

Abstract
Applications of remote sensing data to monitor bird migration usher a new understanding of magnitude and extent of movements across entire flyways. Millions of birds move through the western USA, yet this region is understudied as a migratory corridor. Characterizing movements in the Pacific Flyway offers a unique opportunity to study complementary patterns to those recently highlighted in the Atlantic and Central Flyways. We use weather surveillance radar data from spring and autumn (1995–2018) to examine migrants' behaviours in relation to winds in the Pacific Flyway. Overall, spring migrants tended to drift on winds, but less so at northern latitudes and farther inland from the Pacific coastline. Relationships between winds and autumn flight behaviours were less striking, with no latitudinal or coastal dependencies. Differences in the preferred direction of movement (PDM) and wind direction predicted drift patterns during spring and autumn, with increased drift when wind direction and PDM differences were high. We also observed greater total flight activity through the Pacific Flyway during the spring when compared with the autumn. Such complex relationships among birds’ flight strategies, winds and seasonality highlight the variation within a migration system. Characterizations at these scales complement our understanding of strategies to clarify aerial animal movements.
Keywords: aeroecology, bird migration, Pacific Flyway, radar, remote sensing, wind drift
1. Introduction
Each spring and autumn, billions of migratory birds navigate to and from their breeding and wintering ranges [1–3]. Yet, flight strategies employed during these flights, which can be at high altitudes and often occur at night, are still poorly understood. New technologies are beginning to provide detailed insights into annual activity patterns [4,5], yet still, sampling in-flight behaviours of speed and direction are challenging to acquire, especially of small-bodied birds. These measures are especially important when considering birds' responses to changing atmospheric conditions (e.g. winds aloft). Wind speed and direction are primary drivers for nightly flight initiation [6]; however, en route birds must also contend with being blown off their preferred direction of movement (PDM) by crosswinds [7,8]. To counter the influence of crosswinds, in-flight migrants can compensate by increasing their airspeeds or adjusting their headings (i.e. body axis orientation) to counter wind drift [9]. While heading adjustments minimize overall flight distances, they can lower groundspeeds, thereby slowing the pace of migration and potentially increasing energy expenditure. However, the consequences of drifting off course to unhospitable or deadly environments (e.g. open water) could have ultimate fitness consequences. Season [10,11], topographic barriers [12] and proximity to the breeding destination [13] can shape these flight strategies and fitness consequences.
In North America, the Pacific Flyway—in this case representing bird migration over the land area primarily west of the Rocky Mountains—is generally underrepresented in studies of bird migration [14], with significantly more research occurring on movements east of the Rockies. This region presents a unique opportunity to test how migrants vary flight strategies in complex and geographically diverse wind patterns [15], varied topography and season-dependent resource availability [16] across an extensive latitudinal gradient (greater than 15°). To capture the macro-scale orientation strategies of nocturnally migrating birds moving through the Pacific Flyway, we leverage the US weather surveillance radar network [17]. This network allows for a multi-decadal examination of these behaviours, amassing hundreds of thousands of individual measurements of flight activity. These measurements can reveal seasonal timing of migration, the spatial distribution of migrants and the degree of behavioural plasticity in flight strategies (i.e. the extent to which birds can compensate and drift in changing wind patterns). We hypothesize that the degree of wind drift depends upon proximity to the Pacific coastline, season and latitude (as an indication of distance to end destination). We predict that migrants drift more further from the Pacific coastline because the risks associated with being blown off course diminishes moving away from open water [12]. We predict greater levels of drift during the autumn because of the abundance of young, inexperienced migrants, and that migrants will drift less as they near their end destinations [13].
2. Material and methods
(a). Weather surveillance radar
We characterized migration intensity, speed, track and heading across the Pacific Flyway of the USA (figure 1) by extracting migrant flight data from 19 west coast weather surveillance radar (WSR) stations during spring (1 March to 15 June) and autumn (1 August to 15 November) from spring 1995 to spring 2018. We acquired radar data through the NEXRAD Level-II archive available publicly on Amazon Web Services (AWS) and processed Level-II data using WSR-LIB [18]. To characterize migratory activity through the night, we subsampled scans every half hour between local sunset and sunrise. We sampled data between 5 and 37.5 km from the radar in the 0.5–4.5° elevation sweeps and constructed vertical profiles of bird activity from 0 to 3 km above ground level at 100 m intervals [19]. We derived migrant intensity from reflectivity (η), calculated by converting reflectivity factor following: η[dB] = Z[dBZ] + β, where β = 10 log10(103π5|Km|2/λ4) [20]. We used an average WSR-88D wavelength (λ) of 10.7 cm and |Km|2 for liquid water of 0.93, the dielectric constant. For each vertical profile, we calculated the mean η across the entire profile and used a cube root transformation on η [6]. We calculated nightly means to characterize migrant flight activity and aggregated these means across sites and seasons to examine spatial differences in migrant activity. We used velocity azimuth displays (VAD) on radial velocity [21] to determine migrant track and groundspeed. When necessary, we used the methodology of [22] to de-alias radial velocity measures. These vertical profiles reflect aggregated nocturnal movements that cannot be attributed to specific species.
Figure 1.

Spring (1 March to 15 June, a,b) and autumn (1 August to 15 November, c,d) distributions of migrant track (pink), heading (blue) and wind direction (grey) at weather surveillance radar (WSR) stations summarized from 1995 to 2018 between sunset and sunrise. Points show WSR station location and are shaded by station latitude. Station points (a,c) are scaled to the cube root of the average summed seasonal migration intensity and station points (b,d) are scaled by the rho of wind direction (small points = low directionality). Flight distributions are weighted by the cube root of flight activity and wind direction by the product of the cube root of flight activity and wind speed. Arrows' directions denote PDM (a,c) and average wind direction (b,d). PDM arrows’ lengths (a,c) are scaled to average migrant groundspeed and wind arrows' lengths (b,d) are scaled to the average wind speed. Note, all distributions are scaled to the same size and number of observations, but exhibit differing maxima. Rose diagrams are summarized in 5° sectors. (Online version in colour.)
We removed precipitation contamination on a per-pixel level within the sampling range using a deep-learning classification algorithm, MISTNET, that leverages a convolutional neural network trained on 239 128 samples, with per-pixel accuracy of 97. 3% (precision 98.7%; recall 95.9%) [23]. We used North American Regional Reanalysis (NARR) [24] data to calculate wind speed and direction within the radar sampling region and linked radar and wind variables to their respective height bins. NARR data are assembled at 3 h temporal intervals, 32 km spatial resolution and are modelled at 25 hPa vertical intervals. We calculated airspeed and heading direction using vector subtraction from ground speed and track direction (derived from VAD). We removed 100 m vertical sampling bins with targets with airspeeds less than 5 m s−1 to mitigate the influence of insects in our analysis [25].
(b). Statistics
We estimated the date of peak migration for each WSR station by fitting a generalized additive mixed model to reflectivity with ordinal date as the predictor variable [26]. We included year as a random effect and termed peak migration as the date at maximum predicted reflectivity. To examine the dependence of peak migration date on latitude, we fit a least-squares linear model with latitude as the predictor variable. We used two paired t-tests to examine seasonal differences in (i) summed migration activity and (ii) propensity of drift (i.e. slope of α; see below for calculation).
To quantify the degree of wind drift, we used a mixed-model approach, regressing track (°) on the difference between track and heading (termed α, °) [27]. This approach generated two metrics describing migrant flight behaviour: (i) slope of α, a measure of drift propensity (0, complete wind drift compensation; 1, complete wind drift); and (ii) y-intercept, a measure of PDM (the composite track direction) under no crosswind drift (PDM) [9]. PDM in our analysis is the aggregate direction of many species (i.e. the nocturnal migration system). We included random intercept terms of station, date, station × date and station × year [13,28]. We included one random slope term, of α varying on station. We weighted our analysis by the cube root of radar reflectivity. We weighted summaries of migratory track, heading and measures of drift by the cube root of migratory intensity to prioritize dominant periods of movement [6] and only included data between the α range of −90° to 90° (11.7% of data outside this range).
To summarize the winds migrants used, we weighted wind directions by the product of the cube root of migratory intensity (η) and wind speed (m s−1). Our weighting procedure prioritized winds used by migrants aloft, both by including migrant intensity (i.e. winds used by migrants) and wind speed (i.e. winds with large effects on migrants). This procedure prevented equal weighting of all wind measures.
To examine spatial differences in flight behaviours, we fit a least-squares linear model to slope of α with latitude, distance to coastline and the interaction of latitude and distance to coastline as predictors. Lastly, to determine the dependence of slope of α on differences of PDM and wind direction, we fit a generalized addition model (GAM). These models were fit for each season.
3. Results
We sampled 2475 nights during spring (458 545 30 min samples) and 2429 nights (503 735 samples) during autumn, totalling 962 280 samples. Spring migration traffic peaked between 28 April and 17 May (5 May ± 5.4 days, mean ± s.d.) and autumn migration between 9 September and 7 October (23 September ± 6.4 days, mean ± s.d.). Latitude predicted the date of peak migration during spring (F1,17 = 17.79, p < 0.001, R2 = 0.51) but not during autumn (F1,17 = 0.0015, p = 0.969, R2 < 0.001). Most sites showed higher summed activity in spring compared to autumn (13 of 19 sites, figure 1a,c), and the overall mean activity was significantly higher in spring (paired t-test, t18 = 2.364, p = 0.0295).
Spring preferred directions of movement (PDM) ranged between 304.2°(KVTX, see figure 1a for radar labels) and 23.7°(KPDT) and averaged 347.3° ± 22.1°(±s.d.). Autumn PDMs ranged between 136.9°(KSOX) and 182.2°(KBHX) and averaged 156.5° ± 12.6°(±s.d.) (figure 1c).
There was no evidence that drift differed between seasons: spring (mean = 0.42 ± 0.25, s.d.), autumn (mean = 0.27 ± 0.25, s.d.) (paired t-test, t18 = 1.443, p = 0.1662, figure 2a). During spring, latitude (p < 0.01) and distance to coastline (p < 0.05) significantly affected the propensity of drift (table 1). Birds generally drifted less with increasing latitude and farther from the coastline (electronic supplementary material, figure S1). Changes in drift behaviours in relation to distance to coastline were more prominent at southern latitudes (p < 0.05, see interaction plot; electronic supplementary material, figure S1). During autumn, we did not find that the propensity of drift changed with latitude (p = 0.177) or distance to coastline (p = 0.453), nor any interaction (p = 0.553, table 1).
Figure 2.
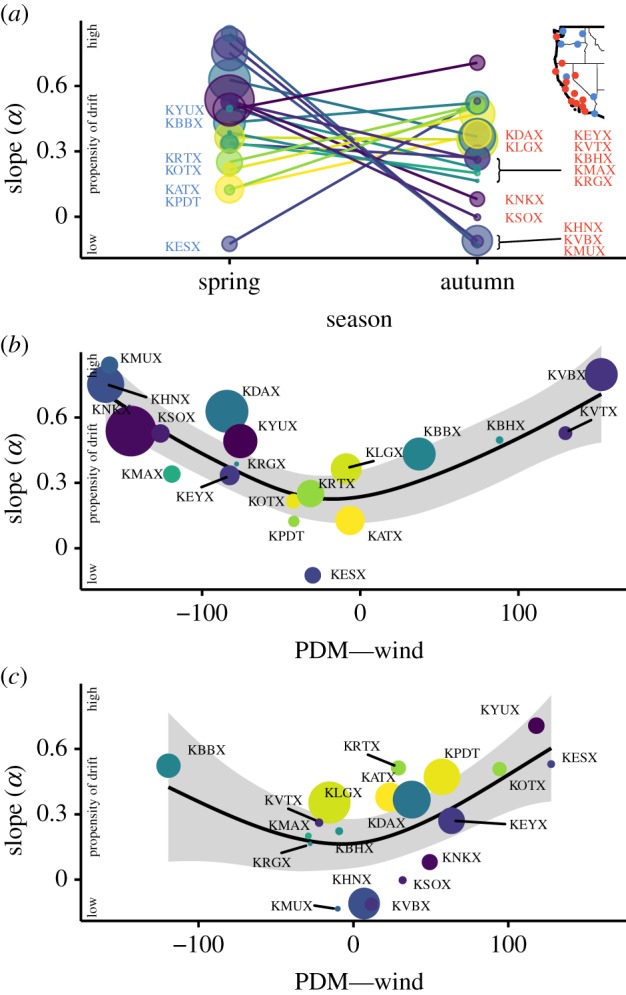
(a) Seasonal propensity of drift (i.e. slope of α). Radar site labels shaded in blue show increasing slope of α values (i.e. more drift) from spring to aututmn. Those in red show decreasing values (i.e. less drift). Inset shows the locations of decreasing (red) and increasing (blue) drift. (b) Spring and (c) autumn mean difference in PDM and wind direction against the propensity of drift (i.e. slope of α). Grey error bars represent 95% confidence intervals from generalized additive model. All points are shaded by WSR station latitude and the size is scaled to the cube root of migration intensity. (Online version in colour.)
Table 1.
Summary of two-way interaction least-squares linear model predicting slope of α across 19 weather surveillance stations during spring and autumn migratory periods.
| parameter | estimate | s.e. | t-value | P (>|t|) |
|---|---|---|---|---|
| spring | ||||
| intercept | 1.775 | 0.376 | 4.718 | 0.000275 |
| latitude | −0.0310 | 0.00951 | −3.262 | 0.00525 |
| distance to coastline (km) | −0.00640 | 0.00247 | −2.594 | 0.0203 |
| latitude: distance to coastline (km) | 0.000130 | 0.0000590 | 2.199 | 0.0440 |
| autumn | ||||
| intercept | −0.556 | 0.537 | −1.035 | 0.317 |
| latitude | 0.0192 | 0.0136 | 1.416 | 0.177 |
| distance to coastline (km) | 0.00271 | 0.00352 | 0.771 | 0.453 |
| latitude: distance to coastline (km) | 0.0000510 | 0.0000840 | −0.607 | 0.553 |
Migrants regularly used winds that opposed their seasonal PDMs (figure 1b,d). The absolute value of spring differences between wind direction and PDM decreased with increasing latitude (linear regression, F1,17 = 15.82, slope = −6.759, p = 0.0010, R2 = 0.482) but did not change with latitude during the autumn (linear regression, F1,17 = 0.3767, p = 0.547, R2 = 0.0217). The differences between PDM and wind direction were predictive of slope of α-values, both during the spring (GAM, F2.692 = 8.367, p = 0.00107, deviance explained = 67.2%, figure 2b) and the autumn (GAM, F2.221 = 3.909, p = 0.0283, deviance explained = 45.3%, figure 2c), with greater levels of drift when wind direction and PDM differences were high.
4. Discussion
We quantify flight strategies employed by migratory birds passing through the Pacific Flyway by leveraging over two decades of weather surveillance data to characterize migrant track, heading, speed and density. Migrants varied their aerial behaviours to the complex assemblage of available winds and altered their strategies across gradients of distance to the coastline, season and latitude.
(a). Geographical and seasonal variation in flight strategies
We observed a general decrease in wind drift with increasing latitudes during spring, supporting our prediction of an optimal migration strategy in which birds reduce wind drift upon approaching breeding destinations [7,8,13]. Contrary to our prediction, drift decreased farther from the coastline, but only during spring. These behavioural changes also coincided with geographically variable wind patterns. Winds tended to be in greater opposition to northward passage along the southern coastline. Differences between PDM and wind direction may exceed migrants’ abilities to compensate and result in elevated levels of drift. These findings contrast behaviours of migrants in the Atlantic Flyway, where birds tend to compensate more at coastal sites to avoid being blown over the ocean [12]. In our analysis, one site in spring (KESX) and three in autumn (KHNX, KMUX and KVBX) exhibited slopes of α below zero, indicating overcompensation behaviours (figure 2; electronic supplementary material, figure S2). Although the magnitude of the offsets (track minus heading) at the three autumn locations were small (e.g. KHNX, KMUX and KVBX), they are consistent and may imply birds were correcting for previous drift or topographical effects [29]. However, local effects of winds related to variation in height, topography (e.g. orographic lift) and geography (e.g. distance from the Pacific) may be inherent sources of error in our methods, requiring more detailed analysis with higher resolution sensors (i.e. small-scale radar, local wind data).
(b). Differential migratory paths through western North America
There was 14.5% less migratory activity through the region in autumn compared to spring, suggesting that the Pacific Flyway is mainly used in spring. While a decrease in migratory activity is not immediately indicative of a looped pattern of migration, few other scenarios are plausible. Unless fewer birds were leaving their breeding grounds than arriving—a net decrease in population during the breeding season—migrants must be taking alternative, easterly or westerly, routes in autumn and thereby escaping detection by Pacific Flyway radars [16].
Surprisingly, spring wind patterns were in strong opposition to northward passage, particularly in regions with the greatest activity (i.e. Pacific southwest). This finding suggests that other factors are responsible for shaping this seasonally dependent migratory route (e.g. food resources) and not winds aloft, as they are in other systems (e.g. Central and Eastern Flyways) [30]. Contrary to other examinations [10,11], wind drift did not vary significantly across seasons, which may reflect first-year migrants using more eastern routes not captured by the 19 Pacific radars. A future investigation examining a larger set of radars, east of the Pacific Flyway, is needed to reveal the interconnectedness of these migratory routes. This result highlights the importance of full annual cycle monitoring and the utility of radar to capture large-scale population dynamics of migratory birds.
5. Conclusion
Flight strategies of migrating birds are fundamental components of their life histories, informed by endogenous (i.e. sensory and physiological) and exogenous (e.g. winds) cues that facilitate navigation and orientation required for their movements in the atmosphere. Radar, with a role as a remote sensing tool for understanding animal movements in the atmosphere, is critical for studying these patterns, particularly in understudied areas such as the Pacific Flyway. The integral ways in which birds and wind coexist at local and regional scales, in particular the variability in this coexistence, is only one part of a much larger system. Continental-scale characterizations of these behaviours will soon be a reality, linking diverse strategies of individuals and species with complex meteorological and climatological phenomena that can inform new perspectives about the efficiencies and evolution of migration itself. A critical understanding of what shapes these migration systems of which they are parts at many scales is more important than ever, given human alterations to ecosystems and climates and the position of bellwethers that birds occupy as proxies for environmental health.
Supplementary Material
Supplementary Material
Acknowledgements
We thank Jessie Barry, Chris Wood and Ian Davies for their role in organizing the Young Birder Event, providing a once-in-a-lifetime opportunity for young birders and connecting the authors of this paper. We thank Heiko Schmaljohann and two anonymous reviewers for constructive feedback on this manuscript.
Data accessibility
The dataset is provided as electronic supplementary material.
Authors' contributions
P.B.N., C.N., A.F. and K.G.H. worked to conceive, design and draft the manuscript. T.-Y.L., K.W., G.B., S.M., D.S. and K.G.H. designed radar algorithms, processed and summarized radar data. K.G.H. generated figures. All authors provided editorial advice, approved the final version of this manuscript and are in agreement to be accountable for all aspects of the work.
Competing interests
We declare we have no competing interests.
Funding
Funding for this project was provided by NSF (grant nos. IIS-1633206, DBI-1661329, DBI-1661259), Leon Levy Foundation and Rose Postdoctoral Fellowships.
References
- 1.Dokter AM, Farnsworth A, Fink D, Ruiz-Gutierrez V, Hochachka WM, Sorte FAL, Robinson OJ, Rosenberg KV, Kelling S. 2018. Seasonal abundance and survival of North America's migratory avifauna determined by weather radar. Nat. Ecol. Evol. 2, 1603–1609. ( 10.1038/s41559-018-0666-4) [DOI] [PubMed] [Google Scholar]
- 2.Hahn S, Bauer S, Liechti F. 2009. The natural link between Europe and Africa—2.1 billion birds on migration. Oikos 118, 624–626. ( 10.1111/j.1600-0706.2008.17309.x) [DOI] [Google Scholar]
- 3.Horton KG, Doren BMV, La Sorte FA, Cohen EB, Clipp HL, Buler JJ, Fink D, Kelly JF, Farnsworth A.. 2019. Holding steady: little change in intensity or timing of bird migration over the Gulf of Mexico. Glob. Change Biol. 25, 1106–1118. ( 10.1111/gcb.14540) [DOI] [PubMed] [Google Scholar]
- 4.Liechti F, Bauer S, Dhanjal-Adams KL, Emmenegger T, Zehtindjiev P, Hahn S. 2018. Miniaturized multi-sensor loggers provide new insight into year-round flight behaviour of small trans-Sahara avian migrants. Mov. Ecol. 6, 19 ( 10.1186/s40462-018-0137-1) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sjöberg S, et al. 2018. Barometer logging reveals new dimensions of individual songbird migration. J. Avian Biol. 49, e01821 ( 10.1111/jav.01821) [DOI] [Google Scholar]
- 6.Van Doren BM, Horton KG.. 2018. A continental system for forecasting bird migration. Science 361, 1115–1118. ( 10.1126/science.aat7526) [DOI] [PubMed] [Google Scholar]
- 7.Liechti F. 2006. Birds: blowin’ by the wind? J. Ornithol. 147, 202–211. ( 10.1007/s10336-006-0061-9) [DOI] [Google Scholar]
- 8.Alerstam T. 1979. Wind as selective agent in bird migration. Ornis Scand. 10, 76–93. ( 10.2307/3676347) [DOI] [Google Scholar]
- 9.Chapman JW, Klaassen RHG, Drake VA, Fossette S, Hays GC, Metcalfe JD, Reynolds AM, Reynolds DR, Alerstam T. 2011. Animal orientation strategies for movement in flows. Curr. Biol. 21, R861–R870. ( 10.1016/j.cub.2011.08.014) [DOI] [PubMed] [Google Scholar]
- 10.Horton KG, Van Doren BM, Stepanian PM, Farnsworth A, Kelly JF.. 2016. Seasonal differences in landbird migration strategies. Auk 133, 761–769. ( 10.1642/auk-16-105.1) [DOI] [Google Scholar]
- 11.Nilsson C, Klaassen RHG, Alerstam T. 2013. Differences in speed and duration of bird migration between spring and autumn. Am. Nat. 181, 837–845. ( 10.1086/670335) [DOI] [PubMed] [Google Scholar]
- 12.Horton KG, Van Doren BM, Stepanian PM, Hochachka WM, Farnsworth A, Kelly JF.. 2016. Nocturnally migrating songbirds drift when they can and compensate when they must. Sci. Rep. 6, 21249 ( 10.1038/srep21249) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Horton KG, Van Doren BM, La Sorte FA, Fink D, Sheldon D, Farnsworth A, Kelly JF.. 2018. Navigating north: how body mass and winds shape avian flight behaviours across a North American migratory flyway. Ecol. Lett. 21, 1055–1064. ( 10.1111/ele.12971) [DOI] [PubMed] [Google Scholar]
- 14.Carlisle JD, Skagen SK, Kus BE, Riper CV, Paxtons KL, Kelly JF. 2009. Landbird migration in the American west: recent progress and future research directions. Condor Ornithol. Appl. 111, 211–225. ( 10.1525/cond.2009.080096) [DOI] [Google Scholar]
- 15.La Sorte FA, Horton KG, Nilsson C, Dokter AM.. 2018. Projected changes in wind assistance under climate change for nocturnally migrating bird populations. Glob. Change Biol. 25, 589–601. ( 10.1111/gcb.14531) [DOI] [PubMed] [Google Scholar]
- 16.La Sorte FA, Fink D, Hochachka WM, DeLong JP, Kelling S.. 2014. Spring phenology of ecological productivity contributes to the use of looped migration strategies by birds. Proc. R. Soc. B 281, 20140984 ( 10.1098/rspb.2014.0984) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kelly JF, Horton KG. 2016. Toward a predictive macrosystems framework for migration ecology. Glob. Ecol. Biogeogr. 25, 1159–1165. ( 10.1111/geb.12473) [DOI] [Google Scholar]
- 18.Sheldon D. 2015. WSRLIB: MATLAB toolbox for weather surveillance radar. See http://bitbucket.org/dsheldon/wsrlib.
- 19.Farnsworth A, Van Doren BM, Hochachka WM, Sheldon D, Winner K, Irvine J, Geevarghese J, Kelling S.. 2016. A characterization of autumn nocturnal migration detected by weather surveillance radars in the northeastern US. Ecol. Appl. 26, 752–770. ( 10.1890/15-0023) [DOI] [PubMed] [Google Scholar]
- 20.Chilson PB, Frick WF, Stepanian PM, Shipley JR, Kunz TH, Kelly JF. 2012. Estimating animal densities in the aerosphere using weather radar: to Z or not to Z? Ecosphere 3, 72 ( 10.1890/ES12-00027.1) [DOI] [Google Scholar]
- 21.Browning KA, Wexler R. 1968. The determination of kinematic properties of a wind field using Doppler radar. J. Appl. Meteorol. 7, 105–113. () [DOI] [Google Scholar]
- 22.Sheldon D, Farnsworth A, Irvine J, Van Doren B, Webb K, Dietterich TG, Kelling S.. 2013. Approximate Bayesian inference for reconstructing velocities of migrating birds from weather radar. Association for the Advancement of Artificial Intelligence, pp. 1334–1340. [Google Scholar]
- 23.Lin T-Y, et al. 2019. MISTNET: measuring historical bird migration in the US using archived weather radar data and convolutional neural networks. Methods Ecol. Evol. 2041-210X.13280. ( 10.1111/2041-210X.13280) [DOI]
- 24.Mesinger F, et al. 2006. North American regional reanalysis. Bull. Am. Meteorol. Soc. 87, 343–360. ( 10.1175/BAMS-87-3-343) [DOI] [Google Scholar]
- 25.Larkin RP. 1991. Flight speeds observed with radar, a correction: slow ‘birds’ are insects. Behav. Ecol. Sociobiol. 29, 221–224. ( 10.1007/bf00166405) [DOI] [Google Scholar]
- 26.Wood S. 2015. mgcv: Mixed GAM Computation Vehicle with GCV/AIC/REML Smoothness Estimation. See http://cran.r-project.org/web/packages/mgcv/index.html.
- 27.Green M, Alerstam T. 2002. The problem of estimating wind drift in migrating birds. J. Theor. Biol. 218, 485–496. ( 10.1006/jtbi.2002.3094) [DOI] [PubMed] [Google Scholar]
- 28.Van Doren BM, Horton KG, Stepanian PM, Mizrahi DS, Farnsworth A.. 2016. Wind drift explains the reoriented morning flights of songbirds. Behav. Ecol. 27, 1122–1131. ( 10.1093/beheco/arw021) [DOI] [Google Scholar]
- 29.Klaassen RHG, Hake M, Strandberg R, Alerstam T. 2011. Geographical and temporal flexibility in the response to crosswinds by migrating raptors. Proc. R. Soc. B 278, 1339–1346. ( 10.1098/rspb.2010.2106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.La Sorte FA, et al. 2014. The role of atmospheric conditions in the seasonal dynamics of North American migration flyways. J. Biogeogr. 41, 1685–1696. ( 10.1111/jbi.12328) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The dataset is provided as electronic supplementary material.