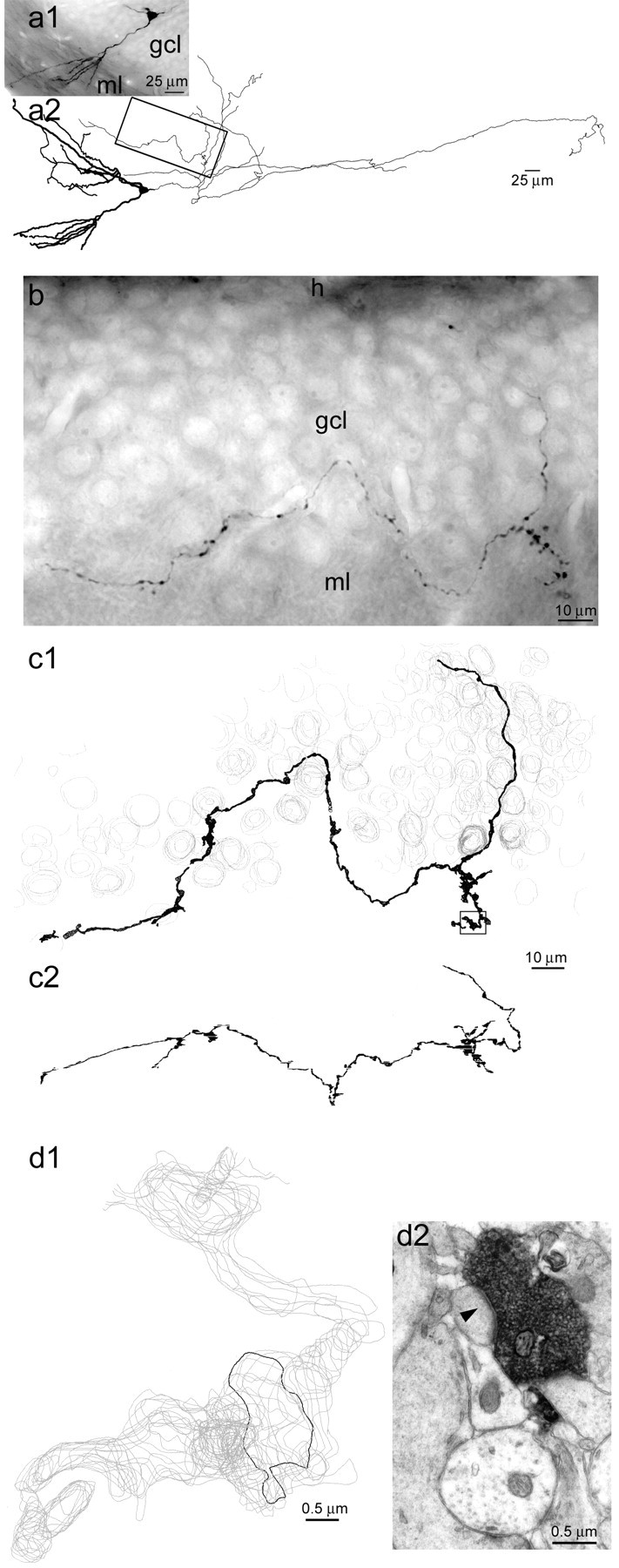
EM reconstruction of a sprouted mossy fiber in an epileptic rat. a, A granule cell labeled with biocytinin vivo in an epileptic rat. Photograph of section containing the soma (a1) and a light-microscopic reconstruction (a2) of the cell. Dendrites are thick; axon is thin. All of the axon shown in this reconstruction is within the dentate gyrus, and most is within the hilus. Several sections away from the soma, an axon collateral projected from the hilus through the granule cell layer and into the molecular layer. A segment of that axon collateral (box) was selected for reconstruction at the EM level.h, Hilus; gcl, granule cell layer;ml, molecular layer. b, Photograph of the selected axon segment. c1, EM reconstruction of the selected axon segment (black). Gray contours indicate cell nuclei that were used as landmarks. Thearea in the box is shown at high magnification in d. c2, Side view of three-dimensionally reconstructed axon segment demonstrating how the axon projected through the thickness of the section. d1, Magnified view of boxed region in c1demonstrates that the reconstructed axon segment consists of aligned serial contours (gray) that outlined biocytin-labeled axon profiles in serial electron micrographs. Theblack contour outlined the biocytin-labeled axon (black) in d2, which forms a synapse (arrowhead) with a dendritic spine.
Fig. 3.
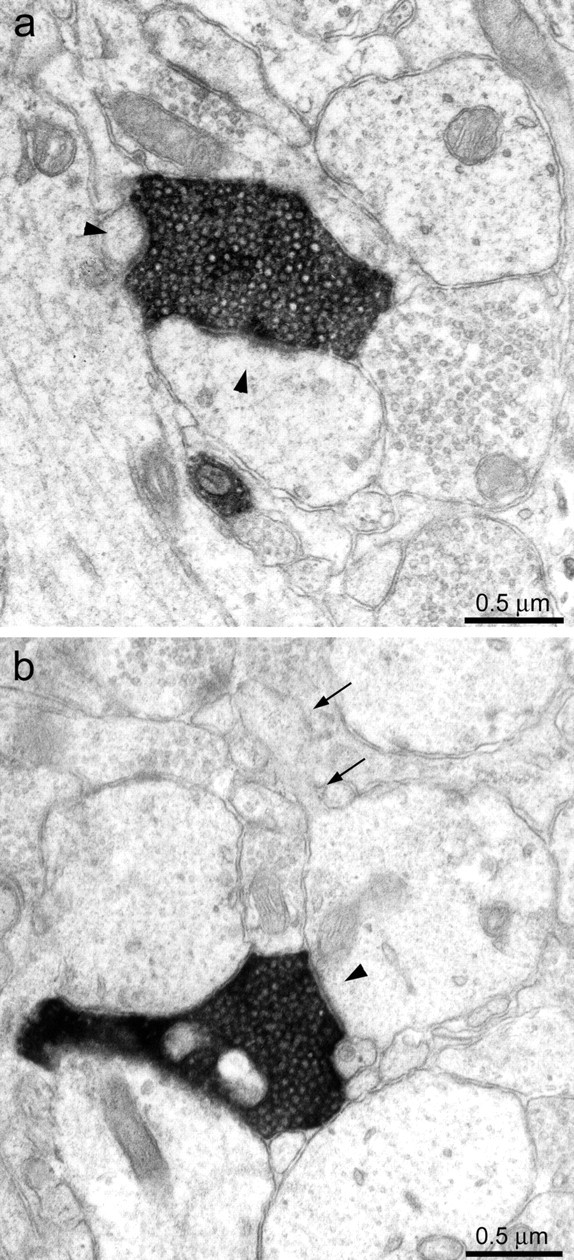
Sprouted mossy fibers synapsed with dendritic spines and dendritic shafts. a, A biocytin-labeled axon (black) in the molecular layer formed synapses (arrowheads) with a large and a small spine.b, A biocytin-labeled axon (black) in the molecular layer formed a synapse (arrowhead) with the shaft of a spiny (arrows) dendrite.
Fig. 4.

Sprouted mossy fibers synapsed preferentially with dendritic spines. Reconstructed sprouted mossy fibers with synaptic contacts indicated by markers. Squaresindicate that the postsynaptic target was a dendritic spine;circles indicate a dendritic shaft. The identity of each reconstruction (rat and segment) corresponds to Table 1. Borders between strata (h, hilus; gcl, granule cell layer; ml, molecular layer) are indicated bylines.
Table 1.
EM-reconstructed sprouted mossy fibers in epileptic rats
| Rat and segment | Molecular layer (#synapses) | Granule cell layer (# synapses) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Length (μm) | Spine | Shaft | GABA negative | GABA positive | Length (μm) | Spine | Shaft | GABA negative | GABA positive | |
| 1A | 84 | 32 | 4 | 32 | 4 | 42 | 6 | 0 | 6 | 0 |
| 1B | 297 | 98 | 11 | 99 | 3 | 122 | 14 | 3 | 16 | 1 |
| 2 | 97 | 41 | 0 | 40 | 1 | 9 | 1 | 1 | 1 | 1 |
| 3 | 107 | 32 | 4 | 30 | 4 | – | – | – | – | – |
| 4A | 157 | 23 | 3 | – | – | 79 | 7 | 2 | – | – |
| 4B | 41 | 13 | 2 | – | – | – | – | – | – | – |
| 5A | 97 | 20 | 3 | – | – | – | – | – | – | – |
| 6A | 93 | 28 | 4 | – | – | 60 | 2 | 4 | – | – |
| 6B | – | 70 | 3 | 73 | 0 | – | 2 | 1 | 3 | 0 |
| 5B | – | 33 | 0 | 33 | 0 | – | 2 | 0 | 2 | 0 |
| 4C | – | 47 | 0 | 46 | 1 | – | – | – | – | – |
| Total | 973 | 437 | 34 | 353 | 13 | 312 | 34 | 11 | 28 | 2 |
Sprouted mossy fibers preferentially synapse with GABA-negative dendritic spines
Inhibitory interneurons synthesize and express the neurotransmitter GABA (Ribak et al., 1978; Sloviter and Nilaver, 1987), and their cell bodies and dendrites are GABA-positive (Halasy and Somogyi, 1993). Granule cells usually are not GABA positive (Sloviter and Nilaver, 1987), but, under special circumstances, they may be (Sloviter et al., 1996). In our material, all granule cells appeared to be GABA-negative. However, if some did have GABA-positive dendrites, we would have overestimated the number of synapses formed by sprouted mossy fibers with interneurons. Specifically GABA-labeled aspiny dendrites were identified in the molecular layer of each batch of immunolabeled sections. In addition, whenever GABA-negative postsynaptic targets were identified, other GABA-positive structures were evident in the same section (Fig.5a). Adjacent sections were examined to verify neurochemical identity of postsynaptic targets. These positive controls reduced the likelihood of false-negative results.
Fig. 5.
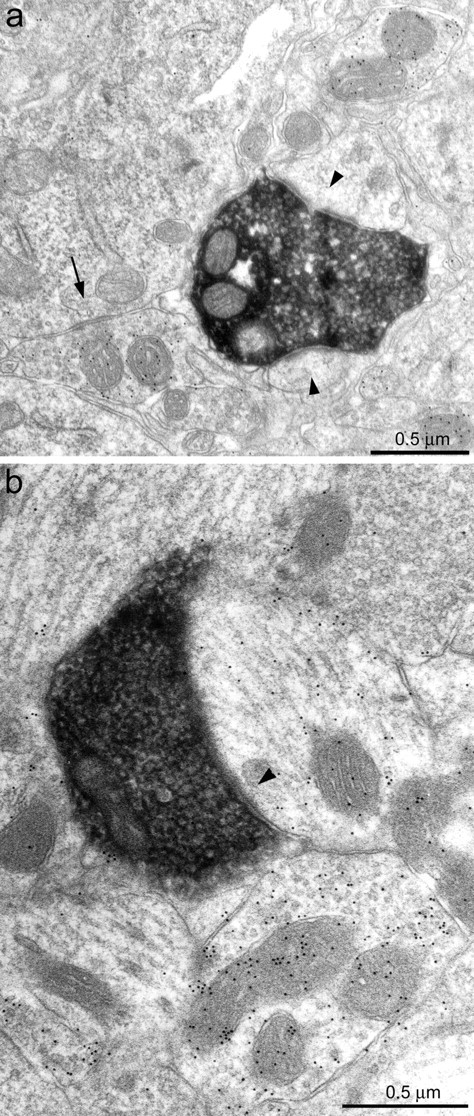
Sprouted mossy fibers synapsed with GABA-negative and GABA-positive targets. a, A biocytin-labeled axon (black) formed synaptic contacts (arrowheads) with two GABA-negative spines. Nearby GABA-positive structures were labeled with 10-nm-diameter colloidal gold particles, and a GABA-positive axon terminal formed a symmetric synapse (arrow) with a granule cell body.b, A biocytin-labeled axon (black) in the molecular layer formed a synaptic contact (arrowhead) with a GABA-positive dendritic shaft.
Sprouted mossy fibers formed asymmetric synaptic contacts with GABA-negative and GABA-positive postsynaptic targets (Fig. 5). The proportion of synapses formed with GABA-negative versus GABA-positive targets was measured from a subset of the reconstructed sprouted mossy fibers described above (Table 1). From three epileptic rats, four reconstructed sprouted mossy fibers had a summed length of 173 μm in the granule cell layer and 585 μm in the molecular layer (Fig.6). In addition, another three axon segments from three different rats were examined for synaptic contacts but were not reconstructed. In the granule cell layer, 30 synaptic contacts were identified: 93% with GABA-negative targets and 7% with GABA-positive targets. In the molecular layer, 366 synaptic contacts were identified: 96% with GABA-negative targets and 4% with GABA-positive targets. These findings indicate that sprouted mossy fibers preferentially synapse with GABA-negative dendritic spines. For sprouted mossy fibers in both the granule cell layer and molecular layer, 67% of the postsynaptic dendritic shafts and 98% of the postsynaptic dendritic spines were GABA-negative.
Fig. 6.

Sprouted mossy fibers synapsed preferentially with GABA-negative dendritic spines. Reconstructed sprouted mossy fibers with synaptic contacts indicated by markers.Squares indicate that the postsynaptic target was a dendritic spine; circles indicate a dendritic shaft.Open markers indicate that the postsynaptic target was GABA-negative; filled markers indicate GABA-positive. The identity of each reconstruction (rat and segment) corresponds to Table 1. Borders between strata (h, hilus;gcl, granule cell layer; ml, molecular layer) are indicated by lines.
Number of new synapses
Synaptic density was measured from the EM reconstructions of sprouted mossy fibers (Figs. 4, 6). For the eight reconstructed axon segments in the molecular layer, an average of one synapse was formed per 3 μm (0.17–0.43 synapses/μm). For the five reconstructed axon segments in the granule cell layer, an average of one synapse was formed every 7 μm (0.10–0.22 synapses/μm). Synaptic density was significantly higher in the molecular layer than in the granule cell layer (p < 0.002; t test). Most presynaptic profiles formed only one synapse, but it was not uncommon to find a large presynaptic bouton that formed multiple synapses. Only very rarely could we verify multiple synaptic contacts between a sprouted mossy fiber and a postsynaptic neuron. However, this was difficult to determine, because postsynaptic neurons were not labeled individually.
Integrating our light microscopic data on sprouted mossy fiber length per granule cell with our EM data on synaptic density, we estimate that the average granule cell formed 140 new synapses in the granule cell layer and 370 new synapses in the molecular layer. Furthermore, we estimate that >88% of the new synapses were with dendritic spines and >95% were with GABA-negative targets.
DISCUSSION
Previous studies provide evidence that sprouted mossy fibers synapse with spines of excitatory granule cells (Frotscher and Zimmer, 1983; Represa et al., 1993; Franck et al., 1995; Okazaki et al., 1995;Zhang and Houser, 1999; Wenzel et al., 2000). However, other studies suggest that sprouted mossy fibers preferentially synapse with inhibitory interneurons (Sloviter, 1992; Kotti et al., 1997). We addressed this unsettled issue by intracellularly labeling granule cells in epileptic rats in vivo, reconstructing sprouted axon segments at the EM level, identifying postsynaptic targets with GABA immunocytochemistry, and quantifying the number of new synapses formed by sprouted mossy fibers. We found that sprouted mossy fibers synapse almost exclusively with GABA-negative dendritic spines.
Granule cells are the predominant synaptic target of sprouted mossy fibers
It is highly likely that sprouted mossy fibers synapsed with the GABA-negative dendritic spines of granule cells. Granule cells are the predominant GABA-negative neurons in the dentate gyrus, and their spiny dendrites project through the granule cell layer and into the molecular layer in which they extend and ramify (Ramón y Cajal, 1995). Hilar mossy cells, like granule cells, are GABA-negative (Soriano and Frotscher, 1994), but, in rats, mossy cell dendrites are mostly confined to the hilus and only occasionally extend into the molecular layer (Amaral, 1978). There are GABA-negative cholinergic neurons in the dentate gyrus, but they are extremely rare (Frotscher et al., 2000). Therefore, granule cells account for the vast majority of GABA-negative neuronal structures in the granule cell layer and molecular layer.
Only 4% of the synaptic contacts examined were with GABA-immunoreactive neurons. This finding suggests that sprouted mossy fibers only occasionally synapsed with inhibitory interneurons. The Timm's stain has been used to demonstrate appositions and synaptic contacts between mossy fiber terminals and basket cells in the granule cell layer of control (Ribak and Peterson, 1991) and epileptic (Sloviter, 1992; Kotti et al., 1997) rats. Qualitatively, it appears that basket cells receive more synaptic input from Timm's-positive terminals after mossy fiber sprouting (Sloviter, 1992; Kotti et al., 1997). Quantitative analyses are needed, but this finding suggests that basket cells may receive more direct synaptic input from granule cells after mossy fiber sprouting. However, it does not necessarily follow that basket cells receive more excitatory synaptic contacts, because Timm's-positive inputs may have only replaced Timm's-negative excitatory inputs that were lost during epileptogenic injuries. Regardless of the changes in excitatory synaptic input to basket cells, our findings suggest that, for each new synapse formed by a sprouted mossy fiber with a GABA-positive neuron, >20 new synapses are formed with granule cells.
Functional implications
Our findings support the hypothesis that granule cell axon reorganization produces a novel recurrent excitatory network that might generate seizures in patients and models of temporal lobe epilepsy (Nadler et al., 1980; Tauck and Nadler, 1985). According to that hypothesis, sprouted mossy fibers replace the excitatory synaptic input to granule cell proximal dendrites that is lost after hilar mossy cells are killed by epileptogenic injuries. Mossy cells are the predominant neuron in the hilus (Amaral, 1978; Buckmaster and Jongen-Rêlo, 1999). In patients and models of temporal lobe epilepsy, the loss of hilar neurons is correlated with the extent of mossy fiber sprouting (Babb et al., 1991; Masukawa et al., 1995; Buckmaster and Dudek, 1997b;Nissinen et al., 2001). Like sprouted mossy fibers, axons of mossy cells are glutamatergic, and they synapse almost exclusively with GABA-negative dendritic spines in the granule cell layer and molecular layer (Buckmaster et al., 1996; Wenzel et al., 1997). However, sprouted mossy fibers project locally (within 400 μm of the parent cell body) (Sutula et al., 1998; Buckmaster and Dudek, 1999), whereas mossy cell axons project distally (beyond 600 μm of the parent cell body) before making the vast majority of their synaptic contacts (Buckmaster et al., 1996). Therefore, the loss of mossy cells and the sprouting of mossy fibers does not simply replace one recurrent excitatory network with another. Rather, it replaces a distally distributed and disynaptic excitatory feedback circuit with one that is local and monosynaptic. Previous slice experiments provide functional evidence of recurrent excitation between granule cells after mossy fiber sprouting in models of temporal lobe epilepsy (Wuarin and Dudek, 1996, 2001; Molnár and Nadler, 1999; Lynch and Sutula, 2000). In addition, mossy fiber sprouting brings synaptically releasable zinc into the granule cell layer and molecular layer where it might diffuse to inhibitory synapses and impair GABAA receptor-mediated inhibition (Buhl et al., 1996; Shumate et al., 1998).
The consequences of synaptic reorganization in the dentate gyrus may be especially dire. It has been proposed that normally the dentate gyrus acts like a gate and prevents seizures by filtering neuronal activity between highly seizurogenic regions in the hippocampus and entorhinal cortex (Stringer et al., 1989; Lothman et al., 1991). In kainate-induced epileptic rats, however, the threshold for maximal dentate activation is increased, not decreased (Buckmaster and Dudek, 1997a). Nevertheless, the formation of a novel excitatory feedback circuit between granule cells may reduce the filtering capability of the dentate and increase its propensity to amplify neuronal activity and propagate seizures through the hippocampus and to other limbic structures.
Our findings support the recurrent excitation hypothesis, but many questions persist. Mossy fiber sprouting apparent as aberrant Timm's staining in the granule cell layer and molecular layer is a common finding in patients and models of temporal lobe epilepsy. The extent of aberrant Timm's staining, however, does not correlate with seizure frequency (Buckmaster and Dudek, 1997b; Timofeeva and Peterson, 1999;Nissinen et al., 2001). Longo and Mello (1997, 1998) reported that, in models of temporal lobe epilepsy, treatment with cycloheximide blocks mossy fiber sprouting, but the rats develop epilepsy nevertheless. However, other investigators have not been able to replicate that controversial result (Williams et al., 2002). Mossy fiber sprouting is likely to be one of many factors that contribute to temporal lobe epileptogenesis. Some other likely contributors include changes in neurotransmitter expression by granule cells (Shumate et al., 1998), altered patterns of synaptic input from the entorhinal cortex after layer III neuron loss (Du et al., 1993), and the loss of GABAergic interneurons in the dentate gyrus (de Lanerolle et al., 1989; Sloviter et al., 1991; Mathern et al., 1995; Zhu et al., 1997; Maglóczky et al., 2000).
Another persistent question is the effect of mossy fiber sprouting within the hilus. The summed length per granule cell of mossy fiber collaterals within the hilus is greater in epileptic versus control rats (Sutula et al., 1998; Buckmaster and Dudek, 1999; Wenzel et al., 2000). Within the hilus of control rats, there are a variety of different cell types that mossy fibers might synapse with (Amaral, 1978). In patients (de Lanerolle et al., 1989; Sloviter et al., 1991;Mathern et al., 1995; Zhu et al., 1997; Maglóczky et al., 2000) and models of temporal lobe epilepsy (Sloviter, 1987; Obenaus et al., 1993; Buckmaster and Dudek, 1997b; Buckmaster and Jongen-Rêlo, 1999), hilar neuron loss changes the number and proportion of synaptic targets available to mossy fibers within the hilus. The formation of novel basal dendrites by granule cells (Spigelman et al., 1998;Buckmaster and Dudek, 1999) provides another synaptic target within the hilus and another avenue for excitatory feedback between granule cells (Ribak et al., 2000). Future studies could address these issues by reconstructing mossy fibers in the hilus and identifying their postsynaptic targets.
The present study shows that sprouting mossy fibers synapse almost exclusively with excitatory neurons in the granule cell layer and molecular layer of the dentate gyrus. Lesioning the synaptic input from the entorhinal cortex to granule cells also triggers mossy fiber sprouting and synaptogenesis in adult rats (Laurberg and Zimmer, 1981;Frotscher and Zimmer, 1983). A variety of experimental treatments that produce epilepsy also induce axon sprouting in other brain regions (Salin et al., 1995; Perez et al., 1996; McKinney et al., 1997;Esclapez et al., 1999). These findings highlight the remarkable plasticity of the adult CNS and suggest that the formation of novel recurrent excitatory circuits may be a common contributing factor to epileptogenesis.
Footnotes
This work was supported by National Institutes of Health/National Institute of Neurological Disorders and Stroke Grants NS40276 and NS39110.
Correspondence should be addressed to Paul Buckmaster, 300 Pasteur Drive, R102 Edwards Building, Department of Comparative Medicine, Stanford University, Stanford, CA 94305-5330. E-mail:psb@stanford.edu.
REFERENCES
- 1.Acsády L, Kamondi A, Sik A, Freund T, Buzsáki G. GABAergic cells are the major postsynaptic target of mossy fibers in the rat hippocampus. J Neurosci. 1998;18:3386–3403. doi: 10.1523/JNEUROSCI.18-09-03386.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Amaral DG. A Golgi study of cell types in the hilar region of the hippocampus in the rat. J Comp Neurol. 1978;182:851–914. doi: 10.1002/cne.901820508. [DOI] [PubMed] [Google Scholar]
- 3.Babb TL, Kupfer WR, Pretorius JK, Crandall PH, Levesque MF. Synaptic reorganization by mossy fibers in human epileptic fascia dentata. Neuroscience. 1991;42:351–363. doi: 10.1016/0306-4522(91)90380-7. [DOI] [PubMed] [Google Scholar]
- 4.Buckmaster PS, Dudek FE. Network properties of the dentate gyrus in epileptic rats with hilar neuron loss and granule cell axon reorganization. J Neurophysiol. 1997a;77:2685–2696. doi: 10.1152/jn.1997.77.5.2685. [DOI] [PubMed] [Google Scholar]
- 5.Buckmaster PS, Dudek FE. Neuron loss, granule cell axon reorganization, and functional changes in the dentate gyrus of epileptic kainate-treated rats. J Comp Neurol. 1997b;385:385–404. [PubMed] [Google Scholar]
- 6.Buckmaster PS, Dudek FE. In vivo intracellular analysis of granule cell axon reorganization in epileptic rats. J Neurophysiol. 1999;81:712–721. doi: 10.1152/jn.1999.81.2.712. [DOI] [PubMed] [Google Scholar]
- 7.Buckmaster PS, Jongen-Rêlo AL. Highly specific neuron loss preserves lateral inhibitory circuits in the dentate gyrus of kainate-induced epileptic rats. J Neurosci. 1999;19:9519–9529. doi: 10.1523/JNEUROSCI.19-21-09519.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Buckmaster PS, Wenzel HJ, Kunkel DD, Schwartzkroin PA. Axon arbors and synaptic connections of hippocampal mossy cells in the rat in vivo. J Comp Neurol. 1996;366:270–292. doi: 10.1002/(sici)1096-9861(19960304)366:2<270::aid-cne7>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- 9.Buckmaster PS, Yamawaki R, Zhang GF. Axon arbors and synaptic connections of a vulnerable population of interneurons in the dentate gyrus in vivo. J Comp Neurol. 2002;445:360–373. doi: 10.1002/cne.10183. [DOI] [PubMed] [Google Scholar]
- 10.Buhl EH, Otis TS, Mody I. Zinc-induced collapse of augmented inhibition by GABA in a temporal lobe epilepsy model. Science. 1996;271:369–373. doi: 10.1126/science.271.5247.369. [DOI] [PubMed] [Google Scholar]
- 11.Cronin J, Dudek FE. Chronic seizures and collateral sprouting of dentate mossy fibers after kainic acid treatment in rats. Brain Res. 1988;474:181–184. doi: 10.1016/0006-8993(88)90681-6. [DOI] [PubMed] [Google Scholar]
- 12.de Lanerolle NC, Kim JH, Robbins RJ, Spencer DD. Hippocampal interneuron loss and plasticity in human temporal lobe epilepsy. Brain Res. 1989;495:387–395. doi: 10.1016/0006-8993(89)90234-5. [DOI] [PubMed] [Google Scholar]
- 13.Du F, Whetsell WO, Jr, Abou-Khalil B, Blumenkopf B, Lothman EW, Schwarcz R. Preferential neuronal loss in layer III of the entorhinal cortex in patients with temporal lobe epilepsy. Epilepsy Res. 1993;16:223–233. doi: 10.1016/0920-1211(93)90083-j. [DOI] [PubMed] [Google Scholar]
- 14.Engel J, Jr, Williamson PD, Wieser H-G. Mesial temporal lobe epilepsy. In: Engel J Jr, Pedley TA, editors. Epilepsy: a comprehensive textbook. Lippincott-Raven; Philadelphia: 1997. pp. 2417–2426. [Google Scholar]
- 15.Esclapez M, Hirsch JC, Ben-Ari Y, Bernard C. Newly formed excitatory pathways provide a substrate for hyperexcitability in experimental temporal lobe epilepsy. J Comp Neurol. 1999;408:449–460. doi: 10.1002/(sici)1096-9861(19990614)408:4<449::aid-cne1>3.0.co;2-r. [DOI] [PubMed] [Google Scholar]
- 16.Franck JE, Pokorny J, Kunkel DD, Schwartzkroin PA. Physiologic and morphologic characteristics of granule cell circuitry in human epileptic hippocampus. Epilepsia. 1995;36:543–558. doi: 10.1111/j.1528-1157.1995.tb02566.x. [DOI] [PubMed] [Google Scholar]
- 17.Frotscher M, Zimmer J. Lesion-induced mossy fibers to the molecular layer of the rat fascia dentata: identification of postsynaptic granule cells by the Golgi-EM technique. J Comp Neurol. 1983;215:299–311. doi: 10.1002/cne.902150306. [DOI] [PubMed] [Google Scholar]
- 18.Frotscher M, Vida I, Bender R. Evidence for the existence of non-GABAergic, cholinergic interneurons in the rodent hippocampus. Neuroscience. 2000;96:27–31. doi: 10.1016/s0306-4522(99)00525-4. [DOI] [PubMed] [Google Scholar]
- 19.Gray EG. Axo-somatic and axo-dendritic synapses of the cerebral cortex. J Anat. 1959;93:420–433. [PMC free article] [PubMed] [Google Scholar]
- 20.Halasy K, Somogyi P. Subdivisions in the multiple GABAergic innervation of granule cells in the dentate gyrus of the rat hippocampus. Eur J Neurosci. 1993;5:411–429. doi: 10.1111/j.1460-9568.1993.tb00508.x. [DOI] [PubMed] [Google Scholar]
- 21.Houser CR, Miyashiro JE, Swartz BE, Walsh GO, Rich JR, Delgado-Escueta AV. Altered patterns of dynorphin immunoreactivity suggest mossy fiber reorganization in human hippocampal epilepsy. J Neurosci. 1990;10:267–282. doi: 10.1523/JNEUROSCI.10-01-00267.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Isokawa M, Levesque MF, Babb TL, Engel J., Jr Single mossy fiber axonal systems of human dentate granule cells studied in hippocampal slices from patients with temporal lobe epilepsy. J Neurosci. 1993;13:1511–1522. doi: 10.1523/JNEUROSCI.13-04-01511.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kotti T, Riekkinen PJ, Miettinen R. Characterization of target cells for aberrant mossy fiber collaterals in the dentate gyrus of epileptic rat. Exp Neurol. 1997;146:323–330. doi: 10.1006/exnr.1997.6553. [DOI] [PubMed] [Google Scholar]
- 24.Laurberg S, Zimmer J. Lesion-induced sprouting of hippocampal mossy fiber collaterals to the fascia dentata in developing and adult rats. J Comp Neurol. 1981;200:433–459. doi: 10.1002/cne.902000310. [DOI] [PubMed] [Google Scholar]
- 25.Longo BM, Mello LEAM. Blockade of pilocarpine- or kainate-induced mossy fiber sprouting by cycloheximide does not prevent subsequent epileptogenesis in rats. Neurosci Lett. 1997;226:163–166. doi: 10.1016/s0304-3940(97)00267-x. [DOI] [PubMed] [Google Scholar]
- 26.Longo BM, Mello LEAM. Supragranular mossy fiber sprouting is not necessary for spontaneous seizures in the intrahippocampal kainate model of epilepsy in the rat. Epilepsy Res. 1998;32:172–182. doi: 10.1016/s0920-1211(98)00049-7. [DOI] [PubMed] [Google Scholar]
- 27.Lothman EW, Bertram EH, III, Stringer JL. Functional anatomy of hippocampal seizures. Prog Neurobiol. 1991;37:1–82. doi: 10.1016/0301-0082(91)90011-o. [DOI] [PubMed] [Google Scholar]
- 28.Lynch M, Sutula T. Recurrent excitatory connectivity in the dentate gyrus of kindled and kainic acid-treated rats. J Neurophysiol. 2000;83:693–704. doi: 10.1152/jn.2000.83.2.693. [DOI] [PubMed] [Google Scholar]
- 29.Maglóczky ZS, Wittner L, Borhegyi ZS, Halász P, Vajda J, Czirják S, Freund TF. Changes in the distribution and connectivity of interneurons in the epileptic human dentate gyrus. Neuroscience. 2000;96:7–25. doi: 10.1016/s0306-4522(99)00474-1. [DOI] [PubMed] [Google Scholar]
- 30.Masukawa LM, O'Connor WM, Lynott J, Burdette LJ, Uruno K, McGonigle P, O'Connor MJ. Longitudinal variation in cell density and mossy fiber reorganization in the dentate gyrus from temporal lobe epileptic patients. Brain Res. 1995;678:65–75. doi: 10.1016/0006-8993(95)00167-o. [DOI] [PubMed] [Google Scholar]
- 31.Mathern GW, Babb TL, Pretorius JK, Leite JP. Reactive synaptogenesis and neuron densities for neuropeptide Y, somatostatin, and glutamate decarboxylase immunoreactivity in the epileptogenic human fascia dentata. J Neurosci. 1995;15:3990–4004. doi: 10.1523/JNEUROSCI.15-05-03990.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.McKinney RA, Debanne D, Gähwiler BH, Thompson SM. Lesion-induced axonal sprouting and hyperexcitability in the hippocampus in vitro: implications for the genesis of posttraumatic epilepsy. Nat Med. 1997;3:990–996. doi: 10.1038/nm0997-990. [DOI] [PubMed] [Google Scholar]
- 33.Mello LEAM, Cavalheiro EA, Tan AM, Kupfer WR, Pretorius JK, Babb TL, Finch DM. Circuit mechanisms of seizures in the pilocarpine model of chronic epilepsy: cell loss and mossy fiber sprouting. Epilepsia. 1993;34:985–995. doi: 10.1111/j.1528-1157.1993.tb02123.x. [DOI] [PubMed] [Google Scholar]
- 34.Molnár P, Nadler JV. Mossy fiber–granule cell synapses in the normal and epileptic rat dentate gyrus studied with minimal laser photostimulation. J Neurophysiol. 1999;82:1883–1894. doi: 10.1152/jn.1999.82.4.1883. [DOI] [PubMed] [Google Scholar]
- 35.Nadler JV, Perry BW, Cotman CW. Selective reinnervation of hippocampal area CA1 and the fascia dentata after destruction of CA3-CA4 afferents with kainic acid. Brain Res. 1980;182:1–9. doi: 10.1016/0006-8993(80)90825-2. [DOI] [PubMed] [Google Scholar]
- 36.Nissinen J, Lukasiuk K, Pitkänen A. Is mossy fiber sprouting present at the time of the first spontaneous seizures in rat experimental temporal lobe epilepsy? Hippocampus. 2001;11:299–310. doi: 10.1002/hipo.1044. [DOI] [PubMed] [Google Scholar]
- 37.Obenaus A, Esclapez M, Houser CR. Loss of glutamate decarboxylase mRNA-containing neurons in the rat dentate gyrus following pilocarpine-induced seizures. J Neurosci. 1993;13:4470–4485. doi: 10.1523/JNEUROSCI.13-10-04470.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Okazaki MM, Evenson DA, Nadler JV. Hippocampal mossy fiber sprouting and synapse formation after status epilepticus in rats: visualization after retrograde transport of biocytin. J Comp Neurol. 1995;352:515–534. doi: 10.1002/cne.903520404. [DOI] [PubMed] [Google Scholar]
- 39.Perez Y, Morin F, Beaulieu C, Lacaille J-C. Axonal sprouting of CA1 pyramidal cells in hyperexcitable hippocampal slices of kainate-treated rats. Eur J Neurosci. 1996;8:736–748. doi: 10.1111/j.1460-9568.1996.tb01259.x. [DOI] [PubMed] [Google Scholar]
- 40.Peters A, Palay S, Webster HD. The fine structure of the nervous system, Ed 3, pp 138–211. Oxford UP; New York: 1991. [Google Scholar]
- 41.Ramón y Cajal S. Histology of the nervous system of man and vertebrates, Vol 2 (Swanson N, Swanson LW, translators), pp 614–625. Oxford UP; New York: 1995. [Google Scholar]
- 42.Represa A, Jorquera I, Le Gal La Salle G, Ben-Ari Y. Epilepsy induced collateral sprouting of hippocampal mossy fibers: does it induce the development of ectopic synapses with granule cell dendrites? Hippocampus. 1993;3:257–268. doi: 10.1002/hipo.450030303. [DOI] [PubMed] [Google Scholar]
- 43.Ribak CE, Peterson GM. Intragranular mossy fibers in rats and gerbils form synapses with the somata and proximal dendrites of basket cells in the dentate gyrus. Hippocampus. 1991;1:355–364. doi: 10.1002/hipo.450010403. [DOI] [PubMed] [Google Scholar]
- 44.Ribak CE, Vaughn JE, Saito K. Immunocytochemical localization of glutamic acid decarboxylase in neuronal somata following colchicine inhibition of axonal transport. Brain Res. 1978;140:315–332. doi: 10.1016/0006-8993(78)90463-8. [DOI] [PubMed] [Google Scholar]
- 45.Ribak CE, Tran PH, Spigelman I, Okazaki MM, Nadler JV. Status epilepticus-induced hilar basal dendrites on rodent granule cells contribute to recurrent excitatory circuitry. J Comp Neurol. 2000;428:240–253. doi: 10.1002/1096-9861(20001211)428:2<240::aid-cne4>3.0.co;2-q. [DOI] [PubMed] [Google Scholar]
- 46.Salin P, Tseng G-F, Hoffman S, Parada I, Prince DA. Axonal sprouting in layer V pyramidal neurons of chronically injured cerebral cortex. J Neurosci. 1995;15:8234–8245. doi: 10.1523/JNEUROSCI.15-12-08234.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Shumate MD, Lin DD, Gibbs JW, III, Holloway KL, Coulter DA. GABAA receptor function in epileptic human dentate granule cells: comparison to epileptic and control rat. Epilepsy Res. 1998;32:114–128. doi: 10.1016/s0920-1211(98)00045-x. [DOI] [PubMed] [Google Scholar]
- 48.Sik A, Penttonen M, Buzsáki G. Interneurons in the hippocampal dentate gyrus: an in vivo intracellular study. Eur J Neurosci. 1996;9:573–588. doi: 10.1111/j.1460-9568.1997.tb01634.x. [DOI] [PubMed] [Google Scholar]
- 49.Sloviter RS. Decreased hippocampal inhibition and a selective loss of interneurons in experimental epilepsy. Science. 1987;235:73–76. doi: 10.1126/science.2879352. [DOI] [PubMed] [Google Scholar]
- 50.Sloviter RS. Possible functional consequences of synaptic reorganization in the dentate gyrus of kainate-treated rats. Neurosci Lett. 1992;137:91–96. doi: 10.1016/0304-3940(92)90306-r. [DOI] [PubMed] [Google Scholar]
- 51.Sloviter RS, Nilaver G. Immunocytochemical localization of GABA-, cholecystokinin-, vasoactive intestinal polypeptide-, and somatostatin-like immunoreactivity in the area dentata and hippocampus of the rat. J Comp Neurol. 1987;256:42–60. doi: 10.1002/cne.902560105. [DOI] [PubMed] [Google Scholar]
- 52.Sloviter RS, Sollas AL, Barbaro NM, Laxer KD. Calcium-binding protein (calbindin-D28K) and parvalbumin immunocytochemistry in the normal and epileptic human hippocampus. J Comp Neurol. 1991;308:381–396. doi: 10.1002/cne.903080306. [DOI] [PubMed] [Google Scholar]
- 53.Sloviter RS, Dichter MA, Rachinsky TL, Dean E, Goodman JH, Sollas AL, Martin DL. Basal expression and induction of glutamate decarboxylase and GABA in excitatory granule cells of the rat and monkey hippocampal dentate gyrus. J Comp Neurol. 1996;373:593–618. doi: 10.1002/(SICI)1096-9861(19960930)373:4<593::AID-CNE8>3.0.CO;2-X. [DOI] [PubMed] [Google Scholar]
- 54.Soriano E, Frotscher M. Mossy cells of the rat fascia dentata are glutamate-immunoreactive. Hippocampus. 1994;4:65–69. doi: 10.1002/hipo.450040108. [DOI] [PubMed] [Google Scholar]
- 55.Spigelman I, Yan X-X, Obenaus A, Lee Y-S, Wasterlain CG, Ribak CE. Dentate granule cells form novel basal dendrites in a rat model of temporal lobe epilepsy. Neuroscience. 1998;86:109–120. doi: 10.1016/s0306-4522(98)00028-1. [DOI] [PubMed] [Google Scholar]
- 56.Stringer JL, Williamson JM, Lothman EW. Induction of paroxysmal discharges in the dentate gyrus: frequency dependence and relationship to afterdischarge production. J Neurophysiol. 1989;62:126–135. doi: 10.1152/jn.1989.62.1.126. [DOI] [PubMed] [Google Scholar]
- 57.Sutula T, Xiao-Xian H, Cavazos J, Scott G. Synaptic reorganization in the hippocampus induced by abnormal functional activity. Science. 1988;239:1147–1150. doi: 10.1126/science.2449733. [DOI] [PubMed] [Google Scholar]
- 58.Sutula T, Cascino G, Cavazos J, Parada I, Ramirez L. Mossy fiber synaptic reorganization in the epileptic human temporal lobe. Ann Neurol. 1989;26:321–330. doi: 10.1002/ana.410260303. [DOI] [PubMed] [Google Scholar]
- 59.Sutula T, Zhang P, Lynch M, Sayin U, Golarai G, Rod R. Synaptic and axonal remodeling of mossy fibers in the hilus and supragranular region of the dentate gyrus in kainate-treated rats. J Comp Neurol. 1998;390:578–594. doi: 10.1002/(sici)1096-9861(19980126)390:4<578::aid-cne9>3.0.co;2-y. [DOI] [PubMed] [Google Scholar]
- 60.Tauck DL, Nadler JV. Evidence of functional mossy fiber sprouting in hippocampal formation of kainic acid-treated rats. J Neurosci. 1985;5:1016–1022. doi: 10.1523/JNEUROSCI.05-04-01016.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Timofeeva OA, Peterson GM. Dissociation of mossy fiber sprouting and electrically-induced seizure sensitivity: rapid kindling versus adaptation. Epilepsy Res. 1999;33:99–115. doi: 10.1016/s0920-1211(98)00078-3. [DOI] [PubMed] [Google Scholar]
- 62.Wenzel HJ, Buckmaster PS, Anderson NL, Wenzel ME, Schwartzkroin PA. Ultrastructural localization of neurotransmitter immunoreactivity in mossy cell axons and their synaptic targets in the rat dentate gyrus. Hippocampus. 1997;7:559–570. doi: 10.1002/(SICI)1098-1063(1997)7:5<559::AID-HIPO11>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 63.Wenzel HJ, Woolley CS, Robbins CA, Schwartzkroin PA. Kainic acid-induced mossy fiber sprouting and synapse formation in the dentate gyrus of rats. Hippocampus. 2000;10:244–260. doi: 10.1002/1098-1063(2000)10:3<244::AID-HIPO5>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 64.Williams PA, Wuarin JP, Dou P, Ferraro DJ, Dudek FE (2002) A reassessment of the effects of cycloheximide on mossy fiber sprouting and epileptogenesis in the pilocorpine model of temporal lobe epilepsy. J Neurophysiol, in press. [DOI] [PubMed]
- 65.Wuarin J-P, Dudek FE. Electrographic seizures and new recurrent excitatory circuits in the dentate gyrus of hippocampal slices from kainate-treated epileptic rats. J Neurosci. 1996;16:4438–4448. doi: 10.1523/JNEUROSCI.16-14-04438.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wuarin J-P, Dudek FE. Excitatory synaptic input to granule cells increases with time after kainate treatment. J Neurophysiol. 2001;85:1067–1077. doi: 10.1152/jn.2001.85.3.1067. [DOI] [PubMed] [Google Scholar]
- 67.Zhang N, Houser CR. Ultrastructural localization of dynorphin in the dentate gyrus in human temporal lobe epilepsy: a study of reorganized mossy fiber synapses. J Comp Neurol. 1999;405:472–490. [PubMed] [Google Scholar]
- 68.Zhu Z-Q, Armstrong DL, Hamilton WJ, Grossman RG. Disproportionate loss of CA4 parvalbumin-immunoreactive interneurons in patients with Ammon's horn sclerosis. J Neuropathol Exp Neurol. 1997;56:988–998. doi: 10.1097/00005072-199709000-00004. [DOI] [PubMed] [Google Scholar]