

Abstract
Glial cell-line derived neurotrophic factor (GDNF) and its relative neurturin (NTN) are potent trophic factors for motoneurons. They exert their biological effects by activating the RET tyrosine kinase in the presence of a glycosyl-phosphatidylinositol-linked co-receptor, either GFRα1 or GFRα2. By whole-mount in situ hybridization on embryonic mouse spinal cord, we demonstrate that whereas Ret is expressed by nearly all motoneurons, Gfra1 and Gfra2 exhibit complex and distinct patterns of expression. Most motoneurons purified from Gfra1 null mutant mice had lost their responsiveness to both GDNF and NTN. However, a minority of them (∼25%) retained their ability to respond to both factors, perhaps because they express GFRα2. Surprisingly,Gfra2−/−motoneurons showed normal survival responses to both GDNF and NTN. Thus, GFRα1, but not GFRα2, is absolutely required for the survival response of a majority of motoneurons to both GDNF and NTN. In accordance with the phenotype of the mutant motoneurons observed in culture we found the loss of distinct groups of motoneurons, identified by several markers, in theGfra1−/− spinal cords but no gross defects in theGfra2−/− mutant. During their natural programmed cell death period, motoneurons in theGfra1−/− mutant mice undertook increased apoptosis. Taken together these findings support the existence of subpopulations of motoneuron with different trophic requirements, some of them being dependent on the GDNF family.
Keywords: motoneuron subpopulations, motoneuron survival, neurotrophic factors, GDNF, NTN, Ret, Gfra1, Gfra2, in situhybridization, mutant mice
Spinal motoneurons are organized into discrete longitudinal columns that contain pools of motoneurons distributed along the dorsoventral, rostrocaudal and mediolateral body axes according to their target muscles. In the embryo, motor columns can be distinguished by the combinatorial expression of members of a family of LIM homeodomain proteins, called the LIM code (Tsuchida et al., 1994). Recently, transcription factors belonging to the Ets family have been shown to be expressed by individual motor pools (Lin et al., 1998).
Because different subpopulations of motoneuron can also be distinguished by the receptors for neurotrophic factors they synthesize (Yamamoto et al., 1997), we hypothesized that these different subpopulations of motoneuron might have different trophic requirements. To test this hypothesis we focused on two members of the glial cell line-derived neurotrophic factor (GDNF) family: GDNF and neurturin (NTN). GDNF is the original member of a family distantly related to transforming growth factor-β, which at present includes three additional members: NTN, persephin (PSP), and artemin (ART) (Lin et al., 1993; Kotzbauer et al., 1996; Baloh et al., 1998; Milbrandt et al., 1998). Members of this family are potent survival factors for several populations of central and peripheral neurons. In particular, GDNF, NTN, and PSP have potent effects on the survival of motoneurons either in vivo or in vitro (Henderson et al., 1994; Oppenheim et al., 1995; Yan et al., 1995; Klein et al., 1997;Milbrandt et al., 1998).
The actions of GDNF and its relatives are mediated by a receptor complex consisting of the tyrosine kinase receptor RET and a ligand-binding glycosyl-phosphatidylinositol (GPI)-linked protein (GFRα). There are multiple GFRα proteins: GFRα1, 2, 3 and 4. In vitro experiments indicate that GFRα1 and 2 are the favored receptors for GDNF and NTN, respectively. Comparison of the phenotypes of mutant mice lacking either the ligand or the GFRα receptor highlighted the preferential interaction between GDNF and GFRα1 and NTN and GFRα2 (for review, see Airaksinen et al., 1999). However, at high concentrations GDNF can signal after binding to GFRα2 (Baloh et al., 1997; Jing et al., 1997; Sanicola et al., 1997), and NTN can bind to GFRα1 (Creedon et al., 1997; Jing et al., 1997). In accordance with the biological activities of GDNF and NTN,Gfra1 and 2 are widely distributed in both the PNS and CNS (Treanor et al., 1996; Baloh et al., 1997; Klein et al., 1997; Trupp et al., 1997; Widenfalk et al., 1997; Yu et al., 1998;Soler et al., 1999).
In the chick, Gfra receptors are expressed in different subpopulations of motoneuron (Soler et al., 1999) in accordance with the complementary and sometimes overlapping patterns of expression ofGfra1 and Gfra2 we have observed in the rat embryonic spinal cord (our unpublished observations). In particular, in the rat lumbar region there are subpopulations of motoneuron that express mRNA coding for one receptor but not for the other. These results raised the possibility that there might be two classes of lumbar motoneurons: one sustained by GDNF acting through GFRα1 and the other by NTN acting through GFRα2. The best way of testing this hypothesis is by genetic analysis in the mouse. We therefore chose to study the patterns of GDNF family receptor expression in mouse spinal cord and to analyze the effects of null mutations in Gfra1 and Gfra2 on survival of specific motoneuron subpopulations.
It has been indeed reported that Gdnf and Gfra1knock-out mice show a loss of 25% of motoneurons in lumbar spinal cord, but the missing population has not been identified (Moore et al., 1996; Sanchez et al., 1996; Cacalano et al., 1998). We show that GFRα1 is required for survival signaling by both GDNF and NTN in spinal motoneurons. Furthermore, prominent motoneuron groups that strongly express Gfra1 (but not Gfra2) are lost in theGfra1−/−mutant, whereas other motoneurons are apparently unaffected. Thus, GFRα1 is critical for the survival of specific groups of motoneuron during development, and different motoneurons have clearly different trophic requirements in vivo.
MATERIALS AND METHODS
Animals and genotype analysis. Embryos were collected from either wild-type or mutant mice. Vaginal positive plug was recorded as embryonic day 0.5 (E0.5).Gfra2−/−mice, previously described (Rossi et al., 1999), were bred as homozygous mice like C57Bl/6, their wild-type counterparts. Heterozygous Gfra1+/− mice were generously provided by A. Rosenthal (Genentech, South San Francisco). They were back-crossed to C57Bl/6 mice for at least four generations and were mated to obtain homozygous embryos; wild-type embryos from the same litters were used as controls.
Genotype analysis of the Gfra1 allele was performed using a previously described PCR technique (Arce et al., 1999). The primers CAGCTTCCTACCTAATCTG, GTTGTAGAGAGACTTCTGC and GGAGCAAAGCTGCTATTGG were used to amplify the targeted allele (422 bp band) and theGfra1 wild-type allele (342 bp band). When used for motoneuron purification, embryos were kept during genotyping at 4°C in Hybernate E medium supplemented with B-27 (Life Technologies, Cergy Pontoise, France).
Motoneuron purification and culture. Ventral spinal cords from E12.5 or E13.5 mouse embryos were dissected and dissociated, and motoneurons were isolated as described previously (Arce et al., 1999). Briefly, motoneurons were purified by a combination of metrizamide density-gradient centrifugation and indirect magnetic cell sorting with an antibody that recognizes the p75 low-affinity NGF receptor, a specific marker for motoneurons at this stage. A rat anti-mouse p75 purchased from Chemicon (Temecula, CA) was used. We used either E12.5 motoneurons purified by this procedure or E13.5 motoneurons purified only by a metrizamide density gradient. Identical results were obtained with motoneurons purified by both methods, irrespective of stages (data not shown). Purified motoneurons (800 or 1000 neurons per well) were plated in four-well tissue culture dishes (Nunc, Roskilde, Denmark). Wells had been previously coated with polyornithine/laminin (Henderson et al., 1995). Culture medium (basal medium) was Neurobasal, supplemented with the B-27 supplement (Life Technologies), horse serum (2% v/v), l-glutamine (0.5 mm), l-glutamate (25 μm), and 2-mercaptoethanol (25 μm).
To evaluate motoneuron survival after 3 d in culture, wells were filled with warm L-15 medium, and the cover of the dish was replaced. The number of large phase-bright neurons with long axonal processes was counted in duplicate either in the total area of the well or across two diameters of the well. To allow for comparison of values from different experiments or from different sets of embryo, survival values were corrected for the value in basal medium (taken as 0%) and expressed relative to the survival in 1 ng/ml BDNF (taken as 100%).
Neurotrophic factors. Recombinant neurotrophic factors were added at the time of seeding. Rat GDNF was purchased from Sigma (St. Louis, MO), and human BDNF was purchased from R & D Systems (Minneapolis, MN). Human NTN was either generously provided by Genentech or purchased from Peprotech (London, UK). Neurotrophic factors were prepared as stock solutions (1–100 μg/ml) in PBS supplemented with 0.5% BSA (Sigma) and kept in aliquots at −70°C. Once thawed, aliquots were kept at 4°C and used within 1 week.
Probes. Plasmid cDNA clones were linearized and transcribed with T7 or T3 polymerase using digoxigenin (DIG)-labeling reagents (Roche Diagnostics, Meylan, France). Probes were used at a concentration between 10 and 500 ng/ml. The Gfra1 andRet clones were as previously described (Arce et al., 1998). The rat Gfra2 probe corresponded to nt 1–1297 (GenBank accession number AF005226). The rat Islet1 probe (Pfaff et al., 1996) and a 600 bp fragment of the mouseRaldh2 cDNA were kindly provided by T. M. Jessell. The mouse EphA4 corresponded to nt 652–1834 and was generously given by P. Charnay.
In situ hybridization. Whole-mount in situhybridization (ISH) was performed as described by Henrique et al. (1995). Spinal cords were dissected, and fixed overnight at 4°C in 4% (w/v) paraformaldehyde in PBT (0.1% Tween 20 in PBS), progressively dehydrated in increasing concentrations of ethanol/PBT, stored at −20°C, then rehydrated in decreasing concentrations of ethanol/PBT and treated with proteinase K (10 μg/ml in PBS). They were then post-fixed for 20 min in 4% paraformaldehyde, 0.1% glutaraldehyde, and 0.1% Tween 20 and prehybridized 1 hr at 70°C in 1.3× SSC, 50% formamide, 2% Tween 20, 0.5% 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonic acid, 5 mm EDTA, and 50 μg/ml yeast RNA. Hybridization was performed overnight with DIG-labeled riboprobes in the same buffer. Washes with hybridization buffer were followed by RNase A treatment (10 μg/ml in 0.5 m NaCl, 10 mm Tris, pH 7.5, and 0.1% Tween 20, 1 hr at 37°C), and subsequent washes with hybridization buffer at 65°C. Spinal cords were then blocked in MABT (0.1 mmaleate, 0.15 m NaCl, and 0.1% Tween 20, pH 7.5) containing 20% sheep serum, and incubated overnight at 4°C with anti-DIG-alkaline phosphatase (AP)-conjugate (Roche Diagnostics) diluted 1: 2000 in MABT with 2% sheep serum. After extensive washes with MABT, revelation was performed using nitro blue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate (Roche Diagnostics) in 0.1m NaCl, 50 mmMgCl2, 0.1% Tween 20, and 0.1m Tris pH 9.5. After staining, the spinal cords were flat-mounted as open-book preparations in 80% glycerol and examined by at least two observers to whom the genotype of the embryos was unknown. Some preparations were subsequently embedded in 30% albumin, 0.5% gelatin, and 1.5% glutaraldehyde in 0.12m phosphate buffer, and 30 μm sections were cut with a Leica (Nussloch, Germany) vibratome.
Whole-mount terminal deoxynucleotidyl transferase-mediated biotinylated dUTP nick end labeling. Whole-mount terminal deoxynucleotidyl transferase-mediated biotinylated dUTP nick end labeling (TUNEL) and double TUNEL/Islet labeling were performed essentially as described by Yamamoto and Henderson (1999). Spinal cords were fixed, dehydrated, and rehydrated through graded ethanol concentrations into PBS and then stained with the ApopTag kit (Oncor). Briefly, spinal cords were incubated in ApopTag equilibration buffer for >5 min at room temperature and transferred to the working strength TdT enzyme solution for 12 hr at 4°C, followed by 2 hr at 37°C. The reaction was stopped by incubating the spinal cords in ApopTag stop solution for 40 min at 37°C. After washing in TBST (0.14m NaCl, 10 mm KCl, 25 mm Tris, pH 7.0, and 0.1% Tween 20), endogenous AP was inactivated by incubating the spinal cords in TBST for 20 min at 65°C. The spinal cords were then incubated in blocking solution (10% goat serum, 1% BSA in PBS) followed by incubation with anti-DIG-AP conjugate (Roche Diagnostics; diluted 1:2000 in blocking solution) overnight at 30°C. They were then extensively washed in MABT, stained as described for the whole-mount ISH, and examined under transillumination. For double TUNEL/Islet staining, spinal cords were blocked and then incubated with biotin-SP-conjugated anti-digoxigenin antibodies (Jackson ImmunoResearch, West Grove, PA) diluted; 1:1000) and Islet 1/2 monoclonal antibodies 2D6 and 4D5 (Developmental Hybridoma Bank, Baltimore, MD). After several washes, the spinal cords were incubated with Cy3-conjugated anti mouse antibodies (Jackson ImmunoResearch; 1:1000) and Cy2-conjugated streptavidin (Amersham Pharmacia Biotech, Orsay, France; 1:500) and washed with MABT. The spinal cords were flattened into open-book preparations and analyzed with a Zeiss LSM 400 confocal microscope. At least two spinal cords from each genotype were examined for each stage. For each analysis, wild-type and mutant tissues were dissected from embryos belonging to the same littermate and processed in parallel.
RESULTS
Distinct expression patterns of Gfra1 andGfra2 within the mouse spinal cord
To get an overall view of the expression of GDNF family receptor components in different subpopulations of mouse motoneuron, whole-mount ISH was performed on predissected E13.5 or E14.5 spinal cords usingRet, Gfra1, and Gfra2 probes. Spinal cords were then flattened into an “open-book” configuration, in which the ventral midline lies medial and the dorsal edges of the neural tube lie lateral. This allows the complex array of motor columns and pools to be visualized in the stereomicroscope. Different regions of the spinal cord were then examined. Identical results were obtained at both stages, thus only E13.5 stages are presented.
Within the brachial region, most motor columns expressedIslet1, Ret, and Gfra1 (Fig.1A–C). The expression of Gfra2 was more restricted and found only in some columns. The most lateral one was the most widespread along the rostrocaudal axis (Figs. 1D,2C, arrows) and was one of the columns positive for Islet1, Ret, andGfra1 (Figs. 1A–C,2A,B). Because the whole-mount spinal cords were flattened, some columns appearing medial are actually lateral columns. This is the case for the Gfra2-positive column that was positive for Ret and Islet2 but negative for Islet1 with few cells positive forGfra1 (Figs. 1A–D, yellow lines, 2A–C, circle; data not shown). Immediately caudal to this column was another column that was negative forGfra2 but positive for the other probes (Figs.1A–D, white lines, 2D–F, circle).
Fig. 1.

Distinct expression patterns ofGfra1 and Gfra2 in the E13.5 mouse spinal cord. Whole-mount ISH performed using probes to Islet1(A, E), Ret (B, F),Gfra1 (C, G), or Gfra2(D, H) on brachial (A-D) and lumbar (E-H) regions shows thatRet is widely expressed, whereas the GFRα-subunits are expressed in different subpopulations of motoneuron. The arrangement of the different motor columns is shown: circled inwhite are the groups of motoneurons that strongly express Gfra1, and in yellow are those that expresses Gfra2.Arrows point to motoneurons that express both Gfra1 andGfra2. The white-circled column belongs to lateral motor column (LMC); however, flattening of the cord in this preparation leads to it appearing close to the midline.
In the thoracic region, expression of Ret, Gfra,1and Gfra2 was detected in the median motor column (MMC) (Fig. 1B–D), which can be further subdivided into lateral and medial columns (MMCl and MMCm, respectively). A careful analysis of sections from whole-mount spinal cord revealed that both columns were stained using the Gfra1 and the Retprobes, whereas only very few cells in the MMCl were positive forGfra2 (Fig. 2G–I). Comparison of the staining observed with the Ret probe (Fig. 2G) and the Gfra1 probe (Fig. 2H) revealed that only part of the MMCl was positive for Gfra1.
Fig. 2.
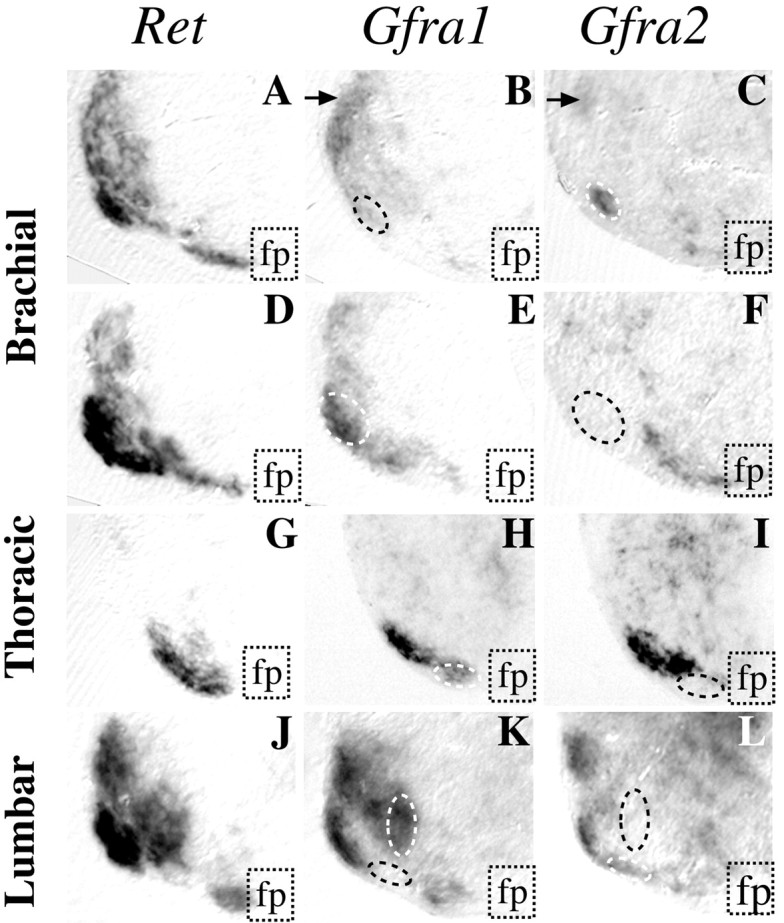
Gfra1 is more widely expressed in motoneurons than Gfra2. Whole-mount ISH was performed on E13.5 spinal cords that were subsequently sectioned. Transverse sections were performed at brachial (A-F), thoracic (G–I), or lumbar (J–L) levels. In the brachial region sections were performed either at the level of the yellow circle (A–C) or thewhite circle (D–F) drawn on Figure 1. Panels show half-ventral horn of each section;fp indicates the floor plate. Nearly all motoneurons are stained using a Ret probe (A, D, G, andJ), whereas a Gfra1 probe (B, E, H, and K) stained more motoneurons than hybridization using a Gfra2 probe (C, F, I, and L). Arrowsin B and C indicate the sameGfra1- and Gfra2-positive column as in Figure 1, C and D. Dashed lines delineate motoneuron groups that are positive for only one α-receptor.
It was more difficult to analyze the lumbar motor columns because of their relatively compact organization in the mouse. However, whereasRet was expressed in most motoneurons (Fig.1F), the pattern of expression of Gfra1and Gfra2 was different (Fig. 1G,H). For example, a group of motoneurons localized in the rostral part of the lumbar region were positive for Gfra1, Ret, andIslet1 (Fig. 1E–G, white lines) and negative for Gfra2 (Fig. 1H). Sections performed at the lumbar spinal cord levels after whole-mount in situ hybridization confirmed the distinct patterns of expression of Gfra1 and Gfra2 (Fig. 2K,L). In the mouse, Gfra2 was expressed in fewer motoneurons than was Gfra1 (Fig. 2, compare K, L). However, some groups of motoneurons were positive for Gfra2and negative for Gfra1 (Fig. 2K,L). In conclusion, in the mouse spinal cord, most motoneurons expressGfra1, some of which are also positive for Gfra2, whereas few motoneurons express only Gfra2.
Differential responses of Gfra1 orGfra2 null mutant motoneurons to GDNF and NTN
The distinct patterns of expression of Gfra1 andGfra2 within the spinal cord raised the possibility that these receptor components might have distinct roles in motoneuron development. Because GDNF and NTN both have survival activity on motoneurons, we first examined the ability of mutant motoneurons lacking either GFRα1 or GFRα2 to respond to GDNF and NTN. GFRα2 is the favored receptor for NTN, and so we studied the NTN responsiveness in vitro of motoneurons purified from total spinal cords of E12.5 or E13.5 Gfra2 null mutant (Gfra2−/−) mice. Surprisingly, in three independent experiments, motoneurons lacking Gfra2 retained their responsiveness to NTN and GDNF at both low and high concentrations (Fig.3A), suggesting that in motoneurons, survival responses are not mediated by GFRα2.
Fig. 3.

Survival of embryonic motoneurons from wild-type,Gfra2−/−, andGfra1−/− mice in the presence of GDNF and NTN. A, Survival of motoneurons from wild-type (WT) andGfra2−/− E12.5 embryos in the presence of different concentrations of GDNF and NTN.B, Survival of motoneurons from wild-type (WT) andGfra1−/− E13.5 embryos in the presence of different concentrations of GDNF and NTN. Survival values (mean ± SEM; n = 3) are normalized for survival in basal medium [Neurobasal (NB) defined as 0%] and expressed for each region as percentage of the number of motoneurons that developed in 1 ng/ml BDNF (defined as 100%).
To identify the GFRα receptor essential for the GDNF and NTN survival responses, we next studied knock-out mice deficient for GFRα1 (Gfra1−/−). In three independent experiments, motoneurons from E13.5Gfra1−/−mutants retained normal responsiveness to BDNF but no longer responded to low concentrations of GDNF (≤0.1 ng/ml) (Fig. 3B), whereas the same concentration of GDNF (0.1 ng/ml) prevented the death of 75 ± 2.6% of motoneurons purified from wild-type littermates. More surprisingly,Gfra1−/−motoneurons were also less responsive to NTN. Only 24 ± 3% of the Gfra1 null mutant motoneurons were rescued by 1 ng/ml NTN, and this figure did not increase with higher concentrations of NTN, although in the same experiment 1 ng/ml NTN saved 80 ± 2% of wild-type motoneurons (Fig. 3B). At higher concentrations (1 and 10 ng/ml) GDNF rescued 27 ± 3.5% and 35 ± 1% of mutant motoneurons, respectively, compared to 98 ± 0.1% and 74 ± 17% of wild-type motoneurons. We presumed thatGfra1−/−motoneurons kept alive by GDNF and NTN expressed GFRα2. Our results identify GFRα1 as the principal survival co-receptor for GDNF and NTN.
Loss of specific subpopulations of motoneurons in theGfra1 knock-out mice
Our in vitro results showing that GFRα2 was not necessary to induce motoneuron survival in the presence of GDNF or NTN corroborated the absence of gross abnormalities observed in theGfra2−/−spinal cords (Rossi et al., 1999). Likewise, the absolute requirement for GFRα1 in the survival activity of GDNF on a subpopulation of motoneurons in culture was in accordance with the loss of motoneurons observed in the spinal cord of mutant mice lacking Gfra1(Cacalano et al., 1998). Thus, we sought to characterize motoneuron subpopulations potentially lost inGfra1−/−andGfra2−/−mutant mice. Spinal cords were dissected from wild-type,Gfra1−/−, andGfra2−/−embryos at E15.5, the end of the programmed cell death (PCD) period in mouse motoneurons (Yamamoto and Henderson, 1999). To get an overall view of the motoneuron organization in these mice, we first used ISH on whole-mount spinal cord using a probe to the motoneuron markerIslet1. At the brachial level (Fig.4A–C), comparison of mutant and wild-type spinal cord staining clearly showed that a group of motoneurons was present in the wild-type and theGfra2−/−samples (Fig. 4A,B) but was missing in theGfra1−/−mutant (Fig. 4C). This group of motoneurons belonged to the lateral motor column (LMC) and could be easily distinguished in whole-mount spinal cords by its characteristic position in the brachial swelling of the neural tube. It corresponds to the column shown to be strongly positive for Gfra1 expression but negative forGfra2 (Fig. 1C,D, white lines). This was further confirmed using probes to ChAT (data not shown) and toRet: wild-type andGfra2−/−spinal cords showed similar labeling patterns (Fig.4D,E), but the same column was missing inGfra1−/−spinal cords (Fig. 4F, orange lines).
Fig. 4.
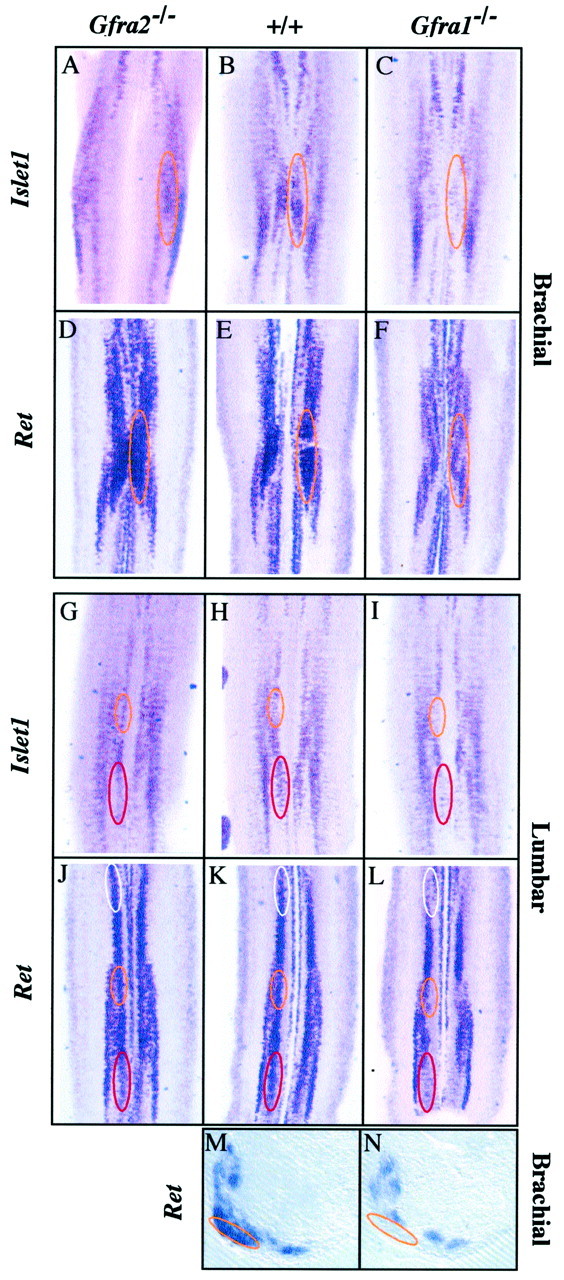
Loss of motoneurons inGfra1−/− spinal cords. Whole-mount ISH was performed using probes toIslet1 (A–C andG–I) or Ret (D–Fand J–L) on E15.5 spinal cords from wild-type (B, E, H, and K),Gfra1−/−(C, F, I, and L), orGfra2−/−(A, D, G, and J) mice. Analysis of brachial (A–F) and lumbar (G–L) regions shows that several groups of motoneuron are missing in theGfra1−/− mutant mice (orange, red, and white lines). At the brachial level the Ret staining apparent within theorange lines corresponds toIslet1-negative motoneurons present on a different focal plane within the whole-mount views. The negative column is localized in the brachial swelling that corresponds to LMC; this has been confirmed by transverse sections of whole-mount ISH that clearly show that the lateral motor column circled orange in the wild-type (M) is missing in the Gfra1−/−mutant mice (N). Only partial loss (red lines) is observed in theGfra2−/− lumbar spinal cords. Note that in A, the most lateral part of the object represents the lateral limit of the ventral horn. Each panel is representative of at least four spinal cords.
In the lumbar region, Islet1 and Ret staining revealed the possible loss of three groups of motoneurons in theGfra1−/−mutant (Fig. 4I,L) compared to their wild-type counterparts (Fig. 4H,K). In theGfra2−/−mutant we observed a small decrease in the expression ofIslet1 and Ret in only one of these groups of motoneuron (Fig. 4G,J, red lines).
Because several groups of motoneuron are no longer detectable in theGfra1−/−spinal cords but only a few groups of motoneuron are missing in theGfra2−/−spinal cords, we further focused on theGfra1−/−mutant mice. This possible motoneuron loss in this mutant was studied by ISH using markers of subpopulation of motoneurons. We used two probes that have been shown to be markers of chick LMC motoneurons localized in the limb-innervating regions: EphA4 (Ohta et al., 1996) and Raldh2 (Zhao et al., 1996; Sockanathan and Jessell, 1998). In both brachial and lumbar regions, the number ofEphA4-expressing motoneurons was dramatically reduced in theGfra1−/−mutant compared to the wild-type (Fig.5A–D). Similar results were obtained after staining using the Raldh2 probe. In the brachial region, the motoneurons stained by Raldh2 that appear the most medial in this view (Fig. 5E) were absent in theGfra1−/−mutant (Fig. 5F). In the lumbar region, a group of motoneurons localized in the rostral part (Fig. 5G) was lost in theGfra1−/−mutant (Fig. 5H), whereas the level of expression ofRaldh2 decreased in a group of motoneurons that appeared more caudal (Fig. 5G,H).
Fig. 5.

EphA4- and Raldh2- expressing motoneurons are lost inGfra1−/− mutant mice. Whole-mount ISH was performed using probes toEphA4 (A–D) orRaldh2 (E–H) on E15.5 spinal cords from wild-type (A, E, C, and G) orGfra1−/−(B, F, D, and H) mice. Analysis of brachial (A, B and E, F) and lumbar (C, D and G, H) regions shows that the number of motoneurons expressing EphA4decreased in the mutant as compared to the wild-type (arrows). Raldh2 expression is greatly reduced in several groups of motoneuron in theGfra1−/− mutant mice (rostral, dashed lines), and the staining is also decreased in a further group of motoneurons (caudal, dashed lines). Each panel is representative of at least four spinal cords.
Increased apoptosis of motoneurons lost in theGfra1−/− mice
The absence of motoneurons in theGfra1−/−mutant spinal cord suggested that they had died owing to the loss of GDNF or NTN signaling. At E13.5, the moment of the naturally occurring motoneuron cell death (Yamamoto and Henderson, 1999), TUNEL-positive cells were apparent in wild-type spinal cord (Fig.6A). InGfra1−/−spinal cords the number of TUNEL-positive cells increased dramatically in the lumbar region (Fig. 6B). At E14.5, wild-type PCD had decreased as compared to E13.5 (Fig. 6A,C) but in the mutant, TUNEL staining was still higher than in the wild-type (Fig. 6C,D). At E15.5, when normal motoneuron PCD is nearly complete (Yamamoto and Henderson, 1999), increased TUNEL staining was no longer apparent in the mutant (data not shown). We also did not observe any significant changes in the number of apoptotic cells in the brachial region from E12.5 to E15.5 (data not shown). To identify the cells dying by apoptosis, spinal cords were double-stained with TUNEL and antibodies to Islet1/2. Confocal analysis of these preparations showed that, in both wild-type and mutant spinal cords, most TUNEL-positive nuclei represent apoptotic motoneurons (Fig.6E–H). Therefore, the loss of lumbar motoneurons observed in the Gfra1 mutants could be attributed, at least in part, to an increase in cell death during the period of motoneuron PCD.
Fig. 6.
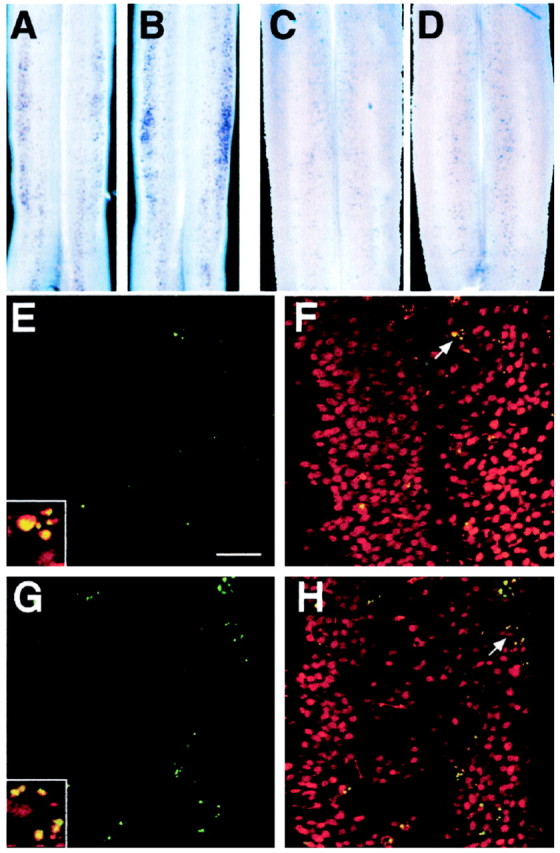
Increased apoptotic cell death of motoneurons in the Gfra1−/−mutant detected by whole-mount TUNEL labeling. Top panels, Lumbar spinal cords from wild-type (A, C) orGfra1−/−(B, D) mice from stage E13.5 (A, B) or E14.5 (C, D). Note the increased intensity of staining in mutant spinal cords as compared to wild-type. Because inA and B spinal cords are not completely flattened, the most lateral part of the image in A andB represents the lateral limit of the ventral horn, but in C and D represents the dorsal limit of the spinal cord. Middle and bottom panels, Confocal micrographs of double-labeled lumbar spinal cord preparations from wild-type (E, F) orGfra1−/−(G, H) E13.5 embryos stained by TUNEL (ingreen) and antibodies to Islet 1/2 (inred). TUNEL-positive nuclei are more numerous in the mutant (G) than in the wild type (E). Superposition of TUNEL and Islet1/2 fluorescence images (F, H, and insets) shows that nearly all TUNEL-positive nuclei represent apoptotic motoneurons. Scale bar (for E–H), 50 μm.
DISCUSSION
We have shown that subpopulations of spinal motoneuron can be distinguished by the combinatorial expression of the ligand-binding subunits GFRα1 and GFRα2, at the stage at which their programmed cell death is about to begin. Contrary to expectation, analysis of mutant mice in which Gfra1 has been deleted clearly showed that GFRα1 is required for survival activity of both GDNF and NTN on most purified motoneurons. Accordingly, groups of motoneuron that strongly express Gfra1 but not Gfra2 identified by specific independent markers are missing in theGfra1−/−mice. This loss, at least in part, reflects an increase in cell death during the period of naturally occurring PCD. These findings substantiate the hypothesis that there are different subpopulations of motoneuron with different trophic requirements and demonstrate that GFRα1 signaling plays a vital role in the development of specific motoneuron pools.
Distinct expression patterns of Gfra1 andGfra2 in spinal motoneurons
As a first step to gain insight into the role of GDNF and NTN on motoneuron development, we studied the expression of their receptor complex components in normal mice. To get an overview of the spinal cord and to better distinguish the organization of motoneurons expressing these receptors, we used whole-mount ISH on predissected spinal cord, which provides complete and detailed information on the receptor expression patterns. Within the mouse spinal cord, most motoneurons express Gfra1, and some are also positive forGfra2, whereas few motoneurons express onlyGfra2. Distinct patterns of expression of Gfra1and Gfra2 have already been observed in the nervous system or in developing organs (Treanor et al., 1996; Baloh et al., 1997; Jing et al., 1997; Klein et al., 1997; Trupp et al., 1997; Widenfalk et al., 1997; Yu et al., 1998; Soler et al., 1999). However, our results highlight the heterogeneity within a single type of central neurons and strengthen parallels between motoneuron subpopulations and those already described in peripheral sensory ganglia (for review, see Snider and Wright, 1996; Davies, 1997).
During the period of motoneuron cell death, GDNF and NTN are expressed in the environment of motoneurons. GDNF is expressed by floor-plate, limb bud mesenchyme, and by Schwann cells and, at later stages, by some muscles (Henderson et al., 1994; Sanchez et al., 1996; Wright and Snider, 1996; Golden et al., 1999; our unpublished results). NTN has been found in limb bud and skeletal muscles (Golden et al., 1999). Because different subpopulations of motoneuron can be distinguished by the Gfra receptor they express, we assessed whether these different subpopulations might exhibit different responsiveness to GDNF or NTN, and we determined which receptors are involved in the survival response to which factor. To this end, we studied the responsiveness to GDNF and NTN of motoneurons purified from mutant mice in whichGfra1 or Gfra2 had been deleted.
GFRα2 is not essential for motoneuron survival
Surprisingly, motoneurons lacking Gfra2 respond equally well in culture to GDNF and NTN. This result suggests that in motoneurons both GDNF and NTN signal for survival mainly through the complexes formed by RET and GFRα1 receptor. This is in accordance with the report that RET-GFRα1-expressing fibroblasts respond similarly to both GDNF and NTN (Baloh et al., 1997). One might predict that motoneurons that normally express Gfra2 but notGfra1 should not respond to GDNF or NTN when cultured fromGfra2−/−embryos. However, their total number is probably too small to be detected in our survival assay, and Gfra1 may be upregulatedin vitro in the presence of GDNF or NTN. Alternatively, at the stage of motoneuron purification, motoneurons dependent on GFRα2 for their survival are already lost. This is unlikely because there are no gross defects in spinal motoneurons in these mutants (Rossi et al., 1999; our results). In addition, we cannot rule out the role of another GFRα family member, GFRα4, for example (Thompson et al., 1998).
The normal survival response of motoneurons lacking Gfra2 in vitro is consistent with the absence of gross motoneuron defects inGfra2−/−orNtn−/−mice (Airaksinen et al., 1999; Heuckeroth et al., 1999). However, the possibility of minor defects cannot be ruled out as demonstrated by the decrease in Islet1 and Ret expression in some lumbar motoneurons ofGfra2−/−mice.
GFRα1 is required for development of distinct subpopulations of motoneuron
Most of the motoneurons purified from mice lackingGfra1 did not respond to GDNF or NTN in vitro. Thus, in the majority of motoneurons isolated by our purification procedure, GFRα1 is absolutely required for the survival activity of either GDNF or NTN. NTN and GDNF rescued ∼25% of theGfra1−/−motoneurons, so this subpopulation of motoneurons might express GFRα2. Altogether these results suggest a lack of α-receptor/ligand specificity in purified motoneurons in contrast to the in vivo specific α-receptor/ligand interaction observed in DRG neurons and in other neurons (Leitner et al., 1999; for review, seeAiraksinen et al., 1999). In addition, the low percentage of motoneurons expressing Gfra2 observed by in situhybridization could be in accordance with the low percentage (∼ 25%) ofGfra1−/−motoneurons saved by NTN or GDNF.
Our results with motoneurons differ from those obtained using dopaminergic and nodose neurons isolated fromGfra1−/− mice. E12Gfra1−/− nodose neurons do not respond to GDNF even at high concentrations, even though these neurons express Gfra2 at E15 and E18 (M. S. Airaksinen, unpublished observations). It will be important to determine whetherGfra2 is expressed byGfra1−/− nodose neurons at E12.Gfra1−/− dopaminergic neurons do not respond to either GDNF or NTN whatever the concentration (Cacalano et al., 1998), in accordance with the lack of expression ofGfra2 in these neurons (Horger et al., 1998). Motoneurons are thus the only neurons described, which, in the absence of GFRα1, retain a partial responsiveness to both GDNF and NTN.
In vivo, mice lacking Gdnf or Gfra1show a significant loss of lumbar motoneurons (Moore et al., 1996;Sanchez et al., 1996; Cacalano et al., 1998; Oppenheim et al., 2000) in accordance with our results showing that Gfra1 is required for GDNF survival effects. Nevertheless, ∼75% of the motoneurons survive at birth in contrast to the ∼75% of the motoneurons, which die in vitro in the presence of GDNF alone. Several hypotheses might be put forward to explain this observation: (1) motoneurons still alive inGfra1−/− mice expressGfra2 and are saved by NTN. However, the fact that only 25% of purified motoneurons from theGfra1−/− mice were saved by NTN does not argue for this hypothesis; (2) although in wild-type mice <75% of motoneurons express Gfra2, in vivo some GFRα2 molecules can act in “trans” in knock-out mice (Trupp et al., 1997; Yu et al., 1998); (3) in vivo neurotrophic factors from other families may also compensate.
To characterize the motoneurons lost in the mutant we performed whole-mount ISH using several markers. This study revealed that groups of motoneuron expressing several markers such as ChAT,Islet1, Ret, EphA4, andRaldh2 are lacking in the mutant. Although we cannot completely exclude the possibility that the lack of these markers in the mutant is attributable to their downregulation in the absence of GDNF signaling, this is unlikely because this implicates all these markers in the same GDNF-dependent pathway. The lack of some subpopulations of motoneuron in the mutant might be attributable to an increase in apoptotic death, as suggested by the increased levels of TUNEL labeling in lumbar regions of E13.5 and E14.5 mutant spinal cords. Our double-staining experiments identifying dying cells as motoneurons are in accordance with the increased number of pyknotic motoneurons observed by Oppenheim et al. (2000) during the PCD inGdnf−/−mice. This observation indicates that the loss of motoneurons inGfra1−/−spinal cord takes place during the period of PCD. Altogether these observations implicate GDNF signaling through GFRα1 as a major actor in survival of subpopulations of motoneuron, especially in lumbar spinal cord. The absence of increased TUNEL staining in the brachial regions of mutant spinal cords where motoneurons are clearly missing raises an additional hypothesis that GDNF signaling is a critical factor for earlier motoneuron development processes. Further studies are needed to explore this possibility.
Conclusion
We have shown that Gfra1 and Gfra2 exhibit complex expression patterns in mouse spinal cord. In vitroand in vivo GFRα1 is necessary for the survival of subpopulations of motoneuron as demonstrated using purifiedGfra1−/−motoneurons and by observation of increased cell death during the period of PCD in theGfra1−/−mice. The absence of distinct groups of motoneuron expressing several markers in theGfra1−/−mice demonstrates for the first time that signaling through a neurotrophic factor receptor is absolutely required for the development of specific subpopulations of motoneuron.
Footnotes
This work was supported by Institut National de la Santé et de la Recherche Médicale (INSERM), Centre National de la Recherche Scientifique (CNRS), the Association Française contre les Myopathies (AFM), the Institut pour la Recherche sur la Moelle Epinière (IRME), Academy of Finland, and European Community contract CT960433. A.G. and J.L. were supported by French Ministère de la Recherche et de la Technologie, and G.H. by AFM and the Fondation pour la Recherche Médicale. A.G. was further supported by Association de Recherche contre le Cancer. We thank Chris Henderson and members of INSERM U.382 for many helpful discussions and encouraging support. We acknowledge generous gifts ofGfra1+/− mice from A. Rosenthal (Genentech, San Francisco, CA) and A. Davies (University of St. Andrews).
Correspondence should be addressed to O. deLapeyrière, Institut National de la Santé et de la Recherche Médicale U.382, Institut de Biologie du Développement de Marseille, Campus de Luminy, Case 907, 13288 Marseille Cedex 09, France. E-mail:delapeyr@ibdm.univ-mrs.fr.
REFERENCES
- 1.Airaksinen MS, Titievsky A, Saarma M. GDNF family neurotrophic factor signaling: four masters, one servant. Mol Cell Neurosci. 1999;13:313–325. doi: 10.1006/mcne.1999.0754. [DOI] [PubMed] [Google Scholar]
- 2.Arce V, Pollock RA, Philippe JM, Pennica D, Henderson CE, deLapeyrière O. Synergistic effects of Schwann- and muscle-derived factors on motoneuron survival involve GDNF and cardiotrophin-1 (CT-1). J Neurosci. 1998;18:1440–1448. doi: 10.1523/JNEUROSCI.18-04-01440.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Arce V, Garces A, de Bovis B, Filippi P, Henderson C, Pettmann B, deLapeyrière O. Cardiotrophin-1 requires LIFRbeta to promote survival of mouse motoneurons purified by a novel technique. J Neurosci Res. 1999;55:119–126. doi: 10.1002/(SICI)1097-4547(19990101)55:1<119::AID-JNR13>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 4.Baloh RH, Tansey MG, Golden JP, Creedon DJ, Heuckeroth RO, Keck CL, Zimonjic DB, Popescu NC, Johnson EM, Jr, Milbrandt J. TrnR2, a novel receptor that mediates neurturin and GDNF signaling through Ret. Neuron. 1997;18:793–802. doi: 10.1016/s0896-6273(00)80318-9. [DOI] [PubMed] [Google Scholar]
- 5.Baloh RH, Tansey MG, Lampe PA, Fahrner TJ, Enomoto H, Simburger KS, Leitner ML, Araki T, Johnson EM, Jr, Milbrandt J. Artemin, a novel member of the GDNF ligand family, supports peripheral and central neurons and signals through the GFRalpha3-RET receptor complex. Neuron. 1998;21:1291–1302. doi: 10.1016/s0896-6273(00)80649-2. [DOI] [PubMed] [Google Scholar]
- 6.Cacalano G, Farinas I, Wang LC, Hagler K, Forgie A, Moore M, Armanini M, Phillips H, Ryan AM, Reichardt LF, Hynes M, Davies A, Rosenthal A. GFRalpha1 is an essential receptor component for GDNF in the developing nervous system and kidney. Neuron. 1998;21:53–62. doi: 10.1016/s0896-6273(00)80514-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Creedon DJ, Tansey MG, Baloh RH, Osborne PA, Lampe PA, Fahrner TJ, Heuckeroth RO, Milbrandt J, Johnson EM., Jr Neurturin shares receptors and signal transduction pathways with glial cell line-derived neurotrophic factor in sympathetic neurons. Proc Natl Acad Sci USA. 1997;94:7018–7023. doi: 10.1073/pnas.94.13.7018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Davies AM. Neurotrophin switching: where does it stand? Curr Opin Neurobiol. 1997;7:110–118. doi: 10.1016/s0959-4388(97)80128-6. [DOI] [PubMed] [Google Scholar]
- 9.Golden JP, DeMaro JA, Osborne PA, Milbrandt J, Johnson EM., Jr Expression of neurturin, GDNF, and GDNF family-receptor mRNA in the developing and mature mouse. Exp Neurol. 1999;158:504–528. doi: 10.1006/exnr.1999.7127. [DOI] [PubMed] [Google Scholar]
- 10.Henderson CE, Phillips HS, Pollock RA, Davies AM, Lemeulle C, Armanini M, Simpson LC, Moffet B, Vandlen RA, Koliatsos VE, Rosenthal A. GDNF: a potent survival factor for motoneurons present in peripheral nerve and muscle. Science. 1994;266:1062–1064. doi: 10.1126/science.7973664. [DOI] [PubMed] [Google Scholar]
- 11.Henderson CE, Bloch-Gallego E, Camu W. Purified embryonic motoneurons In: Nerve cell culture: a practical approach (Cohen J, Wilkin G, eds), pp 69–81. Oxford UP; London: 1995. [Google Scholar]
- 12.Henrique D, Adam J, Myat A, Chitnis A, Lewis J, Ish-Horowicz D. Expression of a Delta homologue in prospective neurons in the chick. Nature. 1995;375:787–790. doi: 10.1038/375787a0. [DOI] [PubMed] [Google Scholar]
- 13.Heuckeroth RO, Enomoto H, Grider JR, Golden JP, Hanke JA, Jackman A, Molliver DC, Bardgett ME, Snider WD, Johnson EM, Jr, Milbrandt J. Gene targeting reveals a critical role for neurturin in the development and maintenance of enteric, sensory, and parasympathetic neurons. Neuron. 1999;22:253–263. doi: 10.1016/s0896-6273(00)81087-9. [DOI] [PubMed] [Google Scholar]
- 14.Horger BA, Nishimura MC, Armanini MP, Wang LC, Poulsen KT, Rosenblad C, Kirik D, Moffat B, Simmons L, Johnson EM, Jr, Milbrandt J, Rosenthal A, Bjorklund A, Vandlen RA, Hynes MA, Phillips HS. Neurturin exerts potent actions on survival and function of midbrain dopaminergic neurons. J Neurosci. 1998;18:4929–4937. doi: 10.1523/JNEUROSCI.18-13-04929.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jing S, Yu Y, Fang M, Hu Z, Holst PL, Boone T, Delaney J, Schultz H, Zhou R, Fox GM. GFRalpha-2 and GFRalpha-3 are two new receptors for ligands of the GDNF family. J Biol Chem. 1997;272:33111–33117. doi: 10.1074/jbc.272.52.33111. [DOI] [PubMed] [Google Scholar]
- 16.Klein RD, Sherman D, Ho WH, Stone D, Bennett GL, Moffat B, Vandlen R, Simmons L, Gu Q, Hongo JA, Devaux B, Poulsen K, Armanini M, Nozaki C, Asai N, Goddard A, Phillips H, Henderson CE, Takahashi M, Rosenthal A. A GPI-linked protein that interacts with Ret to form a candidate neurturin receptor. Nature. 1997;387:717–721. doi: 10.1038/42722. [DOI] [PubMed] [Google Scholar]
- 17.Kotzbauer PT, Lampe PA, Heuckeroth RO, Golden JP, Creedon DJ, Johnson EM, Jr, Milbrandt J. Neurturin, a relative of glial-cell-line-derived neurotrophic factor. Nature. 1996;384:467–70. doi: 10.1038/384467a0. [DOI] [PubMed] [Google Scholar]
- 18.Leitner ML, Molliver DC, Osborne PA, Vejsada R, Golden JP, Lampe PA, Kato AC, Milbrandt J, Johnson EM., Jr Analysis of the retrograde transport of glial cell line-derived neurotrophic factor (GDNF), neurturin, and persephin suggests that in vivo signaling for the GDNF family is GFRalpha coreceptor-specific. J Neurosci. 1999;19:9322–9331. doi: 10.1523/JNEUROSCI.19-21-09322.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lin LH, Doherty DH, Lile JD, Bektesh S, Collins F. GDNF: a glial cell-derived neurotrophic factor for midbrain dopaminergic neurons. Science. 1993;260:1130–1132. doi: 10.1126/science.8493557. [DOI] [PubMed] [Google Scholar]
- 20.Lin JH, Saito T, Anderson DJ, Lance-Jones C, Jessell TM, Arber S. Functionally related motor neuron pool and muscle sensory afferent subtypes defined by coordinate ETS gene expression. Cell. 1998;95:393–407. doi: 10.1016/s0092-8674(00)81770-5. [DOI] [PubMed] [Google Scholar]
- 21.Milbrandt J, de Sauvage F, Fahrner TL, Baloh RH, Leitner ML, Tansey ML, Lampe PA, Heuckeroth RO, Kotzbauer PT, Simburger KS, Golden JP, Davies JA, Vejsada R, Kato AC, Hynes M, Sherman D, Nishimura M, Wang L-C, Vandlen R, Moffat B, Klein RD, Poulsen K, Gray C, Garces A, Henderson CE, Phillips HS, Johnson EM. Persephin, a novel neurotrophic factor related to GDNF and neurturin. Neuron. 1998;20:245–253. doi: 10.1016/s0896-6273(00)80453-5. [DOI] [PubMed] [Google Scholar]
- 22.Moore MW, Klein RD, Farinas I, Sauer H, Armanini M, Phillips H, Reichardt LF, Ryan AM, Carver-Moore K, Rosenthal A. Renal and neuronal abnormalities in mice lacking GDNF. Nature. 1996;382:76–79. doi: 10.1038/382076a0. [DOI] [PubMed] [Google Scholar]
- 23.Ohta K, Nakamura M, Hirokawa K, Tanaka S, Iwama A, Suda T, Ando M, Tanaka H. The receptor tyrosine kinase, Cek8, is transiently expressed on subtypes of motoneurons in the spinal cord during development. Mech Dev. 1996;54:59–69. doi: 10.1016/0925-4773(95)00461-0. [DOI] [PubMed] [Google Scholar]
- 24.Oppenheim RW, Houenou LJ, Johnson JE, Lin LF, Li L, Lo AC, Newsome AL, Prevette DM, Wang S. Developing motor neurons rescued from programmed and axotomy-induced cell death by GDNF. Nature. 1995;3730:344–346. doi: 10.1038/373344a0. [DOI] [PubMed] [Google Scholar]
- 25.Oppenheim RW, Houenou LJ, Parsadanian AS, Prevette D, Snider WD, Shen L. GDNF and developing mammalian motoneurrons: regulation of programmed cell death among motoneurons subtypes. J Neurosci. 2000;20:5001–5011. doi: 10.1523/JNEUROSCI.20-13-05001.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pfaff SL, Mendelsohn M, Stewart CL, Edlund T, Jessell TM. Requirement for LIM homeobox gene Isl1 in motor neuron generation reveals a motor neuron-dependent step in interneuron differentiation. Cell. 1996;84:309–320. doi: 10.1016/s0092-8674(00)80985-x. [DOI] [PubMed] [Google Scholar]
- 27.Rossi J, Luukko K, Poteryaev D, Laurikainen A, Sun YF, Laakso T, Eerikainen S, Tuominen R, Lakso M, Rauvala H, Arumae U, Pasternack M, Saarma M, Airaksinen MS. Retarded growth and deficits in the enteric and parasympathetic nervous system in mice lacking GFR alpha2, a functional neurturin receptor. Neuron. 1999;22:243–252. doi: 10.1016/s0896-6273(00)81086-7. [DOI] [PubMed] [Google Scholar]
- 28.Sanchez MP, Silos-Santiago I, Frisen J, He B, Lira SA, Barbacid M. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 1996;382:70–73. doi: 10.1038/382070a0. [DOI] [PubMed] [Google Scholar]
- 29.Sanicola M, Hession C, Worley D, Carmillo P, Ehrenfels C, Walus L, Robinson S, Jaworski G, Wei H, Tizard R, Whitty A, Pepinsky RB, Cate RL. Glial cell line-derived neurotrophic factor-dependent RET activation can be mediated by two different cell-surface accessory proteins. Proc Natl Acad Sci USA. 1997;94:6238–43. doi: 10.1073/pnas.94.12.6238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Snider WD, Wright DE. Neurotrophins cause a new sensation. Neuron. 1996;16:229–232. doi: 10.1016/s0896-6273(00)80039-2. [DOI] [PubMed] [Google Scholar]
- 31.Sockanathan S, Jessell TM. Motor neuron-derived retinoid signaling specifies the subtype identity of spinal motor neurons. Cell. 1998;94:503–514. doi: 10.1016/s0092-8674(00)81591-3. [DOI] [PubMed] [Google Scholar]
- 32.Soler RM, Dolcet X, Encinas M, Egea J, Bayascas JR, Comella JX. Receptors of the glial cell line-derived neurotrophic factor family of neurotrophic factors signal cell survival through the phosphatidylinositol 3-kinase pathway in spinal cord motoneurons. J Neurosci. 1999;19:9160–9169. doi: 10.1523/JNEUROSCI.19-21-09160.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Thompson J, Doxakis E, Pinon LG, Strachan P, Buj-Bello A, Wyatt S, Buchman VL, Davies AM. GFRalpha-4, a new GDNF family receptor. Mol Cell Neurosci. 1998;11:117–126. doi: 10.1006/mcne.1998.0682. [DOI] [PubMed] [Google Scholar]
- 34.Treanor JJ, Goodman L, de Sauvage F, Stone DM, Poulsen KT, Beck CD, Gray C, Armanini MP, Pollock RA, Hefti F, Phillips HS, Goddard A, Moore MW, Buj-Bello A, Davies AM, Asai N, Takahashi M, Vandlen R, Henderson CE, Rosenthal A. Characterization of a multicomponent receptor for GDNF. Nature. 1996;382:80–83. doi: 10.1038/382080a0. [DOI] [PubMed] [Google Scholar]
- 35.Trupp M, Belluardo N, Funakoshi H, Ibanez CF. Complementary and overlapping expression of glial cell line-derived neurotrophic factor (GDNF), c-ret proto-oncogene, and GDNF receptor- alpha indicates multiple mechanisms of trophic actions in the adult rat CNS. J Neurosci. 1997;17:3554–3567. doi: 10.1523/JNEUROSCI.17-10-03554.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tsuchida T, Ensini M, Morton SB, Baldassare M, Edlund T, Jessell TM, Pfaff SL. Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes. Cell. 1994;79:957–970. doi: 10.1016/0092-8674(94)90027-2. [DOI] [PubMed] [Google Scholar]
- 37.Widenfalk J, Nosrat C, Tomac A, Westphal H, Hoffer B, Olson L. Neurturin and glial cell line-derived neurotrophic factor receptor-beta (GDNFR-beta), novel proteins related to GDNF and GDNFR-alpha with specific cellular patterns of expression suggesting roles in the developing and adult nervous system and in peripheral organs. J Neurosci. 1997;17:8506–8519. doi: 10.1523/JNEUROSCI.17-21-08506.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wright DE, Snider WD. Focal expression of glial cell line-derived neurotrophic factor in developing mouse limb bud. Cell Tissue Res. 1996;286:209–217. doi: 10.1007/s004410050689. [DOI] [PubMed] [Google Scholar]
- 39.Yamamoto Y, Henderson CE. Patterns of programmed cell death in populations of developing spinal motoneurons in chicken, mouse, and rat. Dev Biol. 1999;214:60–71. doi: 10.1006/dbio.1999.9413. [DOI] [PubMed] [Google Scholar]
- 40.Yamamoto Y, Livet J, Pollock RA, Garces A, Arce V, deLapeyrière O, Henderson CE. Hepatocyte growth factor (HGF/SF) is a muscle-derived survival factor for a subpopulation of embryonic motoneurons. Development. 1997;124:2903–2913. doi: 10.1242/dev.124.15.2903. [DOI] [PubMed] [Google Scholar]
- 41.Yan Q, Matheson C, Lopez OT. In vivo neurotrophic effects of GDNF on neonatal and adult facial motor neurons. Nature. 1995;373:341–344. doi: 10.1038/373341a0. [DOI] [PubMed] [Google Scholar]
- 42.Yu T, Scully S, Yu Y, Fox GM, Jing S, Zhou R. Expression of GDNF family receptor components during development: implications in the mechanisms of interaction. J Neurosci. 1998;18:4684–4696. doi: 10.1523/JNEUROSCI.18-12-04684.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhao D, McCaffery P, Ivins KJ, Neve RL, Hogan P, Chin WW, Drager UC. Molecular identification of a major retinoic-acid-synthesizing enzyme, a retinaldehyde-specific dehydrogenase. Eur J Biochem. 1996;240:15–22. doi: 10.1111/j.1432-1033.1996.0015h.x. [DOI] [PubMed] [Google Scholar]