

Abstract
In the ears of mammals, hair cell loss results in permanent hearing and balance deficits, whereas in fish, amphibians, and birds, the production of replacement hair cells can restore those modalities. In avian ears, continuous exposures to forskolin trigger cell proliferation and the regeneration of hair cells, so we investigated the effect of forskolin on sensory epithelia cultured from the ears of mammals. Continuous 72 hr exposures to forskolin failed to induce proliferation in neonatal rat utricles, but brief (≤1 hr) exposures to forskolin or Br-cAMP did. Proliferation occurred only in media that contained serum. Forskolin also augmented the mitogenic effects of glial growth factor 2. The S-phase entry induced by forskolin was blocked by monensin and bafilomycin, two compounds that can inhibit the recycling of membrane receptors. The results are consistent with the hypothesis that in mammalian vestibular epithelia elevated cAMP induces S-phase entry by increasing the number of growth factor receptors at the plasma membrane.
Keywords: regeneration, hair cells, cell proliferation, hearing, cAMP, receptor recycling
Fish, amphibians, and birds quickly recover from hearing and balance deficits that would be permanent for a mammal. In those species, supporting cells in the sensory epithelia divide to produce progeny that can differentiate as replacement hair cells (for review, see Corwin and Oberholtzer, 1997). Although mammals do not have a robust capacity for spontaneous hair cell regeneration, their balance epithelia are capable of some self-repair (Forge et al., 1993). After hair cell loss, limited numbers of supporting cells divide in those epithelia, and some of the progeny appear to differentiate as hair cells (Warchol et al., 1993; Lambert et al., 1997; Kuntz and Oesterle, 1998). The capacity for supporting-cell division in mammals can be enhanced by treatments with mitogenic growth factors, which are most effective in epithelia from neonates (Lambert, 1994;Yamashita and Oesterle, 1995; Zheng et al., 1997; Kuntz and Oesterle, 1998) (R. Gu, M. Montcouquiol, M. Marchionni, and J. T. Corwin, unpublished observations).
The cell proliferation that underlies regeneration in the hearing organs of birds is believed to be mediated via the second messenger cAMP (Navaratnam et al., 1996). In vitro treatments with high levels of forskolin result in increased incorporation of tritiated thymidine in the supporting cells and hair cells when auditory epithelia from chickens are exposed to that activator of adenylyl cyclase, which catalyzes the production of cAMP (for review, see Shaywitz and Greenberg, 1999). Incubations of the epithelia with inhibitors of the cAMP-dependent protein kinase (PKA) block the effect, suggesting that activation of PKA is required for the forskolin-induced regeneration of hair cells in birds.
To assess the potential for forskolin-mediated proliferation in mammalian ears, we cultured utricular hair cell epithelia from neonatal rats in the presence of forskolin or a cAMP analog. Continuous exposure to forskolin alone did not induce proliferation in rat epithelia, but brief exposures to forskolin or the cAMP analog strongly stimulated S-phase entry. When combined with a continuous exposure to recombinant human glial growth factor 2 (rhGGF2), those treatments significantly increased the mitogenic effect of forskolin. The responses were blocked by monensin and bafilomycin, two compounds that can block receptor recycling to the plasma membrane. The results suggest that in the ears of mammals the mitogenic effects of cAMP may depend on its capacity to increase receptor recycling, similar to its survival-promoting effects on spinal motor neurons and retinal ganglion cells (Meyer-Franke et al., 1998).
MATERIALS AND METHODS
Epithelial cell cultures. Experiments were conducted in accordance with an approved animal use protocol that adhered to practices in the National Institutes of Health Guide for the Care and Use of Laboratory Animals under the supervision of the University of Virginia Animal Care Advisory Committee. The study used 92 2-d-old (P2) Sprague Dawley rats that were anesthetized with carbon dioxide and decapitated. Heads were skinned and placed in ice-cold 70% ethanol for 10 min. The vestibular organs were dissected out from both ears and transferred to ice-cold DMEM/F12 medium (Life Technologies, Gaithersburg, MD), and the otoconia and otolithic membrane were removed. To separate the sensory epithelium from the underlying tissue, utricles were incubated in thermolysin at 0.5 mg/ml (Sigma, St Louis, MO) in DMEM/F12 for 45 min at 37°C in a 5% CO2atmosphere (Saffer et al., 1996). Then they were transferred to ice-cold DMEM/F12 containing 5% fetal bovine serum (FBS; HyClone, Logan, UT) to stop the enzymatic digestion. The epithelium was removed with fine forceps, and the outer edges that formed a boundary with the surrounding nonsensory epithelium were trimmed away with a diamond microscalpel and discarded. The remaining piece of pure sensory epithelium was composed of only supporting cells and hair cells and was cut into four approximately equal pieces. The pieces of epithelium were transferred to a glass-bottom culture dish (MatTek Corporation, Ashland, MA) that had been precoated with poly-l-lysine (5 μg/ml for 1 hr at 37°C; Sigma) and fibronectin (100 μg/ml overnight at 37°C; Sigma). The epithelia were allowed to adhere for 1 hr at 37°C in a 5% CO2 atmosphere in medium that contained 5% FBS. Then they were cultured for 72 hr in the standard medium, DMEM/F12 containing 5-bromo-2-deoxyuridine (BrdU; 3 μg/ml; Sigma), 2.5% FBS, and one of the test compounds or 1% dimethylsulfoxide (DMSO) as a vehicle control.
Test compounds and media. To activate cAMP-dependent cascades, pieces of epithelium were incubated with forskolin (Sigma) or cAMP, 8-bromo-sodium salt (Br-cAMP; Calbiochem, La Jolla, CA). In the “continuous exposure” experiments the test compounds were present throughout the 72 hr culture period. In the “brief exposure” experiments cultures were treated with forskolin (15 min) or Br-cAMP (1 hr) in the standard medium with BrdU, and then the media were replaced by either standard medium alone or medium containing rhGGF2 at 50 ng/ml (Cambridge NeuroScience, Inc., Cambridge, MA) for 72 hr.
To assess the involvement of PKA, pieces of epithelia were incubated with the PKA inhibitor KT5720 (Calbiochem) for 1 hr before a 15 min treatment with forskolin in the absence of BrdU. The medium was then replaced by the standard medium that also contained KT5720, and the culture was maintained for 72 hr.
To inhibit receptor recycling to the membrane, cultures were pretreated with 0.5 μm monensin or 0.1 μm bafilomycin A1 (Calbiochem) for 30 min before the addition of forskolin, followed by a combined treatment with forskolin and monensin for 15 min. Then, the medium was replaced for 30 min by standard medium containing monensin or bafilomycin A1 before it was replaced by the standard medium without monensin or bafilomycin A1 for the remaining 71.5 hr. In one experiment rhGGF2 was used in combination with a 75 min preincubation with 0.5 μm monensin (30 + 15 + 30 min). In that experiment, cultures were treated for 30 min with 0.5 μm monensin in the standard medium but without BrdU. The medium was then replaced by the standard medium containing monensin for 15 min, that was then replaced by the standard medium that also contained 50 ng/ml rhGGF2, and the pieces of epithelium were cultured for 71.25 hr more. In some figures (see Figs. 1, 4, 6, 7), the control bar represents the same data (from 24 pieces of epithelium). The same is true for the GGF2 bar (see Figs. 2, 3, 5; from 37 pieces of epithelium) and for the 15 min forskolin bar (see Figs. 4, 6, 7, 9; from 20 pieces of epithelium).
Fig. 1.
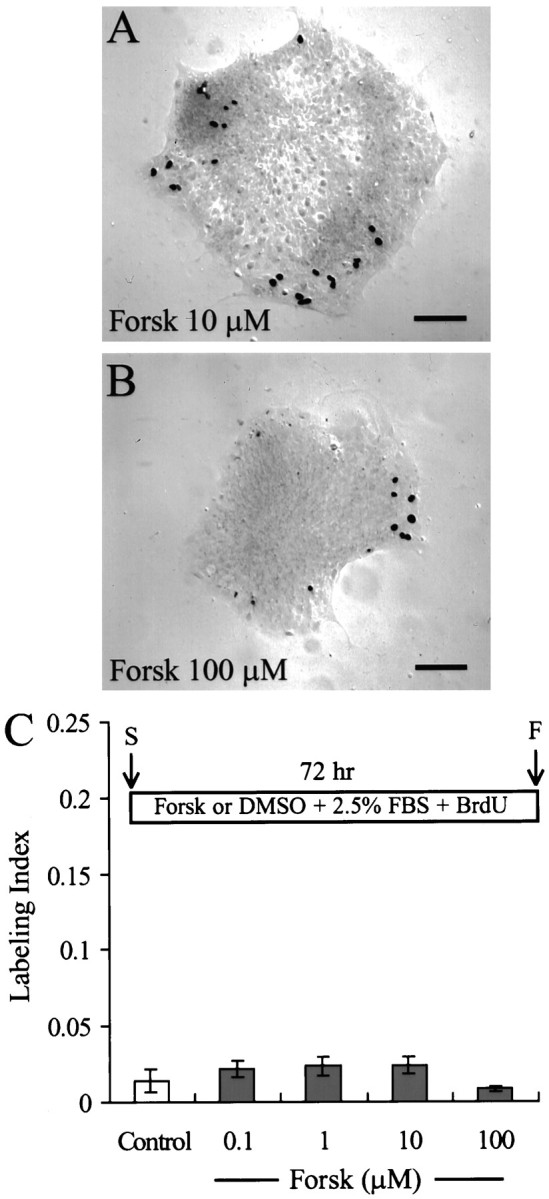
Continuous exposure to 0.1–100 μmforskolin does not stimulate S-phase entry in mammalian utricle cells.A, B, Pieces of utricular sensory epithelium fixed after 72 hr in culture in the standard medium in the continuous presence of 10 μm (A) or 100 μm(B) forskolin (Forsk). Nuclei of cells that entered S-phase and incorporated BrdU were stainedblack after immunocytochemistry and were visualized by differential interference contrast (DIC) microscopy. C, Histogram of the level of BrdU labeling in epithelia showing no significant increase in S-phase entry after a 72 hr treatment with forskolin when compared with control cultures in 1% DMSO.Inset, Timeline. The standard medium consisted of 2.5% FBS and 3 μg/ml BrdU with either 1% DMSO (Control) or forskolin at 0.1–100 μm present from the start of the experiment (S). After 72 hr in culture the tissues were fixed (F) and processed for immunocytochemistry. Scale bars, 100 μm.
Fig. 4.

Brief treatments with forskolin or Br-cAMP induce 11- to 12-fold increases in S-phase entry. A, Timeline is shown. Pieces of epithelium were treated for 15 min with 1 μm forskolin (Forsk) or for 1 hr with 0.5 mm Br-cAMP in a standard medium. The forskolin- and Br-cAMP-containing media were then replaced with a standard medium for the remainder of the 72 hr culture period. In this and all following figures, discontinuity indicators show that the timelines are schematic and not to scale. B, C, When pieces of utricles were treated briefly with forskolin (Forsk 15 min), the level of S-phase entry was ∼12-fold higher than that in control epithelia. When other pieces of epithelium were treated for 1 hr with Br-cAMP (Br-cAMP 1 hr), the level of S-phase entry was ∼11-fold higher than that in the control epithelia. *, Significant difference when compared with Control(p < 0.05). Scale bars, 100 μm.F, Fixed; S, scale.
Fig. 6.
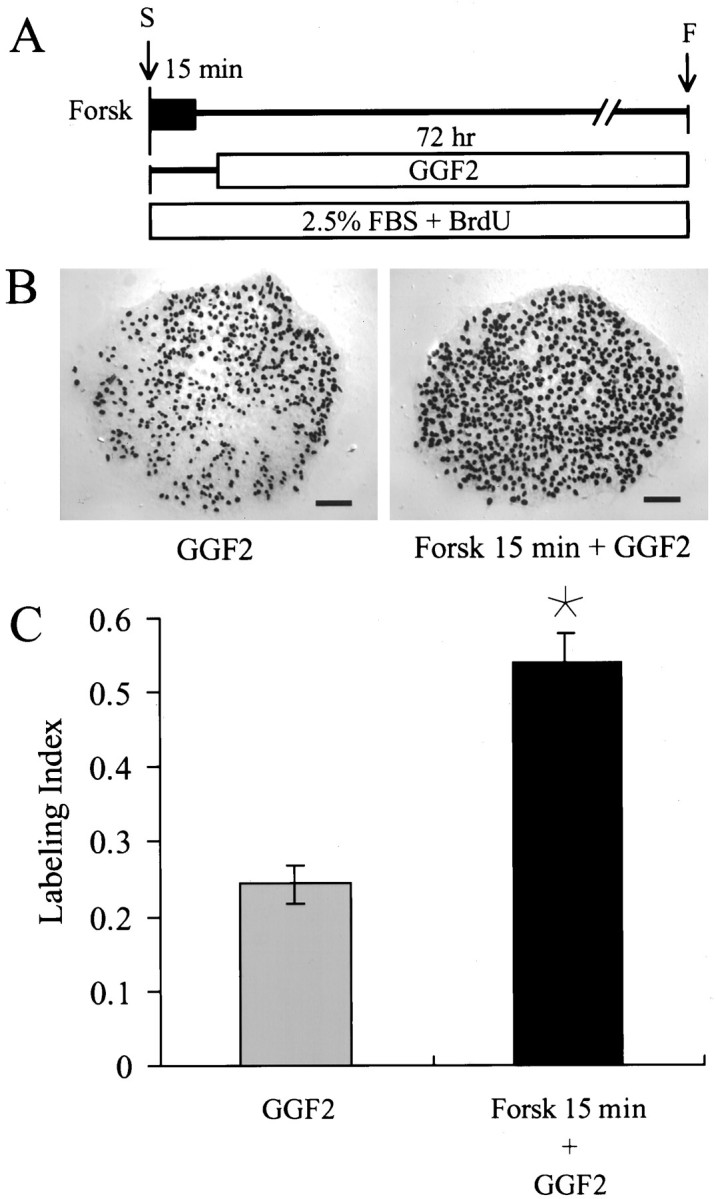
A 15 min treatment with 1 μmforskolin more than doubles the magnitude of S-phase response induced by rhGGF2. A, Timeline is shown. Pieces of epithelium were incubated for 15 min with 1 μm forskolin (Forsk) in a standard medium. The medium was then replaced by a medium containing rhGGF2 with BrdU and FBS for the remainder of the 72 hr culture period. B, DIC micrographs show many black BrdU-labeled nuclei in a piece of utricular epithelium treated with rhGGF2 (GGF2) and the strong increase induced by the brief treatment with forskolin (Forsk 15 min + GGF2). C, The brief treatment with forskolin increased the rhGGF2-induced entry in S-phase up to 52%. *, Significant difference when compared withGGF2 (p < 0.001). Scale bars, 100 μm. F, Fixed; S, start.
Fig. 7.
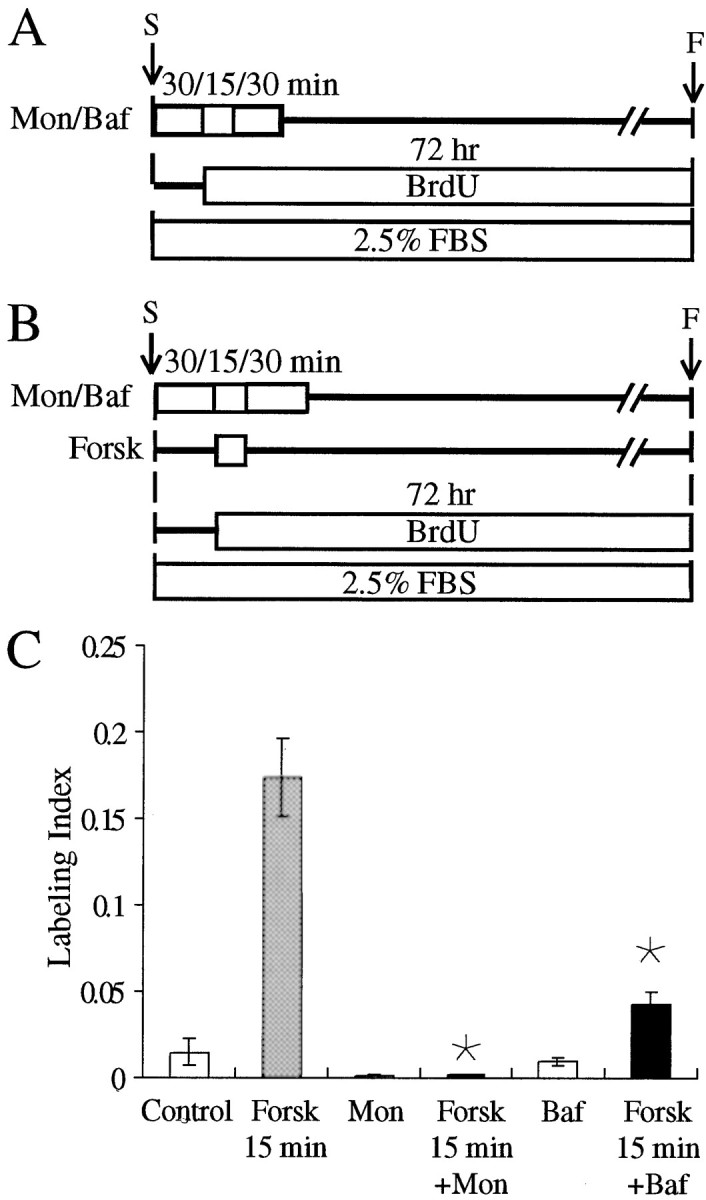
Receptor recycling prevents S-phase induction triggered by brief treatment with forskolin. A, B, Timelines are shown. A, Pieces of epithelium were preincubated for 75 min (30/15/30 min) with 0.5 μm monensin (Mon) or 0.1 μmbafilomycin A1 (Baf). During the first 30 min, the medium did not contain BrdU. For the next 45 min, the medium contained FBS and BrdU. The pieces were then maintained in an FBS-containing medium with BrdU for the remainder of the 72 hr culture.B, Other pieces were pretreated with 0.5 μm monensin (Mon) or 0.1 μmbafilomycin A1 (Baf) for 30 min in a medium devoid of BrdU. The medium was then changed for a standard medium containing 1 μm forskolin (Forsk) for 15 min and replaced by a medium containing 0.5 μm monensin or 0.1 μm bafilomycin A1 and FBS and BrdU for 30 additional minutes. Finally, the monensin or the bafilomycin A1 was removed, and the pieces of utricle were cultured in a standard medium for the remainder of the 72 hr culture period. C, The 75 min (30/15/30 min) treatment with 0.5 μmmonensin or 0.1 μm bafilomycin A1 reduced the level of S-phase entry as compared with the control, but this was not significant. Treatment with monensin inhibited the 15 min forskolin effect up to 99% (Forsk 15 min + Mon), whereas treatment with bafilomycin A1 induced a 70.5% inhibition (Forsk 15 min + Baf). * Significant difference when compared with Forsk 15 min (p < 0.001). F, Fixed; S, start.
Fig. 2.

Continuous exposures to forskolin at 0.1 and 1 μm enhanced rhGGF2-mediated induction of S-phase entry, whereas exposures to 10 and 100 μm forskolin reduced the induction of S-phase that was evoked by rhGGF2. A, Timeline is shown. Pieces of epithelium were cultured for 72 hr in a standard medium containing either 50 ng/ml rhGGF2 (GGF2) or forskolin (Forsk) at concentrations ranging from 0.1 to 100 μm. B, Forskolin (1 μm) potentiated the rhGGF2 effect, whereas at higher concentrations (10 and 100 μm), forskolin significantly inhibited the rhGGF2-induced S-phase entry. *, Significant difference from cultures treated with GGF2(p < 0.05). F, Fixed;S, start.
Fig. 3.

Continuous exposure to Br-cAMP significantly inhibits rhGGF2-induced S-phase entry. A, Timeline is shown. Pieces of epithelium were cultured for 72 hr in a standard medium containing either rhGGF2 alone (GGF2) or Br-cAMP from 0.1 to 1 mm in the presence of rhGGF2.B, At 0.1, 0.5, or 1 mm, the Br-cAMP significantly inhibited the rhGGF2-induced S-phase entry. *, Significant difference when compared with GGF2(p < 0.001). F, Fixed;S, start.
Fig. 5.

Treatment with a PKA inhibitor suggests dependence on PKA for the increase in S-phase induced by brief treatment with forskolin. A, B, Timelines are shown. A, Pieces of epithelium were incubated for 15 min with 1 μmforskolin (Forsk) and then maintained in a standard medium for the remainder of the 72 hr culture period. Other pieces of utricle were preincubated for 60 min with 0.5, 1, or 5 μmKT5720 in a medium containing 2.5% FBS but no BrdU. The medium was then changed for a standard medium containing the inhibitor and 1 μm forskolin for 15 min. The forskolin was removed, and the pieces of epithelium were maintained in a standard medium containing KT5720 for 72 hr. B, Pieces of epithelium were treated for 1 hr with 0.5 mm Br-cAMP in a standard medium. The Br-cAMP-containing medium was then replaced with a standard medium for the remainder of the 72 hr culture period. Other pieces of epithelium were preincubated for 60 min with 5 μm KT5720 in a medium containing 2.5% FBS but no BrdU. The medium was then changed for a standard medium containing the inhibitor and 0.5 mm Br-cAMP for 1 hr. The Br-cAMP was removed, and the pieces of epithelium were maintained in a standard medium containing 5 μm KT5720 for 72 hr. C, Exposure to KT5720 induced a dose-dependent inhibition, reducing the forskolin-induced effect by 52% at 1 μm KT5720 and by 92.4% at 5 μm KT5720. Treatment with 5 μm KT5720 reduced the short-term Br-cAMP effect by 85.2% *, Significant difference when compared with Forsk 15 min(p < 0.05). **, Significant difference when compared with Br-cAMP 1 hr (p< 0.001). F, Fixed; S, start.
Fig. 9.

Absence of serum prevents S-phase induction triggered by brief treatment with forskolin. A, B, Timelines are shown. Pieces of epithelium were treated for 15 min with 1 μm forskolin (Forsk) in a serum-containing medium (A) or in a defined medium (B) composed of DMEM/F12 supplemented with transferrin, sodium selenite, hydrocortisone, and insulin in the presence of BrdU. The medium was then changed for fresh medium containing BrdU for the reminder of the 72 hr culture period.C, The level of forskolin-induced S-phase entry was strongly reduced in the defined medium (Forsk 15 min defined) when compared with the standard medium (Forsk 15 min 2.5% FBS). *, Significant difference when compared withForsk 15 min 2.5% FBS (p < 0.05). F, Fixed; S, start.
Live/dead assays (Molecular Probes, Eugene, OR) were performed according to the manufacturer's recommendations on 24 pieces of utricle cultured in standard medium in the presence of 100 μm forskolin for 24, 48, and 72 hr (eight pieces each).
Bromodeoxyuridine labeling. After culture, epithelia were fixed in 4% paraformaldehyde for 30 min, rinsed three times in PBS, and then immersed in 1N HCl for 15 min to denature the nucleic acids. Immunocytochemical identification of nuclei that had incorporated BrdU was performed at room temperature. The cultures were preincubated for 1 hr in PBS with 10% normal horse serum (NHS) and then incubated for 2 hr in a mouse monoclonal antibody against BrdU (Becton Dickinson, San Jose, CA) diluted 1:50 in PBS with 2% NHS and 0.2% Triton X-100. After three rinses, the specimens were incubated for 30 min in a secondary antibody solution containing biotinylated rat-adsorbed anti-mouse IgG (Vector Laboratories, Burlingame, CA) diluted 1:100 in PBS with 2% NHS and 0.2% Triton X-100. Then they were reacted with diaminobenzidine using an Elite ABC kit (Vector Laboratories) with nickel intensification. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; Molecular Probes) at 10 μg/ml in PBS with 0.1% Triton X-100 for 30 min. After three rinses with PBS, the epithelia were examined by epifluorescence microscopy.
MetaMorph software (Universal Imaging Corporation, Media, PA) was used to acquire images from a cooled CCD camera interfaced to a ZeissAxiovert 135. All the nuclei that were stained by DAPI and all the nuclei that were labeled by BrdU in each sheet of epithelium were counted. The labeling index was calculated for each piece of sensory epithelium by dividing the number of BrdU-labeled cells by the total number of cells. For each test condition, 20–37 pieces of utricular sensory epithelium were analyzed, and the average labeling index and the SEM were calculated. Statistical significance was determined using the two-tailed Student's t test, and n values indicate the number of pieces of epithelium analyzed per condition.
For calretinin immunocytochemistry, cultures immunostained previously for BrdU were incubated for 30 min in PBS with 10% normal goat serum (NGS) and then incubated for 1 hr in a rabbit polyclonal anti-calretinin (Chemicon, Temecula, CA) diluted 1:500 in PBS with 2% NGS and 0.2% Triton X-100. After three rinses, the specimens were incubated for 30 min with an anti-rabbit indocarbocyanine-conjugated secondary antibody (Vector Laboratories) diluted 1:500 in PBS with 2% NGS and 0.2% Triton X-100. After three rinses with PBS, the epithelia were examined by epifluorescence microscopy.
RESULTS
Continuous incubation with forskolin
Continuous 72 hr incubations with forskolin at concentrations of 100 nm, 1 μm, 10 μm, and 100 μm (n = 20, 20, 21, and 22, respectively) did not induce significant increases in S-phase entry in utricular epithelia from P2 rats (Fig. 1). Live/dead assays performed on eight pieces of epithelium cultured for 24 hr, eight pieces for 48 hr, and eight pieces for 72 hr in the presence of 100 μm forskolin revealed less than two dead cells per piece (data not shown). However, when treatments with low concentrations of forskolin were combined with treatments with the mitogenic growth factor rhGGF2, significant numbers of cells entered S-phase (Fig. 2). Approximately 36.8 ± 0.4% of the cells were labeled with BrdU during 72 hr in the presence of both 1 μm forskolin and rhGGF2 (n = 24 pieces of epithelium), a level of labeling that was significantly higher than that induced by rhGGF2 alone (24.3 ± 2.4%; n = 37; p < 0.05). Higher concentrations of forskolin resulted in dose-dependent inhibition of the rhGGF2-induced S-phase entry. The mitogenic response to rhGG2 was reduced by 26% in the presence of 10 μmforskolin (17.9 ± 0.15%; n = 28) and by 65% in 100 μm forskolin (8.5 ± 0.11%;n = 30) (p < 0.05 for each).
Like the treatments with 10 and 100 μm forskolin, long-term (72 hr) treatments with high concentrations of a membrane-permeant analog of cAMP, Br-cAMP, reduced the level of BrdU labeling that was induced in the presence of rhGGF2 (Fig.3). In the presence of rhGGF2, Br-cAMP at 100 μm (n = 22), 500 μm (n = 26), and 1 mm (n = 20) reduced the level of rhGGF2-induced S-phase entry by 70.8–80.3% when compared with epithelia that were cultured with rhGGF2 alone (p < 0.001; Fig. 3). However, similar to the combination of 1 μm forskolin and rhGGF2, a low concentration of Br-cAMP (4 μm) and rhGGF2 resulted in 39.2 ± 0.03% S-phase entry (n = 21;p < 0.01), a level significantly higher than the 24.3 ± 2.4% that was induced by rhGGF2 alone.
Brief incubation with forskolin or Br-cAMP stimulates proliferation
Treating epithelia for just 15 min with 1 μmforskolin in the standard medium resulted in a 12-fold increase in the level of S-phase entry during the subsequent 72 hr culture period, as indicated by the BrdU labeling of 17.3 ± 2.2% of the cells (n = 20). Other epithelia that were cultured in the same medium for the same time, but without the 15 min forskolin treatment, contained 1.5 ± 0.7% BrdU-labeled cells by comparison (n = 28) (Fig. 4).
To confirm that a brief elevation in intracellular cAMP level induces S-phase entry, we used the membrane-permeant cAMP analog Br-cAMP. A 60 min treatment with 0.5 mm Br-cAMP increased the level of BrdU labeling during the subsequent 72 hr culture period to 15.6 ± 2.3% (n = 23), which was more than a 10-fold increase from the level of BrdU labeling in epithelia cultured without such a 60 min treatment.
The role of PKA in the intracellular events induced by the brief forskolin treatment was assessed by treating epithelia with the PKA inhibitor KT5720. KT5720 induced a dose-dependent inhibition, reducing the effect of the brief exposure to forskolin by 54% at 1 μm (n = 20; p < 0.05) and by 92.4% at 5 μm (n = 20;p < 0.001) (Fig.5). Treatment with 5 μm KT5720 also inhibited the effect of the 1 hr treatment with 0.5 mm Br-AMP by 85.2% (n = 20; p < 0.001) (Fig. 5).
To investigate the hypothesis that the brief forskolin treatments might increase the level of membrane receptors, epithelia were treated with 1 μm forskolin for 15 min followed by incubation with rhGGF2 in the standard medium for 72 hr. During that treatment, 54.1 ± 3.8% of the cells were labeled with BrdU (n = 20), a level of labeling that was more than two times the level that was induced by a 72 hr treatment with rhGGF2 alone (24.3 ± 2.4%; p < 0.001) (Fig.6).
As a test for the unlikely possibility that forskolin and rhGGF2 were able to stimulate direct proliferation in hair cells, eight pieces of utricle epithelium were processed for immunocytochemistry with an anti-calretinin antibody (a hair cell marker) and the anti-BrdU antibody after the combined forskolin and rhGGF2 treatment; 201 ± 11 cells were labeled by anti-calretinin, on average, but none were double-labeled for BrdU, confirming that hair cells did not enter S-phase after such a treatment (data not shown).
Inhibition of receptor recycling blocks forskolin stimulation of proliferation
To assess whether the short-term effect of forskolin could result from increased receptor density, sheets of epithelia were treated with monensin, an ionophore, and bafilomycin A1, a macrolide antibiotic, which both can block or slow down the recycling of receptors to the plasma membrane (Basu et al., 1981; Mellman et al., 1986; Johnson et al., 1993; Presley et al., 1997). Treatment of the epithelia with 0.5 μm monensin abolished the increase in S-phase entry that was triggered by a 15 min treatment with 1 μm forskolin (0.18 ± 0.08%; n = 20; p< 0.001) (Fig. 7). Treatment with 0.1 μm bafilomycin A inhibited the forskolin effect by 75% (4.3 ± 0.7%; n = 20; p< 0.001) (Fig. 7).
To control for the possibility of cytotoxic effects of monensin, pieces of epithelia (n = 20) were treated with monensin for 75 min before 72 hr of culture in the standard medium plus rhGGF2. The 75 min period corresponded to the total time that epithelia were in the presence of monensin during the experiments that established the ability of monensin to block the effects of a 15 min forskolin treatment. The results demonstrated that epithelia treated with monensin were viable and responded to mitogenic stimulation by rhGGF2, but their levels of BrdU labeling were reduced by ∼50% (11.1 ± 1.6%; n = 20) from the levels in epithelia cultured in rhGGF2 without the monensin pretreatment (p < 0.05; Fig. 8). This established that the nearly complete block of the proliferative effect of the brief forskolin treatment was not simply the result of a cytotoxic action of monensin.
Fig. 8.
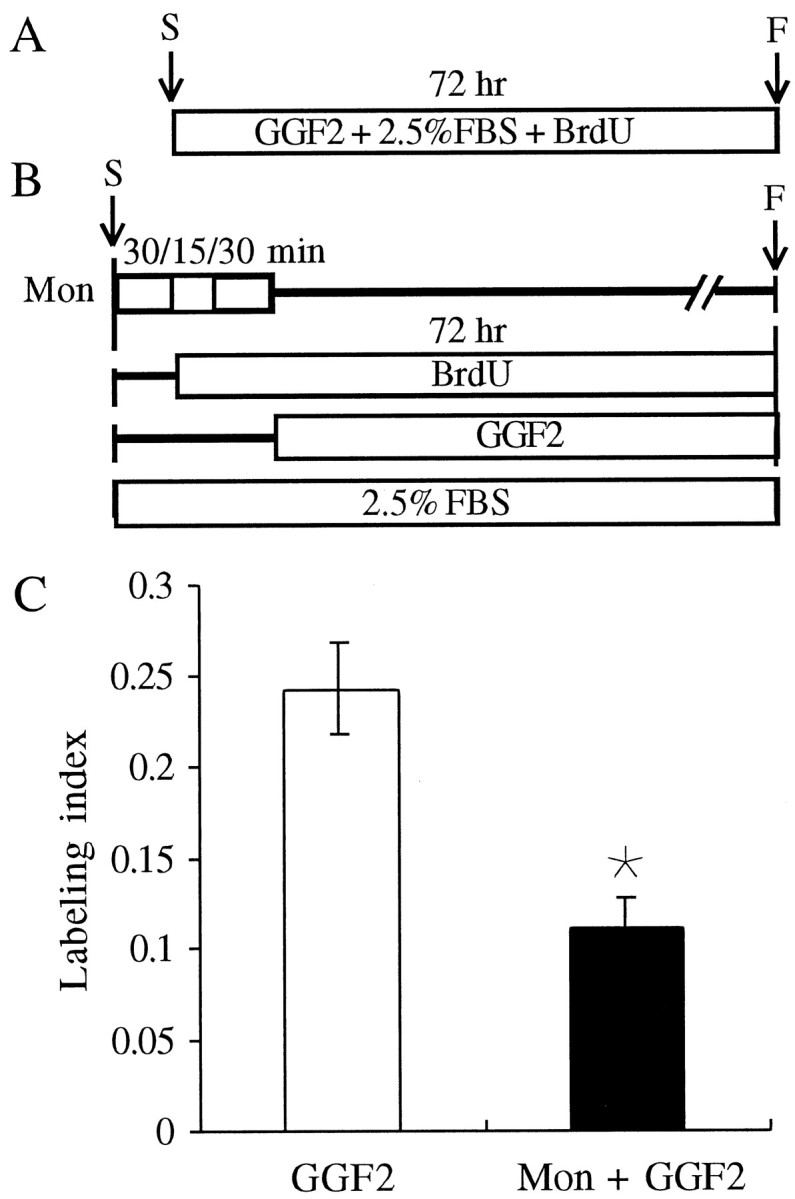
Monensin treatment inhibits the rhGGF2-induced S-phase entry by half. A, B, Timelines are shown.A, Pieces of epithelia were cultured in a standard medium containing 50 ng/ml rhGGF2 (GGF2) for 72 hr.B, Twenty pieces were pretreated with 0.5 μm monensin (Mon) for 30 min in a medium containing FBS but no BrdU. The medium was then changed for a fresh medium containing FBS and BrdU for 45 min. Then, the medium was replaced by a medium containing rhGGF2, FBS, and BrdU for the remainder of the 72 hr culture period. C, Treatment with monensin resulted in a significant reduction of the rhGGF2-induced effect, but we were still able to see ∼50% of the effect triggered by rhGGF2 alone. *, Significant difference when compared with GGF2(p < 0.05). F, Fixed;S, start.
Brief treatments with forskolin do not trigger S-phase entry in the absence of growth factors
To determine whether serum components are necessary for the forskolin-induced increases in S-phase entry, comparisons were made between sensory epithelia that were treated with forskolin in serum containing medium and others that were treated with forskolin in a defined medium without exogenous growth factors. The defined medium was composed of DMEM/F12 supplemented with 50 μg/ml transferrin, 5.2 ng/ml sodium selenite, 3.6 ng/ml hydrocortisone, and 10 ng/ml insulin. The cultures were treated with a 15 min pulse of 1 μmforskolin as described previously, but the entire experiment was performed using the defined medium in place of the standard medium that contained 2.5% FBS. In the absence of the exogenous growth factors in serum, the 15 min treatment did not induce increased S-phase entry (1.3 ± 0.1%; n = 20), although the same 15 min treatment with forskolin induced 17.3 ± 2.2% of the cells to enter S-phase when cultured in the presence of 2.5% serum as described above (Fig. 9).
DISCUSSION
Numerous studies associate the second messenger cAMP with the inhibition of cell proliferation in vitro. In this study, data show that cAMP can affect the level of S-phase entry of mammalian utricular supporting cells in a complex way.
Continuous cAMP treatment inhibits proliferation
When forskolin was present in the medium for the entire 72 hr culture period, we saw no proliferative response in utricular sensory epithelia. Moreover, treatments with high concentrations of forskolin (10 and 100 μm) significantly inhibited the rhGGF2-induced proliferation. The antiproliferative effect of forskolin was also observed after treatments with Br-cAMP, a membrane-permeant cAMP analog. In pheochromocytoma 12 (PC12) cells, 0.1 μmforskolin generates a 50–80% increase in cAMP as compared with basal levels (Mark et al., 1995), and 10 μm forskolin has been reported to raise the level of intracellular cAMP by 15-fold in the retina (Rehen et al., 1996). It is conceivable therefore that 10 or 100 μm forskolin raised the concentration of cAMP in the cells to levels beyond the physiological range and perhaps to levels that are toxic to the cells.
In numerous systems, an increase in cAMP is known to inhibit the mitogen-activated protein kinase (MAPK) cascade (for review, seeBurgering and Bos, 1995). Inhibition of the MAPK cascade can be exerted by cAMP at the level of raf, MAPK kinase (MEK), or MAPK in rat1HER fibroblasts (Wu et al., 1993). cAMP also has been reported to regulate negatively the activity of the mammalian target of rapamycin (mTOR) (Lin and Lawrence, 1996; Scott and Lawrence, 1998) and p70 S6K (Monfar et al., 1995), two kinases that appear to play pivotal roles in rhGGF2-induced S-phase entry in the mammalian utricle (Montcouquiol and Corwin, 2001). Forskolin (Kato et al., 1994) and Br-cAMP (Onishi and Hruska, 1997) have also been shown to increase levels of the cyclin-dependent kinase inhibitor p27 KIP1, a widely expressed tumor suppressor. The long-term exposure to high concentrations of forskolin may have suppressed S-phase entry in cells of the sensory epithelium by inhibiting one or more of those enzymes (see Fig.10).
Fig. 10.

A working model of forskolin regulation of S-phase entry in mammalian supporting cells. An increase in intracellular cAMP is achieved either via forskolin activating the AC or via Br-cAMP crossing the plasma membrane. Prolonged treatment (72 hr) with forskolin or Br-cAMP alone did not induce stimulation of S-phase entry. Prolonged treatments with Br-cAMP or high doses of forskolin (10–100 μm) in the presence of rhGGF2 may lead to the inhibition of components of the MAPK cascade such as Raf, MEK, or MAPK. Other intracellular intermediates such as mTOR or p70 S6K may also be inhibited by long-term treatments. These inhibitory cascades might involve the cAMP-dependent kinase PKA, which can in turn translocate to the nucleus to phosphorylate CREB. An exception occurred for 72 hr treatments with 1 μm forskolin that potentiated the rhGGF2 effect. This effect could be caused by the activation of the Rap1 kinase, which in turn can activate the MAPK cascade, or by stimulation of receptor recycling. The inhibition of the rhGGF2 effect by both forskolin and Br-cAMP could also be mediated via the increase in expression of a cyclin-dependent kinase inhibitor such as p27 KIP1. Brief treatments with forskolin (15 min) or Br-cAMP (1 hr) could increase the density of RTK, such as ErbB receptors, inserted at the membrane surface, allowing the binding of GF such as GGF2. This effect may be partly mediated by PKA-dependent activation and a nonidentified cAMP-dependent pathway. The inhibitors used are shown in boxes. AC, Adenylyl cyclase; CREB, cAMP-responsive element-binding protein;ErbBs, receptor tyrosine kinases of the ErbB family;GF, growth factor; PI3-K, phosphatidylinositol 3-kinase; p70 S6K, the70 kDa S6 protein kinase; RTK, receptor tyrosine kinase.
Short cAMP treatment stimulates proliferation
Although continuous treatments with forskolin or Br-cAMP alone failed to induce S-phase entry, continuous exposures to 1 μm forskolin or 4 μm Br-cAMP significantly potentiated the S-phase-inducing effect of rhGGF2. These results are consistent with evidence of synergistic effects of forskolin and another neuregulin in Schwann cells (Kim et al., 1997). Rahmatullah et al. (1998) also showed that the continuous presence of forskolin and heregulin (a growth factor related to GGF2) for at least 32 hr is necessary for induction of proliferation. Their results suggest that both factors are required to induce phosphorylation of CREB; each factor alone failed to induce significant phosphorylation.
In fact, combined application of cAMP and a growth factor such as platelet-derived growth factor (PDGF), epidermal growth factor, fibroblast growth factor, or GGF2 can promote a synergistic stimulation of cell proliferation (Westermark et al., 1986; Porter et al., 1987;Roger et al., 1987). Weinmaster and Lemke (1990) suggested that the synergistic proliferative effect obtained from the combination of cAMP and growth factors may result from the cAMP-mediated transcription of growth factor receptors. The authors further noted that the induction of the receptors could account for the observed mitogenicity of cAMP alone, because in vitro proliferation assays are typically conducted in the presence of serum, which contains low concentrations of PDGF and other growth factors. Indeed, Raff et al. (1978) had noted that elevation of intracellular cAMP does not trigger Schwann cell division if those cells are cultured in media containing 1% FBS or less, suggesting that growth factors in the serum were required for the cAMP-induced mitogenicity.
In our experiments, strong increases in S-phase entry were induced by a 15 min treatment with forskolin and by a 1 hr treatment with Br-cAMP. Both effects were strongly inhibited by KT5720, suggesting the involvement of PKA. Meyer-Franke et al. (1998) observed that a 1 hr treatment with forskolin induced a large, rapid increase in the number of tyrosine kinase receptors on the plasma membrane of retinal ganglion cells. The authors suggested that the cAMP elevation triggered the translocation of receptors from a vesicular pool to the plasma membrane. The increase in the receptors on the membrane enhanced the responsiveness of the retinal ganglion cells to trophic factors, thereby promoting increased survival. Our results with monensin and bafilomycin A1, drugs known to block receptor recycling, are consistent with a similar hypothesis. Inhibition of endosome acidification has been associated with reduced endocytic recycling and intracellular retention of recycling receptors (for review, see Mellman et al., 1986; Stevens and Forgac, 1997). Monensin exchanges sodium ions with protons across membranes, disrupting the proton gradient that maintains the acid pH (Mollenhauer et al., 1990), and bafilomycin A1 is an inhibitor of the vacuolar H+-ATPase (Bowman et al., 1988). When monensin was added to the culture medium before the short-term forskolin treatment, the cAMP-dependent increase in S-phase entry was strongly inhibited. Addition of bafilomycin A1 before the short-term forskolin treatment gave similar results. Both experiments support the hypothesis that the short-term forskolin treatment led to increased S-phase entry by increasing the density of as yet unidentified growth factor receptors on the plasma membrane. This could amplify the signal activated by growth factors in serum, leading to the proliferation.
Although receptor recycling that is independent of acidic endosomal pH has been reported (Mellman et al., 1984; Romanek et al., 1993), in this study tests with two acidification-neutralizing drugs resulted in strong inhibition of the forskolin effect. We acknowledge that monensin and bafilomycin A1 could have various side effects and that those effects could underlie the inhibition of BrdU labeling observed in these experiments (Tartakoff, 1983; Mollenhauer et al., 1990; Dinter and Berger, 1998). However, this reduction of BrdU incorporation could also reflect the inhibition of a basal level of receptor recycling involved in the response to growth factors in serum. In agreement with this, S-phase entry was abolished in cultures in serum-free medium without added growth factors (Fig. 9).
A prediction from the hypothesis that the increase in intracellular cAMP enhanced the density of growth factor receptors on the plasma membrane is that a short-term treatment with forskolin followed by incubation in a growth factor for one of the expressed receptors would produce a stronger effect than would exposure to the growth factor alone. The result obtained with rhGGF2 in combination with a short-term treatment with forskolin matched the prediction (Fig. 6). Also consistent with the hypothesis, the short-term forskolin treatment in serum-free medium failed to induce a significant increase in S-phase entry, which occurred when serum was present.
Our experimental results do not exclude the possibility of direct activation of intracellular signaling cascades by cAMP. cAMP can induce Ras-independent activation of MAPK in PC12 cells (Vossler et al., 1997), apparently via activation of the Ras-related protein Rap1 and PKA. Moreover cAMP can activate mTOR and is mitogenic in Swiss 3T3 cells (Withers et al., 1995) and protein kinase B (Sable et al., 1997).
Our results show that cAMP triggers a complex range of responses in mammalian utricular cells as compared with just one response reported for chick cochlear cells. In the chick auditory system, Navaratnam et al. (1996) showed that a 72–96 hr exposure to 100 μmforskolin led to increased supporting-cell proliferation and thereby to regeneration. In the rat utricle, an increase of S-phase entry occurred only when the epithelia are treated for a short period. In contrast to the results of Navaratnam et al. (1996), continuous exposure to forskolin alone, whatever the concentration, failed to induce significant S-phase entry (Fig. 1). An explanation for the discrepant findings may reside in the different culture conditions used.Navaratnam et al. (1996) used free-floating organ cultures and 10% fetal calf serum. Another possibility is that the 100 μmforskolin simply induced hair cell death in the chick's epithelia, where the loss of hair cells triggers spontaneous supporting-cell proliferation and regeneration (Corwin and Cotanche, 1988; Ryals and Rubel, 1988; Warchol and Corwin, 1996). In mammals, the loss of hair cells does not trigger strong levels of supporting-cell proliferation (Warchol et al., 1993), so even if the 100 μm forskolin killed hair cells, that would not have triggered a strong S-phase entry response. Moreover, a live/dead assay revealed no evidence of cell death induced by 100 μm forskolin. It is possible that forskolin activated signaling cascades in hair cells, but in our tests, cells that were immunoreactive for the hair cell marker calretinin were never BrdU-positive, consistent with a large literature suggesting that differentiated hair cells are effectively postmitotic (for review, seeCorwin and Oberholtzer, 1997).
It is conceivable that different signaling pathways in our system are activated depending on the dose and the duration of the forskolin treatment. Continuous treatment with low concentrations (0.1 or 1 μm) of the drug could have acted synergistically with rhGGF2 to induce S-phase entry by stimulating recycling and/or synthesis of growth factor receptors. When used at 10 or 100 μm, forskolin might inhibit one or multiple pathways, because of the very high level of cAMP in the cell, and lead to the inhibition of the rhGGF2-induced effect. In the brief treatment with forskolin, the cAMP could principally increase the density of growth factor receptors on the plasma membrane, and this effect might be mediated at least partly via PKA activation. In the long-term treatment, PKA might be strongly and continuously activated, leading mainly to inhibition of multiple intracellular signaling cascades (Fig.10).
Finally, the differences in the responses to forskolin between mammals and birds may also reflect important differences between species that do and those that do not regenerate hair cells throughout life. Understanding and using those differences may help us to find better approaches for inducing regeneration in the mammalian inner ear.
Footnotes
This work was supported by National Institute on Deafness and Other Communication Disorders Grant R01-DC00200. We thank Sherri Smith and Larry Phillips for their technical help, as well as Mark Witte, Kambiz Karimi, Jason Meyers, David Lenzi, and Veena Vasandani for helpful comments and critical reading of this manuscript. We also thank David Brautigan for valuable suggestions and Mark Marchionni of Cambridge NeuroScience, Inc., for the generous gift of rhGGF2.
Correspondence should be addressed to Dr. Mireille Montcouquiol, Department of Otolaryngology–Head, Neck, and Surgery and Department of Neuroscience, University of Virginia, School of Medicine, HSC Box 396, Cobb Hall, Charlottesville, VA 22908. E-mail:mm7vy@virginia.edu.
REFERENCES
- 1.Basu SK, Goldstein JL, Anderson RG, Brown MS. Monensin interrupts the recycling of low density lipoprotein receptors in human fibroblasts. Cell. 1981;24:493–502. doi: 10.1016/0092-8674(81)90340-8. [DOI] [PubMed] [Google Scholar]
- 2.Bowman EJ, Siebers A, Altendorf K. Bafilomycins: a class of inhibitors of membrane ATPases from microorganisms, animal cells and plant cells. Proc Natl Acad Sci USA. 1988;85:7972–7976. doi: 10.1073/pnas.85.21.7972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Burgering BM, Bos JL. Regulation of Ras-mediated signalling: more than one way to skin a cat. Trends Biochem Sci. 1995;20:18–22. doi: 10.1016/s0968-0004(00)88944-6. [DOI] [PubMed] [Google Scholar]
- 4.Corwin JT, Cotanche DA. Regeneration of sensory hair cells after acoustic trauma. Science. 1988;240:1772–1774. doi: 10.1126/science.3381100. [DOI] [PubMed] [Google Scholar]
- 5.Corwin JT, Oberholtzer JC. Fish n' chicks: model recipes for hair-cell regeneration? Neuron. 1997;19:951–954. doi: 10.1016/s0896-6273(00)80386-4. [DOI] [PubMed] [Google Scholar]
- 6.Dinter A, Berger EG. Golgi-disturbing agents. Histochem Cell Biol. 1998;109:571–590. doi: 10.1007/s004180050256. [DOI] [PubMed] [Google Scholar]
- 7.Forge A, Li L, Corwin JT, Nevill G. Ultrastructural evidence for hair cell regeneration in the mammalian inner ear. Science. 1993;259:1616–1619. doi: 10.1126/science.8456284. [DOI] [PubMed] [Google Scholar]
- 8.Johnson LS, Dunn KW, Pytowski B, McGraw TE. Endosome acidification and receptor trafficking: bafilomycin A1 slows receptor externalization by a mechanism involving the receptor's internalization motif. Mol Biol Cell. 1993;4:1251–1266. doi: 10.1091/mbc.4.12.1251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kato JY, Matsuoka M, Polyak K, Massague J, Sherr CJ. Cyclic AMP-induced G1 phase arrest mediated by an inhibitor (p27Kip1) of cyclin-dependent kinase 4 activation. Cell. 1994;79:487–496. doi: 10.1016/0092-8674(94)90257-7. [DOI] [PubMed] [Google Scholar]
- 10.Kim HA, DeClue JE, Ratner N. cAMP-dependent protein kinase A is required for Schwann cell growth: interactions between the cAMP and neuregulin/tyrosine kinase pathways. J Neurosci Res. 1997;49:236–234. [PubMed] [Google Scholar]
- 11.Kuntz AL, Oesterle EC. Transforming growth factor alpha with insulin stimulates cell proliferation in vivo in adult rat vestibular sensory epithelium. J Comp Neurol. 1998;399:413–423. [PubMed] [Google Scholar]
- 12.Lambert PR. Inner ear hair cell regeneration in a mammal: identification of a triggering factor. Laryngoscope. 1994;104:701–718. doi: 10.1288/00005537-199406000-00010. [DOI] [PubMed] [Google Scholar]
- 13.Lambert PR, Gu R, Corwin JT. Analysis of small hair bundles in the utricles of mature guinea pigs. Am J Otol. 1997;18:637–643. [PubMed] [Google Scholar]
- 14.Lin TA, Lawrence JC., Jr Control of the translational regulators PHAS-I and PHAS-II by insulin and cAMP in 3T3–L1 adipocytes. J Biol Chem. 1996;271:30199–30204. doi: 10.1074/jbc.271.47.30199. [DOI] [PubMed] [Google Scholar]
- 15.Mark MD, Liu Y, Wong ST, Hinds TR, Storm DR. Stimulation of neurite outgrowth in PC12 cells by EGF and KCl depolarization: a Ca(2+)-independent phenomenon. J Cell Biol. 1995;130:701–710. doi: 10.1083/jcb.130.3.701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mellman I, Plutner H, Ukkonen P. Internalization and rapid recycling of macrophage Fc receptors tagged with monovalent antireceptor antibody: possible role of a prelysosomal compartment. J Cell Biol. 1984;98:1163–1169. doi: 10.1083/jcb.98.4.1163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mellman I, Fuchs R, Helenius A. Acidification of the endocytic and exocytic pathways. Annu Rev Biochem. 1986;55:663–700. doi: 10.1146/annurev.bi.55.070186.003311. [DOI] [PubMed] [Google Scholar]
- 18.Meyer-Franke A, Wilkinson GA, Kruttgen A, Hu M, Munro E, Hanson MG, Jr, Reichardt LF, Barres BA. Depolarization and cAMP elevation rapidly recruit TrkB to the plasma membrane of CNS neurons. Neuron. 1998;21:681–693. doi: 10.1016/s0896-6273(00)80586-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mollenhauer HH, Morre DJ, Rowe LD. Alteration of intracellular traffic by monensin; mechanism, specificity and relationship to toxicity. Biochim Biophys Acta. 1990;1031:225–246. doi: 10.1016/0304-4157(90)90008-Z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Monfar M, Lemon KP, Grammer TC, Cheatham L, Chung J, Vlahos CJ, Blenis J. Activation of pp70/85 S6 kinases in interleukin-2-responsive lymphoid cells is mediated by phosphatidylinositol 3-kinase and inhibited by cyclic AMP. Mol Cell Biol. 1995;15:326–337. doi: 10.1128/mcb.15.1.326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Montcouquiol M, Corwin JT (2001) Intracellular signals that control cell proliferation in mammalian balance epithelia: key roles for phosphatidylinositol-3 kinase, mammalian target of rapamycin, and S6 kinases in preference to calcium protein kinase C, and mitogen-activated protein kinase. J Neurosci, in press. [DOI] [PMC free article] [PubMed]
- 22.Navaratnam DS, Su HS, Scott S-P, Oberholtzer C. Proliferation in the auditory receptor epithelium mediated by cyclic AMP-dependent signaling pathway. Nat Med. 1996;2:1136–1139. doi: 10.1038/nm1096-1136. [DOI] [PubMed] [Google Scholar]
- 23.Onishi T, Hruska K. Expression of p27Kip1 in osteoblast-like cells during differentiation with parathyroid hormone. Endocrinology. 1997;138:1995–2004. doi: 10.1210/endo.138.5.5146. [DOI] [PubMed] [Google Scholar]
- 24.Porter S, Glaser L, Bunge RP. Release of autocrine growth factor by primary and immortalized Schwann cells. Proc Natl Acad Sci USA. 1987;84:7768–7772. doi: 10.1073/pnas.84.21.7768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Presley JF, Mayaor S, McGraw TE, Dunn KW, Maxfield FR. Bafilomycin A1 treatment retards transferrin receptor recycling more than bulk membrane recycling. J Biol Chem. 1997;272:13929–13936. doi: 10.1074/jbc.272.21.13929. [DOI] [PubMed] [Google Scholar]
- 26.Raff MC, Hornby-Smith A, Brockes JP. Cyclic AMP as a mitogenic signal for cultured rat Schwann cells. Nature. 1978;273:672–673. doi: 10.1038/273672a0. [DOI] [PubMed] [Google Scholar]
- 27.Rahmatullah M, Schroering A, Rothblum K, Stahl RC, Urban B, Carey DJ. Synergistic regulation of Schwann cell proliferation by heregulin and forskolin. Mol Cell Biol. 1998;18:6245–6252. doi: 10.1128/mcb.18.11.6245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Rehen SK, Varella MH, Freitas FG, Moraes MO, Linden R. Contrasting effects of protein synthesis inhibition and of cyclic AMP on apoptosis in the developing retina. Development. 1996;122:1439–1448. doi: 10.1242/dev.122.5.1439. [DOI] [PubMed] [Google Scholar]
- 29.Roger PP, Servais P, Dumont JE. Induction of DNA synthesis in dog thyrocytes in primary culture: synergistic effects of thyrotropin and cyclic AMP with epidermal growth factor and insulin. J Cell Physiol. 1987;130:58–67. doi: 10.1002/jcp.1041300110. [DOI] [PubMed] [Google Scholar]
- 30.Romanek R, Sargeant R, Paquet MR, Gluck S, Klip A, Grinstein S. Chloroquine inhibits glucose-transporter recruitment induced by insulin in rat adipocytes independently of its action on endomembrane pH. Biochem J. 1993;296:321–327. doi: 10.1042/bj2960321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ryals BM, Rubel EW. Hair cell regeneration after acoustic trauma in adult Coturnix quail. Science. 1988;240:1774–1776. doi: 10.1126/science.3381101. [DOI] [PubMed] [Google Scholar]
- 32.Sable CL, Filippa N, Hemmings B, Van Obberghen E. cAMP stimulates protein kinase B in a Wortmannin-insensitive manner. FEBS Lett. 1997;409:253–257. doi: 10.1016/s0014-5793(97)00518-8. [DOI] [PubMed] [Google Scholar]
- 33.Saffer LD, Gu R, Corwin JT. An RT-PCR analysis of mRNA for growth factor receptors in damaged and control sensory epithelia of rat utricles. Hear Res. 1996;94:14–23. doi: 10.1016/0378-5955(95)00228-6. [DOI] [PubMed] [Google Scholar]
- 34.Scott PH, Lawrence JC., Jr Attenuation of mammalian target of rapamycin activity by increased cAMP in 3T3–L1 adipocytes. J Biol Chem. 1998;273:34496–34501. doi: 10.1074/jbc.273.51.34496. [DOI] [PubMed] [Google Scholar]
- 35.Shaywitz AJ, Greenberg ME. CREB: a stimulus-induced transcription factor activated by a diverse array of extracellular signals. Annu Rev Biochem. 1999;68:821–861. doi: 10.1146/annurev.biochem.68.1.821. [DOI] [PubMed] [Google Scholar]
- 36.Stevens TH, Forgac M. Structure, function and regulation of the vacuolar (H+)-ATPase. Annu Rev Cell Dev Biol. 1997;13:779–808. doi: 10.1146/annurev.cellbio.13.1.779. [DOI] [PubMed] [Google Scholar]
- 37.Tartakoff AM. Perturbation of vesicular traffic with the carboxylic ionophore monensin. Cell. 1983;32:1026–1028. doi: 10.1016/0092-8674(83)90286-6. [DOI] [PubMed] [Google Scholar]
- 38.Vossler MR, Yao H, York RD, Pan MG, Rim CS, Stork PJ. cAMP activates MAP kinase and Elk-1 through a B-Raf- and Rap1-dependent pathway. Cell. 1997;89:73–82. doi: 10.1016/s0092-8674(00)80184-1. [DOI] [PubMed] [Google Scholar]
- 39.Warchol ME, Corwin JT. Regenerative proliferation in organ cultures of the avian cochlea: identification of the initial progenitors and determination of the latency of the proliferative response. J Neurosci. 1996;16:5466–5477. doi: 10.1523/JNEUROSCI.16-17-05466.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Warchol ME, Lambert PR, Goldstein BJ, Forge A, Corwin JT. Regenerative proliferation in inner ear sensory epithelia from adult guinea pigs and humans. Science. 1993;259:1619–1622. doi: 10.1126/science.8456285. [DOI] [PubMed] [Google Scholar]
- 41.Weinmaster G, Lemke G. Cell-specific cyclic AMP-mediated induction of the PDGF receptor. EMBO J. 1990;9:915–920. doi: 10.1002/j.1460-2075.1990.tb08189.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Westermark K, Westermark B, Karlsson FA, Ericson LE. Location of epidermal growth factor receptors on porcine thyroid follicle cells and receptor regulation by thyrotropin. Endocrinology. 1986;118:1040–1046. doi: 10.1210/endo-118-3-1040. [DOI] [PubMed] [Google Scholar]
- 43.Withers DJ, Bloom SR, Rozengurt E. Dissociation of cAMP-stimulated mitogenesis from activation of the mitogen-activated protein kinase cascade in Swiss 3T3 cells. J Biol Chem. 1995;270:21411–21419. doi: 10.1074/jbc.270.36.21411. [DOI] [PubMed] [Google Scholar]
- 44.Wu J, Dent P, Jelinek T, Wolfman A, Weber MJ, Sturgill TW. Inhibition of the EGF-activated MAP kinase signaling pathway by adenosine 3′,5′-monophosphate. Science. 1993;262:1065–1069. doi: 10.1126/science.7694366. [DOI] [PubMed] [Google Scholar]
- 45.Yamashita H, Oesterle EC. Induction of cell proliferation in mammalian inner ear sensory epithelia by transforming growth factor α and epidermal growth factor. Proc Natl Acad Sci USA. 1995;92:3152–3155. doi: 10.1073/pnas.92.8.3152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zheng JL, Helbig C, Gao W-Q. Induction of cell proliferation by fibroblast and insulin-like growth factors in pure rat inner ear epithelial cell cultures. J Neurosci. 1997;17:216–226. doi: 10.1523/JNEUROSCI.17-01-00216.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]