

ABSTRACT
Salicylic acid (SA) is involved in several plant processes including responses to abiotic stresses. Although SA is thought to interact with other regulatory molecules in a complex way, currently, little information is available regarding its molecular mechanisms of action in response to abiotic stresses. In a previous work, we observed that drought-resistant oat plants significantly increased their SA levels as compared with a susceptible cultivar. Furthermore, exogenous SA treatment alleviated drought symptoms. Here, we investigated the interaction between SA and polyamine biosynthesis during drought responses in oat and revealed that SA regulated polyamine biosynthesis through changes in polyamine gene expression. Overall, SA treatment decreased the levels of putrescine under drought conditions while increased those of spermine. This correlates with the downregulation of the ADC gene and upregulation of the AdoMetDC gene. Based on the presented results, we propose that SA modulates drought responses in oat by regulating polyamine content and biosynthesis.
KEYWORDS: Drought, oat, polyamine, salicylic acid
Salicylic acid (SA) is an important plant hormone naturally occurring in plants. Its role in plant–pathogen interactions has been widely documented.1 In addition, SA is involved in the regulation of plant development and growth, flowering, and ripening.2 More recently, it was also reported to be involved in plant responses to abiotic stresses.3 Thus, SA mediates the response to salinity, low temperature, and drought.4 Consequently, SA has been proposed to have a great agronomic potential to improve stress tolerance in economically important crops. However, only sparse information is available to date regarding the molecular mechanisms of SA in response to abiotic stresses. Furthermore, the effect of SA in tolerance to abiotic stresses is under controversy due to the concentration-dependent effect of this hormone. Accordingly, low concentrations of applied SA alleviate the sensitivity to abiotic stresses, whereas high concentrations lead to a decreased tolerance due to the induction of oxidative stress.3 In a previous metabolomic study,5 we observed an induction of the SA biosynthetic pathway in a drought-resistant oat cultivar (Patones) as compared with a susceptible cultivar (Flega). Further studies showed that the resistant cultivar increased SA content when subjected to gradual water depletion compared with well-watered or susceptible plants and that exogenous SA applications at 100 µM alleviated drought symptoms in part due to a reduction of stomatal conductance.5
SA is thought to interact in a complex way with other hormonal compounds.6 It has been reported to interact with ethylene during heat stress,7 and it is also among the major phytohormones interacting with nitric oxide (NO).8 Both ethylene and NO are directly related to polyamine content, and our previous data indicated a complex interplay between ethylene, NO, and polyamines during drought tolerance responses.9 Indeed, most common polyamines such as the diamine, putrescine (Put), the triamine, spermidine (Spd), and the tetra-amine, spermine (Spm), act as regulatory molecules in many fundamental regulatory processes as well as in stress responses. However, there is little information on the direct SA–polyamine interaction during drought stress responses apart from a study from Németh et al.10 in maize in which SA caused sensitivity to drought, maybe due to the relatively high concentration applied. We hypothesized that the SA-mediated alleviation of drought symptoms observed in oat might be due to its interaction with polyamines.
To test this hypothesis, oat plants from the resistant Patones and susceptible Flega cultivars were grown and subjected to drought as previously.5,11,12 At the beginning of the drought treatment, the plants were sprayed with 100 μM SA or 4-hydroxibenzoic acid (4-hBA), an isomer of SA with no biological activity13 as control. As previously,5 we observed an alleviation of drought symptoms in SA-treated plants. Quantification of polyamines by high-pressure liquid chromatography revealed significantly reduced levels of putrescine in resistant Patones plants under drought conditions as compared with Flega (p= 0.009). Furthermore, SA treatment reduced the amount of this polyamine in both cultivars (p < 0.001) compared with 4-hBA-treated plants under drought conditions but not under the well-watered conditions (Figure 1). Differential effect of plant hormones under well-watered or drought conditions has been previously reported. For instance, the accumulation of Abscisic acid (ABA) in roots helps to maintain growth under drought stress conditions but not when applying ABA to well-watered plants.14 Increased level of putrescine accelerates senescence and has been associated with chlorophyll loss,15 features that were also observed in plants under water stress. Thus, the reduction of putrescine observed in SA-treated plants supports the drought symptom alleviation observed. Interestingly, SA reduced the levels of spermidine in resistant Patones plants but not in susceptible Flega. This could be due to a different sensitivity of the two cultivars as also reported in tomato during salt stress, where exogenous SA affected polyamine production in the resistant cultivar but not in the susceptible.16 On the other hand, SA significantly increased spermine content in both cultivars. Spermine has been associated with drought tolerance in other species. Thus, spermine pretreatment reversed the drought hypersensitive phenotype of the Arabidopsis acl5/spms mutant otherwise unable to produce spermine.17 This polyamine may modulate the activity of certain ion channels increasing Ca2+ and regulating stomatal closure.17 Thus, spermine could contribute to the previously observed reduction of stomatal conductance mediated by SA.5 In addition, a role for spermine increasing the antioxidative capacity has also been reported.18 In this sense, the increase in spermine observed would also, at least partially, explain the increase in antioxidant capacity and related activities observed in Patones under drought.5
Figure 1.
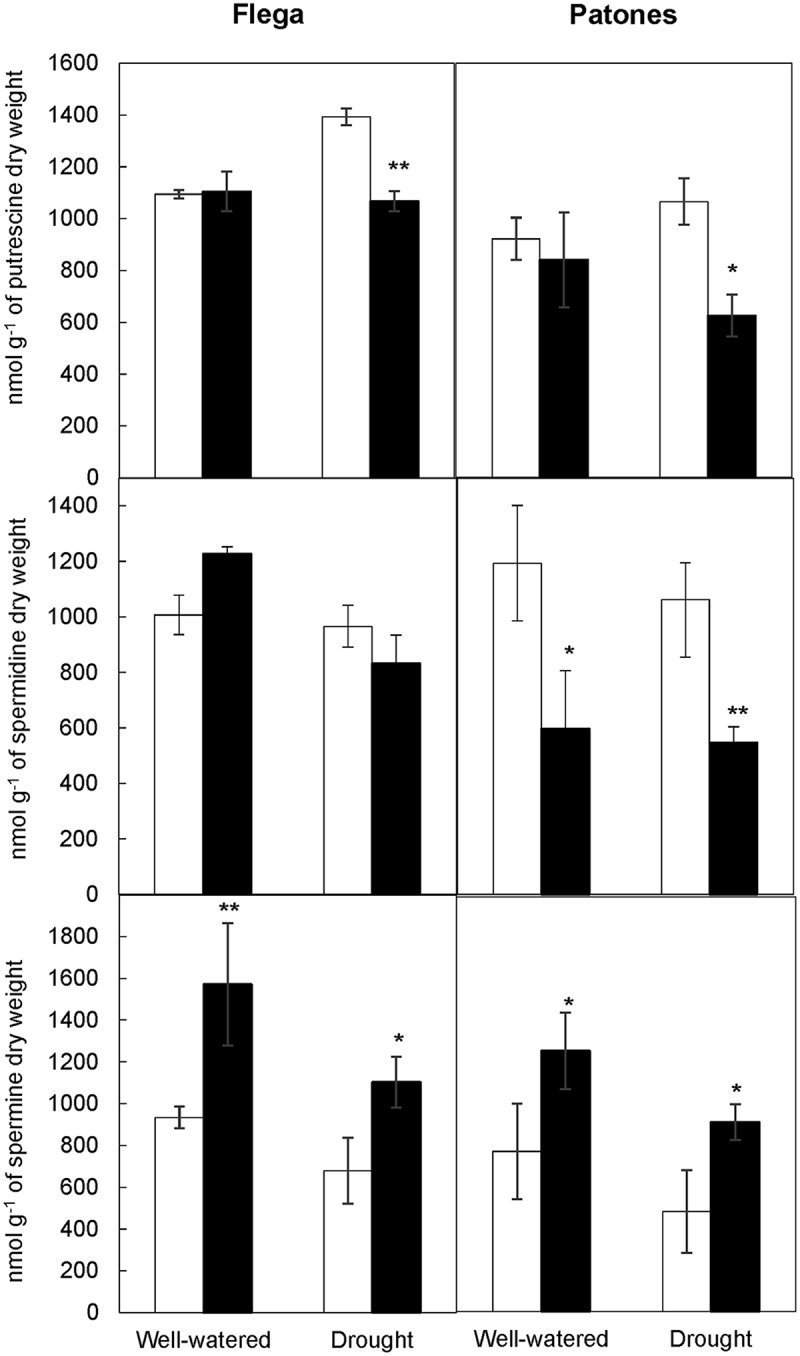
Effect of salicylate on polyamine content. Putrescine, spermidine, and spermine were measured in well-watered or droughted Flega (drought susceptible) and Patones (drought resistant) plants treated with salicylate 100 µM (black bars) or with the biologically inactive isomer 4-hydroxybenzoic acid (4-hBA) (control, white bars). Data are mean of five replicates ± standard error. * and ** indicate significant differences between treatments at p < 0.05 and 0.01, respectively.
Expression studies of the main genes involved in polyamine biosynthesis by qRT-PCR supported the previous conclusions. Thus, the downregulation of ADC gene during drought stress might contribute to the low levels of putrescine observed (Figure 2). Surprisingly, whereas ADC expression increased in SA-treated well-watered plants, putrescine levels were not altered. This suggests that under well-watered conditions, the increase in ADC expression might lead to accumulation of polyamine-related compounds such as agmatine, which acts as reservoir and precursor of polyamines or be derived to other related biosynthetic pathways such as the tricarboxylic acid pathway.9 The increased levels of spermine observed were supported by an overall upregulation of S-adenosylmethionine decarboxylase (AdoMetDC) gene, which is directly involved in spermine biosynthesis. Interestingly, Flega cultivar also showed a differential behavior since SA-treated plants under drought did not show any change on AdoMetDC expression. Nevertheless, the level of spermine also increased in this cultivar following SA treatment. This suggested that SA could influence other polyamine-related processes such as induction of enzymatic activities or post-translational modifications leading to spermine production, as it has been reported with other enzymes.19 Indeed, it has been described that spermine itself is able to inhibit the post-translational modification required by ADC to form putrescine, decreasing putrescine accumulation and the associated senescence.20 Some controversy exists regarding the effect of SA on ethylene, with works reporting its inhibition, stimulation, or absence of an effect. Our data show no significant changes in methionine adenosyltransferase expression following SA treatment. This would support a role for SA during drought-regulating polyamine biosynthesis without affecting ethylene production, which has been related to drought-associated senescence symptoms.
Figure 2.
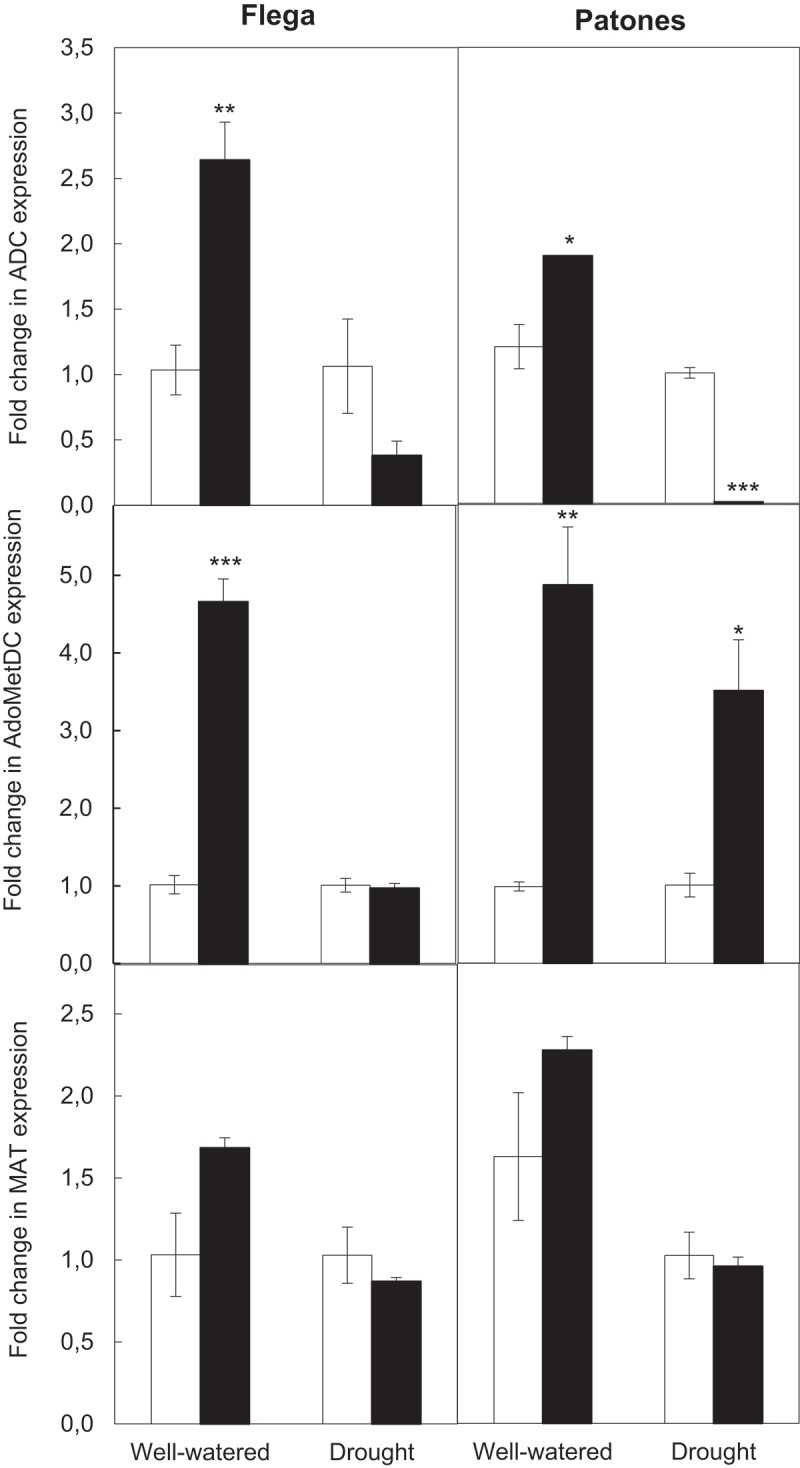
Effect of salicylate on the expression of several genes involved in polyamine biosynthesis. Expression of arginine decarboxylase (ADC), S-adenosylmethionine decarboxylase (AdoMetDC), and methionine adenosyltransferase (MAT) was measured in well-watered or droughted Flega (drought susceptible) and Patones (drought resistant) plants treated with salicylate 100 µM (black bars) or with the biologically inactive isomer 4-hydroxybenzoic acid (4-hBA) (control, white bars). Oat GAPDH gene was used as an internal control to normalize gene expression. Data, expressed as fold change in expression of SA-treated plants with respect to controls (4-hBA-treated plants), are mean of at least three independent biological plus three technical replications ± standard error. *, **, and *** indicate significant differences with respect to treatments at p < 0.05, 0.01, and 0.001, respectively.
In summary, we observed that SA modulated polyamine content under drought conditions by influencing the expression of its main biosynthetic genes. These data shed light on the complex interaction of SA with polyamines and may help to build the complex scheme of signaling networks, which could contribute to alleviate drought adverse effect in plants.
Funding Statement
This work was supported by the Spanish Ministry of Economy and Competitiveness [AGL2016-78965AGR], regional government through the AGR-253 group, and the European Regional and Social Development Funds. FJCC is the holder of an FPI fellowship BES-2014-071044 from the Spanish Ministry of Economy and Competitiveness.
Acknowledgments
We thank the Spanish Ministry of Economy and Competitiveness, the regional government, and the European Regional and Social Development Funds for jointly sponsoring this study.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
References
- 1.Dempsey DA, Vlot AC, Wildermuth MC, Klessig DF.. Salicylic acid biosynthesis and metabolism, in Arabidopsis book. Vol. 9 2011. e0156. doi: 10.1199/tab.0156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Vicente MRS, Plasencia J.. Salicylic acid beyond defence: its role in plant growth and development. J Exp Bot. 2011;62:3321–3338. doi: 10.1093/jxb/err031. [DOI] [PubMed] [Google Scholar]
- 3.Hara M, Furukawa J, Sato A, Mizoguchi T, Miura K. Abiotic stress and role of salicylic acid in plants In: Ahmad P, Prasad MNV, editors. Abiotic stress responses in plants: metabolism, productivity and sustainability. 2012. p. 235–251. [Google Scholar]
- 4.Miura K, Tada Y. Regulation of water, salinity, and cold stress responses by salicylic acid. Front Plant Sci. 2014;5. doi: 10.3389/fpls.2014.00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sánchez-Martín J, Heald J, Kingston-Smith A, Winters A, Rubiales D, Sanz M, Mur LAJ, Prats E. A metabolomic study in oats (Avena sativa) highlights a drought tolerance mechanism based upon salicylate signalling pathways and the modulation of carbon, antioxidant and photo-oxidative metabolism. Plant Cell Environ. 2015;38:1434–1452. doi: 10.1111/pce.12501. [DOI] [PubMed] [Google Scholar]
- 6.Khan MIR, Fatma M, Per TS, Anjum NA, Khan NA. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front Plant Sci. 2015;6. doi: 10.3389/fpls.2015.00462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Khan MI, Iqbal N, Masood A, Per TS, Khan NA. Salicylic acid alleviates adverse effects of heat stress on photosynthesis through changes in proline production and ethylene formation. Plant Signal Behav. 2013;8:e26374. doi: 10.4161/psb.26374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Alavi SMN, Arvin MJ, Kalantari KM. Salicylic acid and nitric oxide alleviate osmotic stress in wheat (Triticum aestivum L.) seedlings. J Plant Interact. 2014;9:683–688. doi: 10.1080/17429145.2014.900120. [DOI] [Google Scholar]
- 9.Montilla-Bascón G, Rubiales D, Hebelstrup KH, Mandon J, Harren FJM, Cristescu SM, Mur LAJ, Prats E. Reduced nitric oxide levels during drought stress promote drought tolerance in barley and is associated with elevated polyamine biosynthesis. Sci Rep. 2017;7:13311. doi: 10.1038/s41598-017-13458-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nemeth M, Janda T, Horvath E, Paldi E, Szalai G. Exogenous salicylic acid increases polyamine content but may decrease drought tolerance in maize. Plant Sci. 2002;162:569–574. doi: 10.1016/S0168-9452(01)00593-3. [DOI] [Google Scholar]
- 11.Sánchez-Martín J, Canales FJ, Tweed JKS, Lee MRF, Rubiales D, Gomez-Cadenas A, Arbona V, Mur LAJ, Prats E. Fatty acid profile changes during gradual soil water depletion in oats suggests a role for jasmonates in coping with drought. Front Plant Sci. 2018;9. doi: 10.3389/fpls.2018.01077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sánchez-Martín J, Mur LAJ, Rubiales D, Prats E. Targeting sources of drought tolerance within an Avena spp. collection through multivariate approaches. Planta. 2012;236:1529–1545. doi: 10.1007/s00425-012-1709-8. [DOI] [PubMed] [Google Scholar]
- 13.Medina-Puche L, Castello MJ, Canet JV, Lamilla J, Colombo ML, Tornero P. Beta-carbonic anhydrases play a role in salicylic acid perception in Arabidopsis. PLoS One. 2017;12. doi: 10.1371/journal.pone.0181820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sharp RE, Wu YJ, Voetberg GS, Saab IN, Lenoble ME. Confirmation that abscisic-acid accumulation is required for maize primary root elongation at low water potentials. J Exp Bot. 1994;45:1743–1751. doi: 10.1093/jxb/45.Special_Issue.1743. [DOI] [Google Scholar]
- 15.Capell T, Escobar C, Liu H, Burtin D, Lepri O, Christou P. Over-expression of the oat arginine decarboxylase cDNA in transgenic rice (Oryza sativa L.) affects normal development patterns in vitro and results in putrescine accumulation in transgenic plants. Theor Appl Genet. 1998;97:246–254. doi: 10.1007/s001220050892. [DOI] [Google Scholar]
- 16.Gharbi E, Martinez JP, Benahmed H, Fauconnier ML, Lutts S, Quinet M. Salicylic acid differently impacts ethylene and polyamine synthesis in the glycophyte Solanum lycopersicum and the wild-related halophyte Solanum chilense exposed to mild salt stress. Physiol Plant. 2016;158:152–167. doi: 10.1111/ppl.12458. [DOI] [PubMed] [Google Scholar]
- 17.Yamaguchi K, Takahashi Y, Berberich T, Imai A, Takahashi T, Michael AJ, Kusano T. A protective role for the polyamine spermine against drought stress in Arabidopsis. Biochem Biophys Res Commun. 2007;352:486–490. doi: 10.1016/j.bbrc.2006.11.041. [DOI] [PubMed] [Google Scholar]
- 18.Fu XZ, Xing F, Wang NQ, Peng LZ, Chun CP, Cao L, Ling LL, Jiang CL. Exogenous spermine pretreatment confers tolerance to combined high-temperature and drought stress in vitro in trifoliate orange seedlings via modulation of antioxidative capacity and expression of stress-related genes. Biotechnol Biotec Eq. 2014;28:192–198. doi: 10.1080/13102818.2014.909152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Caarls L, Pieterse CMJ, Van Wees SCM. How salicylic acid takes transcriptional control over jasmonic acid signaling. Front Plant Sci. 2015;6. doi: 10.3389/fpls.2015.00170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Capell T, Bassie L, Christou P. Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress. Proc Natl Acad Sci U S A. 2004;101:9909–9914. doi: 10.1073/pnas.0306974101. [DOI] [PMC free article] [PubMed] [Google Scholar]