

Abstract
In Drosophila photoreceptors the multivalent PDZ protein INAD organizes the phototransduction cascade into a macromolecular signaling complex containing the effector PLC, the light-activated TRP channels, and a regulatory PKC. Previously, we showed that the subcellular localization of INAD signaling complexes is critical for signaling. Now we have examined how INAD complexes are anchored and assembled in photoreceptor cells. We find thattrp mutants, or transgenic flies expressinginaD alleles that disrupt the interaction between INAD and TRP, cause the mislocalization of the entire transduction complex. The INAD–TRP interaction is not required for targeting but rather for anchoring of complexes, because INAD and TRP can be targeted independently of each other. We also show that, in addition to its scaffold role, INAD functions to preassemble transduction complexes. Preassembly of signaling complexes helps to ensure that transduction complexes with the appropriate composition end up in the proper location. This may be a general mechanism used by cells to target different signaling machinery to the pertinent subcellular location.
Keywords: INAD, signaling complex, transducisome, Drosophila, phototransduction, subcellular localization, signal transduction, anchoring, assembly
Every cell must sort through a vast array of external signals and transduce them into the appropriate intracellular responses. Because many intracellular signaling cascades share common downstream components, an important strategy for maintaining specificity within one pathway, while avoiding unwanted cross-talk between different pathways, is the organization of transduction pathways into distinct signaling complexes (Pawson and Scott, 1997; Tsunoda et al., 1998). Scaffold proteins function as organizers of transduction pathways, bringing together signaling molecules into physically defined signaling units. This strategy enables a cell to promote specificity and selectivity while maximizing the speed of signaling.
In Drosophila phototransduction, speed of signaling is critical for achieving the temporal resolution necessary for a flying organism. Phototransduction in Drosophila is the fastest known G-protein-coupled signaling cascade, taking just a few tens of milliseconds to go from light activation of rhodopsin to the generation of a receptor potential and <100 msec to terminate the response (Ranganathan et al., 1995). In this pathway, light stimulation of rhodopsin activates a G-protein, which then activates a PLC. PLC catalyzes the hydrolysis of phosphatidylinositol-4,5-bisphosphate (PIP2) into inositol trisphosphate (IP3) and diacylglycerol (DAG), leading to the eventual opening and modulation of the light-activated ion channels, transient receptor potential (TRP), and TRP-like (TRPL). Calcium-dependent regulatory processes involving the activation of an eye-specific protein kinase C (eye-PKC), calmodulin (cam), arrestin, and cam-kinase then mediate deactivation of the light response (Ranganathan et al., 1991; Smith et al., 1991; Hardie et al., 1993).
An important strategy used by Drosophila photoreceptors to attain an extremely high speed of signaling is the organization of signaling components into transduction complexes by the scaffold protein INAD (these complexes are referred to as transducisomes; Huber et al., 1996; Shieh and Zhu, 1996; Chevesich et al., 1997; Tsunoda et al., 1997). INAD contains five PDZ domains, each binding a component of the phototransduction cascade: PDZ1 and PDZ5 bind PLC (Shieh et al., 1997; Tsunoda et al., 1997; van Huizen et al., 1998), PDZ2 and PDZ4 bind eye-PKC (Tsunoda et al., 1997; Adamski et al., 1998), and PDZ3 binds TRP (Shieh and Zhu, 1996; Tsunoda et al., 1997). This organization brings signaling components into close proximity, thus promoting (1) efficient signaling and (2) the creation of small microdomains in which localized changes in the level of intracellular Ca2+ can exert exquisite modulation of the light response (Hardie, 1991; Ranganathan et al., 1991; Scott and Zuker, 1997; Scott et al., 1997).
In wild-type flies, the INAD transducisomes localize to the rhabdomeres, a subcellular compartment consisting of 60,000 microvilli that house the ∼108 molecules of rhodopsin found in Drosophila photoreceptors. IninaD null mutants PLC, TRP, and eye-PKC are distributed randomly in the photoreceptors; this mislocalization leads to a near-complete loss of signaling (Tsunoda et al., 1997). These findings demonstrate that the localization of signaling components is dependent on the presence of the INAD scaffold protein and that it is not the mere presence of signaling molecules that is critical for effective signaling, but rather their location.
Studies in a number of other systems have validated the importance of PDZ proteins in the subcellular localization of signaling components and cellular signaling (Kim et al., 1995, 1996; Brenman et al., 1996;Dong et al., 1997; Tejedor et al., 1997; Zito et al., 1997; Torres et al., 1998) (for review, see Sheng, 1996; Kornau et al., 1997; Craven and Bredt, 1998; O'Brien et al., 1998; Tsunoda et al., 1998). For example, in Caenorhabditis elegans, the development of the vulva is dependent on a Ras signaling pathway that is mediated by the LET-23 EGF receptor. LIN-2, LIN-7, and LIN-10 are all PDZ-containing proteins essential for localizing the receptor to the basolateral membrane of epithelial cells; mutations inlin-2, lin-7, or lin-10 lead to the mislocalization of LET-23 receptors and result in a vulvaless phenotype (Hoskins et al., 1996; Simske et al., 1996; Kaech et al., 1998). InDrosophila, the polarization of epithelial cells is dependent on the PDZ scaffold protein Disks Lost (Dlt), which interacts with Crumbs and Neurexin IV (Bhat et al., 1999). Similarly, the PDZ protein Bazooka interacts with the Inscutable protein and is required for the apical localization of Inscutable and proper asymmetric division in Drosophila neuroblasts (Schober et al., 1999). Target proteins also play a role in localizing scaffold proteins. For instance, when PSD-95 is transfected into rat pyramidal cells, it localizes only to the dendrites; however, when it is cotransfected with Kv1.4 potassium channel, it also localizes to the axons (Arnold and Clapham, 1999).
In this study, we show that the TRP ion channel is essential for the localization of the INAD scaffold protein and that the TRP–INAD interaction is required for maintaining transduction complexes in the rhabdomeres. We also examined how transduction complexes are assembled and show that signaling complexes are preassembled before they are targeted to the rhabdomeres. The preassembly of transduction complexes may emerge as a powerful strategy in the organization of cellular signaling.
MATERIALS AND METHODS
Immunostaining cross sections of photoreceptors.INAD, TRP, eye-PKC, PLC, rhodopsin, and Dgq were detected by using polyclonal antibodies as previously described (Smith et al., 1991;Scott et al., 1995; Niemeyer et al., 1996; Tsunoda et al., 1997). For immunostaining cross sections of photoreceptors, we first fixed fly heads in 3% paraformaldehyde in PBS for 1 hr on ice and infiltrated them with 2.3 m sucrose in PBS overnight; finally, they were frozen and cut into 1-μm-thick sections. Tissue sections were treated with a blocking solution of 1% BSA and 0.1% saponin in PBS (PBS-S) for 30 min and then incubated with antibody either at 4°C overnight (INAD, TRP, Dgq, rhodopsin) or for 2 hr at room temperature (eye-PKC, PLC). Antibodies were diluted 1:300 (INAD), 1:20 (eye-PKC), and 1:100 (rhodopsin and Dgq) in PBS-S; the TRP and PLC antibodies were preabsorbed first with a homogenate of trpor norpA mutant heads to reduce background staining and were used at a final dilution of 1:100 (TRP) or 1:500 (PLC). FITC and lissamine rhodamine-conjugated secondary antibodies (Jackson ImmunoResearch, West Grove, PA) were used at 1:500 (FITC) or 1:200 (rhodamine) for a 1 hr incubation at room temperature.
Electron microscopic localization of INAD. Fly heads were bisected longitudinally and fixed in periodate–lysine–paraformaldehyde (PLP) solution (McLean and Nakane, 1974) for 1.5 hr at room temperature. Then the specimens were dehydrated partially with 90% ethanol, embedded in LR White medium (Polysciences, Warrington, PA), and cut into 80-nm-thick sections. The sections were picked up on Formvar-coated nickel grids, etched with saturated aqueous sodium meta-periodate (Sigma, St. Louis, MO) for 15 min, treated with a blocking solution of 5% normal goat serum and 0.05% Tween 20 (Wako Pure Chemical Industries, Osaka, Japan) in PBS (5NGS/PBST) for 20 min, and then incubated with anti-INAD antibody diluted 1:100 with 5NGS/PBST at 6°C overnight. After several rinses with PBST the sections were incubated with 10 nm colloidal gold-conjugated secondary antibody (British BioCell International, Cardiff, UK) diluted 1:50 with 5NGS/PBST for 2 hr at room temperature. The sections were counterstained with 2% aqueous uranyl acetate and Reynolds' lead solution (Reynolds, 1963) and then examined in a JEM 1200EX electron microscope (Jeol, Tokyo, Japan).
DNA constructs and transgenic flies. For inaD PDZ mutants three amino acid substitutions were made in each PDZ domain: PDZ1 (phe28ala, ile30ala, ile32ala), PDZ2 (leu260ala, leu262ala, leu264ala), PDZ3 (leu375ala, ile377ala, val379ala), and PDZ4 (met499ala, val501ala, val503ala). Mutant inaD constructs were cloned into a P-element-mediated transformation vector containing five Glass-binding sites derived from the ninaE promoter (pGMR; Hay et al., 1994). For hs-inaD flies, inaDcDNA was cloned into a P-element transformation vector under the control of the heat shock promoter (Baker et al., 1994). FortrpC34 flies a truncatedtrp (with coding deleted for the C-terminal 34 amino acids) cDNA was cloned into the pGMR transformation vector.Drosophila P-element-mediated transformations and further genetic manipulations were performed with standard techniques.
Expression and analysis of proteins in Chinese hamster ovary (CHO) cells. inaD and mutant inaD cDNAs were cloned into the pcDNA3 transfection vector (Invitrogen, San Diego, CA), which uses the cytomegalovirus immediate-early (CMV) promoter for expression. CHO cells were transfected with Lipofectamine reagent (Life Technologies, Grand Island, NY) and grown for 24 hr in 10% fetal bovine serum, 0.5 μg/ml amphotericin B, and 100 μg/ml gentamycin in MEM α media (Life Technologies) at 37°C; then they were grown for an additional 24 hr in 10% fetal bovine serum, 100 U/ml penicillin, and 100 μg/ml streptomycin in Leibovitz's L-15 media (Life Technologies) at room temperature. Cells were fixed with 2% paraformaldehyde in PBS for 40 min and stained similarly to tissue sections (described above). Confocal images were collected on a Nikon/Bio-Rad 1024 confocal microscope and imported into Photoshop 5.0 (Adobe Systems, Mountainview, CA) for presentation.
Immunoprecipitation. Frozen heads (500–2000) were homogenized in 1 ml of Buffer A [containing (in mm) 20 HEPES, 30 NaCl, and 1 EDTA, pH 7.5, plus protease inhibitors] with a glass–glass homogenizer. The homogenate was centrifuged at 4000 × g for 1 min to remove chitinous material. Membranes were isolated by centrifugation at 100,000 × g for 30 min at 4°C and resuspended to a final equivalent concentration of 2 heads/μl (wild-type, inaD mutants).
For the cross-linking anti-INAD antibody to protein A beads, every 50 μl of anti-INAD antibody was incubated with 30 μl of ImmunoPure-immobilized protein A beads (Pierce, Rockford, IL) in a final volume of 170 μl of ImmunoPure IgG binding buffer (Pierce) overnight at 4°C. Antibody and beads then were cross-linked in 20 mm dimethyl dimelimidate (DMP) and 0.2 mmsodium borate, pH 9.0, for 30 min. Reaction was stopped with 0.2m ethanolamine, pH 8.0. Beads were stored in PBS.
For immunoprecipitations, the membranes (wild-type, inaDmutant, equivalent of membranes from 50 heads; hs-inaD, equivalent of 500 heads) were solubilized in 1% Triton X-100 and incubated with anti-INAD antibody cross-linked to protein A beads (bed volume of 20 μl) in a 100 μl total volume overnight at 4°C. Then the beads were washed in Buffer A, resuspended in SDS buffer, and boiled; the entire immunoprecipitate was fractionated by SDS-PAGE.
RESULTS
Subcellular localization of INAD signaling complexes requires the TRP channel
Given that the subcellular localization of signaling components is essential for proper signaling (Tsunoda et al., 1997), we set out to investigate how INAD signaling complexes are maintained (anchored) in the rhabdomeres. To test whether INAD is anchored through one of its known partners, we examined the subcellular distribution of INAD in mutants lacking individual partners. We used null alleles oftrp, inaC, and norpA to eliminate TRP, eye-PKC, and PLC, respectively. Figure1A shows that, much like in wild-type cells, INAD is localized correctly to the rhabdomeres in inaC and norpA mutant photoreceptors. In contrast, INAD is severely mislocalized in null trp mutants, with most of the protein found in the cell body instead of the rhabdomeres (Fig. 1A). Because INAD itself is essential for the localization of TRP, eye-PKC, and PLC (Tsunoda et al., 1997), we investigated whether eye-PKC and PLC are mislocalized in null trp alleles. Indeed, immunolocalization studies demonstrated that eye-PKC and PLC are mislocalized in trpmutant photoreceptors (Fig. 1B). Interestingly, eye-PKC and PLC are still coimmunoprecipitated with INAD intrp null mutants (data not shown), suggesting that complexes are stable even when they are mislocalized (see below).
Fig. 1.

TRP is required for the localization of INAD signaling complexes. A, Anti-INAD immunofluorescent staining of cross sections (1 μm thick) of wild-type (wt), inaC209,norpAP41, andtrp343 null mutant photoreceptors. Normal rhabdomeric localization of INAD is seen ininaC209 andnorpAP41 mutants, whereas it is mislocalized severely in trp343mutants. B, INAD, PLC, and eye-PKC also are mislocalized in trp mutants. Shown is immunofluorescent staining of cross sections of trp343 null photoreceptors. Note that Gα, the G-protein protein that shuttles between activated rhodopsin and transducisomes (Bahner et al., 2000), shows normal rhabdomeric localization. C, EM immunogold localization of INAD in wild-type andtrp343 rhabdomeres, demonstrating that ∼25% of the INAD seen in wild-type rhabdomeres still remains intrp mutant rhabdomeres. We cannot exclude the possibility that this small amount of INAD may be binding to TRPL channels in the rhabdomeres.
To demonstrate that it is the specific interaction of INAD with TRP that is required for the rhabdomeric localization of INAD transducisomes, we generated two transgenic lines expressing mutant proteins predicted to disrupt the TRP–INAD interaction. In the first line, inaDPDZ3, three conserved residues in the third PDZ domain of INAD (PDZ3; the site of TRP binding) were changed to alanines (leu375ala, ile377ala, val379ala). These mutations are expected to disrupt the interaction between PDZ domains and their targets (Doyle et al., 1996). The second transgenic line, trpC34, expresses a mutant TRP protein that lacks its INAD binding site (a truncation of the C-terminal 34 amino acids; Shieh and Zhu, 1996). As predicted, TRP and INAD are mislocalized in inaDPDZ3and trpC34 mutants (Fig.2). Because INAD is mislocalized ininaDPDZ3 andtrpC34 mutants, eye-PKC and PLC are mislocalized also (data not shown). Taken all together, these results substantiate the requirement for TRP in the maintenance of INAD complexes in the rhabdomeres and strongly suggest that part of thetrp phenotype may be attributable to the mislocalization of transducisomes (see below).
Fig. 2.
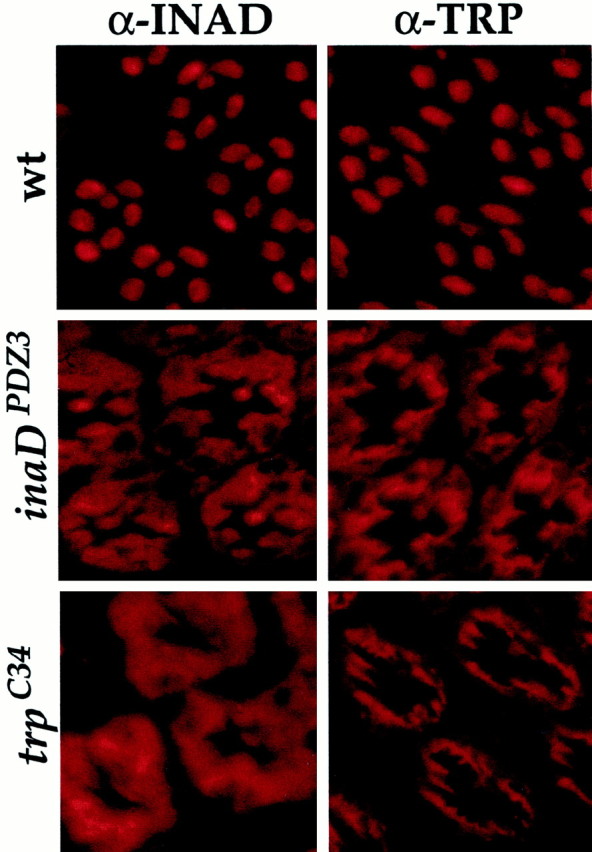
Mutations that disrupt the INAD–TRP interaction display mislocalization of INAD and TRP. Shown is immunofluorescent staining for INAD and TRP in cross sections (1 μm thick) of wild-type (wt), inaDPDZ3, andtrpC34 transgenic photoreceptors (as indicated). inaDPDZ3 flies express an INAD protein containing three point mutations expected to disrupt the INAD–TRP interaction (see Materials and Methods).trpC34 flies express a truncated TRP protein that is missing its PDZ binding site (C-terminal 34 amino acids deleted). INAD and TRP both are mislocalized severely ininaDPDZ3 andtrpC34 transformants. Given that rhodopsin (Rh1) is not part of the INAD signaling complex (Tsunoda et al., 1997; Huber et al., 1998; B. H. Shieh, personal communication) and that Gα shuttles between Rh1 and the transducisome (Tsunoda et al., 1997; Bahner et al., 2000), we examined the localization of Rh1 and Gα as controls for normal rhabdomeric labeling. Indeed, rhodopsin and Gα are localized normally intrp, inaDPDZ3, andtrpC34 mutant backgrounds (data not shown).
INAD and TRP are targeted independently to the rhabdomeres
Because INAD is mislocalized in trp mutants and TRP is mislocalized in inaD mutants, we wondered about the relationship between TRP and INAD for targeting/anchoring to the rhabdomeres. For example, does one protein depend on the other for targeting to the rhabdomeres, and/or does one require the other for maintenance in the rhabdomeres? To investigate whether either TRP or INAD relies on the other for targeting to the rhabdomeres, we examined INAD and TRP at a time when rhabdomeres are completing their maturation and transduction proteins are first synthesized. Figure3 shows that, in trp mutant photoreceptors, INAD initially is delivered and localized to the rhabdomeres of pupae but then becomes mislocalized by the time of eclosion. Similarly, in inaD null photoreceptors, TRP initially is targeted to the rhabdomeres of late pupae but becomes mislocalized in newly eclosed flies (Fig. 3). These results demonstrate that TRP and INAD do not depend on each other to be targeted to the rhabdomeres, but they need each other to remain in the rhabdomeres.
Fig. 3.
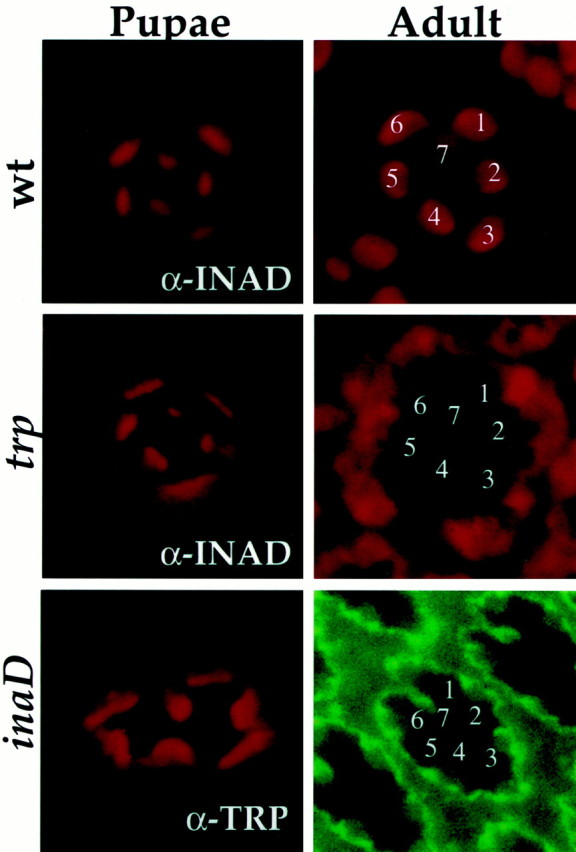
INAD and TRP are targeted independently to the rhabdomeres in pupae but require each other to be maintained in the rhabdomeres. Shown is anti-INAD and anti-TRP (as indicated) immunofluorescent staining of cross sections (1 μm thick) of wild-type (wt),trp343, andinaD1 null photoreceptors from pupae and adult flies. In wild type the INAD (top row) and TRP (data not shown) are targeted to the rhabdomeres in pupae and remain localized in the rhabdomeres of adult photoreceptors. Intrp343 mutants (middle row) INAD is targeted to the rhabdomeres in pupae but becomes mislocalized in adult photoreceptors. Similarly, ininaD1 null mutants (bottom row) TRP is targeted to the rhabdomeres in pupae but becomes mislocalized in adult photoreceptors. Rhabdomeres of individual R1–R7 photoreceptors are indicated by numbers.
PDZ1 also is required for localization of INAD
Examination of the signaling properties of inaD andtrp mutants reveals a salient difference between the two mutants. Although both inaD and trp null mutants show severe mislocalization of signaling components, light responses are robust in trp mutants but are extraordinarily poor ininaD mutants (Hardie and Minke, 1992; Tsunoda et al., 1997). Because signaling is fully dependent on the rhabdomeric localization of transduction components (Tsunoda et al., 1997), we reasoned thattrp mutants may have a small amount of INAD signaling complexes remaining in the rhabdomeres (like theinaD215 allele; see Tsunoda et al., 1997), and this may be sufficient to activate the TRPL channels (this is the light-activated channel remaining in trp mutants;Niemeyer et al., 1996). To test this postulate, we performed electron microscopic immunogold labeling (immunoEM) of INAD. Figure1C shows that trp mutants indeed have ∼25% of the level of rhabdomeric INAD of wild-type controls; this likely underlies the signaling seen in trp mutants.
Given that there is 25% of INAD in the rhabdomeres of trpmutants, we wondered whether INAD might be anchored to the rhabdomeric membrane through another one of its PDZ domains. We generated transgenic animals expressing INAD proteins each containing point mutations in one of the five PDZ domains and examined their localization. inaDPDZ2 andinaDPDZ5 mutants displayed normal rhabdomeric localization of INAD proteins (data not shown), suggesting that PDZ2 and PDZ5 do not play a role in anchoring. As expected,inaDPDZ3 mutants showed a mislocalization of INAD protein (see Fig. 2).inaDPDZ1 andinaDPDZ4 transformants, however, did not produce stable proteins in vivo and we were unable to test them. To investigate the membrane association of these two PDZ mutants in a different system, we used a tissue culture assay to monitor membrane localization of INAD and its targets.
We expressed inaD in CHO cells and examined its subcellular localization with confocal microscopy. Wild-type INAD localizes to the periphery of the cell, in close proximity to the plasma membrane (Fig.4A,E). Because this happens in the absence of TRP, this result suggests that membrane association does not depend on TRP–INAD interactions and perhaps would be expected to occur even in PDZ3 mutants. Indeed, when CHO cells are transfected with a PDZ3 mutant construct, they are nearly indistinguishable from wild-type controls (data not shown). These cells also show complex formation. eye-PKC exhibits punctate, perinuclear localization (Fig. 4D). However, when wild-type INAD and eye-PKC are coexpressed, eye-PKC now colocalizes with INAD to the periphery (Fig. 4E) and coimmunoprecipitates with INAD (data not shown). We next examined the localization of all five different PDZ mutants. In contrast to wild-type INAD and mutations in PDZ-2, PDZ-3, PDZ-4, and PDZ-5 domains (Fig. 4C,G), the PDZ1 mutant protein does not localize to the periphery. Instead, it is found expressed diffusely throughout the cell (Fig.4B,F). To ensure that INADPDZ1 is a functional protein, we examined its interaction with eye-PKC. Figure 4Fconfirms that PDZ1 still can interact with and redistribute eye-PKC. These results indicate that PDZ1 is essential for membrane localization and suggest that INAD is anchored by interactions between PDZ3 and TRP and between PDZ1 and a membrane target.
Fig. 4.

PDZ1 is required for localizing INAD at the membrane of CHO cells. A–C, Wild-type and mutantinaD constructs containing point mutations (see Materials and Methods) in PDZ1, PDZ2, PDZ3, PDZ4, or PDZ5 were transfected into CHO cells. Shown are confocal images of immunofluorescently stained CHO cells transfected with wild-typeinaD (INAD; A),inaDPDZ1 (PDZ1;B), and inaDPDZ2(PDZ2; C). Left, Anti-INAD staining. Right, Anti-INAD (green) superimposed with rhodamine-conjugated phalloidin staining (red). INAD shows membrane-associated staining, as seen in A and E, in 72.7% of transfected cells (n = 414), whereas PDZ1 shows membrane-associated staining in only 19.9% of transfected cells (n = 272). In most cells PDZ1 (B, F) is expressed diffusely throughout the cell. The percentage of transfected cells displaying membrane-associated localization was 63.8% for PDZ2 (n = 315), 74.8% for PDZ3 (n = 302), 42.4% for PDZ4 (n = 85), and 40.6% for PDZ5 (n = 256). D–G, Wild-type and mutant INADs redistribute eye-PKC when cotransfected. Shown is immunofluorescent staining of CHO cells cotransfected withinaD (INAD),inaDPDZ1 (PDZ1), orinaDPDZ2 (PDZ2) with eye-PKC. Left, Anti-INAD staining. Right, Anti-PKC staining. PKC transfected alone displays a punctate, perinuclear expression pattern (D). When cotransfected with inaD (E),inaDPDZ1(F), orinaDPDZ2 (G), PKC is redistributed into an expression pattern like that of INAD, PDZ1, or PDZ2, respectively.
Signaling complexes are assembled before they are targeted to the rhabdomeres
How are the “soluble” partners of INAD (eye-PKC and PLC) targeted to the rhabdomeres? To test whether eye-PKC and PLC could be targeted to the rhabdomeres in the absence of INAD, we examined their initial subcellular localization in wild-type, trp, andinaD animals from the earliest times that we could detect expression. Figure 5 shows that eye-PKC and PLC localize specifically to the rhabdomeres of wild-type andtrp mutant pupae but remain mainly in the cell bodies ofinaD mutant photoreceptors. These findings indicate that INAD is required for the targeting of PLC and eye-PKC and suggest that INAD may be preassembling signaling complexes before targeting to the rhabdomeres.
Fig. 5.

PKC and PLC require INAD to be targeted to the rhabdomeres. Shown is anti-PKC and anti-PLC immunofluorescent staining of cross sections (1 μm thick) oftrp343 andinaD1 pupal photoreceptors. Pupae were aged and sectioned at the earliest times of anti-PKC and anti-PLC detection. PKC and PLC are targeted to the rhabdomeres intrp mutants but were mislocalized at similar times ininaD mutants.
To determine whether INAD assembles its soluble partners into complexes before they are delivered to the rhabdomeres, we designed an experiment that would allow us to follow the fate of a pulse of newly synthesized INAD and its association with PLC and eye-PKC. We generated transgenic flies expressing INAD under the control of an inducible heat shock promoter (hs-inaD) and examined the formation of signaling complexes, rhabdomeric targeting, and electrophysiological recovery of light responses as a function of time after a pulse of INAD expression. Uninduced hs-inaDtransgenic flies are virtually identical to inaD null mutants: TRP, eye-PKC, and PLC are unstable (Fig.6) and mislocalized (data not shown), and light responses have severe defects with very poor amplitudes and kinetics of activation and deactivation (Fig.7A; Tsunoda et al., 1997). A 2 hr heat shock at 37°C induces a pulse of INAD expression, which is followed by the restabilization of TRP, eye-PKC, and PLC (see Fig. 6). By 7 hr after heat shock INAD has reached the rhabdomeres, and ERG recordings begin to resemble wild-type responses (Fig.7A).
Fig. 6.

Induction of INAD expression in vivo. Shown is immunoblot analysis of transduction proteins in wild-type (wt) and hs-inaD transformant flies. hs-inaD flies without heat shock (no hs) did not express INAD, and levels of TRP, PLC, and eye-PKC were reduced dramatically (Tsunoda et al., 1997).hs-inaD flies were given a 2 hr heat shock (hs) at 37°C, and levels of transduction proteins were assayed at 12 hr intervals after heat shock (hs + 0, hs + 12 hr, etc.). Induction of INAD expression can be seen immediately after heat shock; this pulse of INAD protein expression was followed by a rise in the levels of TRP, eye-PKC, and PLC.
Fig. 7.

INAD assembles PKC and PLC into complexes before they are targeted to the rhabdomeres.A, Top Left, Electroretinogram (ERG) recordings from wild-type and hs-inaD flies.hs-inaD flies either were uninduced (no hs) or were heat-shocked for 2 hr at 37°C and then recorded at 3 hr (+3hr) or 7 hr (+7hr) after heat shock. The stimulus was a 5 sec pulse of orange light (570 nm long-pass filter). Note the rescue of signaling at 7 hr after heat shock.Top Right to Bottom, Electron microscopic immunogold localization of INAD in hs-inaD rhabdomeres. At 3 hr after heat shock (+3hr) INAD protein had not yet arrived in the rhabdomeres, whereas INAD was present at 7 hr after heat shock (+7hr). Particles per square micrometer that were counted included the following: no heat shock, 1.29 ± 1.90 (n = 28 rhabdomeres); 3 hr, 1.86 ± 2.30 (n = 28); 7 hr, 19.2 ± 5.29 (n = 40). B, Membrane extracts from heads of wild-type (wt), hs-inaD, andinaD1 flies were immunoprecipitated (50 heads for wt and inaD1; 500 heads for hs-inaD) by using an anti-INAD antibody.hs-inaD flies either were uninduced (no hs) or were heat-shocked for 2 hr at 37°C and then assayed at 3 hr after heat shock (+3hr). Immunoprecipitated proteins were separated by SDS-PAGE, transferred to nitrocellulose, and probed with antibodies specific for INAD, PLC, eye-PKC, and rhodopsin (Rh1). Fly mb refers to wild-type membranes before immunoprecipitation. As expected, PLC and eye-PKC coimmunoprecipitated with INAD in wild-type membranes but did not precipitate from inaD null or wild-type membranes incubated without antibody (wt, no Ab). Fromhs-inaD flies, PKC and PLC coimmunoprecipitated with anti-INAD antibodies 3 hr after heat shock, a time when signaling still is not restored and INAD has not yet reached the rhabdomeres (A); it should be noted that it is possible that immunoEM and immunoprecipitation may have different sensitivities. Rhodopsin, which is not a part of the signaling complex, did not coimmunoprecipitate with INAD in any genotype.
To assay for preassembly, we immunoprecipitated INAD from heads of heat-shocked hs-inaD flies at 3 hr after heat shock, a time before INAD has reached the rhabdomeres and long before light responses have recovered (Fig. 7A). Figure 7B shows that INAD directly interacts with its soluble partners, eye-PKC and PLC, and that these complexes are preassembled. Control immunoprecipitations from uninduced flies, or inaD null mutants, showed only background levels of eye-PKC and PLC. As expected, rhodopsin is not found in the complex, although it is >1000-fold more abundant than any of these proteins. These results show that INAD preassembles eye-PKC and PLC into complexes and support a model in which signaling complexes, once assembled, are targeted as a whole to the rhabdomeres.
DISCUSSION
The strategic placement of the proper signaling machinery at the appropriate subcellular location is crucial for normal signaling and cellular function. For example, in neurons, ion channels and receptors are not distributed randomly throughout the cell membrane but are clustered at relevant subcellular sites: Na+ channels at nodes of Ranvier, K+ and Ca+2channels at presynaptic terminals, and acetylcholine and glutamate receptors at postsynaptic sites. Similarly, the appropriate localization of receptors to the apical or basolateral membranes of epithelial cells is necessary for receiving and responding to various environmental cues.
In Drosophila photoreceptors, high sensitivity to light is achieved by having an extremely large number of light receptor molecules (∼100,000,000 rhodopsins). All of these rhodopsin molecules are housed in a specialized subcellular compartment called the rhabdomere. Each rhabdomere consists of ∼60,000 microvilli, which provide the huge membrane surface needed to accommodate the large number of receptor molecules and signaling components. INAD acts as a scaffold protein, organizing signaling components downstream of rhodopsin (TRP, PLC, eye-PKC; see introductory remarks) into discreet signaling complexes or transducisomes. Previously, we showed that INAD is essential for the rhabdomeric localization and organization of signaling complexes (Tsunoda et al., 1997). In this study we used INAD transducisomes as a model for examining how signaling complexes are assembled and anchored.
Anchoring of INAD signaling complexes
INAD, like many PDZ–scaffold proteins, is a soluble protein, yet it functions as an organizer of membrane-associated complexes. Thus, we wondered how INAD is anchored to the membrane. Because TRP is a transmembrane protein, we investigated whether TRP anchors INAD. Results from this study show that the interaction of INAD with the TRP channel indeed is required for anchoring signaling complexes in the rhabdomeres. TRP then may interact with the cytoskeleton, securing INAD and the whole complex to the membrane; ankyrin repeats on the N terminus of TRP could play a role in linking TRP to the cytoskeleton. Another possibility is that the INAD–TRP interaction reveals, or unmasks, sites on TRP or INAD that are important for membrane anchoring.
ImmunoEM studies showed, however, that ∼25% of INAD remains in the rhabdomeres of trp null mutants, suggesting that TRP is not the only anchor of INAD. We have shown that PDZ1 of INAD may play a role in anchoring INAD to the membrane. PDZ1 has been proposed to bind an unconventional myosin III, encoded by the ninaC gene, that could link INAD to the actin cytoskeleton (Wes et al., 1999). INAD, however, is localized normally in ninaCmutants (Wes et al., 1999), suggesting that it is unlikely that NINAC anchors INAD. Possibly, PDZ1 interacts with a yet unidentified target protein that interacts with the cytoskeleton and anchors INAD. Notably, PDZ domains have been shown to bind directly to cytoskeletal-associated elements, such as α-actinin (Mandai et al., 1999), actin (Xia et al., 1997), protein 4.1 (Marfatia et al., 1997), neuroligins (Irie et al., 1997), and dystrophin (Brenman et al., 1996).
Assembly of INAD signaling complexes
How are signaling components initially targeted to the rhabdomeres? Where are signaling complexes assembled? We envision two potential scenarios: (1) signaling components are targeted independently to the rhabdomeres where they are assembled into complexes, and (2) signaling complexes are “preassembled” and subsequently targeted as a whole to the rhabdomeres. In this report, we present evidence supporting the latter strategy. Preassembly of INAD signaling complexes offers the advantage of minimizing the number of stray signaling components in the rhabdomeres while maximizing the number of complete, functional complexes. It is also worth noting that, in the absence of INAD, its targets TRP, PLC, and eye-PKC are very unstable and nearly undetectable (see Fig. 6 and Tsunoda et al., 1997). Interestingly, a pulse of INAD expression leads to their restabilization, likely via their assembly into transduction complexes. It will be of interest to determine whether this represents a regulatory strategy to ensure that “loose” signaling molecules do not wander about the cell.
Preassembly of macromolecular complexes has been documented for KATP channels (Zerangue et al., 1999) and mammalian T-cell receptor complexes (Klausner et al., 1990), as well as in Chlamydomonas and Paramecium, in which the outer dynein arms of the flagellum are preassembled in the cytoplasm before they are transported to the tip of the growing flagellum (Fowkes and Mitchell, 1998; Rosenbaum et al., 1999). Preassembly then may be a mechanism common to many cells for targeting different signaling machinery or macromolecular structures to their pertinent subcellular domains. Preassembly and the regulation of complex assembly/targeting may be particularly important in the nervous system where a single neuron must target distinct ion channel or receptor complexes, as well as the relevant regulatory machinery, to hundreds or thousands of different pre- and postsynaptic sites.
It will be important to understand where the complexes are assembled and how this assembly is performed. Strategies for enforcing preassembly may be similar to some immune receptors and KATP channel subunits that have been shown to contain endoplasmic reticulum (ER) retention signals that are concealed when subunits are assembled, allowing whole receptor or channel complexes to be targeted to the membrane, whereas individual subunits are degraded or retained in the ER (Bonifacino et al., 1990a,b;Zerangue et al., 1999). Although we have shown that INAD is required for targeting complexes to the rhabdomeres, virtually nothing is known about how targeting is accomplished. Genetic screens that track the localization of INAD may provide a fruitful means for identifying components involved in assembly and targeting. With the identification of proteins that play a role in these processes, we may begin to build a picture of how signaling complexes are assembled, how assembly is regulated, and how signaling complexes are targeted to the proper location.
Footnotes
This work was funded in part by a grant from the National Eye Institute to C.S.Z. S.T. is a fellow of the American Cancer Society and C.S.Z. is an investigator of the Howard Hughes Medical Institute. We thank Ann Becker for generating and purifying antibodies used in this study, Ruth Bodner for making the originaltrpC34 construct, and members of the Zuker lab for many helpful discussions.
Correspondence should be addressed to Dr. Susan Tsunoda, Howard Hughes Medical Institute and Departments of Biology and Neurosciences, University of California at San Diego, La Jolla, CA 92093-0649. E-mail:susan@flyeye.ucsd.edu.
REFERENCES
- 1.Adamski FM, Zhu M-Y, Bahiraei F, Shieh B-H. Interaction of eye protein kinase C and INAD in Drosophila. J Biol Chem. 1998;273:17713–17719. doi: 10.1074/jbc.273.28.17713. [DOI] [PubMed] [Google Scholar]
- 2.Arnold DB, Clapham DE. Molecular determinants for subcellular localization of PSD-95 with an interacting K+ channel. Neuron. 1999;23:149–157. doi: 10.1016/s0896-6273(00)80761-8. [DOI] [PubMed] [Google Scholar]
- 3.Bahner M, Sander P, Paulsen R, Huber A. The visual G-protein of fly photoreceptors interacts with the PDZ domain-assembled INAD signaling complex via direct binding of activated Gαq to phospholipase Cβ. J Biol Chem. 2000;275:2901–2904. doi: 10.1074/jbc.275.4.2901. [DOI] [PubMed] [Google Scholar]
- 4.Baker E, Colley N, Zuker C. The cyclophilin homolog ninaA functions as a chaperone forming a stable complex in vivo, with its protein target, rhodopsin. EMBO J. 1994;13:4886–4895. doi: 10.1002/j.1460-2075.1994.tb06816.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bhat MA, Izaddoost S, Lu Y, Cho K-O, Choi K-W, Bellen HJ. Discs lost, a novel multi-PDZ domain protein, establishes and maintains epithelial polarity. Cell. 1999;96:833–845. doi: 10.1016/s0092-8674(00)80593-0. [DOI] [PubMed] [Google Scholar]
- 6.Bonifacino JS, Cosson P, Klausner RD. Colocalized transmembrane determinants for ER degradation and subunit assembly explain the intracellular fate of TCR chains. Cell. 1990a;63:503–513. doi: 10.1016/0092-8674(90)90447-m. [DOI] [PubMed] [Google Scholar]
- 7.Bonifacino JS, Suzuki CK, Klausner RD. A peptide sequence confers retention and rapid degradation in the endoplasmic reticulum. Science. 1990b;247:79–82. doi: 10.1126/science.2294595. [DOI] [PubMed] [Google Scholar]
- 8.Brenman JE, Chao DS, Gee SH, McGee AW, Craven SE, Santillano DR, Wu Z, Huang F, Xia H, Peters MF, Froehner SC, Bredt DS. Interaction of nitric oxide synthase with the postsynaptic density protein PSD-95 and α1-syntrophin mediated by PDZ domains. Cell. 1996;84:757–767. doi: 10.1016/s0092-8674(00)81053-3. [DOI] [PubMed] [Google Scholar]
- 9.Chevesich J, Kreuz AJ, Montell C. Requirement for the PDZ domain protein, INAD, for localization of the TRP store-operated channel to a signaling complex. Neuron. 1997;18:95–105. doi: 10.1016/s0896-6273(01)80049-0. [DOI] [PubMed] [Google Scholar]
- 10.Craven SE, Bredt DS. PDZ proteins organize synaptic signaling pathways. Cell. 1998;93:495–498. doi: 10.1016/s0092-8674(00)81179-4. [DOI] [PubMed] [Google Scholar]
- 11.Dong H, O'Brien RJ, Fung ET, Lanahan AA, Worley PF, Huganir RL. GRIP: a synaptic PDZ domain-containing protein that interacts with AMPA receptors. Nature. 1997;386:279–284. doi: 10.1038/386279a0. [DOI] [PubMed] [Google Scholar]
- 12.Doyle DA, Lee A, Lewis J, Kim E, Sheng M, MacKinnon R. Crystal structures of a complexed and peptide-free membrane protein-binding domain: molecular basis of peptide recognition by PDZ. Cell. 1996;85:1067–1076. doi: 10.1016/s0092-8674(00)81307-0. [DOI] [PubMed] [Google Scholar]
- 13.Fowkes ME, Mitchell DR. The role of preassembled cytoplasmic complexes in assembly of flagellar dynein subunits. Mol Biol Cell. 1998;9:2337–2347. doi: 10.1091/mbc.9.9.2337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hardie R. Whole-cell recordings of the light-induced current in dissociated Drosophila photoreceptors: evidence for feedback by calcium permeating the light-sensitive channels. Proc R Soc Lond [Biol] 1991;245:203–210. [Google Scholar]
- 15.Hardie RC, Minke B. The trp gene is essential for a light-activated Ca2+ channel in Drosophila photoreceptors. Neuron. 1992;8:643–651. doi: 10.1016/0896-6273(92)90086-s. [DOI] [PubMed] [Google Scholar]
- 16.Hardie RC, Peretz A, Suss-Toby E, Rom GA, Bishop SA, Selinger Z, Minke B. Protein kinase C is required for light adaptation in Drosophila photoreceptors. Nature. 1993;363:634–637. doi: 10.1038/363634a0. [DOI] [PubMed] [Google Scholar]
- 17.Hay BA, Wolff T, Rubin GM. Expression of baculovirus P35 prevents cell death in Drosophila. Development. 1994;120:2121–2129. doi: 10.1242/dev.120.8.2121. [DOI] [PubMed] [Google Scholar]
- 18.Hoskins R, Hajnal AF, Harp SA, Kim SK. The C. elegans vulval induction gene lin-2 encodes a member of the MAGUK family of cell junction proteins. Development. 1996;122:97–111. doi: 10.1242/dev.122.1.97. [DOI] [PubMed] [Google Scholar]
- 19.Huber A, Sander P, Gobert A, Bahner M, Hermann R, Paulsen R. The transient receptor potential protein (Trp), a putative store-operated Ca2+ channel essential for phosphoinositide-mediated photoreception, forms a signaling complex with NorpA, InaC, and InaD. EMBO J. 1996;15:7036–7045. [PMC free article] [PubMed] [Google Scholar]
- 20.Huber A, Sander P, Bahner M, Paulsen R. The TRP Ca2+ channel assembled in a signaling complex by the PDZ protein INAD is phosphorylated through the interaction with protein kinase C (ePKC). FEBS Lett. 1998;425:317–322. doi: 10.1016/s0014-5793(98)00248-8. [DOI] [PubMed] [Google Scholar]
- 21.Irie M, Hata Y, Takeuchi M, Ichtchenko K, Toyoda A, Hirao K, Takai Y, Rosahl TW, Sudof TC. Binding of neuroligins to PSD-95. Science. 1997;277:1511–1514. doi: 10.1126/science.277.5331.1511. [DOI] [PubMed] [Google Scholar]
- 22.Kaech SM, Whitfield CW, Kim SK. The LIN-2/LIN-7/LIN-10 complex mediates basolateral membrane localization of the C. elegans EGF receptor LET-23 in vulval epithelial cells. Cell. 1998;94:761–771. doi: 10.1016/s0092-8674(00)81735-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kim E, Niethhammer M, Rothschild A, Jan YN, Sheng M. Clustering of Shaker-type K+ channels by interaction with a family of membrane-associated guanylate kinases. Nature. 1995;378:85–88. doi: 10.1038/378085a0. [DOI] [PubMed] [Google Scholar]
- 24.Kim E, Cho K-O, Rothschild A, Sheng M. Heteromultimerization and NMDA receptor clustering activity of Chapsyn-110, a member of the PSD-95 family of proteins. Neuron. 1996;17:103–113. doi: 10.1016/s0896-6273(00)80284-6. [DOI] [PubMed] [Google Scholar]
- 25.Klausner RD, Lippincott-Schwartz J, Bonifacino JS. The T-cell antigen receptor: insights into organelle biology. Annu Rev Cell Biol. 1990;6:403–431. doi: 10.1146/annurev.cb.06.110190.002155. [DOI] [PubMed] [Google Scholar]
- 26.Kornau HC, Seeburg PH, Kennedy MB. Interaction of ion channels and receptors with PDZ domain proteins. Curr Opin Neurobiol. 1997;7:368–373. doi: 10.1016/s0959-4388(97)80064-5. [DOI] [PubMed] [Google Scholar]
- 27.Mandai K, Nakanishi H, Satoh A, Obaishi H, Wada M, Nishioka H, Itoh M, Mizoguchi A, Aoki T, Fujimoto T, Matsuda Y, Tsukita S, Takai Y. Afadin: a novel actin filament-binding protein with one PDZ domain localized at cadherin-based cell-to-cell adherens junction. J Cell Biol. 1999;139:517–528. doi: 10.1083/jcb.139.2.517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Marfatia SM, Morais-Cabral JH, Kim AC, Byron O, Chishti AH. The PDZ domain of human erythrocyte p55 mediates its binding to the cytoplasmic carboxyl terminus of glycophorin C. J Biol Chem. 1997;272:24191–24197. doi: 10.1074/jbc.272.39.24191. [DOI] [PubMed] [Google Scholar]
- 29.McLean IW, Nakane PK. Periodate–lysine–paraformaldehyde fixative. J Histochem Cytochem. 1974;22:1077–1083. doi: 10.1177/22.12.1077. [DOI] [PubMed] [Google Scholar]
- 30.Niemeyer BA, Suzuki E, Scott K, Jalink K, Zuker CS. The Drosophila light-activated conductance is composed of the two channels, TRP and TRPL. Cell. 1996;85:651–659. doi: 10.1016/s0092-8674(00)81232-5. [DOI] [PubMed] [Google Scholar]
- 31.O'Brien RJ, Lau LF, Huganir RL. Molecular mechanisms of glutamate receptor clustering at excitatory synapses. Curr Opin Neurobiol. 1998;8:364–369. doi: 10.1016/s0959-4388(98)80062-7. [DOI] [PubMed] [Google Scholar]
- 32.Pawson T, Scott JT. Signaling through scaffold, anchoring, and adaptor proteins. Science. 1997;278:2075–2080. doi: 10.1126/science.278.5346.2075. [DOI] [PubMed] [Google Scholar]
- 33.Ranganathan R, Harris GL, Stevens CF, Zuker CS. A Drosophila mutant defective in extracellular calcium-dependent photoreceptor inactivation and rapid desensitization. Nature. 1991;354:230–232. doi: 10.1038/354230a0. [DOI] [PubMed] [Google Scholar]
- 34.Ranganathan R, Malicki DM, Zuker CS. Signal transduction in Drosophila photoreceptors. Annu Rev Neurosci. 1995;18:283–317. doi: 10.1146/annurev.ne.18.030195.001435. [DOI] [PubMed] [Google Scholar]
- 35.Reynolds ES. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J Cell Biol. 1963;17:208–212. doi: 10.1083/jcb.17.1.208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rosenbaum JL, Cole DG, Diener DR. Intraflagellar transport: the eyes have it. J Cell Biol. 1999;144:385–388. doi: 10.1083/jcb.144.3.385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schober M, Schaefer M, Knoblich JA. Bazooka recruits Inscuteable to orient asymmetric cell divisions in Drosophila neuroblasts. Nature. 1999;402:548–551. doi: 10.1038/990135. [DOI] [PubMed] [Google Scholar]
- 38.Scott K, Zuker C. Lights out: deactivation of the phototransduction cascade. Trends Biochem Sci. 1997;22:350–354. doi: 10.1016/s0968-0004(97)01100-6. [DOI] [PubMed] [Google Scholar]
- 39.Scott K, Leslie A, Sun Y, Hardy R, Zuker C. Gαq protein function in vivo: genetic dissection of its role in photoreceptor cell physiology. Neuron. 1995;15:919–927. doi: 10.1016/0896-6273(95)90182-5. [DOI] [PubMed] [Google Scholar]
- 40.Scott K, Sun YM, Beckingham K, Zuker CS. Calmodulin regulation of light-activated channels and receptor function mediates termination of the light response in vivo. Cell. 1997;91:375–383. doi: 10.1016/s0092-8674(00)80421-3. [DOI] [PubMed] [Google Scholar]
- 41.Sheng M. PDZs and receptor/channel clustering: rounding up the latest suspects. Neuron. 1996;17:575–578. doi: 10.1016/s0896-6273(00)80190-7. [DOI] [PubMed] [Google Scholar]
- 42.Shieh BH, Zhu MY. Regulation of the TRP Ca2+ channel by INAD in Drosophila photoreceptors. Neuron. 1996;16:991–998. doi: 10.1016/s0896-6273(00)80122-1. [DOI] [PubMed] [Google Scholar]
- 43.Shieh BH, Zhu MY, Lee JK, Kelly IM, Bahiraei F. Association of INAD with NORPA is essential for controlled activation and deactivation of Drosophila phototransduction in vivo. Proc Natl Acad Sci USA. 1997;11:12682–12687. doi: 10.1073/pnas.94.23.12682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Simske JS, Kaech SM, Harp SA, Kim SK. LET-23 receptor localization by the cell junction protein LIN-7 during C. elegans vulval induction. Cell. 1996;85:195–204. doi: 10.1016/s0092-8674(00)81096-x. [DOI] [PubMed] [Google Scholar]
- 45.Smith DP, Ranganathan R, Hardy RW, Marx J, Tsuchida T, Zuker CS. Photoreceptor deactivation and retinal degeneration mediated by a photoreceptor-specific protein kinase C. Science. 1991;254:1478–1484. doi: 10.1126/science.1962207. [DOI] [PubMed] [Google Scholar]
- 46.Tejedor FJ, Bokhari A, Rogero O, Gorczyca M, Zhang J, Kim E, Sheng M, Budnik V. Essential role for dlg in synaptic clustering of Shaker K+ channels in vivo. J Neurosci. 1997;17:152–159. doi: 10.1523/JNEUROSCI.17-01-00152.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Torres R, Firestein BL, Dong H, Staudinger J, Olson EN, Huganir RL, Bredt DSG, NW, Yancopoulos GD. PDZ proteins bind, cluster, and synaptically colocalize with Eph receptors and their ephrin ligands. Neuron. 1998;21:1453–1463. doi: 10.1016/s0896-6273(00)80663-7. [DOI] [PubMed] [Google Scholar]
- 48.Tsunoda S, Sierralta J, Sun Y, Bodner R, Suzuki E, Becker A, Socolich M, Zuker CS. A multivalent PDZ-domain protein assembles signaling complexes in a G-protein-coupled cascade. Nature. 1997;388:243–249. doi: 10.1038/40805. [DOI] [PubMed] [Google Scholar]
- 49.Tsunoda S, Sierralta J, Zuker CS. Specificity in signaling pathways: assembly into multimolecular signaling complexes. Curr Opin Genet Dev. 1998;8:419–422. doi: 10.1016/s0959-437x(98)80112-3. [DOI] [PubMed] [Google Scholar]
- 50.van Huizen R, Miller K, Chen D-M, Li Y, Lai Z-C, Raab RW, Stark WS, Shortridge RD, Li M. Two distantly positioned PDZ domains mediate multivalent INAD–phospholipase C interactions essential for G-protein-coupled signaling. EMBO J. 1998;17:2285–2297. doi: 10.1093/emboj/17.8.2285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wes PD, Xu X-ZS, Li H-S, Chien F, Doberstein SK, Montell C. Termination of phototransduction requires binding of the NINAC myosin III and the PDZ protein INAD. Nat Neurosci. 1999;2:447–453. doi: 10.1038/8116. [DOI] [PubMed] [Google Scholar]
- 52.Xia H, Winokur ST, Kuo WL, Altherr MR, Bredt DS. Actinin-associated LIM protein: identification of a domain interaction between PDZ and spectrin-like repeat motifs. J Cell Biol. 1997;139:507–515. doi: 10.1083/jcb.139.2.507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zerangue N, Schwappach B, Jan YN, Jan LY. A new ER trafficking signal regulates the subunit stoichiometry of plasma membrane KATP channels. Neuron. 1999;22:537–548. doi: 10.1016/s0896-6273(00)80708-4. [DOI] [PubMed] [Google Scholar]
- 54.Zito K, Fetter RD, Goodman CS, Isacoff EY. Synaptic clustering of Fasiclin II and Shaker: essential targeting sequences and role of Dlg. Neuron. 1997;19:1007–1016. doi: 10.1016/s0896-6273(00)80393-1. [DOI] [PubMed] [Google Scholar]