

Abstract
Ocean warming associated with global climate change is already inducing geographic range shifts of marine species. Juvenile coral reef fishes transported into temperate latitudes (termed ‘vagrant’ fishes) can experience winter water temperatures below their normal thermal minimum. Such environmental extremes may increase energetic costs for such fishes, resulting in reduced performance, which may be the governing factor that limits the potential for poleward range expansion of such fishes. This study compared the juvenile physiological performance and behaviour of two congeneric tropical damselfishes which settle during austral summer months within temperate eastern Australia: Abudefduf vaigiensis have an extended southern range, and lower threshold survival temperature than the congeneric A. whitleyi. Physiological and behavioural performance parameters that may be affected by cooler temperature regimes at higher latitudes were measured in aquaria. Lower water temperature resulted in reduced growth rates, feeding rates, burst escape speed and metabolic rates of both species, with significantly reduced performance (up to six-fold reductions) for fishes reared at 18 °C relative to 22 °C and 26 °C. However, A. whitleyi exhibited lower growth rates than A. vaigiensis across all temperatures, and lower aerobic capacity at the lowest temperature (18 °C). This difference between species in growth and metabolic capacity suggests that the extended southern distribution and greater overwintering success of A. vaigiensis, in comparison to A. whitleyi is related to thermal performance parameters which are critical in maintaining individual health and survival. Our results support previous findings in the region that water temperature below 22 °C represents a critical physiological threshold for tropical Abudefduf species expatriating into temperate south-eastern Australia.
Subject terms: Animal migration, Marine biology
Introduction
The geographic ranges of species are determined by their ability to successfully disperse into environments and then withstand local conditions1,2. A species’ range can be controlled by a combination of both climatic (e.g. local temperature, levels of precipitation and extreme weather events3,4) and ecological factors (e.g. competition for suitable food and resources5). Near the periphery of a species’ range, conditions may become increasingly stressful, either due to reductions in suitable resources and/or reductions in physiological suitability6. For range restrictions related to thermal physiology, climate change threatens to alter the distribution of organisms globally by shifting them towards cooler climates in a phenomenon referred to as poleward range expansions or ‘range shift’7–9. Range shifts have already been documented for numerous populations of marine organisms10,11 and many of these shifts are occurring faster than the global average of all environments12.
Successful range shifts require the ability for species to disperse to new locations. The majority of marine organisms, including fishes, possess a dispersive life phase during larval development13,14. This allows local or regional currents to potentially facilitate poleward range expansion, including the expatriation of tropical fish species into temperate habitats15. For example, the East Australian Current (EAC) is a western boundary current that transports the larvae of tropical reef fish polewards to temperate environments of Australia’s south-east (SE) coastline, with juveniles of many coral-reef species already observed inhabiting temperate rocky reefs within austral summer months16,17. Range shift capacity of species is currently limited due to high mortality during the cooler winter months18. However, with increased ocean warming and strengthening of the EAC associated with climate change19, it is likely that the occurrence and persistence of expatriated tropical species in this region will increase in the future associated with higher water temperatures within winter months18.
Warming of SE Australian waters has already increased the abundance and diversity of new settlers of a range tropical fishes into temperate habitats, with 47 species recorded during surveys between 2003–200516 and an additional 22 species observed in a study conducted 10 years later17. Species from the damselfish genus Abudefduf have been one of the most commonly observed vagrants, in some locations being observed on 50% of surveys during recruitment months16. From January to May, 5 species of Abudefduf spp. vagrants settle into temperate rocky reef habitats along the New South Wales (NSW) coast16,20,21. The onset of winter results in almost 100% mortality among these tropical fishes when water temperature drops below 17 °C18, therefore over-winter survival appears to be the major limitation to the poleward range expansion of these tropical vagrants into temperate Australia. Previous research has shown that juvenile abundance of these tropical vagrants on temperate reefs varies greatly, not just between unrelated species but also between congenerics14,16. For example, A. vaigiensis, A. sexfaciatus and A. bengalensis are found more frequently, abundantly and further south in SE Australia than A. whitleyi, which has only been observed at a few locations sporadically and have rarely been recorded at latitudes higher than 37°16,17. Over-winter survival also differs among Abudefduf species, with previous modelling indicating a warmer threshold survival temperature for vagrant A. whitleyi (16.5–16.8 °C) relative to both A. vaigiensis and A. bengalensis (12.8–13 °C), and no overwintering recorded for A. sexfasciatus in temperate SE Australia18. It is predicted by the year 2080 that the overwinter survival of tropical vagrants in this region will occur annually due to ocean warming above minimum thermal thresholds18; therefore, it is important to understand which species will most likely successfully recruit and sustain populations.
Both physiological and behavioural performance of marine fishes are linked to ambient thermal conditions22,23. Ectotherm metabolic theory predicts that temperatures close to critical thermal maxima or minima will negatively impact kinetic activity and destabilise heat-shock proteins24,25. When approaching the thermal limits, physiological constraints also occur through reduced aerobic capacity due to limitations of the circulatory and ventilatory systems, consequently reducing an individual’s aerobic scope (i.e. energy available above basic cellular requirements) and impacting all higher functions including growth, feeding, swimming and reproduction26,27. Several studies have already shown the adverse effects on the aerobic capacity of tropical fish after exposure to water temperatures approaching their upper thermal limits28,29, however no research has focused on how individual fitness is compromised at the lower end of their thermal window. Thermal-tolerance limits of individuals are likely to differ among fish species18. A recent laboratory investigation of A. vaigiensis showed that winter-water temperatures of less than 18 °C result in depressed feeding rates, growth and burst escape ability, potentially compromising this species ability to survive and persist in temperate habitats22. Such negative consequences for the feeding and growth of individuals may then prolong their time spent at smaller size classes and ability to escape predators30,31, thereby increasing their risk of predation. Understanding inter-specific differences in thermal performance and whether such temperature-mediated limits may also impact the range of tropical reef fish species dispersing in temperate Australia will be vital in understanding species-specific range shift potential and our ability to predict future range shifts.
This study investigated the thermal performance of two species of juvenile tropical damselfish (A. vaigiensis and A. whitleyi) that recruit into temperate SE Australian rocky reef habitat16,21. The southern distribution of A. whitleyi is not as extensive as A. vaigeinsis which may result from species-specific differences in thermal performance within this cooler water region. To understand the thermal performance of juvenile fishes, growth rate, feeding rate, burst escape behaviour and metabolic rate were determined across temperatures that reflect typical natal coral reef (26 °C) and SE Australia temperate rocky reefs in summer (22 °C) and winter (18 °C). We tested the hypothesis that these performance metrics would be reduced in A. whitleyi relative to A. vaigiensis at lower water temperatures.
Results
Feeding rates
In both species, feeding rates of juveniles increased with water temperature (Fig. 1; F(2,60) = 29.627, p < 0.001). Specifically, there was at least a seven fold increase for both species over the temperature range 18 to 26 °C. The greatest change in feeding rate for both species occurred from 18 to 22 °C, but feeding rate differed significantly between all temperature treatments (Tukey’s test for all treatment comparisons; p < 0.001). Feeding rates did not differ between species (F(1,60) = 0.006, p = 0.938).
Figure 1.
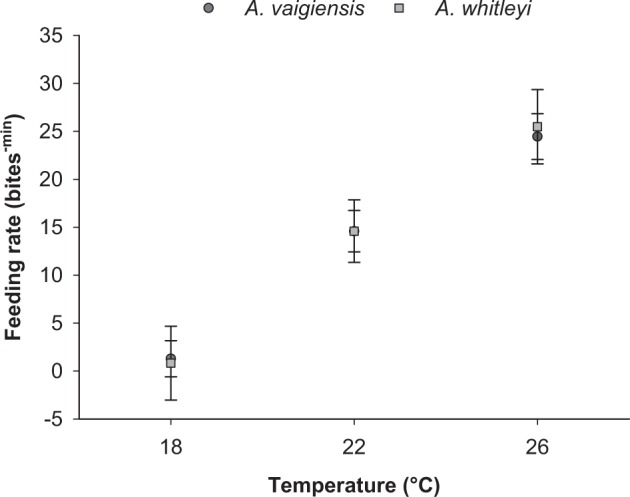
Feeding rate of A. vaigiensis (n: 18 °C = 21, 22 °C = 16, 26 °C = 13) and A. whitleyi (n: 18 °C = 5, 22 °C = 7, 26 °C = 5) maintained at three temperature treatments. Values are least square mean (Mean ± SE) adjusted for the covariate Weight (mean = 0.314 g).
Growth rates
Growth rate, calculated as a change in mass per day, was higher at warmer water temperatures of 22 and 26 °C for both species than at 18 °C (Fig. 2; Temperature: F(2,56) = 15.424, p < 0.001). Fish maintained at 18 °C lost 0.001 g (approximately 1% loss) per day over the testing period while fish at both 22 and 26 °C increased in weight by approximately 0.004 g (approximately 2.5% gain) per day (22 and 26 °C significantly different from 18 °C, Tukey’s tests, p < 0.05). In addition, A.vaigiensis exhibited higher mean growth overall of 0.003 g per day, compared to 0.001 g per day for A. whitleyi (F(1,56) = 4.738, p = 0.0345). While mean growth rate for A. vaigiensis appeared to be similar to A. whitleyi at 18 °C and higher at 22 and 26 °C, this apparent difference did not result in a significant interaction (Temperature * Species: F(1,56) 3.482, p = 0.67). A. whitleyi juveniles were generally smaller at collection, 19.22 mm SL (±0.36 mm SE) compared to 19.79 mm SL for A. vaigiensis (±0.09 mm SE).
Figure 2.
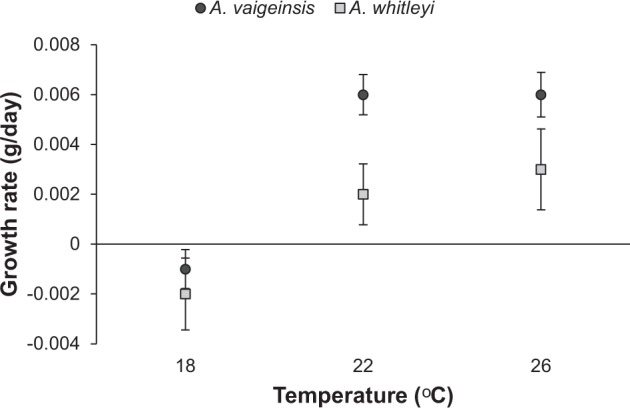
Mean (±SE) growth rate of A. vaigiensis (n: 18 °C = 17, 22 °C = 16, 26 °C = 13) and A. whitleyi (n: 18 °C = 5, 22 °C = 7, 26 °C = 3) maintained at three temperature treatments. Values are least squares mean (Mean ± SE) adjusted for the covariate Weight (mean = 0.32 g).
Burst swim speed
Burst swim speed was greater at higher water temperature for both species (Fig. 3; F(2,60) = 3.349, p = 0.042). Juvenile A. whitleyi exhibited higher burst swim speeds at 22 °C (43.59 bl/s) than 18 °C (27.4) bl/s), but no further increase was seen from 22 to 26 °C (Fig. 3). The significant differences found among temperatures was driven by the 18 °C treatment for both species, which was significantly different from both 22 and 26 °C treatments (Tukey’s tests; 18–22 °C, p = 0.024; 18–26 °C, p = 0.038), while 22 and 26 °C treatments did not differ from each other (Tukey’s test; 22–26 °C, p = 0.935). Burst speed did not differ between the two species, exhibiting similar trends across the 18 to 26 °C temperature range tested (F(1,60) = 0.002, p = 0.966; Temperature * Species: F(2,60) = 2.296, p = 0.745).
Figure 3.
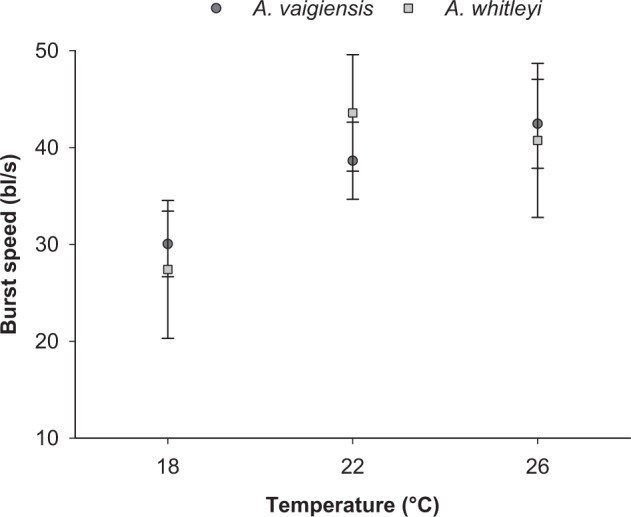
Mean (±SE) burst swimming speed of A. vaigiensis (n: 18 °C = 22, 22 °C = 16, 26 °C = 12) and A. whitelyi (n: 18 °C = 5, 22 °C = 7, 26 °C = 4) maintained at three temperature treatments.
Aerobic performance
All measured aerobic metabolic attributes increased with water temperature for both species (Fig. 4; MO2ROUTINE: F(2,49) = 19.35, p < 0.001; MO2MAX: F(2,48) = 60.101, p < 0.001; Net Scope: F(2,46) = 27.579, p < 0.001). MO2ROUTINE increased approximately 0.014 mg/hr per degree of warming, with each temperature treatment found to be significantly different (Fig. 4A; Tukey’s tests, all p < 0.01). MO2MAX increased at a higher rate to MO2ROUTINE of 0.043 mg/hr per degree Celsius and again all temperature treatments were significantly different within a species (Fig. 4B; Tukey’s tests, all p < 0.05). For both MO2ROUTINE and MO2MAX no significant differences were found between species (MO2ROUTINE: F(2,49) = 0.272, p > 0.05, MO2MAX: F(2,48) = 0.038, p > 0.05), nor the interaction between species and temperature (MO2ROUTINE: F(2,49) = 0.384, p > 0.05, MO2MAX: F(2,48) = 2.088, p > 0.05). However, a significant interaction between species and temperature was observed for aerobic scope (F(2,46) = 27.579, p < 0.001). This was driven by higher aerobic scope at 18 °C and lower at 26 °C for A. vaigiensis relative to A. whitleyi (Fig. 4C).
Figure 4.
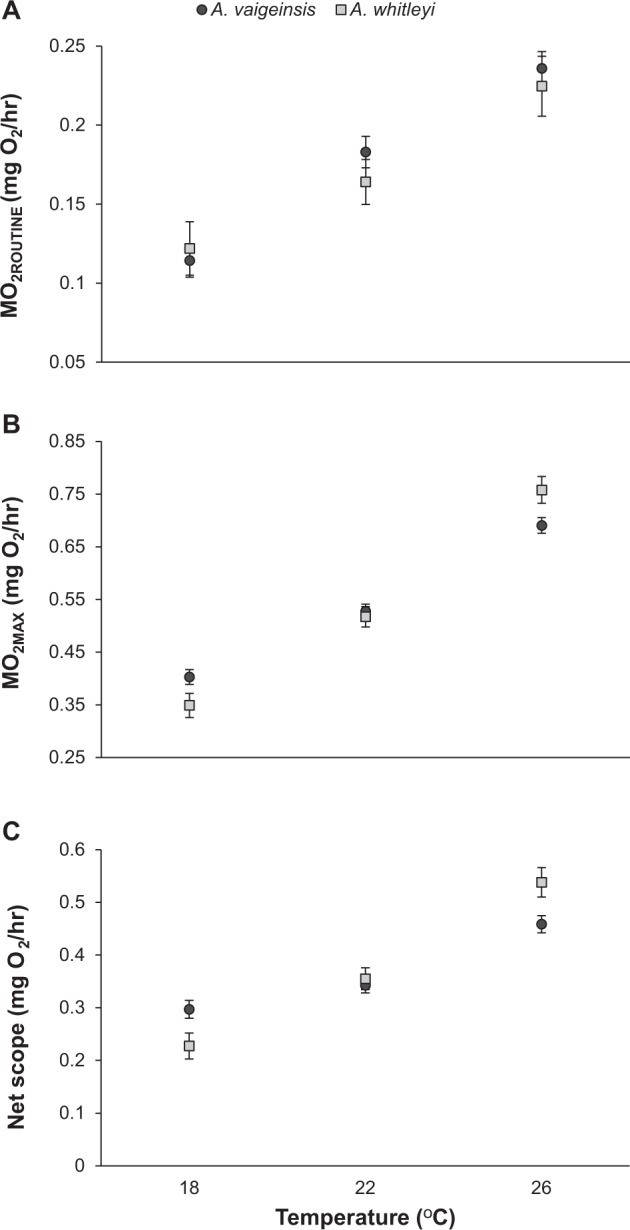
Mean (±SE) (A) MO2ROUTINE, (B) MO2MAX and (C) net aerobic scope of A. vaigiensis (MO2ROUTINE − n 18 °C = 14, 22 °C = 14, 26 °C = 13; MO2MAX − n 18 °C = 16, 22 °C = 15, 26 °C = 15; net aerobic scope - n 18 °C = 13, 22 °C = 12, 26 °C = 12) and A. whitleyi (MO2ROUTINE, MO2MAX and net aerobic scope – n: 18 °C = 5, 22 °C = 7, 26 °C = 4) maintained at three temperature treatments. Values are least square means adjusted for the covariate Weight (mean = 0.38 g).
Discussion
This study shows that reduced water temperatures approaching critical thermal minima impact key performance attributes of the juvenile stages of two vagrant damselfishes, with performance loss observed below 22 °C. Reductions in feeding, growth, burst-swimming performance and aerobic capacity were observed at 18 °C, compared to 22 °C and 26 °C, for both A. vaigiensis and A. whitleyi. Some aspects of performance of A. whitleyi were also reduced relative to A. vaigiensis. Consistent with our hypothesis of reduced performance of A. whitleyi relative to A. vaigiensis at low temperatures, the reduction in aerobic scope for A. whitleyi at 18 °C indicates greater physiological sensitivity at this low temperature compared to A. vaigiensis. A. vaigiensis also exhibited faster growth than A. whitleyi at all temperatures, which may indirectly enhance survival by reducing time spent susceptible to gape-limited predators30. Contrary to our hypothesis, however, feeding rate and burst speed (predator escape capacity) were similar between the species. While patterns of feeding rate across temperatures appeared similar between species, with small errors observed around the mean values, the lack of difference in burst speed may have resulted from low numbers of A. whitleyi. This species is uncommon in the study region, so sample sizes were relatively low, potentially reducing the power of the experiment to detect the apparent reduction in burst speed for A. whitleyi relative to A. vaigiensis at lowest temperature (18 °C). However, the significant differences found among temperature treatments suggest that power was significant enough to detect differences at the 0.05 level. With the exception of aerobic scope, the minimal differences found between the species indicate the overall similarities of their physiological and behavioural responses. Regardless, this research compliments our previous understanding of thermal performance and sensitivity of vagrant tropical fish, and emphasises that ocean temperature may need to warm above the performance threshold of 18 °C, rather than the critical thermal minimum, to allow range shifts.
As temperatures approach the lower thermal threshold, the maximal capacity for oxygen delivery declines faster than the basic oxygen requirements, thus limiting aerobic capacity25,27,32. These thermal minima temperatures also induce the destabilisation of proteins and enzymes that are essential for performance25. Aerobic scope, and the associated MO2ROUTINE and MO2MAX, showed high sensitivity to temperature change with significant differences between all testing temperatures. Interestingly, we found that A. vaigiensis maintained a slightly higher aerobic scope than A. whitleyi at 18 °C; conversely A. whitleyi exhibited slightly higher aerobic scope at 26 °C. Despite only small differences between the species’ aerobic scope, minor changes to aerobic capacity can impact individuals and ultimately populations through energetic trade-offs in behavioural and physical performance27. However, the observed differences between species are small in comparison to the overall reductions observed in relation to water temperature. Over the 8 °C temperature range tested aerobic scope was reduced by approximately 25%. This reduction in aerobic scope is expected to correlate to a reduction in energy available for higher level functions including growth27,32, and may indirectly explain why A. whitleyi distribution is limited at lower latitudes in NSW16. We did observe some differences in growth that matched trends in reduced aerobic capacity from 22 to 18 °C. However, the lack of corresponding differences in growth between 26 and 22 °C, which saw substantial reductions in aerobic capacity, suggests that while we may simplistically expect a direct relationship between energetic costs, energy intake and linear growth there are numerous other aspects of morphological development that we have not measured here that would be energy consumptive.
Similar trends with temperature were observed for burst escape performance, feeding rate and growth in both species, with the largest changes observed between 18–22 °C, rather than 22–26 °C. The broad range of traits affected potentially indicates that a number of critical physiological processes are impacted at water temperatures less than 22 °C. This result is not unexpected, since the average minimum winter water temperature on natal corals reefs in the southern GBR is 21 °C22, and these species may not have evolved to tolerate cooler waters. The observed reductions in performance at 18 °C matches with field observations of seasonal mortality during winter (June-August) in NSW coastal waters, which on average is between 15–17 °C18,22. In combination, these results suggest SE Australia waters would need to warm above 18 °C before range shifts may be thermally possible, and likely higher for substantial overwintering and establishment of populations.
While we observed differences in burst speed among water temperatures, it was the least sensitive performance metric to changes in water temperature. This is probably due to the fact that at high velocity swimming, white anaerobic muscles provide the power needed for burst speed33, whereas the other performance traits measured were powered by the aerobic systems. While burst speed was not as sensitive to temperature change, even small differences in response speed could have severe impacts on survival34. Although no significant differences were detected between species in the current study, at 18 °C A. vaigiensis juveniles swam on average 7 body lengths greater than A. whitelyi juveniles, which, if confirmed as a difference in future studies employing greater replication, may have implications for survival and population establishment in temperate habitats. Even a slight species-specific difference in escape distance may be the determinant of predator avoidance and ultimately survival. There are a number of other components of the burst escape response that were not measured in the study, specifically directionality, C-start latency and formation and Mauthner neurons (M-cell) excitability31,35. Action-potential in one M-cell triggers intracellular responses that initiate a fast-body bend (C-start), or escape response, away from an aversive stimulus36. While not confirmed in the current study, relative burst escape performance warrants further investigation, as a potential driver of differences in range shift potential among vagrant fishes.
The similarities in thermal performance between A. vaigiensis and A. whitleyi for some traits measured indicate that temperature is not likely the sole driver of variation in juvenile range and overwintering likelihood between the two species. Instead, ecological and biophysical interactions with temperature and other environmental factors may govern distribution and overwinter survival21. For example, predation acts as a major driver of population size and is often size-dependent, with higher predation mortality in smaller-size classes37. It has been observed in previous research that in expatriated populations of vagrant damselfish, A. whitleyi are generally smaller that A. vaigiensis38, potentially resulting in more predation of A. whitleyi aggregations. Differences in survival between species may also result from Allee effects39, with the greater abundance of A. vaigiensis compared to A. whitleyi in situ16 potentially contributing to overwintering differences. If population fitness and growth are density-dependent, then the smaller aggregations of A. whitleyi will exhibit reduced growth and spend greater periods of time at vulnerable size classes, increasing the likelihood of predator-induced mortality21.
The current settlement patterns of tropical fish vagrants into temperate SE Australia are correlated with rises in sea surface temperatures, and results obtained in this study indicate that winter temperatures greater than 18 °C could facilitate range shifting of both A. vaigiensis and A. whitleyi. Our findings build on previous studies by highlighting that performance loss can be observed at 18 °C, prior to critical thermal minimum temperatures around 16.5–17.5 °C18. We have shown some support for the expectation that A. whitleyi would be more thermally sensitive to cooler temperatures than A. vaigiensis, based on a more restricted poleward distribution. However, similarities observed for some traits, along with known interactions between environmental and ecological limiters of population size, suggest that future persistence will have more complex mechanisms than thermal performance alone, including aspects unmeasured here such as predation and/or competition, Allee effects and differences in recruit supply16,21,22. Extrinsic factors such as temperature and strengthening poleward ocean currents will likely increase the influx and range shift potential of tropical species into temperate marine ecosystems, and intrinsic factors will likely determine which species will establish viable populations that may influence the ecological dynamics of these systems, including competition and predation15. Further investigation of the lower end (18–22 °C) of tropical vagrants’ thermal minimums is essential for predicting future range shifts as anthropogenic-induced climate change alters the physical variables of marine ecosystems.
Materials and Methods
Experimental design
Juvenile A. vaigiensis (n = 50; standard length (SL) range 13–35 mm) and A. whitleyi (n = 17; SL range 16–31 mm) were collected in several batches between the 27th of February and the 2nd of May, 2013, from rocky reef habitats at 3 locations along the Sydney coast (Collins 33°48′30″S, 151°17′25″E, Freshwater 33°46′54″S, 151°17′39″E, and North Narrabeen 33°42′08″S, 151°18′25″E headlands), using an anaesthetic (clove oil, ethanol and sea water mixture) and hand net40. For A. vaigiensis, batch 1 was caught on the 27th of February, batch on the 26th of March and batch 3 on the 9th of April. For A. whitleyi, batch 1 was caught on the 19th of March, batch 2 on the 26th of March and batch 3 on the 14th of April. Fish were transported live within 3 hours and randomly assigned to 1 of 3 temperature-controlled aquariums at the University of Technology Sydney. Individual fish were placed in opaque-walled 14 L aquaria that were maintained at the ambient ocean temperature from the collection location (~22 °C). Over the course of 5 days, 1 tank was maintained at 22 °C, whereas the others were either lowered or increased in temperature of approximately 1 °C per day depending on their intended treatment tank. Following a habituation period of 5 days, individual fish were assigned randomly into the 3 temperature treatments: 26 °C, 22 °C and 18 °C. Overall for A. vaigeinsis there were 22 assigned to 18 °C, 16 to22 °C and 13 to 26 °C; and for A. whitleyi 5 assigned to 18 °C, 7 to 22 °C and 5 to 26 °C. Treatments were chosen to mimic the different environmental conditions that juvenile Abudefduf species may experience during their life history. The 26 °C treatment mimicked the average early summer temperatures experienced on the Southern Great Barrier Reef (GBR), the most likely source of tropical vagrants in temperate SE Australia, and parallels the irregular maximum sea surface temperatures of SE Australia during the recruitment period35. The summer temperature in the coastal waters of Sydney is approximately 22 °C41, corresponding to arrival and settlement of both Abudefduf species. The 18 °C temperature treatment mirrored the average water temperature at the end of the recruiting season in Sydney (May-June), and approached the lower critical thermal limit of approximately 17 °C previously identified for Abudefduf species22. Temperature treatments took 5 days to reach the desired set points, averaging a change of 0.8 °C daily for the 18 and 26 °C treatments. Water temperature was maintained by 25 W aquarium heaters in each tank and remained at ±0.5 °C of the treatment for 23 days. The tanks maintained at 18 °C were controlled by the thermostatted air-conditioning system, but were still supplied with non-operative heaters, to mimic secondary treatment effects. Tank temperatures were taken daily, and ammonia and pH tests were performed within the first and last weeks of the treatment period. To provide shelter for all experimental animals each tank also contained a 10 cm section of PVC tubing (6 cm diameter).
All fish were fed ad-libitum with 7 Spectrum marine fish food pellets 3 times daily. Uneaten food and faeces were siphoned off daily (prior to first feeding), simultaneously removing approximately 1–2 L of water, which was replaced with clean water (15–20% of the water from each tank).
Feeding rates
To examine feeding rate under standard hunger conditions all fish were starved for 12 hours prior to being supplied with an excess amount of food (following16). The number of bites taken by fish for 60 s post-feeding was counted by observation. All feeding assessments were conducted during the second week of the experiment.
Growth
At the commencement of the experiment, once fish had reached acclimation temperature, all individuals had their standard length (SL) measured to the nearest 0.5 mm, total length (TL) measured to the nearest 0.5 mm (both measured with a mm ruler) and wet weight (WW) measured to the nearest 0.01 g. These were also measured at the completion of the experiment as described above. Somatic growth was assessed by calculating growth rate as a change in WW over the experimental period.
Burst swimming speed
Predation is the major cause of mortality in marine fish populations; therefore the burst swimming speed used for the escape response by a fish is critical for survival42. The burst response of fish was determined by video analysis during the second week, 5–7 days after commencing temperature treatments. Fish were transferred individually to a narrow opaque-walled glass aquarium (10 cm wide, 50 cm long 30 cm tall) that had a gridded background (0.5 cm grids) with water that was maintained within 0.5 °C of their treatment temperature. The sides and back of the tank were blacked out to prevent external behaviour disturbance, and a Panasonic LUMIX DMC-FT4 waterproof digital camera was mounted in front of the tank. The methodology followed the protocols used in22, where individuals were startled using a dropped weight within the tank. After a 5 minute acclimation period, fish were recorded on video for 1 minute to quantify premature startle response before the experiment started. After this minute elapsed, fish were startled by a magnetic weight, which was released in the far side of the tank, and all behaviours recorded for a further minute. Videos were recorded at 24 frames per second, which has been proven adequate in assessing burst swimming speed in similar species22,24. Videos were analysed using Tracker 4.80 Video Analysis and Modelling Tool programs, which allowed videos to be slowed down to 0.04 frames per second. The greatest distance moved between 2 adjacent frames (approximately 0.04 s elapsed time) after the weight was released was taken as the maximum burst swimming speed of a fish in that trial. This measurement was originally taken in mm/s and then converted to body lengths per second (bl/s) by dividing the mm/s travelled by the TL of the fish to account for the effect of body length on distance moved.
Aerobic performance
Metabolic attributes, estimated from oxygen consumption, of A. vaigiensis and A. whitleyi were measured with static daytime respirometry 19 days after commencing temperature treatments, between 08:00–12:0043,44. Static measures produce consistent and reliable measures of MO2ROUTINE for damselfish species, likely due to constant pectoral fin movement45,46. Fish were starved for 12–24 h prior to metabolic testing to remove the effects of digestion on oxygen consumption. To measure MO2ROUTINE, fish were transferred to respirometers (295 mL opaque plastic jar with a 60 mm diameter, covered with black tape except for the bottom), which were submerged in a temperature controlled aquarium that mirrored their temperature treatment. Rate of oxygen usage was measured using the PreSens Fibox contactless sensor system (Precision Sensing GmbH). At the commencement of testing, flow to the respirometers was ceased and oxygen levels in each chamber were measured over a 20 min period. At the conclusion of MO2ROUTINE testing, the respirometers were opened to once again allow oxygenated water flow through the chamber. A pilot study determined that 2 hrs habituation in the respirometers was sufficient time for a fish to recover from handling (recorded lowest rates of oxygen consumption). This recovery time was similar to that found for other damselfish species45,47. To measure MO2MAX a swim respirometry chamber was used; consisting of a Perspex cylinder (150 mm inner diameter and 330 mL volume), below which a magnetic plate was placed to drive the stirring bar in the respirometer, creating a current, which the fish swam against28,43,48. The speed of the water was set so that the fish could maintain a sustained and consistent speed that was close to burst (which is sporadic). MO2MAX was calculated from 5 minutes of oxygen usage and was derived using the slope of the linear regression between oxygen availability and time, per fish. Both MO2ROUTINE and MO2MAX were calculated in mg O2 h−1, and the difference between these attributes was calculated as net aerobic scope (MO2MAX − MO2ROUTINE).
Statistical analysis
Potential differences in burst speed (bl/s) depending on temperature (fixed factor) and species (fixed factor) were tested using a factorial Analysis of Variance (ANOVA). In the case of feeding rates, growth rates (change in WW) and all metabolic variables (MO2ROUTINE, MO2MAX and Net Scope), individual size influenced the trait response (p < 0.05 for all results). Consequently, factorial GLMs were run with temperature and species as fixed factors, and mass (WW) as a covariate to account for the effect on the variable of interest. In the case of growth rate and feeding rate the initial WW of fish was the mass covariate. For all metabolic variables WW at the time of measurement was used as the covariate. To adhere to the linearity assumption, WW and the metabolic response variable were ln(x + 1) transformed prior to running analyses. The relationship between mass and the factor of interest (growth, feeding and respiration) was consistent across the temperatures (all p < 0.05). Least-square means adjusted for the effect of the covariate are presented throughout. Where results indicated significant differences, Tukey’s post hoc tests were run to identify differences between levels within a factor. Data were graphically inspected for normality and homogeneity of variance was examined using Levene’s test. Data conformed in all cases. All statistical analyses were completed using IBM SPSS® Statistics v23.
Compliance with ethical standards
Ethics approval for this study was obtained from the UTS Animal Care and Ethics (Approval Number: UTS ACEC 2012-433A). All applicable international, national, and/or institutional guidelines for the care and use of animal were followed.
Supplementary information
Acknowledgements
We’d like to firstly thank the School of Life Sciences (UTS) for supporting this research and providing the facilities that made the experiment possible. To Gemma Armstrong for her technical support in laboratory set up, and Cian-Foster Thorpe and Dr Nikki Bramwell for their aid in fish husbandry. Finally, a special thanks for Dr. Hayden Beck for his help and advice prior to the beginning of experimentation, and Associate Professor Will Figueira for loaning us the equipment necessary for metabolic testing. This research was conducted under collection permit NSW F94/696 and ethics approval UTS ACEC 2012-433A.
Author Contributions
L.D.D. conducted the experiments and led the first manuscript draft. J.M.D. was primarily responsible for the introduction of metabolic testing; the background, uses, implementation of its experimental procedures and the explanation of results gained. D.J.B. designed and planned the overall scope of the project; its research aims, experimental design and hypotheses to be tested. D.A.F. assisted in the early stages on the project design. Statistical design was overseen by A.M.F.
Data Availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Competing Interests
The University of Technology Sydney provided $1000 to assist in funding the project. With the exception of this all authors declare no conflict of financial and non-financial interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary information accompanies this paper at 10.1038/s41598-019-50303-z.
References
- 1.Bode M, Bode L, Armsworth PR. Different dispersal abilities allow reef fish to coexist. Proc. Nat. Acad. Sci. 2011;108:16317–16321. doi: 10.1073/pnas.1101019108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.García Molinos J, Burrows MT, Poloczanska ES. Ocean currents modify the coupling between climate change and biogeographical shifts. Sci. Reports. 2017 doi: 10.1038/s41598-017-01309-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Morrongiello JR, et al. Climate change and its implications for Australia’s freshwater fish. Mar. Freshw. Res. 2011;62:1082–1098. doi: 10.1071/MF10308. [DOI] [Google Scholar]
- 4.Pratchett MS, et al. Contribution of climate change to degradation and loss of critical fish habitats in Australian marine and freshwater environments. Mar. Freshw. Res. 2011;62:1062–1081. doi: 10.1071/MF10303. [DOI] [Google Scholar]
- 5.Parmesan C, Root TL, Willig MR. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature. 1999;399:579–583. doi: 10.1038/21181. [DOI] [Google Scholar]
- 6.Brown JH, Stevens GC, Kaufman DM. The geographical range: Size, shape, boundaries and internal structure. Annu. Rev. Ecol. Syst. 1996;27:597–623. doi: 10.1146/annurev.ecolsys.27.1.597. [DOI] [Google Scholar]
- 7.Walther GR, et al. Ecological responses to recent climate change. Nature. 2002;416:389–395. doi: 10.1038/416389a. [DOI] [PubMed] [Google Scholar]
- 8.Hickling R, Roy DB, Hill JK, Fox R, Thomas CD. The distributions of a wide range of taxonomic groups are expanding polewards. Global Change Biol. 2006;12:450–455. doi: 10.1111/j.1365-2486.2006.01116.x. [DOI] [Google Scholar]
- 9.Yamano H, Sugihara K, Nomura K. Rapid poleward range expansion of tropical reef corals in response to rising seas surface temperatures. Geo. Res. Lett. 2011;38:464–474. doi: 10.1029/2010GL046474. [DOI] [Google Scholar]
- 10.Drinkwater KF. The responses of Atlantic cod (Gadus morhua) to future climate change. J. Mar. Sci. 2005;62:1327–1337. [Google Scholar]
- 11.Perry AL, Low PJ, Ellis JR, Reynolds JD. Climate change and distribution shifts in marine fishes. Science. 2005;308:1912–1915. doi: 10.1126/science.1111322. [DOI] [PubMed] [Google Scholar]
- 12.Poloczanska ES, et al. Global imprint of climate change on marine life. Nature Clim. Change. 2013;3:919–925. doi: 10.1038/nclimate1958. [DOI] [Google Scholar]
- 13.Pechenik JA. On the advantages and disadvantages of larval stages in benthic marine invertebrate life cycles. Mar. Ecol. Prog. Ser. 1999;177:269–297. doi: 10.3354/meps177269. [DOI] [Google Scholar]
- 14.Booth DJ, Bond N, Macreadie P. Detecting range shifts among Australian fishes in response to climate change. Mar. Freshwater Res. 2011;62:1027–1042. doi: 10.1071/MF10270. [DOI] [Google Scholar]
- 15.Feary DA, et al. Latitudinal shifts in coral reef fishes: why some species do and others do not shift. Fish Fish. 2014;15:593–615. doi: 10.1111/faf.12036. [DOI] [Google Scholar]
- 16.Booth DJ, Figueira WF, Gregson MA, Brown L, Beretta G. Occurrence of tropical fishes in temperate south eastern Australia: role of the East Australian Current. Estuar. Coast Shelf Sci. 2007;72:102–114. doi: 10.1016/j.ecss.2006.10.003. [DOI] [Google Scholar]
- 17.Fowler, A. M., Parkinson, K. & Booth, D. J. New poleward observations of 30 tropical reef fishes in temperate southeastern. Australia. Mar Biodivers (2017).
- 18.Figueira WF, Booth DJ. Increasing ocean temperatures allow tropical fishes to survive overwinter in temperate waters. Global Change Biol. 2010;16:506–516. doi: 10.1111/j.1365-2486.2009.01934.x. [DOI] [Google Scholar]
- 19.Ridgway, K. & Hill, K. In The East Australian Current (eds Poloczanska, E. S., Hobday, A. J. & Richardson, A. J.) A marine climate change impacts and adaptation report card for Australia 2009. Publication 05/09, National Climate Change Adaptation Research Facility, Southport, QLD (2009).
- 20.Beck HJ, Feary DA, Fowler A, Madin E, Booth DJ. Temperature predators and seasonal water temperatures impact feeding of a range expanding tropical fish. Mar. Biol. 2016;163:70. doi: 10.1007/s00227-016-2844-8. [DOI] [Google Scholar]
- 21.Beck HJ, Feary DA, Nakamura Y, Booth DJ. Wave sheltered embayments are recruitment hotspots for tropical fishes. Mar. Ecol. Prog. Ser. 2016;546:197–212. doi: 10.3354/meps11599. [DOI] [Google Scholar]
- 22.Figueira WF, Biro P, Booth DJ, Valenzuela VC. Performance of tropical fish recruiting to temperate habitats: role of ambient temperature and implications of climate change. Mar Ecol Prog Ser. 2009;384:231–239. doi: 10.3354/meps08057. [DOI] [Google Scholar]
- 23.Warren DT, Donelson JM, McCormick MI, Ferrari MCO, Munday PL. Duration of exposure to elevated temperature affects competitive interactions in juvenile reef fishes. PloS One. 2016;11:e0164505. doi: 10.1371/journal.pone.0164505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lyon JP, Ryan TJ, Scroggie MP. Effects of temperature on the fast-start swimming performance of an Australian freshwater fish. Ecol. Freshw. Fish. 2008;17:184–188. doi: 10.1111/j.1600-0633.2007.00244.x. [DOI] [Google Scholar]
- 25.Hofmann GE, Todgham AE. Living in the now: Physiological mechanisms to tolerate a rapidly changing environment. Annu. Rev. Physiol. 2010;72:127–145. doi: 10.1146/annurev-physiol-021909-135900. [DOI] [PubMed] [Google Scholar]
- 26.Pörtner HO. Climate variations and the physiological basis of temperature dependent biogeography: systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. A. 2002;132:739–761. doi: 10.1016/S1095-6433(02)00045-4. [DOI] [PubMed] [Google Scholar]
- 27.Pörtner HO, Knust R. Climate change affects marine fishes through oxygen limitation of thermal tolerance. Science. 2007;315:95–97. doi: 10.1126/science.1135471. [DOI] [PubMed] [Google Scholar]
- 28.Nilsson GE, Crawley N, Lunde IG, Munday PI. Elevated temperature reduces the respiratory scope if coral reef fishes. Glob. Change Biol. 2009;15:1405–1412. doi: 10.1111/j.1365-2486.2008.01767.x. [DOI] [Google Scholar]
- 29.Donelson JM, Munday PL, McCormick MI, Nilsson GE. Acclimation to predicted ocean warming through development plasticity in a tropical reef fish. Global. Change Biol. 2011;17:1712–1719. doi: 10.1111/j.1365-2486.2010.02339.x. [DOI] [Google Scholar]
- 30.Sogard SM. Size-selectivity mortality in the juvenile stages of teleost fishes: a review. Bull. Mar. Sci. 1997;60:1129–1157. [Google Scholar]
- 31.Preuss T, Faber DS. Central cellular mechanisms underlying temperature-dependent changes in the goldfish startle-escape behavior. J. Neurosci. 2003;23:5617–5626. doi: 10.1523/JNEUROSCI.23-13-05617.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pörtner HO, Farrell AP. Physiology and Climate Change. Science. 2008;322:690–692. doi: 10.1126/science.1163156. [DOI] [PubMed] [Google Scholar]
- 33.Videler JJ, Weihs D. Energetic advantages of burst-and-coast swimming of fish at high speeds. J. Exp. Biol. 1982;97:169–178. doi: 10.1242/jeb.97.1.169. [DOI] [PubMed] [Google Scholar]
- 34.Eaton RC, Bomardieri RA, Meyer OH. The Mauthner-initiated startle response in teleost fish. J. Exp. Biol. 1977;66:65–81. doi: 10.1242/jeb.66.1.65. [DOI] [PubMed] [Google Scholar]
- 35.Fay RR, Ream TJ. The effects of temperature change and transient hypoxia on auditory nerve fiber response in the goldfish (Carassius auratus) Hearing Research. 1992;589:9–18. doi: 10.1016/0378-5955(92)90003-6. [DOI] [PubMed] [Google Scholar]
- 36.Eaton RC, Lavender WA, Wieland CM. Identification of Mauthnerinitiated patterns in goldfish: evidence from simultaneous cinematography and electrophysiology. J. Compar. Physiol. A. 1981;144:521–531. doi: 10.1007/BF01326837. [DOI] [Google Scholar]
- 37.Day T, Abrams PA, Chase JM. The role of size‐specific predation in the evolution and diversification of prey life histories. Evolution. 2002;56:877–887. doi: 10.1111/j.0014-3820.2002.tb01401.x. [DOI] [PubMed] [Google Scholar]
- 38.Beck HJ, Feary DA, Figueira WF, Booth DJ. Assessing range shifts of tropical reef fishes: a comparison of belt transect and roaming underwater census methods. Bull. Mar. Sci. 2014;90:705–721. doi: 10.5343/bms.2013.1055. [DOI] [Google Scholar]
- 39.Berec L, Angulo E, Courchamp F. Multiple Allee effects and population management. Trends Ecol. Evol. 2007;22:185–191. doi: 10.1016/j.tree.2006.12.002. [DOI] [PubMed] [Google Scholar]
- 40.Munday PL, Wilson SK. Comparative efficacy of clove oil and other chemicals in anaesthetization of Pomacentrus amboinensis, a coral reef fish. J. Fish. Biol. 1997;51:931–938. [Google Scholar]
- 41.Australian Marine Institute of Science. Bleaching Thresholds: Heron Island, http://data.aims.gov.au/aimsrtds/yearlytrends.xhtml?fromDate=2012&dataLevel=1&wind=false.%20%C2%A9%20Australian%20Institute%20of%20Marine%20Science%202013 (2013).
- 42.Batty RS, Blaxter JHS. The effect of temperature on the burst swimming performance of fish larvae. J. Exp. Biol. 1992;170:1187–201. [Google Scholar]
- 43.Seebacher F, Ward AJW, Wilson RS. Increased aggression during pregnancy comes at a higher metabolic cost. J. Exp. Biol. 2013;216:771–776. doi: 10.1242/jeb.079756. [DOI] [PubMed] [Google Scholar]
- 44.Rodgers GG, Tenzing P, Clark TD. Experimental methods in aquatic respirometry: the importance of mixing devices and accounting for background respiration. J. Fish. Biol. 2016;88:65–80. doi: 10.1111/jfb.12848. [DOI] [PubMed] [Google Scholar]
- 45.Bernal AM, et al. Phenotypic and molecular consequences of stepwise temperature increase across generations in a coral reef fish. Mol Ecol. 2018 doi: 10.1111/mec.14884. [DOI] [PubMed] [Google Scholar]
- 46.Rodgers GG, Rummer JL, Johnson LK, McCormick MI. Impacts of increased ocean temperature on a low-latitude coral reef fish – Processes related to oxygen uptake and delivery. J. Therm. Biol. 2019;79:95–102. doi: 10.1016/j.jtherbio.2018.12.008. [DOI] [PubMed] [Google Scholar]
- 47.Rummer JL, et al. Life on the edge: thermal optima for aerobic scope of equatorial reef fishes are close to current day temperatures. Global Change Biol. 2014;20:1055–1066. doi: 10.1111/gcb.12455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Donelson JM, Munday PL. Thermal sensitivity does not determine acclimation capacity for a tropical reef fish. J. Anim. Ecol. 2012;81:1126–1131. doi: 10.1111/j.1365-2656.2012.01982.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.