

Abstract
A 1:1 coordination between locomotor and respiratory movements has been described in various mammalian species during fast locomotion, and several mechanisms underlying such interactions have been proposed. Here we use an isolated brainstem–spinal cord preparation of the neonatal rat to determine the origin of this coupling, which could derive either from a direct interaction between the central locomotor- and respiratory-generating networks themselves or from an indirect influence via a peripheral mechanism. We demonstrate that during fictive locomotion induced by pharmacological activation of the lumbar locomotor generators, a concomitant increase in spontaneous respiratory rate occurs without any evident form of phase coupling. In contrast, respiratory motor activity can be fully entrained (1:1 coupling) over a range of periodic electrical stimulation applied to low-threshold sensory pathways originating from hindlimb muscles. Our results provide strong support for the existence of pathways between lumbar proprioceptive afferents, medullary respiratory networks, and phrenic motoneurons that could provide the basis of the locomotor–respiratory coupling in many animals. Thus a peripheral sensory system involved in a well defined rhythmic motor function can be responsible for the tight functional interaction between two otherwise independent motor behaviors.
Keywords: locomotion, respiration, neuronal network interactions, newborn rat, brainstem–spinal cord, in vitro, entrainment, sensory–motor integrations, proprioceptive feedback
In freely moving invertebrates and vertebrates, different regions of the CNS and peripheral nervous system interact to elicit coordinated physiological responses to environmental and behavioral changes. In this context, rhythm-generating networks involved in different rhythmic motor behaviors may function independently but can express coordinated activity patterns under particular physiological circumstances (Dickinson, 1995). In vertebrates, for example, breathing frequency is well known to increase immediately at the onset of exercise (Krogh and Lindhard, 1913; Dejours, 1959) and a 1:1 coupling occurs during locomotion when fast gaits have been reached, particularly in quadrupeds (Bramble and Carrier, 1983). The neurogenic mechanisms underlying coupling are hypothesized to be an important part of the physiological response required to maintain a sufficient supply of oxygen during exercise, especially in animals in which there is high potential for mechanical interference between locomotor and respiratory movements.
The mechanisms underlying locomotor–respiratory coordination, however, remain controversial, and a number of different hypotheses have been proposed (for review, see Viala, 1997). For example, the mechanical consequences of visceral mass motion, especially during quadrupedal locomotion (Bramble and Carrier, 1983;Young et al., 1992; Bramble and Jenkins, 1993), could lead to a close interaction between the respiratory rate and locomotor stride frequency by providing a tight 1:1 coordination between limb movements and volume changes of the thoracic cavity. However, although this mechanical “visceral piston” hypothesis (Bramble and Carrier, 1983) may apply to quadrupeds with their horizontal body position, it cannot account for harmonic locomotor–respiratory couplings (for example, in human 2:1, 4:1, and also 3:2 or 5:2) observed in running bipeds (Bechbache and Duffin, 1977; Bramble and Carrier, 1983; Perségol et al., 1991; Banzett et al., 1992; Bernasconi and Kohl, 1993) and in birds during free flight (Butler and Woakes, 1980; Funk et al., 1993).
A purely central neural origin has also been proposed for locomotor–respiratory interactions. One possibility is that a common drive originating from the hypothalamus (Eldridge et al., 1981) or medullary structures (Romaniuk et al., 1994) could simultaneously influence locomotor and respiratory rhythm generators. Direct interactions between the central rhythm-generating networks could also be involved because a close coupling between locomotor and breathing patterns persists in a variety of decerebrate and paralyzed vertebrate preparations (Viala et al., 1987; Perségol et al., 1988; Kawahara et al., 1989; Funk et al., 1992b; Corio et al., 1993). Finally, periodic activation of limb (Iscoe and Polosa, 1976; Palisses et al., 1988) or wing (Funk et al., 1992a) sensory inputs has been proposed to play a supportive role in locomotor–respiratory coordination. From this ensemble of data, therefore, it appears that locomotor–respiratory coupling in vertebrates results from a combination of diverse mechanisms, whose relative contribution to the coordination process remains largely unknown.
To further explore the neurogenic origin of the coordination between locomotion and respiration, we have used a completely isolated in vitro preparation of a neonate mammalian nervous system in which various parameters can be easily controlled. Changes in spontaneous fictive respiratory activity were analyzed both with extracellular motor root and whole-cell patch-clamp recordings during pharmacologically induced fictive locomotor activity. Our results show that central lumbar locomotor networks can modulate the frequency of the respiratory generator but are unable to couple it with locomotion. However a strictly phase-locked locomotor–respiratory pattern can be evoked by rhythmic activation of hindlimb (likely proprioceptive) sensory input pathways that could provide the basis of locomotor–respiratory coupling.
MATERIALS AND METHODS
In vitro brainstem–spinal cord preparations.Experiments were performed on isolated in vitro preparations of brainstem–spinal cord from newborn rats (0–4 d of age; Sprague Dawley) from different litters. Timed pregnant rats were obtained from a breeding center (Iffa Credo-Charles Rivers, L'Arbresle, France). Animals were deeply anesthetized with ether and decerebrated just rostrally to the fifth cranial nerves. The skin and muscles were rapidly removed and preparations were then placed in a 25 ml chamber containing artificial CSF (see composition below) maintained at 10°C during the dissection. The flow rate (5–10 ml/min) was set to change the total chamber volume within 5 min. The dissection was continued under binocular microscopy to gently isolate the brainstem and the entire spinal cord with its dorsal and ventral roots still attached. At the cervical level, the ventral roots of one side of the spinal cord were kept intact and uncut. The phrenic nerve was then located and cut at the diaphragm level to allow inspiratory activity to be recorded from its central cut end. Finally, the neuraxis was fixed on a Sylgard resin block (Dow Corning, Midland, MI) with the ventral surface upward. Preparations were superfused continuously with artificial CSF equilibrated with 95% O2/5% CO2, pH 7.4, and containing (in mm): 113 NaCl, 4.5 KCl, 1 NaH2PO4, 2 CaCl2, 1 MgCl2, 25 NaHCO3, and 11d-glucose.
Recordings. After dissection, preparations were placed in a 10 ml recording chamber partitioned into two baths with independent perfusion systems (Fig.1A). Perfusion rates (3–5 ml/min) of rostral and caudal chambers were set to change the total chamber volume within 2 min. Petroleum jelly bridges allowed the cord to remain intact between the different compartments and watertightness was checked at the end of each experiment by adding dye (Fast Green, Sigma, Saint-Quentin Fallavier, France) to the perfusion medium. The brainstem–spinal cord preparation was partitioned at low thoracic (T10–T11) spinal levels. The temperature of the artificial CSF was then progressively raised to 25°C and both spinal ventral root and nerve activities were recorded using glass suction electrodes. Signals were amplified (×10000) by homemade amplifiers, bandpass filtered (0.1–3 kHz), rectified, integrated (τ = 100 msec), displayed on an oscilloscope (Hameg, Frankfurt, Germany), and stored on a computer hard disk (Spike 2; Cambridge Electronics Design, Cambridge, UK) for off-line analysis.
Fig. 1.
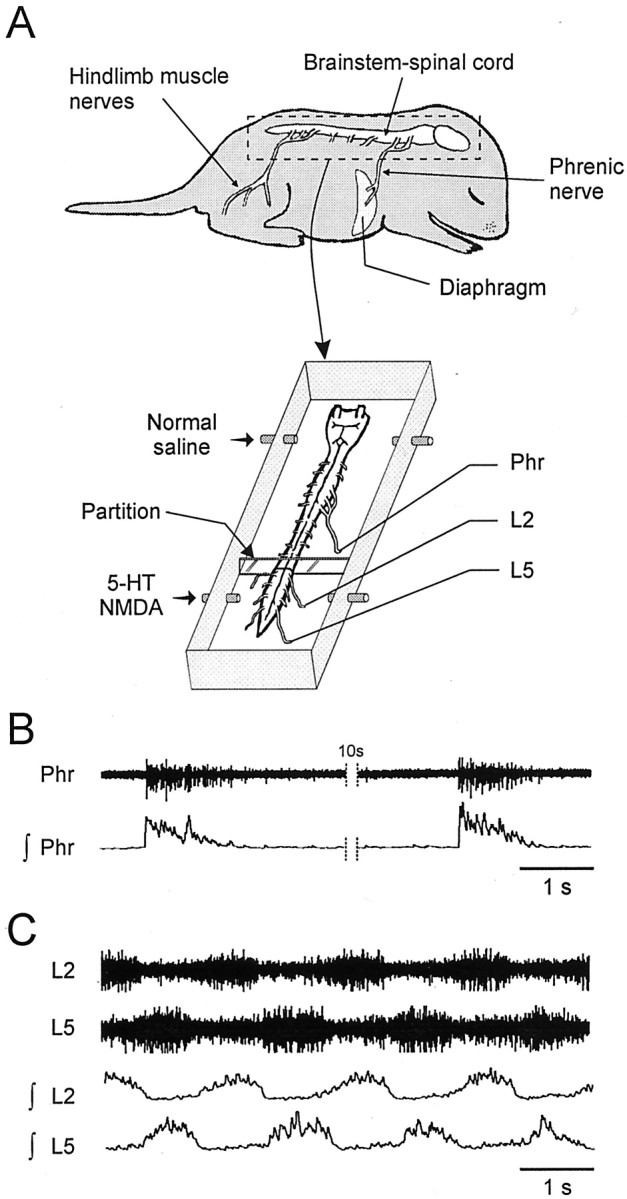
In vitro mammal preparation used to study locomotor–respiratory coupling. A, Schematic drawings of a neonate rat CNS (brainstem–spinal cord) before and after isolation in a recording chamber. Simultaneous recordings were made from a respiratory phrenic nerve (Phr) and locomotor lumbar (L2, L5) ventral roots. Locomotor rhythm-generating networks had to be activated by lumbar cord perfusion of a medium containing 5-HT (10−5m) and NMDA (0.5–2 × 10−5m).B, Raw (top) and integrated (bottom) phrenic nerve activity showing spontaneous respiratory bursts. C, Episode of 5-HT/NMDA-induced locomotor rhythmicity recorded from homolateral lumbar L2 and L5 ventral roots.
Intracellular recordings were made from spinal motoneurons with the blind whole-cell patch-clamp recording technique. Patch electrodes (4.5–6.5 MΩ) were pulled from borosilicate glass (TW150–4; World Precision Instruments, Sarasota, FL) with a vertical puller (PP-83, Narishige, Tokyo, Japan) and filled with a solution containing (in mm): 130 K+-gluconate, 1 CaCl2, 10 HEPES, 2 ATP (Mg2+ salt), and 10 EGTA, pH 7.3, adjusted with KOH. To facilitate tissue penetration by the patch electrode, a thin layer of the ventral part of the white matter was gently scratched. Ventral root activity was checked before and after the scratch procedure to evaluate possible lesioning of the motoneuronal areas. Signals were amplified (Axopatch 1D; Axon Instruments, Foster City, CA) and low-pass filtered (5 kHz; Bessel filter).
Electrical stimulation. Train stimulus pulses over a range of 0.2–5 V with a 0.5 msec duration were applied to spinal roots via glass suction electrodes at 5–20 Hz using an eight channel digital stimulator (A.M.P.I., Jerusalem, Israel).
Drug application and modified saline. Pharmacological substances were bath-applied at least 30 min after the end of dissection by means of gravity supply. The following drugs (all from Sigma) were used: glutamate agonist acting on NMDA receptors (0.5–2 × 10−5m), serotonin (5-HT; 10−5m), and the GABAergic antagonist bicuculline (0.2–2 × 10−5m). In some experiments, a modified saline containing a low Ca2+ concentration (0.1 mm CaCl2, 5 mm MgCl2) was used to reversibly block synaptic transmission.
Histology. Spinal cords were fixed by immersion overnight in 2% paraformaldehyde at 4°C. Selected nervous tissue was cryoprotected at 4°C in 15% (for 24 hr) and then 30% (for 12 hr) sucrose in 0.1 m phosphate buffer. The tissue was quickly frozen, embedded in a plastic resin (Tissue-Tek; Sakura, Tokyo, Japan), and cut transversely with a cryostat at 25 μm thickness. Serial sections were mounted on gelatin-coated glass slides and stained with methylene blue.
Data analysis. Membrane potentials of motoneurons were adjusted for liquid junction potentials (−10 mV for the solution contained in the patch electrode). Statistical values were expressed as mean ± SEM. Differences between means were analyzed using a statistical software package (Sigma Stat) and assessed either by the Student's t test or one-way ANOVA. Changes in mean values for each parameter were taken to be significant at p < 0.05.
RESULTS
Modulation of respiratory rate by fictive locomotion
The isolated in vitro brainstem–spinal cord preparation from neonate rats (Fig. 1A) (for review, see Hilaire and Duron, 1999) is a suitable model for analyzing locomotor–respiratory interactions near birth, because it has the capacity to produce both spontaneous respiratory rhythm originating from the medulla (Fig. 1B) and a pharmacologically induced locomotor rhythm generated in the lumbar spinal cord (Fig.1C). In such a deafferented preparation, therefore, possible interactions between the two rhythm-generating networks can be assessed directly without any involvement of peripheral sensory inputs.
Motor activity recorded from the phrenic motor nerves is considered to be a close reflection of the operation of the medullary network underlying respiratory genesis (Rekling et al., 2000). Under control conditions in our in vitro neonatal preparation, the pattern of this spontaneous activity was stable, with rhythmic bursts occurring at a mean period of 14.6 ± 1.1 sec (ranging from 8 to 21 sec;n = 10 brainstem–spinal cords). In contrast, under the same control conditions, simultaneous recordings from lumbar (L)2 and L5 ventral roots, which carry hindlimb flexor and extensor motor innervation, respectively (Kiehn and Kjaerulff, 1996), invariably lacked spontaneous locomotor rhythmogenic activity (Fig.2A1). However, bath application of a mixture of 5-HT (10−5m) and NMDA (0.5 × 10−5m) consistently elicited a stable rhythm in the L2 and L5 ventral roots (with a mean period of 3.3 ± 0.2 sec;n = 4) (Fig. 2B) (Cazalets et al., 1992; Kiehn and Kjaerulff, 1996). Importantly, this induction of rhythmic activity in the lumbar locomotor networks had no significant effect (p = 0.119) on the ongoing respiratory rhythm period (Fig. 2C). An increase in the NMDA concentration (to 10−5m) in the perfusion cocktail slightly accelerated the locomotor rhythm (to a mean period of 2.8 ± 0.1 sec;n = 11) (Fig. 2B) but did not have any significant effect (p = 0.292) on the respiratory rhythm frequency (Fig.2A2,C). However, perfusion of the lumbar cord with a still higher NMDA concentration (2 × 10−5m) significantly decreased both the mean locomotor (1.7 ± 0.2 sec;n = 6; p < 0.01) (Fig.2A3,B) and respiratory periods (by 31 ± 8.1%; n = 6; p< 0.01) (Fig. 2A3,C). These effects of NMDA and 5-HT, which are reversible (Fig.2A4), therefore suggest that a direct and central influence from the spinal locomotor networks on the upstream respiratory generators occurs only when a “threshold” locomotor frequency has been reached. However, this ascending influence appears to involve a generalized excitability-dependent modulation, because no strict phase coupling between the two rhythms was observed.
Fig. 2.
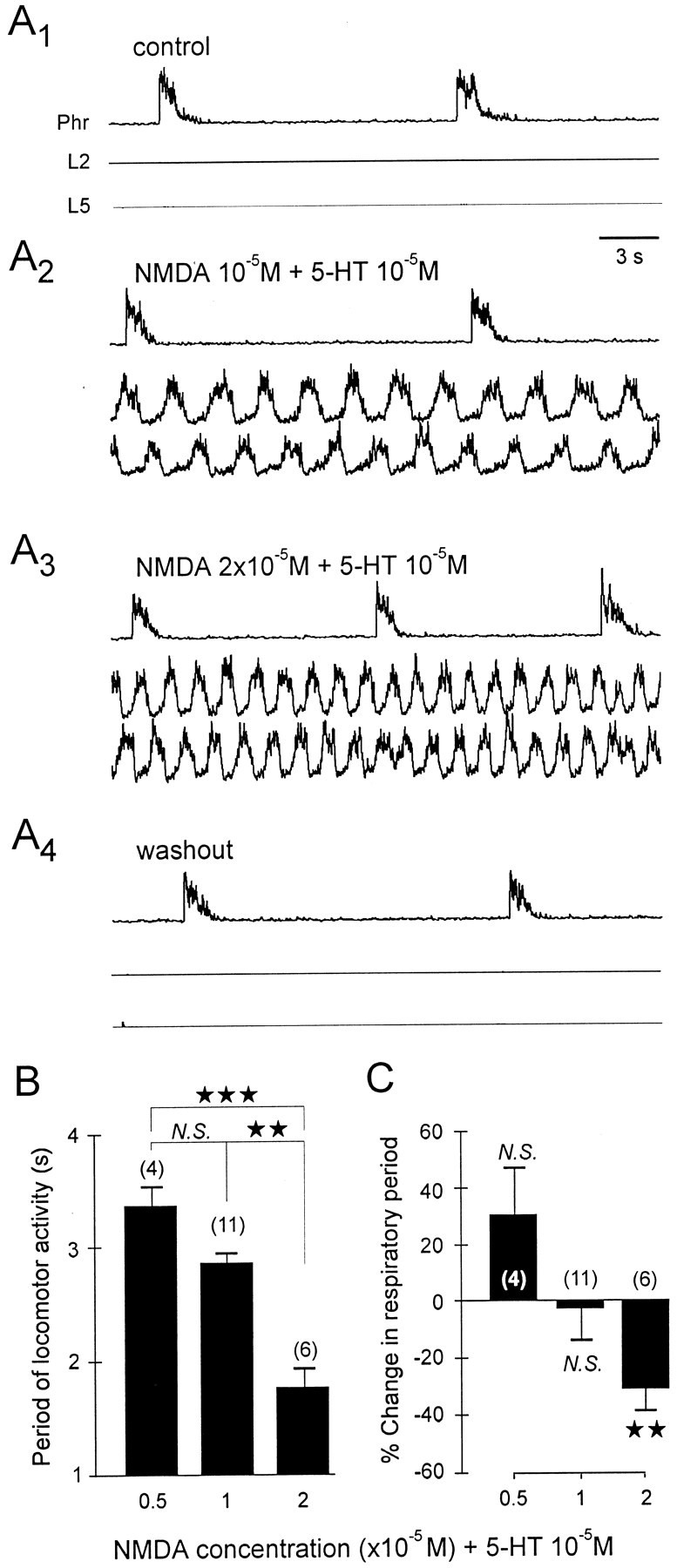
Modulation of respiratory burst frequency by activation of the lumbosacral locomotor generators. A, Phrenic (Phr) and lumbar (L2,L5) integrated activity under control conditions and during application (restricted to the lumbar cord) of constant 5-HT (10−5m) and increasing NMDA concentrations (from 10−5 to 2 × 10−5m). B, Histogram showing the relationship between different NMDA concentrations (0.5–2 × 10−5m plus 10−5m 5-HT) in the lumbosacral bath and the period of the induced locomotor rhythm. C, Histogram showing the resulting change in respiratory rate expressed as percentage of control value in the absence of drugs. Vertical bars indicate mean values; vertical linesindicate the SEM. N.S., Nonsignificant. ★★★p < 0.001; ★★p < 0.01.
Respiratory rhythm resetting by lumbar peripheral afferents
Because close locomotor–respiratory coupling is not mediated by a direct central interaction between the two rhythm-generating networks in this in vitro neonate preparation, we postulated the involvement of a third system in the coupling process. Specifically, we investigated a possible coordinating role played by the lumbar peripheral afferents arising from hindlimb proprioceptors, because this sensory system is functional at birth (Kudo and Yamada, 1987) and is known to be naturally activated during episodes of actual locomotion (Clarac et al., 2000; Duysens et al., 2000; Pearson, 2000).
To activate lumbar sensory inputs in our reduced preparation, electrical stimuli (trains of shocks with a threshold ranging from 0.6 to 1.1 V) were applied via a suction electrode to the distal end of an identified lumbar dorsal root (L1-L5; n = 11 preparations). The evoked incoming volley was recorded more centrally from the same dorsal root as far as possible from the stimulation site to identify which types of afferent were implicated (Fig.3A, inset). Concomitantly, respiratory-like activity was monitored from a phrenic nerve (Fig. 3A–C). Using this paradigm and under control saline perfusion of the whole cord, activation of low-threshold lumbar afferents caused premature triggering of phrenic bursts in the otherwise spontaneous respiratory cycle (Fig. 3C). No significant difference was observed between evoked and spontaneous respiratory bursts (Table 1, left). This respiratory resetting phenomenon was validated first by the lack of significant change in respiratory cycle periods after the occurrence of the triggered phrenic burst (Fig. 3D). Second, as seen in the phase response plot of Figure 3E, the low-threshold afferent activation evoked respiratory rhythm resetting at all phases in the respiratory cycle. Moreover, the phase shift (Δφ) of phrenic respiratory bursting was a strict function of stimulus phase (φ), so that the earlier in a cycle a lumbar afferent stimulus was applied, the greater the phase advance of the subsequent respiratory cycle. This robust effect, which was further confirmed by the value of the coefficient of determination (R2 = 0.98), therefore demonstrated that a strong functional connection exists between lumbar peripheral afferents and the medullary respiratory rhythm generating networks.
Fig. 3.
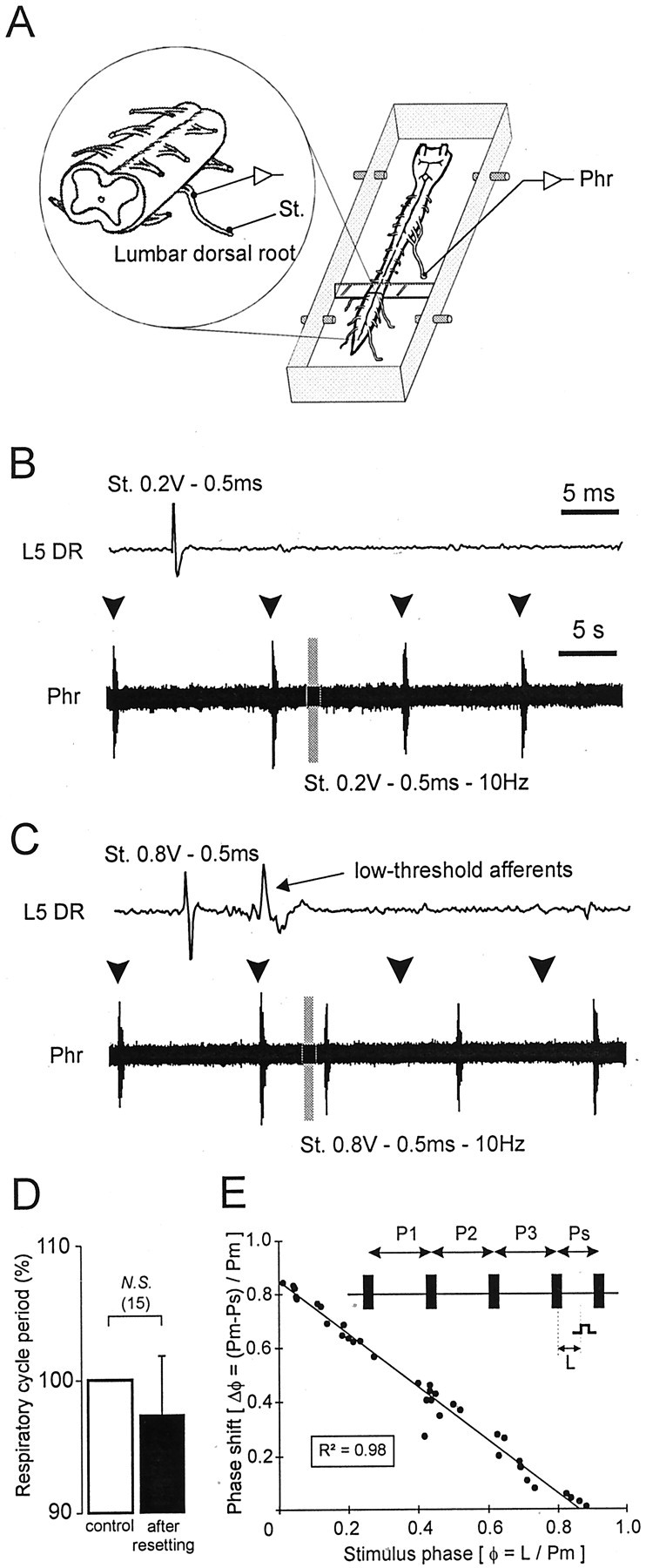
Ability of low-threshold lumbar afferents to reset spontaneous respiratory rhythmicity. A, Schematic representation of the experimental procedure. B, C, Continuous recordings of spontaneous phrenic (Phr) activity during a volley of lumbar (L5) dorsal root (DR) stimulation. Shown above each phrenic trace is a faster time base recording from the corresponding L5 DR during a single shock at the indicated stimulus intensity. The gray barindicates a train stimulation of lumbar afferents. Subthreshold electrical stimulation (0.2 V) of lumbar afferents (B) did not reset the respiratory phrenic rhythmicity, whereas respiratory resetting was obtained when low-threshold lumbar afferents were activated by ≥0.8 V (C). Arrowheads denote the expected time of occurrence of spontaneous phrenic bursts in the absence of resetting. D, Histograms showing lack of significant change in respiratory period (expressed as percentage of the mean control period) after resetting. The control value corresponds to the mean of three successive respiratory periods before the stimulated cycle (white bar), which is compared with the respiratory cycle observed after the stimulated cycle (black bar). N.S., Nonsignificant. E, Phase response plot calculated as follows (also see schematic): the reference period (Pm) was measured from three spontaneous respiratory cycles (P1, P2, and P3); the ratio of the stimulus latency (L) and Pm determined the stimulus phase (φ); the phase shift of the phrenic burst (Δφ) expressed as the difference between Pm and the stimulated period (Ps) and divided again by Pm, was plotted on the ordinate. The solid line indicates linear regression.R2, Coefficient of determination. Standardized data were collected from three different preparations.
Table 1.
Activity in phrenic nerves and phrenic motoneurons during spontaneous (generated endogenously) and evoked (by lumbar dorsal root stimulation) respiratory bursts
| Nerve phrenic bursts | Whole-cell phrenic bursts | ||||
|---|---|---|---|---|---|
| Duration (msec) | Area (a.u.) | Duration (msec) | VSYN (mV) | Firing rate (Hz) | |
| Spontaneous | 558 ± 50 (13) | 143 ± 4 (10) | 870 ± 54 (9) | 19.4 ± 1.6 (9) | 23.3 ± 2.2 (9) |
| Evoked | 593 ± 63 (13) | 130 ± 7 (10) | 855 ± 23 (9) | 23.1 ± 1.1 (9) | 29.6 ± 1.0 (9) |
| pvalue | 0.726 (N.S.) | 0.192 (N.S.) | 0.833 (N.S.) | 0.105 (N.S.) | <0.05* |
Mean ± SEM are presented and compared between spontaneous and evoked groups by Student's t test. a.u., Arbitrary unit; VSYN, respiratory synaptic drive potential; N.S., nonsignificant difference.
*Significant.
In a subsequent step, lesion and pharmacological experiments were performed to determine whether the respiratory resetting action of lumbar afferent stimulation is mediated by a direct neuronal pathway up the spinal cord or is conveyed indirectly via the central lumbar locomotor circuitry itself. A number of experimental arguments favor the first possibility. First, partial transection (dorsal half) of the spinal cord at the C1 level (n = 3) (Fig.4A,B) between the medullary respiratory-generating networks and phrenic motor output prevented dorsal ascending pathways from reaching the medullary respiratory centers. This blocked the ability of the lumbar afferents to reset respiratory rhythm without affecting the ability of the respiratory networks to drive phrenic motoneurons. Second, all synaptic input from lumbar afferents to lower spinal networks was reversibly blocked using a low Ca2+ medium (n = 3) applied selectively to the lumbosacral cord. Under control conditions in these experiments (Fig. 4C,top), stimulation of the low-threshold lumbar afferents induced respiratory resetting and elicited brief locomotor rhythmicity. When the lumbosacral spinal cord was perfused with a low Ca2+ medium (Fig. 4C,middle), respiratory resetting by lumbar afferent stimulation remained, whereas its ability to induce locomotor activity was totally blocked. These results therefore support the conclusion that lumbar sensory inputs have direct access to the higher respiratory centers via a neuronal pathway that bypasses the lumbar locomotor generator networks to ascend in the dorsal spinal cord.
Fig. 4.

Direct influence of low-threshold lumbar afferents on medullary respiratory networks. A, B, Respiratory resetting (A) results from an action on medullary respiratory centers, because transection of the dorsal spinal cord at C1 (B, see histological control; compare withA) suppressed the ability of the same lumbar afferent stimulation to reset phrenic activity (bottom trace). Note that the pattern of phrenic (Phr) motor bursts was similar in the two experimental conditions (see fast time base raw and integrated records at the top right of eachpanel). Arrowheads denote the expected time of occurrence of phrenic bursts in the absence of resetting. The gray bar in A and Bindicate a train stimulation (St., 0.5 msec, 0.8 V, 10 Hz) of lumbar afferents. The dotted line in B shows the part of the spinal cord removed. C, Effects of lumbar afferent activation (gray bar: St., train stimulation, 0.5 msec, 0.7 V, 10 Hz) on both phrenic nerve (Phr) and lumbar ventral root (L5) activity under control conditions (top), during low Ca2+ perfusion of the lumbosacral cord (middle), and after washout with normal saline (bottom). Note that under normal saline perfusion (top and bottom panels), activation of the lumbar afferents also elicited a short sequence of locomotor bursting (arrows).
Priming of phrenic motoneurons for premature activation
To further evaluate the impact of low-threshold lumbar afferents on respiratory output, whole-cell patch-clamp recordings of phrenic motoneurons (n = 9) were performed. As seen above with whole-root recordings, bursting in individual phrenic motoneurons was phase reset by electrical activation of lumbar afferents (Fig.5A). Although the mean firing rate appeared slightly but significantly increased during evoked respiratory bursts, the burst duration and the respiratory synaptic drive potential did not show any significant difference (Table 1, right). During such stimulation, a sequence of different types of postsynaptic potentials was elicited in the recorded motoneuron, consisting of an initial, usually subthreshold EPSP (mean latency, 55.2 ± 1.3 msec; n = 20) (Jahr and Yoshioka, 1986) that was immediately followed by a series of IPSPs and finally by a spike burst driven by a membrane depolarization (Fig. 5B; also see Fig. 7B, numbers 1, 2, and 3, respectively). Injection of constant hyperpolarizing current into these phrenic cells nullified and then reversed the compound IPSP at membrane potentials approximately equal to −70 mV (Fig. 5C,D) in the vicinity of the estimated Cl− equilibrium potential (ECl− = −73.6 mV). Moreover, perfusion of the GABA antagonist bicuculline (0.2–2 × 10−5m; n = 4) on the entire spinal cord reversibly blocked the IPSPs induced by lumbar sensory activation but did not prevent subsequent resetting of the phrenic motor burst (Fig. 5E). We therefore propose that lumbar afferent input has access in parallel to several different levels of the central respiratory circuitry (see Fig. 7B, pathway schematic).
Fig. 5.

Postsynaptic effects of lumbar afferent activation on phrenic motoneurons. A, Simultaneous whole-cell patch-clamp recording of a phrenic motoneuron (Phr Mn) and raw activity of cervical ventral root (C5) under control conditions (top) and during lumbar dorsal root stimulation (bottom, St.). The traces are contiguous (dashed lines). Arrowheadsdenote the expected time of occurrence of phrenic bursts in the absence of resetting. The inset shows motoneuron identification by antidromic electrical stimulation (St.) of corresponding phrenic ventral root. B, Details of postsynaptic events induced in the phrenic motoneuron by lumbar dorsal root stimulation. Vertical bars: St., Train stimulation (0.5 msec, 0.7 V, 10 Hz). Note the initial occurrence of an EPSP followed by series of IPSPs before a spike burst. Thedashed line indicates the resting membrane potential level. C, Hyperpolarizing current-induced reversal of stimulus-evoked compound IPSP. Dashed lines represent the resting membrane potential. D, Scatter plot illustrating the relationship between maximal IPSP amplitude and motoneuron membrane potential. Note the reversal potential at approximately −70 mV (data from 7 phrenic motoneurons). The solid line indicates linear regression. R2, Coefficient of determination. E, Bicuculline application (0.2 × 10−5m) blocks lumbar afferent-evoked inhibition of a phrenic motoneuron. Note that the action potentials inE have been truncated.
Fig. 7.

Lumbar motor-sensory loop as a neural substrate for mammalian locomotor–respiratory coupling. A, Spontaneous respiratory activity of phrenic nerve (Phr) and pharmacologically induced locomotor rhythmicity (L2, lumbar ventral root) in the absence of low-threshold lumbar afferent stimulation. The underlying central networks are schematized on theright. B, bottom left, Repetitive activation of low-threshold lumbar afferents (vertical bars: St.) in time with locomotor bursts immediately drives locomotor–respiratory coupling. The two sets oftraces in A and B are from the same experiment. Related synaptic events recorded from a single phrenic motoneuron are shown in B (upper left). B, right, Schematic representation of proposed circuitry involved in the locomotor–respiratory coupling (hindlimb extensor and flexor muscles, which were removed in our preparations, are included to complete thein vivo motor-sensory loop). Connections responsible for different synaptic influences (arrowheads) on phrenic motoneurons are also numbered. Phr Mn, Phrenic motoneuron; Lumbar Mn, lumbar motoneuron;Resp., respiratory; Loco., locomotor.
Rhythmic lumbar peripheral input can entrain respiratory rhythmicity
Does the resetting action of lumbar afferent stimulation provide a functional substrate for phase-locked locomotor and respiratory coupling? To address this question, we rhythmically activated low-threshold afferents to the lumbar cord with different train stimulation periods (TSPs), to mimic cyclic sensory feedback from the hindlimb during changing locomotor frequencies in the freely moving animal. Because spontaneous respiratory activity in our reduced preparation occurred at minimal periods of ∼8 sec, this value was taken as the maximal TSP applied to the lumbar dorsal roots. With this approach, rhythmic low-threshold lumbar afferent stimulation with a TSP decreasing progressively in 1 sec steps could fully entrain (1:1 coupling) spontaneous respiratory activity at stimulus rate over a range from 8 down to 4 sec (Fig.6A,B). With an additional reduction in TSPs, the 1:1 coupling disappeared (data not shown) and coordination then reappeared as a harmonic (2:1) of the fundamental coupling (Fig. 6A, bottom trace). It is noteworthy that for a given TSP value, the delay to the next phrenic burst remained constant, although it varied for different TSPs (Fig. 6A,C). Together, these data point to a powerful action of lumbar afferents on the medullary respiratory rhythm-generating network, in addition to a direct influence on phrenic motoneurons themselves (see also Perségol et al., 1987).
Fig. 6.

Respiratory rhythm entrainment by rhythmic activation of low-threshold lumbar afferents. A, Recordings of phrenic nerve activity (Phr) during electrical stimulation (St.; 0.9 V, 0.5 msec, 10 Hz) of lumbar afferents with different TSPs. A 1:1 coordination occurs with TSPs of 6 sec and 4 sec but fails at a TSP of 3 sec. B, Scatter plot showing the relationships between respiratory cycle period and lumbar afferent TSPs; the solid line indicates a 1:1 coupling. C, Box plots representing phrenic burst latency (lat.) in relation to the train stimulation period (see schematics above). ★★p< 0.01; ★p < 0.05.
Finally, in an attempt to mimic the situation during real motor behavior, we experimentally “closed” the lumbar motor-sensory loopin vitro by stimulating the low-threshold lumbar afferents in time with fictive locomotor activity (n = 3) (Fig.7B). As reported above, perfusion of the lumbosacral cord with both 5-HT and NMDA elicited locomotor rhythmicity that was completely unrelated to the timing of spontaneous respiratory phrenic activity (Fig. 7A,left traces). However, when the low-threshold lumbar afferents were driven in time with these rhythmic locomotor bursts, tight locomotor–respiratory coupling occurred immediately (Fig.7B, bottom left traces).
DISCUSSION
The results described here from the newborn rat provide new evidence that neural pathways between lumbar proprioceptive inputs and respiratory networks (medullary centers and phrenic motoneurons) exist that could underlie locomotor–respiratory coupling. They also provide, for the first time in a neonate mammal, an insight into the cellular mechanisms and central pathway by which this neurogenic coupling is achieved.
Mechanisms underlying locomotor–respiratory coordination
Locomotor–respiratory coupling in vertebrates is likely to be influenced by a variety of physiological factors acting synergistically. As mentioned previously, various mechanisms (e.g., mechanical, metabolic, and neurogenic interactions) could underlie respiratory entrainment that may vary according to the animal and the locomotor mode used (for review, see Viala, 1997). The use of an isolated in vitro preparation of the vertebrate nervous system restricts analysis to the contribution of purely neurogenic interactions (central and peripheral) and allows a precise control of sensory feedback activation (i.e., dorsal root stimulations). In these reduced preparations, moreover, higher neural centers such as the hypothalamus and the cortex have been removed. Although we do not exclude the possible involvement of these regions in locomotor–respiratory interaction, their ability (through feedforward mechanisms) to provide information in locomotor period that could entrain respiration is doubtful. Clearly they cannot account for the results obtained in the present study.
We report here that pharmacological activation of lumbar locomotor-generating networks leads to an increase in respiratory frequency only when a threshold locomotor rate has been reached (Fig.2). This agrees with previous studies showing that the strength of locomotor–respiratory interaction increases with stepping rate (Kawahara et al., 1989). However in our experiments, this direct network interaction did not manifest any form of phase-coupling, which is also consistent with previous reports (Hill et al., 1988) that rhythmic activity of the lumbar locomotor networks can excite the respiratory centers but is not directly responsible for any cycle-by-cycle coupling.
Proprioceptive hindlimb inputs are known to be rhythmically activated during actual locomotion in a cycle-to-cycle manner, and previous experiments performed on walking cats with intact sensory hindlimb feedback have shown that proprioceptive inputs can crucially influence the timing of stepping movements (for review, see Orlovsky et al., 1999; Duysens et al., 2000). In this way, a number of studies have reported the powerful effect of group I muscle afferents in resetting the locomotor rhythm using in vitro preparations from neonatal rats (Kiehn et al., 1992; Iizuka et al., 1997). Our in vitro results indicate that activation of the lumbar motor-sensory loop also contributes to locomotor–respiratory coupling in quadrupeds, and that the ability of hindlimb somatic afferent inputs (see also below) to reset and entrain the respiratory rhythm at birth is the mechanism underlying this phenomenon (Fig. 7). Although there are no data available regarding locomotor–respiratory coordinations in the freely moving rat, our results indicate that these fictive motor activities in an in vitro neonate preparation can become coupled through stimulation of lumbar proprioceptive afferents. However, we cannot rule out the possibility that in the adult, other mechanisms of coupling (e.g., central relationships) could exist that are not yet functional in the immature neonate animal and that may be more crucial in animals in which these two rhythms show complex coordinations as harmonic couplings. Moreover in quadrupeds, we do not exclude the possibility that the activation of cervical locomotor central pattern generators (Ballion et al., 2001) could be required for the occurrence of a central entrainment.
Although similar data from other vertebrate preparations are thus far unavailable, our findings are consistent with data obtained from a number of invertebrate preparations. For example, in the lobster, rhythmic activation of a proprioceptor enables two otherwise independent rhythms to become coordinated via a rhythm-resetting process (Nagy and Moulins, 1981) and in a related system, a single mechanoreceptor neuron is able to redefine the phase relationships between the activity of two central pattern generators (Combes et al., 1999). Unlike in these systems, however, the capacity for rhythm resetting by afferent stimulation in our experiments was surprisingly strong. Indeed, as illustrated by the phase response curve in Figure3E, electrical activation of the lumbar low-threshold afferents evoked respiratory rhythm resetting at all phases within the respiratory cycle. Moreover, consistent with the ability of an unexpected synaptic input to prematurely reactivate the respiratory rhythm generator in each cycle, a state-dependent (rest, active, or refractory) variation in the latency of the respiratory response to lumbar afferent activation was also evident (Fig. 6C). As reported in disinhibited rat spinal cord, such properties are characteristic of an autoregenerative mechanism (Bracci et al., 1997), and interestingly, the well studied medullary structure, the pre-Bötzinger complex, is known to display such autoregenerative capacities (Smith et al., 1991; Koshiya and Smith, 1999). Because the respiratory burst evoked through electrical stimulation is similar to that generated endogenously (Table 1), we therefore propose that low-threshold lumbar afferents also project to this medullary area thought to be critical for respiratory rhythm generation. In this way, respiratory entrainment, characterized by a 1:1 coupling that does not occur at all imposed rates but escapes to modes of harmonic coupling (2:1), may be achieved through a periodic resetting of the respiratory drive.
Finally, unitary whole-cell recordings revealed that lumbar afferent feedback has synaptic access to several different components of the respiratory system. Phrenic motoneurons receive a characteristic and well defined sequence of synaptic input in response to activation of lumbar sensory afferents. We propose that the barrage of IPSPs mediated by GABAergic synaptic inputs serves to prevent any motoneuronal activity during the activation of lumbar afferents. This suppression of activity may “prepare” the phrenic motor neuron population to receive excitatory command via the medullary respiratory network. Phrenic motoneurons, therefore, would be prepared to respond preferentially to the premature respiratory drive triggered by the lumbar sensory afferent activation (Fig. 7B,right).
Involvement of proprioceptive afferents in the developing locomotor–respiratory system
One question arising from our findings is the identity of the sensory fiber group that is activated by electrical stimulations of lumbar dorsal roots. The very low stimulus intensities used to activate these peripheral sensory inputs point to the involvement of larger-diameter fibers, most likely proprioceptive afferents. Interestingly, previous studies on the development of the stretch reflex pathway in isolated rat lumbar spinal cord have reported very similar stimulation parameters used to activate proprioceptive Ia afferents (Kudo and Yamada, 1987). Even if group II afferents supplying the secondary endings of muscle spindles cannot be totally excluded as potential candidates, it is noteworthy that these fibers are much less abundant than group Ia afferents in the developing rat spinal cord (Snider et al., 1992).
Although there are no data available concerning the patterns of afferent activity during actual locomotion in the neonatal rat, those produced through lumbar dorsal root stimulation in this in vitro study are also effective in inducing episodes of locomotor-like activity (Fig. 4C, arrows). Such a triggering effect on lumbar locomotor-generating networks has been convincingly reported in a number of recent studies on rodents using similar patterns of dorsal root stimulation (Lev-Tov et al., 2000;Whelan et al., 2000; Marchetti et al., 2001). However, the influence of low-threshold sensory inputs on spinal locomotor networks changes during postnatal development (Iizuka et al., 1997), and the patterns of locomotion produced in neonates in vivo and the resultant afferent discharge differ drastically from that generated in adults. On the basis of these data, therefore, we conclude that, at least at birth, neural pathways between lumbar proprioceptive inputs and the respiratory system are present that could provide the basis for the locomotor–respiratory coordination. Consistent with this conclusion, finally, is the abolition of any respiratory resetting after a high cervical transection of the dorsal cord that eliminates proprioceptive effects mediated via dorsal spinal columns. However, we do not exclude the possibility that other afferent systems may have been damaged after this lesion approach.
Surprisingly, our results, performed on isolated preparations from very young neonatal rats (0–4 d of age), have revealed that locomotor–respiratory interactions are established very early during the development. Indeed, although the locomotor-generating networks are functional in fetal rodents and are able to produce alternate locomotor-like bursts as early as embryonic day 18.5 (for review, seeBranchereau et al., 2000; Nishimaru and Kudo, 2000), neonatal rats do not walk spontaneously with an adult gait pattern until postnatal day 10 (Westerga and Gramsbergen, 1990), primarily because of the immaturity of limb postural control (Brocard et al., 1999; Vinay et al., 2000). Therefore, we can hypothesize that the mechanisms underlying locomotor–respiratory coupling could play a role in developing nervous systems because they are present at an early stage of life and are possibly functional during an embryonic period when primary muscle afferents are known to make synaptic contact with spinal neurons (Saito, 1979; Kudo and Yamada, 1985, 1987). Significantly, this timing is well before the stage at which the animal has developed actual locomotion relieved of postural constraints.
Concluding remarks
The use of an isolated mammalian nervous system has provided evidence for a peripheral neural origin of locomotor–respiratory coupling. However, we cannot exclude the possibility that in freely moving animals other factors such as mechanical constraints, central neurogenic control, or metabolic conditions are able in parallel to influence locomotor–respiratory interactions. According to the species, the age, the mode (biped or quadruped), and the strength of locomotion (flight, trot, gallop,… ), one of these factors could become predominant over the others. In any case, locomotor–respiratory interactions are present from insects (Ramirez, 1998) to vertebrates, and our results bring new insights to the functional relationships and integrated physiology of mammalian neural networks involved in different vital functions.
Footnotes
This work was supported by the Université Bordeaux 1, the Centre National de la Recherche Scientifique, and the “Conseil Régional Aquitaine.” We thank Drs. John Simmers, Daniel Cattaert, Jean-René Cazalets, and Gérard Hilaire for their thorough contribution to this manuscript, and Bérangère Ballion for histology. We also acknowledge the skillful assistance of all technical services of our laboratory.
Correspondence should be addressed to Dr. Didier Morin, Laboratoire Neurobiologie des Réseaux, Université Bordeaux 1, Unité Mixte de Recherche Centre National de la Recherche Scientifique 5816, Avenue des Facultés, 33405 Talence Cedex, France. E-mail: d.morin@lnr.u-bordeaux.fr.
REFERENCES
- 1.Ballion B, Morin D, Viala D. Forelimb locomotor generators and quadrupedal locomotion in the neonatal rat. Eur J Neurosci. 2001;14:1727–1738. doi: 10.1046/j.0953-816x.2001.01794.x. [DOI] [PubMed] [Google Scholar]
- 2.Banzett RB, Mead J, Reid MB, Topulos GP. Locomotion in men has no appreciable mechanical effect on breathing. J Appl Physiol. 1992;72:1922–1926. doi: 10.1152/jappl.1992.72.5.1922. [DOI] [PubMed] [Google Scholar]
- 3.Bechbache RR, Duffin J. The entrainment of breathing frequency by exercise rhythm. J Physiol (Lond) 1977;272:553–561. doi: 10.1113/jphysiol.1977.sp012059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bernasconi P, Kohl J. Analysis of co-ordination between breathing and exercise rhythms in man. J Physiol (Lond) 1993;471:693–706. doi: 10.1113/jphysiol.1993.sp019923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bracci E, Beato M, Nistri A. Afferent inputs modulate the activity of a rhythmic burst generator in the rat disinhibited spinal cord in vitro. J Neurophysiol. 1997;77:3157–3167. doi: 10.1152/jn.1997.77.6.3157. [DOI] [PubMed] [Google Scholar]
- 6.Bramble DM, Carrier DR. Running and breathing in mammals. Science. 1983;219:251–256. doi: 10.1126/science.6849136. [DOI] [PubMed] [Google Scholar]
- 7.Bramble DM, Jenkins FA., Jr Mammalian locomotor-respiratory integration: implications for diaphragmatic and pulmonary design. Science. 1993;262:235–240. doi: 10.1126/science.8211141. [DOI] [PubMed] [Google Scholar]
- 8.Branchereau P, Morin D, Bonnot A, Ballion B, Chapron J, Viala D. Development of lumbar rhythmic networks: from embryonic to neonate locomotor-like patterns in the mouse. Brain Res Bull. 2000;53:711–718. doi: 10.1016/s0361-9230(00)00403-2. [DOI] [PubMed] [Google Scholar]
- 9.Brocard F, Vinay L, Clarac F. Development of hindlimb postural control during the first postnatal week in the rat. Brain Res Dev Brain Res. 1999;117:81–89. doi: 10.1016/s0165-3806(99)00101-7. [DOI] [PubMed] [Google Scholar]
- 10.Butler PJ, Woakes AJ. Heart rate, respiratory frequency, and wing beat frequency of free flying barnacle geese, Branta leucopsis. J Exp Biol. 1980;85:213–226. [Google Scholar]
- 11.Cazalets JR, Squalli-Houssaini Y, Clarac F. Activation of the central pattern generators for locomotion by serotonin and excitatory amino acids in neonatal rat. J Physiol (Lond) 1992;455:187–204. doi: 10.1113/jphysiol.1992.sp019296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Clarac F, Cattaert D, Le Ray D. Central control components of a “simple” stretch reflex. Trends Neurosci. 2000;23:199–208. doi: 10.1016/s0166-2236(99)01535-0. [DOI] [PubMed] [Google Scholar]
- 13.Combes D, Meyrand P, Simmers J. Dynamic restructuring of a rhythmic motor program by a single mechanoreceptor neuron in lobster. J Neurosci. 1999;19:3620–3628. doi: 10.1523/JNEUROSCI.19-09-03620.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Corio M, Palisses R, Viala D. Origin of the central entrainment of respiration by locomotion facilitated by MK 801 in the decerebrate rabbit. Exp Brain Res. 1993;95:84–90. doi: 10.1007/BF00229657. [DOI] [PubMed] [Google Scholar]
- 15.Dejours P. La régulation de la ventilation au cours de l'exercice musculaire chez l'homme. J Physiol (Paris) 1959;51:163–261. [PubMed] [Google Scholar]
- 16.Dickinson PS. Interactions among neural networks for behavior. Curr Opin Neurobiol. 1995;5:792–798. doi: 10.1016/0959-4388(95)80108-1. [DOI] [PubMed] [Google Scholar]
- 17.Duysens J, Clarac F, Cruse H. Load-regulating mechanisms in gait and posture: comparative aspects. Physiol Rev. 2000;80:83–133. doi: 10.1152/physrev.2000.80.1.83. [DOI] [PubMed] [Google Scholar]
- 18.Eldridge FL, Millhorn DE, Waldrop TG. Exercise hyperpnea and locomotion: parallel activation from the hypothalamus. Science. 1981;211:844–846. doi: 10.1126/science.7466362. [DOI] [PubMed] [Google Scholar]
- 19.Funk GD, Milsom WK, Steeves JD. Coordination of wingbeat and respiration in the Canada goose. I. Passive wing flapping. J Appl Physiol. 1992a;73:1014–1024. doi: 10.1152/jappl.1992.73.3.1014. [DOI] [PubMed] [Google Scholar]
- 20.Funk GD, Steeves JD, Milsom WK. Coordination of wingbeat and respiration in birds. II. “Fictive” flight. J Appl Physiol. 1992b;73:1025–1033. doi: 10.1152/jappl.1992.73.3.1025. [DOI] [PubMed] [Google Scholar]
- 21.Funk GD, Sholomenko GN, Valenzuela IJ, Steeves JD, Milsom WK. Coordination of wingbeat frequency and respiration in Canada geese during free flight. J Exp Biol. 1993;175:317–323. [Google Scholar]
- 22.Hilaire G, Duron B. Maturation of the mammalian respiratory system. Physiol Rev. 1999;79:325–360. doi: 10.1152/physrev.1999.79.2.325. [DOI] [PubMed] [Google Scholar]
- 23.Hill AR, Adams JM, Parker BE, Rochester DF. Short-term entrainment of ventilation to the walking cycle in humans. J Appl Physiol. 1988;65:570–578. doi: 10.1152/jappl.1988.65.2.570. [DOI] [PubMed] [Google Scholar]
- 24.Iizuka M, Kiehn O, Kudo N. Development in neonatal rats of the sensory resetting of the locomotor rhythm induced by NMDA and 5-HT. Exp Brain Res. 1997;114:193–204. doi: 10.1007/pl00005628. [DOI] [PubMed] [Google Scholar]
- 25.Iscoe S, Polosa C. Synchronization of respiratory frequency by somatic afferent stimulation. J Appl Physiol. 1976;40:138–148. doi: 10.1152/jappl.1976.40.2.138. [DOI] [PubMed] [Google Scholar]
- 26.Jahr CE, Yoshioka K. Ia afferent excitation of motoneurones in the in vitro new-born rat spinal cord is selectively antagonized by kynurenate. J Physiol (Lond) 1986;370:515–530. doi: 10.1113/jphysiol.1986.sp015948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kawahara K, Kumagai S, Nakazono Y, Miyamaoto Y. Coupling between respiratory and stepping rhythms during locomotion in decerebrate cats. J Appl Physiol. 1989;67:110–115. doi: 10.1152/jappl.1989.67.1.110. [DOI] [PubMed] [Google Scholar]
- 28.Kiehn O, Kjaerulff O. Spatiotemporal characteristics of 5HT and dopamine-induced rhythmic hindlimb activity in the in vitro neonatal rat. J Neurophysiol. 1996;75:1472–1482. doi: 10.1152/jn.1996.75.4.1472. [DOI] [PubMed] [Google Scholar]
- 29.Kiehn O, Iizuka M, Kudo N. Resetting from low threshold afferents of N-methyl-d-aspartate-induced locomotor rhythm in the isolated spinal cord-hindlimb preparation from newborn rats. Neurosci Lett. 1992;148:43–46. doi: 10.1016/0304-3940(92)90800-m. [DOI] [PubMed] [Google Scholar]
- 30.Koshiya N, Smith JC. Neuronal pacemaker for breathing visualized in vitro. Nature. 1999;400:360–363. doi: 10.1038/22540. [DOI] [PubMed] [Google Scholar]
- 31.Krogh A, Lindhard J. The regulation of respiration and circulation during the initial stages of muscular work. J Physiol (Lond) 1913;47:112–136. doi: 10.1113/jphysiol.1913.sp001616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kudo N, Yamada T. Development of the monosynaptic stretch reflex in the rat: an in vitro study. J Physiol (Lond) 1985;369:127–144. doi: 10.1113/jphysiol.1985.sp015892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kudo N, Yamada T. Morphological and physiological studies of development of the monosynaptic reflex pathway in the rat lumbar spinal cord. J Physiol (Lond) 1987;389:441–459. doi: 10.1113/jphysiol.1987.sp016665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lev-Tov A, Devolvé I, Kremer E. Sacrocaudal afferents induce rhythmic efferent bursting in isolated spinal cords of neonatal rats. J Neurophysiol. 2000;83:888–894. doi: 10.1152/jn.2000.83.2.888. [DOI] [PubMed] [Google Scholar]
- 35.Marchetti C, Beato M, Nistri A. Alternating rhythmic activity induced by dorsal root stimulation in the neonatal rat spinal cord in vitro. J Physiol (Lond) 2001;530:105–112. doi: 10.1111/j.1469-7793.2001.0105m.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nagy F, Moulins M. Proprioceptive control of the bilaterally organized rhythmic activity of the oesophageal neuronal network in the cape lobster, Jasus lalandii. J Exp Biol. 1981;90:231–251. [Google Scholar]
- 37.Nishimaru H, Kudo N. Formation of the central pattern generator for locomotion in the rat and mouse. Brain Res Bull. 2000;53:661–669. doi: 10.1016/s0361-9230(00)00399-3. [DOI] [PubMed] [Google Scholar]
- 38.Orlovsky GN, Deliagina TG, Grillner S. Neuronal control of locomotion. Oxford UP; New York: 1999. [Google Scholar]
- 39.Palisses R, Perségol L, Viala D, Viala G. Reflex modulation of phrenic activity through hindlimb passive motion in decorticate and spinal rabbit preparation. Neuroscience. 1988;24:719–728. doi: 10.1016/0306-4522(88)90364-8. [DOI] [PubMed] [Google Scholar]
- 40.Pearson K. Motor systems. Curr Opin Neurobiol. 2000;10:649–654. doi: 10.1016/s0959-4388(00)00130-6. [DOI] [PubMed] [Google Scholar]
- 41.Perségol L, Palisses R, Viala D. Different mechanisms involved in supraspinal and spinal reflex regulation of phrenic activity through chest movements. Neuroscience. 1987;23:631–640. doi: 10.1016/0306-4522(87)90081-9. [DOI] [PubMed] [Google Scholar]
- 42.Perségol L, Jordan M, Viala D, Fernandez C. Evidence for central entrainment of the medullary respiratory pattern by the locomotor pattern in the rabbit. Exp Brain Res. 1988;71:153–162. doi: 10.1007/BF00247530. [DOI] [PubMed] [Google Scholar]
- 43.Perségol L, Jordan M, Viala D. Evidence for the entrainment of breathing by locomotor pattern in human. J Physiol (Paris) 1991;85:38–43. [PubMed] [Google Scholar]
- 44.Ramirez J-M. Reconfiguration of the respiratory network at the onset of locust flight. J Neurophysiol. 1998;80:3137–3147. doi: 10.1152/jn.1998.80.6.3137. [DOI] [PubMed] [Google Scholar]
- 45.Rekling JC, Funk GD, Bayliss DA, Dong X-W, Feldman JL. Synaptic control of motoneuronal excitability. Physiol Rev. 2000;80:767–852. doi: 10.1152/physrev.2000.80.2.767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Romaniuk JR, Kasicki S, Kazennikov OV, Selionov VA. Respiratory responses to stimulation of spinal or medullary locomotor structures in decerebrate cats. Acta Neurobiol Exp. 1994;54:11–17. [PubMed] [Google Scholar]
- 47.Saito K. Development of spinal reflexes in the rat fetus studied in vitro. J Physiol (Lond) 1979;294:581–594. doi: 10.1113/jphysiol.1979.sp012947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Smith JC, Ellenberger HH, Ballanyi K, Richter DW, Feldman JL. Pre-Botzinger complex: a brainstem region that may generate respiratory rhythm in mammals. Science. 1991;254:726–729. doi: 10.1126/science.1683005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Snider WD, Zang L, Yusoof S, Gorukanti N, Tsering C. Interactions between dorsal root axons and their target motor neurons in developing mammalian spinal cord. J Neurosci. 1992;12:3494–3508. doi: 10.1523/JNEUROSCI.12-09-03494.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Viala D. Coordination of locomotion and respiration. In: Miller AD, Bianchi AL, Bishop BP, editors. Neural control of the respiratory muscles. CRC; New York: 1997. pp. 285–296. [Google Scholar]
- 51.Viala D, Perségol L, Palisses R. Relationship between phrenic and hindlimb extensor activities during fictive locomotion. Neurosci Lett. 1987;74:49–52. doi: 10.1016/0304-3940(87)90049-8. [DOI] [PubMed] [Google Scholar]
- 52.Vinay L, Brocard F, Pflieger J-F, Simeoni-Alias J, Clarac F. Perinatal development of lumbar motoneurons and their inputs in the rat. Brain Res Bull. 2000;53:635–647. doi: 10.1016/s0361-9230(00)00397-x. [DOI] [PubMed] [Google Scholar]
- 53.Westerga J, Gramsbergen A. The development of locomotion in the rat. Brain Res Dev Brain Res. 1990;57:163–174. doi: 10.1016/0165-3806(90)90042-w. [DOI] [PubMed] [Google Scholar]
- 54.Whelan P, Bonnot A, O'Donovan MJ. Properties of rhythmic activity generated by the isolated spinal cord of the neonatal mouse. J Neurophysiol. 2000;84:2821–2833. doi: 10.1152/jn.2000.84.6.2821. [DOI] [PubMed] [Google Scholar]
- 55.Young IS, Warren RD, Altringham JD. Some properties of the mammalian locomotory and respiratory systems in relation to body mass. J Exp Biol. 1992;164:283–294. doi: 10.1242/jeb.164.1.283. [DOI] [PubMed] [Google Scholar]