

Abstract
Assembly of the SNARE complex and its disassembly caused by the action of soluble N-ethylmaleimide-sensitive factor (NSF) attachment protein (SNAP) and NSF is crucial for the maintenance of vesicular traffic, including fusion of regulated exocytotic vesicles. Various other proteins may also have important roles in the processes leading to membrane fusion via interaction with the SNARE proteins, including the secretory vesicle cysteine string protein (Csp). Here we have examined the effect of overexpression of a dominant negative α-SNAP mutant or Csp on exocytosis of dense-core granules in single chromaffin cells monitored using amperometry to detect released catecholamine. Exocytosis of trans-Golgi network (TGN)-derived dense-core granules was substantially inhibited by expression of α-SNAP(L294A). The amplitude and characteristics of the individual release events were unaffected by expression of α-SNAP(L294A), consistent with an essential role for α-SNAP in early steps of priming but not in the fusion process. In contrast, Csp overexpression, which also inhibited the extent of exocytosis, also modified the kinetics of the individual release events seen as an increase in the rise time and a broadening of the residual amperometric spikes in Csp-transfected cells. These results suggest that unlike α-SNAP, Csp plays a key role in the protein interactions close to the fusion process or fusion pore opening during Ca2+-regulated exocytosis.
Keywords: exocytosis, secretion, SNAP, NSF, Csp, amperometry
An essential role for proteins of the SNARE family in vesicular traffic is well established, and syntaxin 1, synaptosomal associated protein of 25 kDa (SNAP-25), and vesicle-associated membrane protein (VAMP) are essential for vesicle fusion in regulated exocytosis in synapses and in neuroendocrine and endocrine cells (Schiavo et al., 1992; Hay and Scheller, 1997; Burgoyne and Morgan, 1998; Lang, 1999). Syntaxin 1, SNAP-25, and VAMP are able to form a stable complex (the SNARE complex) (Hayashi et al., 1994; Sutton et al., 1998). The assembly of the SNARE complex is regulated by many factors, and considerable attention has been given to the ability of the ATPase NSF to disassemble the SNARE complex after its recruitment by α-solubleN-ethylmaleimide-sensitive factor attachment protein (α-SNAP) (Sollner et al., 1993a,b). The original suggestion that disassembly of the complex by SNAP/NSF provided a driving force for membrane fusion (Sollner et al., 1993b) is inconsistent with various data (Morgan and Burgoyne, 1995a), and a current view is that assembly of the SNARE complex drives fusion (Sutton et al., 1998; Weber et al., 1998).
In the yeast homotypic vacuole fusion system, SNAP/NSF have a role in priming the SNAREs on each vacuole membrane before membrane docking and fusion (Mayer et al., 1996; Nichols et al., 1997; Ungermann et al., 1998). It has been assumed that the role of SNAP/NSF in the synapse is to disassemble the SNARE complex only after synaptic vesicle fusion. This would allow recycling of the SNARE proteins for subsequent rounds of fusion (Lin and Scheller, 1997; Rizo and Sudhof, 1998; Sutton et al., 1998; Weber et al., 1998). Synaptic vesicles are reused up to 1000 times, and factors that affect the recycling of the vesicles and the fusion machinery will have a major functional impact on exocytosis. In contrast, dense-core granule exocytosis in adrenal chromaffin cells involves a one-shot fusion event of a granule derived from the trans-Golgi network (TGN) (Winkler, 1977), which in the absence of its released content cannot effectively resequester catecholamine (Phillips, 1982). Chromaffin cells possess a large pool of mature granules, which in the absence of stimulation are stable, with their catecholamine content having a half-life greater than 15 d (Corcoran et al., 1984). Addition of α-SNAP stimulated dense-core granule exocytosis in permeabilized chromaffin cells (Chamberlain et al., 1995; Morgan and Burgoyne, 1995b), demonstrating that SNAP/NSF-mediated SNARE priming could occur before fusion. Data from patch-clamp capacitance analyses suggest that α-SNAP has an early function in recruitment of vesicles into the releaseable pool (Kibble et al., 1996; Xu et al., 1999); however, an essential requirement for SNAP/NSF function before dense-core granule fusion has not been demonstrated. The α-SNAP mutant, α-SNAP(L294A) (Barnard et al., 1997), which is a dominant negative inhibitor of endosome fusion (Christoforidis et al., 1999), did not inhibit exocytosis in permeabilized (Barnard et al., 1997) or patch-clamped chromaffin cells (Xu et al., 1999), which we have argued is attributable to insufficient time for exchange with endogenous α-SNAP. If SNARE priming by SNAP/NSF is essential in the early steps leading to fusion, then α-SNAP(L294A) should inhibit the extent of exocytosis of the preformed granules if exchange can occur but not affect the characteristics of the fusion process.
Many other proteins are likely to function in exocytosis via effects on the SNARE machinery (Sudhof, 1995; Burgoyne and Morgan, 1998). Among these is the cysteine string protein (Csp) (Zinsmaier et al., 1990), a protein found on synaptic vesicles (Mastrogiacomo et al., 1994) and secretory granules (Chamberlain et al., 1996; Pupier et al., 1997). Studies in Drosophila have shown that Csp is required for viability and for evoked neurotransmission (Umbach et al., 1994;Zinsmaier et al., 1994). It has been suggested that this is attributable to regulation by Csp of Ca2+channel function (Gundersen and Umbach, 1992; Umbach et al., 1998). In contrast, studies on dense-core granule exocytosis have suggested a direct role for Csp, independent of any effects on Ca2+ channels (Chamberlain and Burgoyne, 1998; Zhang et al., 1998, 1999). Csp can interact with Hsc70 Braun et al., 1996; Chamberlain and Burgoyne, 1997a,b) and has the properties of a molecular chaperone (Chamberlain and Burgoyne, 1997b). Its ability to interact with syntaxin (Wu et al., 1999; Nie et al., 1999) and/or VAMP (Leveque et al., 1998) would be consistent with a role in regulating SNARE function. It is not clear, however, whether this would be exerted in vesicle priming, docking, or fusion itself. Csp overexpression has a negative effect in insulin-secreting cells (Brown et al., 1998; Zhang et al., 1999) and in Drosophila (Nie et al., 1999); therefore, we have examined the effect of transient Csp overexpression on exocytosis in chromaffin cells. The results show that both α-SNAP(L294A) and Csp overexpression inhibits exocytosis. Despite the ability of both of these proteins to interact with SNARE proteins, only in the case of Csp was the kinetics of the individual fusion events altered. This is consistent with α-SNAP having an essential function early in the exocytotic pathway but Csp exerting its function close to the fusion event.
MATERIALS AND METHODS
Reagents. Tissue culture reagents were obtained from Life Technologies (Paisley, UK), and high-purity digitonin was from Novabiochem (Nottingham, UK). A plasmid that encodes a fluorescent-enhanced mutant of green fluorescent protein (pEGFP) was obtained from Clontech (Basingstoke, UK), and the expression vector pcDNA3.1 (−) was from Invitrogen (Leek, The Netherlands). All other reagents were obtained from Sigma (Poole, UK).
Buffers. Krebs-Ringer's buffer consisted of the following (in mm): 145 NaCl, 5 KCl, 1.3 MgCl2, 1.2 NaH2PO4, 10 glucose, and 20 HEPES, pH 7.4. Amperometry bath buffer contained (in mm): 139 potassium glutamate, 20 PIPES, 0.2 EGTA, 2 ATP, 2 MgCl2, pH 6.5. Amperometry cell permeabilization/stimulation buffer contained 139 mmpotassium glutamate, 20 mm PIPES, 5 mm EGTA, 2 mm ATP, 2 mm MgCl2, 20 μm digitonin, and 10 μmCa2+, pH 6.5 (total added CaCl2 was 4.16 mm). PBS contained (in mm): 142 NaCl, 2 KCl, 8 Na2HPO4, 1.5 NaH2PO4, pH 7.4. PBT buffer contained PBS and 0.3% BSA and 0.1% Triton X-100.
Plasmid constructs. The his-tagged α-SNAP (L294A) construct was originally created by site-directed mutagenesis in a pQE-9 vector (Barnard et al., 1997). For expression in mammalian cells, the his-tagged construct was amplified by PCR using Pfu polymerase (Stratagene, Amsterdam, The Netherlands) and cloned into pcDNA3.1(−) at EcoRI and HindIII restriction sites to create pα-SNAP(L294A). The wild-type α-SNAP construct was similarly subcloned into pcDNA3.1(−), and in each case the constructs were confirmed by automated sequencing. Csp1 in pcDNA3 was as a myc-tagged construct (Zhang et al., 1999).
Cell culture and transfection. Newly isolated bovine adrenal chromaffin cells (Burgoyne, 1992) were plated on non-tissue culture-treated 10 cm Petri dishes at a density of 1 × 106/ml and left overnight. Nonattached cells were gently pelleted by centrifugation and resuspended in growth medium at a density of 1 × 107/ml. pEGFP (20 μg) and 20 μg of pα-SNAP(L294A), pα-SNAP, or pCsp1 were added per milliliter of cells. Cells and plasmids (1 ml) were electroporated at 250 V and 975 μF for one pulse, using a Bio-Rad Gene Pulser II (Bio-Rad, Hercules, CA) and 4 mm cuvettes. Cells were then rapidly diluted to 1 × 106/ml with fresh growth media. Cells (1 × 106) were added to 13 mm Petri dishes and made up to a volume of 3 ml with fresh growth media and maintained in culture for an additional 3–5 d.
Immunofluorescence. Transfections were performed as described above except that cells were plated onto round glass coverslips (13 mm diameter). After washing twice with PBS, cells were fixed in 3.7% formaldehyde in PBS for 2 hr at room temperature. Cells were then washed twice in PBS, incubated for 30 min in PBT, and incubated overnight with a mouse monoclonal antibody against α-SNAP (clone C1 77.2, Synaptic Systems, Gottingen, Germany) at 1:500 dilution in PBT, anti-Csp antiserum (1:600), or rabbit antiserum against chromogranin A at 1:1000 (a gift from Prof. G. J. Dockray, The Physiological Lab, University of Liverpool). After they were washed three times in PBT, cells were incubated for 1 hr in biotinylated anti-mouse or anti-rabbit IgG (Amersham, Buckinghamshire, UK) at 1:100 in PBT, washed three times with PBT, and finally incubated in Streptavidin Texas Red (Amersham) at 1:50 dilution in PBT for 30 min. After cells were mounted, they were viewed with the appropriate filters to visualize the GPF fluorescence and immunofluorescence.
Amperometric recording. Cells were washed three times with Krebs-Ringer's buffer, incubated in bath buffer, and viewed using a Nikon TE300 inverted microscope. Transfected cells were identified as those fluorescing green (caused by expression of green fluorescent protein) under blue light illumination. A precut 5-μm-diameter carbon fiber electrode (NPI, Tamm, Germany) was positioned close to a cell using an Eppendorf (Hamburg, Germany) PatchMan micromanipulator. The fiber was moved against a cell until visible distortion of the cell membrane could be seen. The fiber was then pulled away until the point at which distortion was no longer visible but the fiber remained in contact with the cell surface. For stimulation, a digitonin-permeabilization protocol (Jankowski et al., 1992) was used. A glass micropipette filled with cell permeabilization/stimulation buffer (with 20 μm digitonin and 10 μm free calcium) was positioned on the opposite side of the cell from the carbon fiber, ∼60 μm from the cell. An Eppendorf Transjector was used to pressure-eject the buffer onto the cell for a 20 sec pulse. A holding voltage of +700 mV was applied across the carbon fiber tip and the Ag/AgCl reference electrode in the bath. Amperometric responses were monitored with a VA-10 amplifier (NPI Electronic),collected at 4 kHz, digitized with a Digidata 1200B acquisition system, and monitored online with the AxoScope program (Axon Instruments). Data were subsequently analyzed using an automated peak detection and analysis protocol with the technical graphics program Origin (Microcal). Spikes were only analyzed in detail if they had a base width greater than 6 msec and an amplitude greater than 40 pA. This amplitude was chosen so that analyses were confined to spikes arising immediately beneath the carbon fiber and to limit effects on the data of diffusion times from distant sites. All data are shown as mean ± SEM, and statistical differences were assessed using an unpaired Student's ttest.
RESULTS
Strategy for transfection and amperometric recording
We have tested whether SNAP/NSF function is essential before fusion of TGN-derived dense-core granules and the role of Csp by using transfection to overexpress the mutant α-SNAP(L294A) protein and Csp. To examine the effects on exocytosis in chromaffin cells, we used a single-cell transfection–amperometry approach (Fisher and Burgoyne, 1999). The cells were cotransfected with control pcDNA3 vector, a plasmid encoding α-SNAP(L294A) or Csp1 and pEGFP to allow visualization of transfected cells for recording. Electroporation resulted in transfection, seen by GFP fluorescence, in ∼1–5% of the chromaffin cells. Cotransfection did not change the percentage of cells that were transfected and had no detectable effect on cell morphology. To confirm cotransfection with two plasmids, transfected cells were fixed and stained with concentrations of antisera determined to be too low to stain control nontransfected cells (Fig.1a).
Fig. 1.

Cotransfection of adrenal chromaffin cells and expression of GFP and α-SNAP. a, Immunofluorescence demonstrates expression of GFP and overexpression of α-SNAP in the same cell. Chromaffin cells were transfected with plasmids encoding GFP and α-SNAP(L294A) and after fixation stained with anti-α-SNAP antiserum at a concentration below that required to stain nontransfected cells. A GFP-expressing cell is shown clearly stained with anti-α-SNAP, but an attached GFP-negative cell (asterisk) was barely stained by anti-α-SNAP. Scale bar, 10 μm. b, Immunofluorescence staining with anti-chromogranin A (CGA) after transfection with plasmids encoding GFP and α-SNAP(L294A). Similar extents of staining were seen in GFP-expressing and nonexpressing cells. c, Scheme of the recording configuration that was used. Note that the ejection pipette was actually positioned ∼60 μm from the cell to be stimulated, and the carbon-fiber electrode was positioned in close contact with the cell surface.
Exocytosis was detected after transfection using a carbon-fiber electrode for amperometric recording of catecholamine release (Wightman et al., 1991) from the transfected (GFP-expressing) cells (Fig.1c), from control nontransfected cells in the same dish, or from control cells transfected with GFP and pcDNA3 vector. The cells were maintained in the absence of Ca2+(with 0.2 mm EGTA in the bath buffer), and exocytosis was evoked using pressure ejection from a pipette containing 20 μm digitonin and 10 μm free Ca2+ to both permeabilize and stimulate the cells directly (Fig.1c). This approach was taken to bypass any effects of transfection and expression on agonist receptors or membrane channels and to allow direct assay of effects of expression on Ca2+-triggered exocytosis. To optimize the time resolution of measurements of amperometric spikes, the carbon fiber was placed in direct contact with the cell surface. Contact did not induce any responses from the cells and in the absence of a stimulus, current spikes were rarely seen. Application of digitonin/Ca2+ evoked a burst of fast, transient current spikes (with most spikes in the first minute after perfusion and fewer at later times) characteristic of the kinetics and amplitude of release of catecholamine from single granule fusion events (Wightman et al., 1991; Chow et al., 1992; Albillos et al., 1997). Preliminary experiments established that no spikes were evoked in the absence of digitonin in the pipette solution (n = 4 cells), demonstrating that permeabilization was essential for responses to be evoked by perfusion with Ca2+. In addition, the spikes were essentially indistinguishable from those evoked in intact cells by agonists and distinct from slow release events attributable to granule lysis seen after prolonged digitonin treatment (Jankowski et al., 1992). Ca2+-induced catecholamine release from populations of digitonin-permeabilized chromaffin cells has been well characterized (Burgoyne, 1991) and shown to occur by bona fide exocytotic machinery. Indeed, under the conditions used here, Ca2+-evoked amperometric spikes were almost completely abolished by cotransfection with botulinum toxins C1 or E (Graham et al., 2000), indicating that they occurred via a SNARE-dependent mechanism.
Expression of α-SNAP(L294A) inhibits exocytosis
Cotransfection with GFP and α-SNAP(L294A) plasmids resulted in detectable expression of α-SNAP(L294A) in GFP-positive cells. Fig.1a shows that GFP-expressing cells were brightly stained with a low concentration of anti-α-SNAP antiserum, indicating overexpression of α-SNAP(L294A) in those cells. Almost all GFP-expressing cells showed α-SNAP staining above the background staining of nontransfected cells. We established that expression of α-SNAP(L294A) did not lead to depletion of secretory granules. Cells cotransfected with plasmids encoding GFP and α-SNAP(L294A) were stained with an antiserum against the granule content protein chromogranin A to a similar extent as control nontransfected cells (Fig. 1b), ruling out the possibility of granule depletion.
It is clear from the example traces shown in Figure2 that expression of the mutant protein α-SNAP(L294A) resulted in a marked reduction in the number of spikes evoked after application of digitonin/Ca2+. From a series of recordings on separate cells, the effect of α-SNAP(L294A) expression on the frequency and characteristics of the spikes was determined. Expression of the α-SNAP(L294A) mutant resulted in a marked (84%,p < 0.001, Student's t test) reduction (to 3.61 ± 1.77 per cell) in the average number of evoked spikes per cell compared with nontransfected cells in the same dishes (Fig.2c). In contrast, transfection with the plasmid encoding GFP alone did not result in any reduction in mean spike number (24.0 ± 2.95 per cell) compared with control nontransfected cells (22.5 ± 3.9 per cell), indicating that the reduction in spike number with α-SNAP(L294A) was not simply a consequence of transfection occurring in a nonfunctional subpopulation of cells or to an inhibitory effect of expression of any exogenous protein in transfected cells. Overexpression of wild-type α-SNAP had no statistically significant effect on spike number compared with control cells (data not shown).
Fig. 2.
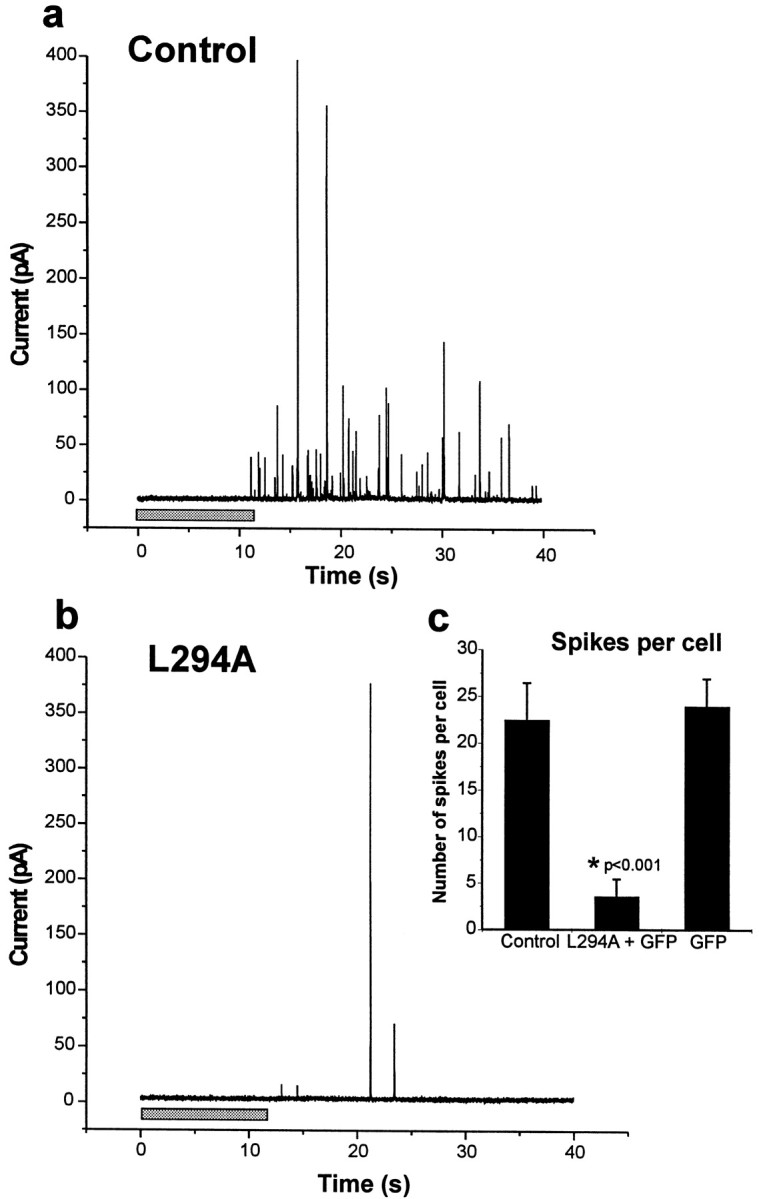
Amperometric recordings from control and α-SNAP(L294A) transfected cells. a,b, The traces shown are examples of control nontransfected cells and cells transfected with GFP and α-SNAP(L294A) assayed in the same dish after stimulation with a 20 sec pulse of digitonin/Ca2+. The recording is shown from near the middle of the stimulation pulse (shaded bar) and continues over the initial period after stimulation.c, The inset shows the mean spikes per cell (as mean ± SEM) over a 2 min period after stimulation for control nontransfected cells (n = 17 cells), cells showing GFP expression from cultures transfected with pEGFP and pαSNAP(L294A) (n = 18 cells), or pEGFP alone (n = 5 cells).
Expression of α-SNAP(L294A) does not affect spike characteristics
Because the extent of exocytosis was reduced by expression of α-SNAP(L294A), the characteristics of the individual residual spikes were examined. The amplitude and time course of spikes (Fig.3a) were similar to those previously reported for both intact and permeabilized chromaffin cells (Wightman et al., 1991; Chow et al., 1992; Jankowski et al., 1992). Despite the reduction in spike number, the peak amplitude and the total charge carried by the spikes that remained were not decreased compared with control cells (Fig. 3c,d), showing that the reduction in spike number was not caused by depletion of granule catecholamine. The overall shape of the spikes did not appear to be affected by expression of α-SNAP(L294A), and the mean values for the half-widths of the spikes was no different from that of control cells. The spike characteristics were also no different in cells recorded in parallel that were transfected with GFP alone. These results indicate that α-SNAP(L294A) expression did not affect the time course of catecholamine release during individual granule fusion events (Fig.3b).
Fig. 3.

Analysis of amperometric spikes from control and α-SNAP (L294A) transfected cells. a, Example spikes from control, nontransfected, and GFP-expressing transfected cells (L294A) are shown on an expanded time base. Mean values of the half-width (b), the total charge carried per spike (c), and the mean value for peak spike height (d) are shown for spikes from control, nontransfected cells (n = 383 spikes) and α-SNAP(L294A) transfected cells (n = 65 spikes). Data are shown as mean ± SEM.
Overexpression of Csp inhibits exocytosis
Overexpression of Csp by transfection with a plasmid encoding Csp1 (Chamberlain and Burgoyne, 1996) was confirmed by immunofluorescence labeling with a low concentration of anti-Csp that labeled GFP-positive but not GFP-negative cells. (Fig.4a).
Fig. 4.

Cotransfection of adrenal chromaffin cells showing GFP expression and overexpression of Csp. a, Immunofluorescence demonstrates expression of GFP and overexpression of Csp in the same cell after staining with a concentration of anti-Csp antiserum too low to stain surrounding nontransfected cells (asterisk). b, Immunofluorescence staining with anti-chromogranin A (CGA) after transfection with plasmids encoding GFP and Csp1 showing similar levels of chromogranin A staining in GFP-expressing and nonexpressing cells. Scale bar, 10 μm.
Based on the presence of chromogranin A staining, contransfection did not deplete the granule population (Fig. 4b). After stimulation, Csp overexpression resulted in a reduction in the number of amperometric spikes induced by digitonin/Ca2+. Figure5a,b shows representative traces from a control cell transfected with the GFP plasmid plus pcDNA3 and from a cell transfected with the GFP and the Csp1 plasmids. Recordings from Csp-transfected and control cells were performed on the same day and with the same carbon-fiber electrodes. In a series of cells from the two conditions recorded in parallel, Csp overexpression reduced the average number of evoked spikes by 82% (p < 0.001, Student's t test), from 17.0 ± 4.2 spikes per cell in cells expressing GFP alone to 3.0 ± 0.9 for cells overexpressing Csp (Fig. 5c).
Fig. 5.

Amperometric recordings from control and Csp-transfected cells. a, b, The traces shown are examples of control GFP-expressing cells and Csp-transfected GFP-expressing cells after stimulation after a 20 sec pulse of digitonin/Ca2+. The inset shows the mean spikes per cell (shown as mean ± SEM) over a 2 min period after stimulation for control cells (n = 18 cells) and Csp-transfected cells (n = 24 cells).
Overexpression of Csp modifies the time course of amperometric spikes
Analysis of the individual amperometric spikes from control cells and the residual spikes from Csp-transfected cells revealed that although the mean amplitude (spike height) was unaffected by Csp overexpression (Fig. 6d), the total charge per spike was significantly increased (p < 0.001, Student's t test) (Fig.6c) from 0.92 ± 0.05 pC for GFP control spikes to 1.49 ± 0.17 pC for spikes from Csp-overexpressing cells. The reason for the increase in total charge was apparent on close inspection of the individual spikes. Those from CSP-transfected cells were often broader than those typically seen in nontransfected (Fig. 3) or control GFP-expressing cells (Fig. 6a), with an overall increase in the mean half-width of the spikes of 60% (p < 0.001, Student's t test) caused by Csp overexpression (Fig.6b). This resulted from a change from a half-width of 6.6 ± 0.24 msec for control GFP spikes to 10.5 ± 0.87 msec for those from Csp-overexpressing cells, whereas the mean spike amplitudes of 115.7 ± 5.6 pA for GFP control spikes and 105.9 ± 8.5 pA for Csp-overexpressing cells were not significantly different. In addition, overlay of the spikes revealed an apparent slowing of the rate of rise (Fig. 7a), and this was reflected in a mean 44% increase (p < 0.001, Student's t test) in the mean rise time in Csp-transfected cells (Fig. 7b) from 5.4 ± 0.16 msec for GFP control spikes to 7.82 ± 0.46 msec for those from Csp-overexpressing cells. The differences in spike kinetics in Csp-overexpressing cells are unlikely to be related to increased instability of granules in these cells after digitonin application because it has been shown that spontaneous lysis of granules attributable to prolonged digitonin exposure, for example, results in slow (half-width 50–250 msec) low-amplitude release events (Jankowski et al., 1992) distinct from those seen in control or overexpressing cells in the present study. In addition, we only analyzed spikes larger than 40 pA, which would exclude such spikes from our analyses. These data, therefore, implicate Csp in events related to fusion pore formation or the control of fusion pore opening.
Fig. 6.

Analysis of amperometric spikes from control and Csp-transfected cells. a, Example spikes representing the average spikes in control GFP-expressing cells and Csp-transfected cells on an expanded time base. Mean values of the half-width of the spikes (b), the total carried per spike (c), and the mean value for peak spike height (d) are shown for control (n= 302 spikes) and Csp-transfected (n = 64 spikes). Data are shown as mean ± SEM.
Fig. 7.
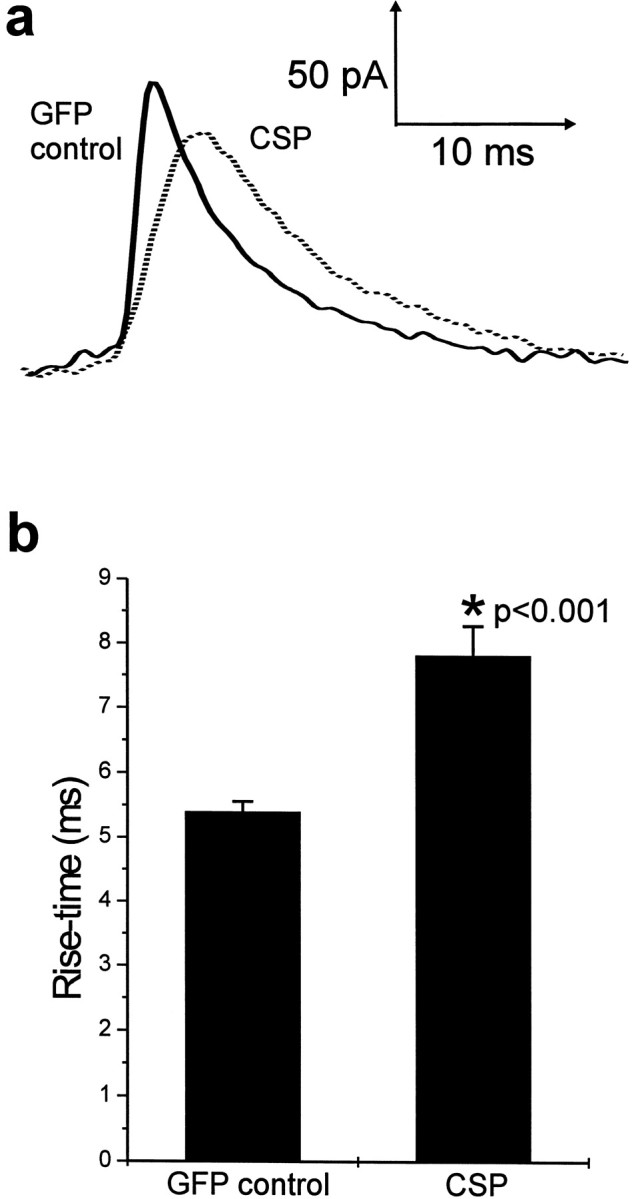
Effect of Csp overexpression on the initial kinetics of amperometric spikes. a, The spikes showing the average characteristics from Figure 6 are overlaid to show the apparent slower rising phase of the spikes from Csp-transfected cells.b, Mean values (shown as mean ± SEM) for the rise time of amperometric spikes from control (n = 302 spikes) and Csp-transfected (n = 64 spikes) cells.
DISCUSSION
Many proteins are required to function in defined steps of the exocytotic pathway to provide the control, speed, and specificity of Ca2+-regulated exocytosis (Sudhof, 1995). The sites of action of most identified proteins in this process are still to be resolved (Burgoyne and Morgan, 1998), and a key question is the identity of proteins involved in the actual fusion process. In this study we have used transfection–amperometry (Fisher and Burgoyne, 1999) to analyze and compare the roles of two proteins, α-SNAP and Csp, and in particular made use of the ability of carbon-fiber amperometry to resolve the kinetics of single fusion events (Wightman et al., 1991; Chow et al., 1992; Jankowski et al., 1992). These two proteins were compared because α-SNAP is likely to function in early priming steps in exocytosis (Chamberlain et al., 1995; Burgoyne and Morgan, 1998; Xu et al., 1999). Csp was earlier suggested to function as a regulator of voltage-dependent Ca2+channels (Gundersen and Umbach, 1992; Umbach et al., 1998); it has recently been implicated as a potential regulator of the exocytotic machinery because of its ability to interact with the same exocyotic SNARE proteins as α-SNAP, but it is not clear whether it acts in early or late stages of the pathway. Analysis of the effect of Csp overexpression suggests that in contrast to α-SNAP, Csp interacts with the exocytotic fusion machinery to play a late role close to the fusion process.
The approach used in this paper depends on efficient cotransfection with two plasmids. We demonstrated a high level of co-transfection using immunofluorescence detection of protein overexpression, and the functional consequences of α-SNAP(L294A) or Csp transfection on generation of amperometric spikes also indicated a high (∼85%) level of cotransfection. The data presented here show that expression of α-SNAP(L294A) or overexpression of Csp in chromaffin cells after transfection significantly inhibited Ca2+-evoked exocytosis. This could not be caused by effects on synthesis of new granules because chromaffin cells maintain a large (30,000 per cell) and stable pool of mature granules (Winkler, 1977; Corcoran et al., 1984) in the absence of stimulation. These have a half-life much longer than the transfection period that was used, and on the basis of chromogranin A staining, transfected cells did not appear to be depleted of secretory granules. Synthesis of new granules in these cells can be readily detected only after massive nonphysiological stimulation to deplete preexisting granules and to activate pathways for granule biogenesis (Winkler and Fischer-Colbrie, 1998). In addition, because the stimulation was based on the use of permeabilized cells with direct activation of exocytosis by Ca2+, the inhibitory effects cannot be attributed to effects on membrane channels.
Study of the temperature-sensitive comatose mutant inDrosophila has produced conflicting data on the site of accumulation of the SNARE complex at the restrictive temperature. This has lead to different interpretations about the site of action of NSF either before or after fusion (Kawasaki et al., 1998; Littleton et al., 1998; Tolar and Pallanck, 1998). It is important that in contrast to the nerve terminal, the extent of exocytosis should not be affected by interference with vesicle membrane or SNARE protein recycling in the chromaffin cell in which there is one-shot usage of TGN-derived granules. α-SNAP(L294A) can recruit NSF to SNAREs but is unable to stimulate the ATPase activity of NSF (Barnard et al., 1997) and so cannot support SNARE priming (Barnard et al., 1997; Christoforidis et al., 1999). The simplest interpretation of the data from α-SNAP(L294A) expression, therefore, is that SNAP-dependent SNARE priming through NSF ATPase activity must be required before fusion of naive secretory granules. Such a priming event could involve priming of SNAREs on the granule or the plasma membrane or both and does not rule out an additional role of SNAP/NSF in SNARE complex disassembly and recycling after fusion that would also be crucial for synaptic transmission.
There is abundant evidence that the SNAREs need not be part of a full complex to act as SNAP receptors because syntaxin alone (Hanson et al., 1995) and also the syntaxin–SNAP-25 dimeric complex (Hayashi et al., 1995) can bind α-SNAP and support SNAP/NSF-mediated disassembly and conformational change. It is likely, therefore, that the SNAP/NSF chaperones (Morgan and Burgoyne, 1995a) interact with SNAREs at multiple points of the vesicle cycle (Burgoyne and Morgan, 1998). SNAREs (Tagaya et al., 1995; Hohne-Zell and Gratzl, 1996; Otto et al., 1997), α-SNAP, and NSF (Hong et al., 1994; Burgoyne and Williams, 1997) are present on chromaffin granules and synaptic vesicles, and so SNAP/NSF-mediated priming could occur on undocked secretory vesicles. Alternatively, priming could occur on vesicles already tethered to the plasma membrane (and seen as morphologically docked) (Banerjee et al., 1996) but in either case would be most likely to occur as a prelude to SNARE complex formation at the site of fusion. It has recently been suggested that NSF and α-SNAP can act directly as membrane fusogens (Otter-Nilsson et al., 1999), but data from chromaffin cells clearly show that late steps in Ca2+-triggered fusion do not require ATP hydrolysis (Parsons et al., 1995; Xu et al., 1999) and are insensitive to N-ethylmaleimide arguing against a late role for NSF (Xu et al., 1999). Expression of α-SNAP(L294A), although depressing the number of release events, did not affect the characteristics of the remaining amperometric spikes, indicating that the α-SNAP mutant did not affect the kinetics of individual fusion events. The use of chromaffin cells allows experimental investigation of effects of expressed proteins on only the outward arm of the exocytotic cycle, and therefore the data demonstrate an essential role for SNAP-mediated SNARE priming before fusion in Ca2+-regulated exocytosis of dense-core granules.
The data from the analysis of Csp overexpression demonstrate that not only does this reduce the extent of exocytosis but it also modifies the kinetics of the residual amperometric spikes, resulting in an increase in the spike half-width and an increase in the mean rise time. An increase in spike half-width could have resulted from an increase in granule size or in the amount of catecholamine content per granule. This is unlikely, however, to explain the changes in the kinetics of the rising phase of the spike attributable to Csp overexpression. The effects of Csp overexpression are also unlikely to be caused by an effect of granule biogenesis given the large number of stable preformed granules in these cells as discussed above. The rise time of the amperometric spike is likely to represent the kinetics of the initial fusion event or more likely the kinetics of fusion pore opening. These results, therefore, functionally demonstrate that Csp exerts its role on the fusion machinery in chromaffin cells. Previous work has shown that stable overexpression of Csp in PC12 cells enhanced exocytosis in permeabilized cells (Chamberlain and Burgoyne, 1998), but as in the present study, transient overexpression in insulin-secreting cells was inhibitory (Zhang et al., 1999). Other examples are known in which a protein required for exocytosis, including syntaxin, is inhibitory when overexpressed (Schulze et al., 1994; Fujita et al., 1998; Wu et al., 1998). The reason for the difference between transient and stable overexpression of Csp in the effect on exocytosis is unclear, but one possible explanation is that stable overexpression of Csp in PC12 cells resulted in compensatory changes in the exocytotic machinery. This difference remains to be resolved. The inhibitory effect of Csp overexpression on exocytosis does not appear to require interaction of Csp with Hsc70. Mutations within the HPD motif of Csp prevent activation of the ATPase activity of Hsc70 (Chamberlain and Burgoyne, 1997b) but do not prevent the inhibitory effect of Csp overexpression on exocytosis (Zhang et al., 1999). Two other studies have examined single fusion events in situations of modified Csp expression without reporting changes in release kinetics. First, no changes in spontaneous release events were reported in Drosophila Csp null mutants (Umbach et al., 1994). Second, overexpression of Csp in INS-1 cells was found to inhibit amperometric events (Brown et al., 1998), but no data on the kinetics of individual release events were described. In both studies the small size of the events may have precluded the type of analysis possible here with examination of the release from large chromaffin granules.
Csp has been subject to debate over whether its primary action in regulated exocytosis is to stimulate Ca2+entry through Ca2+ channels (Gundersen and Umbach, 1992; Umbach et al., 1998), to regulate SNARE assembly at Ca2+ channels (Leveque et al., 1998), or to more directly affect the exocytotic pathway (Chamberlain and Burgoyne, 1998; Zhang et al., 1998, 1999). The data here and from previous studies in which effects are preserved in permeabilized cells (Chamberlain and Burgoyne, 1998; Zhang et al., 1998, 1999) argue for a Ca2+ channel-independent function for Csp. We are now able to extend this further from the analysis of individual amperometric spikes in Csp-overexpressing cells. If the inhibitory effect of Csp was caused by a blockade of vesicle priming or docking, then we would expect to see only a reduction in spike number and no effect on spike kinetics as seen for α-SNAP(L294A). In contrast, the data from Csp-overexpressing cells, demonstrating a change in the kinetics of the release event, would be consistent with a postdocking role for Csp acting directly or indirectly at the level of proteins in the fusion machinery or involved in fusion pore expansion. An effect on the kinetics of the release event has recently been seen resulting from expression of a SNAP-25 mutant (Criado et al., 1999), indicating that such changes reflect modifications to the fusion machinery.
The possession by Csp of a J domain allowing it to interact with Hsc70 and its ability to act as a general molecular chaperone (Braun et al., 1996; Chamberlain and Burgoyne, 1997a,b) suggests interaction with various substrate proteins. Recent work has shown thatDrosophila Csp can interact with syntaxin in vitro (Wu et al., 1999) and in vivo (Nie et al., 1999). The significance of this for the mammalian Csp is unclear because this was reported not to bind to syntaxin but was immunoprecipitated with VAMP (Leveque et al., 1998). The ability of Csp to interact with SNARE proteins does not provide any detailed insight as to how and when its function is exerted and whether this would be seen as changes in vesicle priming, docking, fusion, endocytosis, or recycling. The function of Csp in these steps has been unclear, but we now provide a resolution to this issue and implicate Csp in late stages of the fusion process. It seems probable that a major function of Csp is in the correct folding of SNARE and other key proteins of the exocytotic fusion machinery, and thus it directly influences fusion pore kinetics.
Footnotes
This work was supported by a grant from The Wellcome Trust to R.D.B.
Correspondence should be addressed to Dr. Robert D. Burgoyne, The Physiological Laboratory, The University of Liverpool, Crown Street, Liverpool L69 3BX, UK. E-mail:burgoyne@liv.ac.uk.
REFERENCES
- 1.Albillos A, Dernick G, Horstmann H, Almers W, Alvarez de Toledo G, Lindau M. The exocytotic event in chromaffin cells revealed by patch amperometry. Nature. 1997;389:509–512. doi: 10.1038/39081. [DOI] [PubMed] [Google Scholar]
- 2.Banerjee A, Barry VA, DasGupta BR, Martin TFJ. N-ethylmalemide-sensitive factor acts at a prefusion ATP-dependent step in Ca2+ activated exocytosis. J Biol Chem. 1996;271:20223–20226. doi: 10.1074/jbc.271.34.20223. [DOI] [PubMed] [Google Scholar]
- 3.Barnard RJO, Morgan A, Burgoyne RD. Stimulation of NSF ATPase activity by α-SNAP is essential for SNARE complex disassembly and exocytosis. J Cell Biol. 1997;139:875–883. doi: 10.1083/jcb.139.4.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Braun JEA, Wilbanks SM, Scheller RH. The cysteine string secretory vesicle protein activates Hsc70 ATPase. J Biol Chem. 1996;271:25989–25993. doi: 10.1074/jbc.271.42.25989. [DOI] [PubMed] [Google Scholar]
- 5.Brown H, Larsson O, Branstrom R, Yang S-N, Leibiger B, Leibiger I, Fried G, Moede T, Deeney JT, Brown GR, Jacobsson G, Rhodes CJ, Braun JEA, Scheller RH, Corkey BE, Berggren P-O, Meister B. Cysteine string protein (CSP) is an insulin secretory granule-associated protein regulating β-cell exocytosis. EMBO J. 1998;17:5048–5058. doi: 10.1093/emboj/17.17.5048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Burgoyne RD. Control of exocytosis in adrenal chromaffin cells. Biochim Biophys Acta. 1991;1071:174–201. doi: 10.1016/0304-4157(91)90024-q. [DOI] [PubMed] [Google Scholar]
- 7.Burgoyne RD. Investigation of the intracellular regulators and components of the exocytotic pathway. In: Boulton A, Baker G, Taylor C, editors. Neuromethods, Vol 20: intracellular messengers. Humana; Totowa, NJ: 1992. pp. 433–470. [Google Scholar]
- 8.Burgoyne RD, Morgan A. Analysis of regulated exocytosis in adrenal chromaffin cells: insights into NSF/SNAP/SNARE function. BioEssays. 1998;20:328–335. doi: 10.1002/(SICI)1521-1878(199804)20:4<328::AID-BIES9>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- 9.Burgoyne RD, Williams G. NSF and SNAP are present on adrenal chromaffin granules. FEBS Lett. 1997;414:349–352. doi: 10.1016/s0014-5793(97)01031-4. [DOI] [PubMed] [Google Scholar]
- 10.Chamberlain LH, Burgoyne RD. Identification of a novel cysteine string protein variant and expression of cysteine string proteins in non-neuronal cells. J Biol Chem. 1996;271:7320–7323. doi: 10.1074/jbc.271.13.7320. [DOI] [PubMed] [Google Scholar]
- 11.Chamberlain LH, Burgoyne RD. Activation of the ATPase activity of heat shock proteins Hsc70/Hsp70 by cysteine-string protein. Biochem J. 1997a;322:853–858. doi: 10.1042/bj3220853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chamberlain LH, Burgoyne RD. The molecular chaperone function of the secretory vesicle cysteine string proteins. J Biol Chem. 1997b;272:31420–31426. doi: 10.1074/jbc.272.50.31420. [DOI] [PubMed] [Google Scholar]
- 13.Chamberlain LH, Burgoyne RD. Cysteine string protein functions directly in exocytosis. Mol Biol Cell. 1998;9:2259–2267. doi: 10.1091/mbc.9.8.2259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chamberlain LH, Roth D, Morgan A, Burgoyne RD. Distinct effects of α-SNAP, 14–3-3 proteins and calmodulin on priming and triggering of regulated exocytosis. J Cell Biol. 1995;130:1063–1071. doi: 10.1083/jcb.130.5.1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chamberlain LH, Henry J, Burgoyne RD. Cysteine string proteins are associated with chromaffin granules. J Biol Chem. 1996;271:19514–19517. doi: 10.1074/jbc.271.32.19514. [DOI] [PubMed] [Google Scholar]
- 16.Chow RH, von Ruden L, Neher E. Delay in vesicle fusion revealed by electrochemical monitoring of single secretory events in adrenal chromaffin cells. Nature. 1992;356:60–63. doi: 10.1038/356060a0. [DOI] [PubMed] [Google Scholar]
- 17.Christoforidis S, McBride HM, Burgoyne RD, Zerial M. The Rab5 effector EEA1 is a core component of endosome docking. Nature. 1999;397:621–625. doi: 10.1038/17618. [DOI] [PubMed] [Google Scholar]
- 18.Corcoran JJ, Wilson SP, Kirshner N. Flux of catecholamines through chromaffin vesicles in cultured bovine adrenal medullary cells. J Biol Chem. 1984;259:6208–6214. [PubMed] [Google Scholar]
- 19.Criado M, Gil A, Viniegra S, Gutierrez LM. A single amino acid near the C-terminus of the synaptosome- associated protein of 25kDa (SNAP-25) is essential for exocytosis in chromaffin cells. Proc Natl Acad Sci USA. 1999;96:7256–7261. doi: 10.1073/pnas.96.13.7256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fisher RJ, Burgoyne RD. The effect of transfection with Botulinum neurotoxin C1 light chain in exocytosis measured in cell populations and by single cell amperometry in PC12 cells. Pflügers Arch Eur J Physiol. 1999;437:754–762. doi: 10.1007/s004240050842. [DOI] [PubMed] [Google Scholar]
- 21.Fujita Y, Shirataki H, Sakisaka T, Asakura T, Ohya T, Kotani H, Yokoyama S, Nishioka H, Matsuura Y, Mizoguchi A, Scheller RH, Takai Y. Tomosyn: a syntaxin-1-binding protein that forms a novel complex in the neurotransmitter release process. Neuron. 1998;20:905–915. doi: 10.1016/s0896-6273(00)80472-9. [DOI] [PubMed] [Google Scholar]
- 22.Graham ME, Fisher RJ, Burgoyne RD (2000) Measurement of exocytosis by amperometry in adrenal chromaffin cells: effects of clostridial neurotoxins and activation of protein kinase C on fusion pore kinetics. Biochimie, in press. [DOI] [PubMed]
- 23.Gundersen CB, Umbach JA. Suppression cloning of the cDNA for a candidate subunit of a presynaptic calcium channel. Neuron. 1992;9:527–537. doi: 10.1016/0896-6273(92)90190-o. [DOI] [PubMed] [Google Scholar]
- 24.Hanson PI, Otto H, Barton N, Jahn R. The N-ethylmalemide-sensitive fusion protein and a-SNAP induce a conformational change in syntaxin. J Biol Chem. 1995;270:16955–16961. doi: 10.1074/jbc.270.28.16955. [DOI] [PubMed] [Google Scholar]
- 25.Hay JC, Scheller RH. SNAREs and NSF in targeted membrane fusion. Curr Opin Cell Biol. 1997;9:505–512. doi: 10.1016/s0955-0674(97)80026-9. [DOI] [PubMed] [Google Scholar]
- 26.Hayashi T, McMahon H, Yamasaki S, Binz T, Hata Y, Sudhof TC, Niemann H. Synaptic vesicle membrane fusion complex: action of clostridial neurotoxins on assembly. EMBO J. 1994;13:5051–5061. doi: 10.1002/j.1460-2075.1994.tb06834.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hayashi T, Yamasaki S, Nauenburg S, Binz T, Niemann H. Disassembly of the reconstituted synaptic vesicle membrane complex in vitro. EMBO J. 1995;14:2317–2325. doi: 10.1002/j.1460-2075.1995.tb07226.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hohne-Zell B, Gratzl M. Adrenal chromaffin cells contain functionally different SNAP-25 monomers and SNAP-25/syntaxin heterodimers. FEBS Lett. 1996;394:109–116. doi: 10.1016/0014-5793(96)00931-3. [DOI] [PubMed] [Google Scholar]
- 29.Hong R-M, Mori H, Fukui T, Moriyama Y, Futai M, Yamamoto A, Tashiro Y, Tagaya M. Association of N-ethylmaleimide-sensitive fusion protein with synaptic vesicles. FEBS Lett. 1994;350:253–257. doi: 10.1016/0014-5793(94)00778-0. [DOI] [PubMed] [Google Scholar]
- 30.Jankowski JA, Schroeder TJ, Holz RW, Wightman RM. Quantal secretion of catecholamines measured from individual bovine adrenal medullary cells permeabilized with digitonin. J Biol Chem. 1992;267:18329–18335. doi: 10.21236/ada251716. [DOI] [PubMed] [Google Scholar]
- 31.Kawasaki F, Mattiuz AM, Ordway RW. Synaptic physiology and ultrastructure in comatose mutants define an in vivo role for NSF in neurotransmitter release. J Neurosci. 1998;18:10241–10249. doi: 10.1523/JNEUROSCI.18-24-10241.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kibble AV, Barnard RJO, Burgoyne RD. Patch-clamp capacitance analysis of the effects of alpha-SNAP on exocytosis in adrenal chromaffin cells. J Cell Sci. 1996;109:2417–2422. doi: 10.1242/jcs.109.9.2417. [DOI] [PubMed] [Google Scholar]
- 33.Lang J. Molecular mechanisms and regulation of insulin exocytosis as a paradigm of endocrine secretion. Eur J Biochem. 1999;259:3–17. doi: 10.1046/j.1432-1327.1999.00043.x. [DOI] [PubMed] [Google Scholar]
- 34.Leveque C, Pupier S, Marqueze B, Geslin L, Kataoka M, Takahashi M, DeWaard M, Seagar M. Interaction of cysteine string proteins with the α1A subunit of the P/Q-type calcium channel. J Biol Chem. 1998;272:13488–13492. doi: 10.1074/jbc.273.22.13488. [DOI] [PubMed] [Google Scholar]
- 35.Lin RC, Scheller RH. Structural organisation of the synaptic exocytosis core complex. Neuron. 1997;19:1087–1094. doi: 10.1016/s0896-6273(00)80399-2. [DOI] [PubMed] [Google Scholar]
- 36.Littleton JT, Chapman ER, Kreber R, Garment MB, Carlson SD, Ganetzky B. Temperature-sensitive paralytic mutations demonstrate that synaptic exocytosis requires SNARE complex assembly and disassembly. Neuron. 1998;21:401–413. doi: 10.1016/s0896-6273(00)80549-8. [DOI] [PubMed] [Google Scholar]
- 37.Mastrogiacomo A, Parsons SM, Zampighi GA, Jenden DJ, Umbach JA, Gundersen CB. Cysteine string proteins: a potential link between synaptic vesicles and presynaptic Ca2+ channels. Science. 1994;263:981–982. doi: 10.1126/science.7906056. [DOI] [PubMed] [Google Scholar]
- 38.Mayer A, Wickner W, Haas A. Sec18p (NSF)-driven release of sec17p (α-SNAP) can precede docking and fusion of yeast vacoules. Cell. 1996;85:83–94. doi: 10.1016/s0092-8674(00)81084-3. [DOI] [PubMed] [Google Scholar]
- 39.Morgan A, Burgoyne RD. Is NSF a fusion protein? Trends Cell Biol. 1995a;5:335–339. doi: 10.1016/s0962-8924(00)89059-5. [DOI] [PubMed] [Google Scholar]
- 40.Morgan A, Burgoyne RD. A role for soluble NSF attachment proteins (SNAPs) in regulated exocytosis in adrenal chromaffin cells. EMBO J. 1995b;14:232–239. doi: 10.1002/j.1460-2075.1995.tb06996.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nichols BJ, Ungermann C, Pelham HRB, Wickner WT, Haas A. Homotypic vacuolar fusion mediated by t- and v-SNAREs. Nature. 1997;387:199–202. doi: 10.1038/387199a0. [DOI] [PubMed] [Google Scholar]
- 42.Nie Z, Ranjan R, Wenninger JJ, Hong SN, Bronk P, Zinsmaier K. Overexpression of cysteine string proteins in Drosophila reveals interactions with syntaxin. J Neurosci. 1999;19:10270–10279. doi: 10.1523/JNEUROSCI.19-23-10270.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Otter-Nilsson M, Hendriks R, Pecheur-Huet E-I, Hoekstra D, Nilsson T. Cytosolic ATPases, p97 and NSF, are sufficient to mediate rapid membrane fusion. EMBO J. 1999;18:2074–2083. doi: 10.1093/emboj/18.8.2074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Otto H, Hanson PI, Jahn R. Assembly and disassembly of a ternary complex of synaptobrevin, syntaxin, and SNAP-25 in the membrane of synaptic vesicles. Proc Natl Acad Sci USA. 1997;94:6197–6201. doi: 10.1073/pnas.94.12.6197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Parsons TD, Coorssen JR, Horstmann H, Almers W. Docked granules, the exocytic burst, and the need for ATP hydrolysis in endocrine cells. Neuron. 1995;15:1085–1096. doi: 10.1016/0896-6273(95)90097-7. [DOI] [PubMed] [Google Scholar]
- 46.Phillips JH. Dynamic aspects of chromaffin granule structure. Neuroscience. 1982;7:1595–1609. doi: 10.1016/0306-4522(82)90017-3. [DOI] [PubMed] [Google Scholar]
- 47.Pupier S, Leveque C, Marqueze B, Kataoka M, Takahashi M, Seagar MJ. Cysteine string proteins associated with secretory granules of the rat neurohypophysis. J Neurosci. 1997;17:2722–2727. doi: 10.1523/JNEUROSCI.17-08-02722.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rizo J, Sudhof TC. Mechanics of membrane fusion. Nat Struct Biol. 1998;5:839–842. doi: 10.1038/2280. [DOI] [PubMed] [Google Scholar]
- 49.Schiavo G, Benfenati F, Poulain B, Rossetto O, Polverino de Laureto P, DasGupta BR, Montecucco C. Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature. 1992;359:832–835. doi: 10.1038/359832a0. [DOI] [PubMed] [Google Scholar]
- 50.Schulze KL, Littleton JT, Salzberg A, Halachmi N, Stern M, Lev Z, Bellen HJ. rop, a Drosophila homolog of yeast sec1 and vertebrate n-sec1/munc 18 proteins, is a negative regulator of neurotransmitter release in vivo. Neuron. 1994;13:1099–1108. doi: 10.1016/0896-6273(94)90048-5. [DOI] [PubMed] [Google Scholar]
- 51.Sollner T, Whiteheart SW, Brunner M, Erdjument-Bromage H, Geromanos S, Tempst P, Rothman JE. SNAP receptors implicated in vesicle targeting and fusion. Nature. 1993a;362:318–324. doi: 10.1038/362318a0. [DOI] [PubMed] [Google Scholar]
- 52.Sollner T, Bennett MK, Whiteheart SW, Scheller R, Rothman J. A protein assembly-disassembly pathway in vitro that may correspond to sequential steps of synaptic vesicle docking, activation, and fusion. Cell. 1993b;75:409–418. doi: 10.1016/0092-8674(93)90376-2. [DOI] [PubMed] [Google Scholar]
- 53.Sudhof TC. The synaptic vesicle cycle: a cascade of protein-protein interactions. Nature. 1995;375:645–653. doi: 10.1038/375645a0. [DOI] [PubMed] [Google Scholar]
- 54.Sutton RB, Fasshauer D, Jahn R, Brunger AT. Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4A resolution. Nature. 1998;395:347–353. doi: 10.1038/26412. [DOI] [PubMed] [Google Scholar]
- 55.Tagaya M, Toyonaga S, Takahashi M, Yamamoto A, Fujiwara T, Akagawa K, Moriyama Y, Mizushima S. Syntaxin 1 (HPC-1) is associated with chromaffin granules. J Biol Chem. 1995;270:15930–15933. doi: 10.1074/jbc.270.27.15930. [DOI] [PubMed] [Google Scholar]
- 56.Tolar LA, Pallanck L. NSF function in neurotransmitter release involves rearrangement of the SNARE complex downstream of synaptic vesicle docking. J Neurosci. 1998;18:10250–10256. doi: 10.1523/JNEUROSCI.18-24-10250.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Umbach JA, Zinsmaier KE, Eberle KK, Buchner E, Benzer S, Gundersen CB. Presynaptic dysfunction in Drosophila csp mutants. Neuron. 1994;13:899–907. doi: 10.1016/0896-6273(94)90255-0. [DOI] [PubMed] [Google Scholar]
- 58.Umbach JA, Saitoe M, Kidokoro Y, Gundersen CB. Attenuated influx of calcium ions at nerve endings of csp and shibire mutant Drosophila. J Neurosci. 1998;18:3233–3240. doi: 10.1523/JNEUROSCI.18-09-03233.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ungermann C, Nichols BJ, Pelham HRB, Wickner W. A vacuolar v-t-SNARE complex, the predominant form in vivo and on isolated vacuoles, is disassembled and activated for docking and fusion. J Cell Biol. 1998;140:61–69. doi: 10.1083/jcb.140.1.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Weber T, Zemelman BV, McNew JA, Westermann B, Gmachl M, Parlati F, Sollner TH, Rothman JE. SNAREpins: minimal machinery for membrane fusion. Cell. 1998;92:759–772. doi: 10.1016/s0092-8674(00)81404-x. [DOI] [PubMed] [Google Scholar]
- 61.Wightman RM, Jankowski JA, Kennedy RT, Kawagoe KT, Schroeder TJ, Leszczyszyn DJ, Near JA, Diliberto EJ, Jr, Viveros OH. Temporally resolved catecholamine spikes correspond to single vesicle release from individual chromaffin cells. Proc Natl Acad Sci USA. 1991;88:10754–10758. doi: 10.1073/pnas.88.23.10754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Winkler H. The biogenesis of adrenal chromaffin granules. Neuroscience. 1977;2:657–683. doi: 10.1016/0306-4522(77)90022-7. [DOI] [PubMed] [Google Scholar]
- 63.Winkler H, Fischer-Colbrie R. Regulation of the biosynthesis of large dense-core vesicles in chromaffin cells and neurons. Cell Mol Neurobiol. 1998;18:193–209. doi: 10.1023/A:1022516919932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Wu MN, Littleton JT, Bhat MA, Prokop A, Bellen HJ. ROP, the Drosophila sec1 homolog, interacts with syntaxin and regulates neurotransmitter release in a dosage-dependent manner. EMBO J. 1998;17:127–139. doi: 10.1093/emboj/17.1.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wu MN, Fergestad T, Lloyd TE, He Y, Broadie K, Bellen HJ. Syntaxin 1A interacts with multiple exocytic proteins to regulate neurotransmitter release in vivo. Neuron. 1999;23:593–605. doi: 10.1016/s0896-6273(00)80811-9. [DOI] [PubMed] [Google Scholar]
- 66.Xu T, Ashery U, Burgoyne RD, Neher E. Early requirement for α-SNAP and NSF in the secretory cascade in chromaffin cells. EMBO J. 1999;18:3293–3304. doi: 10.1093/emboj/18.12.3293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Zhang H, Kelley WL, Chamberlain LH, Burgoyne RD, Wollheim CB, Lang J. Cysteine string proteins regulate exocytosis of insulin independent from transmembrane ion fluxes. FEBS Lett. 1998;437:267–272. doi: 10.1016/s0014-5793(98)01233-2. [DOI] [PubMed] [Google Scholar]
- 68.Zhang H, Kelley WL, Chamberlain LH, Burgoyne RD, Lang J. Mutational analysis of cysteine-string protein function in insulin exocytosis. J Cell Sci. 1999;112:1345–1351. doi: 10.1242/jcs.112.9.1345. [DOI] [PubMed] [Google Scholar]
- 69.Zinsmaier KE, Hofbauer A, Heimbeck G, Pflugfeler GO, Buchner S, Buchner E. A cysteine-string protein is expressed in retina and brain of Drosophila. J Neurogenet. 1990;7:15–29. doi: 10.3109/01677069009084150. [DOI] [PubMed] [Google Scholar]
- 70.Zinsmaier KE, Eberle KK, Buchner E, Walter N, Benzer S. Paralysis and early death in cysteine string protein mutants of Drosophila. Science. 1994;263:977–980. doi: 10.1126/science.8310297. [DOI] [PubMed] [Google Scholar]