

Abstract
In mammals, the environmental light/dark cycle strongly synchronizes the circadian clock within the suprachiasmatic nuclei (SCN) to 24 hr. It is well known that not only photic but also nonphotic stimuli can entrain the SCN clock. Actually, many studies have shown that a daytime injection of 8-hydroxy-2-(di-n-propylamino) tetralin (8-OH DPAT), a serotonin 1A/7 receptor agonist, as a nonphotic stimulus induces phase advances in hamster behavioral circadian rhythmsin vivo, as well as the neuron activity rhythm of the SCN in vitro. Recent reports suggest that mammalian homologs of the Drosophila clock gene,Period (Per), are involved in photic entrainment. Therefore, we examined whether phase advances elicited by 8-OH DPAT were associated with a change of Period mRNA levels in the SCN. In this experiment, we cloned partial cDNAs encoding hamster Per1, Per2, andPer3 and observed both circadian oscillation and the light responsiveness of Period. Furthermore, we found that the inhibitory effect of 8-OH DPAT on hamster Per1and Per2 mRNA levels in the SCN occurred only during the hamster's mid-subjective day, but not during the early subjective day or subjective night. The present findings demonstrate that the acute and circadian time-dependent reduction of Per1 and/orPer2 mRNA in the hamster SCN by 8-OH DPAT is strongly correlated with the phase resetting in response to 8-OH DPAT.
Keywords: suprachiasmatic nucleus, 8-OH DPAT, Per mRNA, 5-HT1A/7 receptor, hamster, circadian rhythm, NIH Image
In mammals, the suprachiasmatic nucleus (SCN) of the hypothalamus has been shown to be a primary circadian pacemaker of locomotor activity and various physiological phenomena (Hastings, 1997). Recent studies on the molecular aspects of clock genes have produced a functional model of circadian rhythms (for review, see Dunlap, 1999).
mPer1, mPer2, and mPer3, cloned as mouse homologs of the Drosophila clock gene,Period (Per), exhibit circadian rhythmic expressions in the SCN (Albrecht et al., 1997; Shearman et al., 1997;Sun et al., 1997; Tei et al., 1997; Takumi et al., 1998a,b; Zylka et al., 1998). Brief exposure to light during subjective night results in a large and rapid induction of mPer1 expression (Albrecht et al., 1997; Shigeyoshi et al., 1997). mPer2 mRNA expression in the SCN is also induced in response to light stimuli (Shearman et al., 1997; Takumi et al., 1998a). On the other hand, mPer3mRNA levels do not respond to light during either the subjective night or subjective day (Takumi et al., 1998b; Zylka et al., 1998). Recently, we demonstrated that light-induced phase delays in locomotor activity at CT16 were significantly inhibited when mice were pretreated withmPer1 antisense phosphorothioate oligodeoxynucleotide (ODN) (Akiyama et al., 1999). Therefore, we suggest that the gated expression of mPer1 may be an important step in causing photic entrainment.
On the other hand, nonphotic manipulation such as novel wheel-running (Reebs and Mrosovsky, 1989), social interaction (Mrosovsky, 1988), and saline injection and/or handling (Mead et al., 1992) reportedly causes big phase advances in the hamster circadian clock when performed during subjective day. Additionally, many studies have shown that a daytime injection of 8-hydroxy-2-(di-n-propylamino) tetralin (8-OH DPAT), a serotonin 1A/7 receptor agonist, induces a phase advance in hamster behavioral circadian rhythms in vivo (Tominaga et al., 1992;Edgar et al., 1993; Cutrera et al., 1996; Mintz et al., 1997), as well as the neuron activity rhythm of the SCN in vitro (Shibata et al., 1992; Prosser et al., 1993). Thus, serotonin (5-HT) has been implicated in phase shifts of the circadian system during subjective day in response to nonphotic stimuli. Because light exposure inducesmPer1 and mPer2 expression during subjective night, we questioned whether injection of 8-OH DPAT modifies thePer mRNA levels during subjective day. Almost all behavioral experiments investigating nonphotic-induced phase advances were performed using hamsters, so we attempted to clone partial cDNAs encoding the golden hamster Per1, Per2, andPer3. Therefore, we first established that oscillations and light responses of hamster Per gene mRNA were similar to what has previously been published in the mouse. Second, we found that injection of 8-OH DPAT at CT6, but not at CT1 or CT20, reduced the amount of Per1 and Per2, but not Per3mRNA in the hamster SCN. Third, we demonstrated that nonphotic phase shifting with 8-OH DPAT is strongly correlated with an 8-OH DPAT-dependent, transient decrease in Per1 andPer2 mRNA levels.
MATERIALS AND METHODS
Cloning of partial cDNAs encoding hamster Pergenes. For analysis of Per gene expression by in situ hybridization, we attempted to clone partial cDNAs encoding the golden hamster Per genes. Total RNA was extracted from the golden hamster brain using Trizol (BRL, Bethesda, MD) and was reverse-transcribed using the Superscript one-step RT-PCR system (BRL). RT-PCR was performed using a DNA Thermal Cycler 9600 (Perkin-Elmer, Norwalk, CT) with specific primers derived from mouse and human sequences of Per genes. The sequences of the primers were as follows: Per2 (nucleotide position 822–1601 ofmPer2; GenBank accession number AF035830): 5′-ACACCACCCCTTACAAGCTTCC-3′, 5′-CGCTGGATGATGTCTGGCTC-3′;Per3 (nucleotide position 1956–2754 of mPer3; GenBank accession number AF050182): 5′-GAACTGTATCGACAGTGTCATC-3′, 5′-GGCCATATCTTGGAGGGGAAA-3′. The PCR protocol was executed under the following conditions: cDNA synthesis and predenaturation at 50°C for 30 min followed by 94°C for 2 min, PCR amplification for 35 cycles with denaturation at 94°C for 15 sec, annealing at 55°C for 30 sec, extension at 72°C for 1 min, and final extension at 72°C for 5 min. These PCR products were subcloned into the pGEM-T Easy Vector (Promega, Madison, WI) and were sequenced using ABI PRISM Dye Terminator Cycle Sequencing Ready Reaction Kit (Perkin-Elmer). A partial cDNA encoding hamsterPer1 corresponding to the nucleotide position 726–1367 ofmPer1 (GenBank accession number AB002108) was also cloned (S. Yamamoto and H. Okamura, data not shown).
Animals. Male golden hamsters (Mesocricetus auratus, Tokyo Laboratory Animals Science Co. Ltd., Tokyo, Japan) purchased 6 weeks postpartum were maintained on a 12 hr light/dark cycle with lights on at 8:30 A.M. (room temperature at 23 ± 2°C). Animals were given food and water ad libitum. For assessment of wheel-running activity, hamsters were housed individually in transparent plastic cages (36 × 20 × 20 cm) equipped with a running wheel (13 cm in diameter) that closed a microswitch with each revolution. The number of wheel rotations was measured, and data were stored on a personal computer.
Behavioral experiment. Because 8-OH DPAT has an asymmetrical carbon, this compound has two optical isomers: R(+) andS(−) 8-OH DPAT. A few studies have investigated the relationship between binding affinity of 5-HT1Aand 5-HT7 receptors and the effect of each optical isomer on circadian rhythms (Lovenberg et al., 1993; Eriksson and Evrin, 1996; Miller et al., 1996; Ying and Rusak, 1997). After free-running for 14–20 d in constant darkness, hamsters were randomly assigned to an intraperitoneal injection of 8-OH DPAT (1.0 or 5.0 mg/kg; Research Biochemicals, Natick, MA), (+) 8-OH DPAT (2.5 mg/kg, Research Biochemicals), triazolam (20 mg/kg, Upjohn), or vehicle [sterilized saline for both 8-OH DPAT and (+) 8-OH DPAT or dimethyl sulfoxide (Wako) for triazolam]. Injection was performed at circadian time (CT; CT12: onset time of wheel-running activity) 1, CT6, CT8, CT14, or CT20. Animals were then returned to their individual cages. The phase of the rhythm was assessed visually by applying a straight edge to the onset of activity on successive days before and after drug injection and determining the difference in phases on the day of drug injection (Daan and Pittendrigh, 1976).
In situ hybridization using digoxigenin. In situ hybridization using digoxigenin (DIG) was applied to determine the semiquantity or histochemical distribution ofPer mRNA levels in coronal sections of the hypothalamus. Hamsters were entrained to the light/dark cycle for at least 14 d and then kept in constant dark conditions. On the third day of constant darkness at CT6 or CT20, hamsters were intraperitoneally injected with each drug and then deeply anesthetized with ether 1, 2, or 4 hr after injection and intracardially perfused with 0.1 mphosphate buffer (PB) containing 4% paraformaldehyde (PFA). In some cases, hamster brains were obtained for observation of circadian changes in Per gene expression in the SCN. To make this observation, light (60 lux, 15 min) was applied at CT14 or CT20, and hamsters were killed 1 or 2 hr after the initiation of light exposure. Brains were removed, post-fixed in 0.1 m PB containing 4% PFA for 24 hr at 4°C, and transferred into 20% sucrose in PBS for 24 hr at 4°C. Frontal sections (40 μm thick) were collected and placed in PBS for 30 min, followed by treatment with 6 × SSC for 30 min. Sections were incubated in hybridization buffer [50% formamide, 6 × SSC, 0.1 mg/ml denatured salmon sperm DNA, 1 × Denhardt's solution (0.02% Ficoll, 0.02% polyvinyl pyrolidone, 0.02% bovine serum albumin), and 10% dextran sulfate] containing labeled cRNA probes overnight at 60°C. DIG-UTP (Roche Molecular Biochemicals, Indianapolis, IN)-labeled antisense cRNA was made using a standard protocol for cRNA synthesis. After hybridization, these sections were rinsed in 2 × SSC/50% formamide for 45 min followed by 15 min at 60°C and treated with RNase A for 30 min at 37°C, 2 × SSC/50% formamide for 2 × 15 min at 60°C, and 0.4 × SSC for 30 min at 60°C. Sections were processed for immunocytochemistry by following the DIG nucleic acid detection kit (Roche Molecular Biochemicals) protocol. Photomicrographs were taken with a Fujix digital camera (HC-300, Fujifilm, Tokyo Japan) and captured with photograb-300 (Fujifilm). The density of Per gene expression was semiquantified on a Macintosh computer using the public domain NIH Image program (written by Wayne Rasband, National Institutes of Health).
In situ hybridization using radioisotope. In situ hybridization using radioisotope (RI)-labeled probes was applied to determine the quantity of Per1, Per2, and Per3 mRNA levels in coronal sections of the hypothalamus. Hamsters were entrained to the light/dark cycle for at least 14 d and then kept in constant dark conditions. On the third day of constant darkness at CT1, CT6, or CT20, hamsters were intraperitoneally injected with each drug and then deeply anesthetized for 2 hr after injection and intracardially perfused with 0.1m PB containing 4% PFA. Brains were removed, post-fixed in 0.1 m PB containing 4% PFA for 24 hr at 4°C, and transferred into 20% sucrose in PBS for 24 hr at 4°C. Frontal sections (30 μm thick) were collected and placed in 2 × SSC and then treated with proteinase K (1.0 μg/ml, 10 mm Tris buffer, pH 7.5, 10 mm EDTA) for 10 min at 37°C, 4% PFA in 0.1m PB for 5 min, and 2 × SSC for 5 min followed by 0.25% acetic anhydride in 0.1 mtriethanolamine for 10 min and 2 × SSC for 2 × 5 min. RI [α[33P]UTP (New England Nuclear)]-labeled antisense cRNA was made using a standard protocol for cRNA synthesis. Hybridization and posthybridization washing steps were the same as the protocol for DIG in situ hybridization. RI in situ hybridization images were visualized by autoradiogram and BioMax film (Kodak) and analyzed using an image analyzing system (MCID, Imaging Research Inc.) after conversion into optical density by 14C-autoradiographic microscales (Amersham, Arlington Heights, IL). The values were expressed as means ± SEM. For statistical analysis, one-way ANOVA followed by the Student's t test was applied.
RESULTS
Oscillation and light response of SCN PermRNA expression
To evaluate the topographic differences of expression profile in hamster Per genes of the SCN, in situhybridizations using digoxigenin-labeled probes of hamster Per1, Per2, and Per3 were performed. DIG in situhybridization showed clear signals with the antisense probes in the hamster SCN (Fig. 1). These antisense probe signals were suppressed by competition experiments using the unlabeled antisense probes. Furthermore, the sense probes demonstrated specificity of the antisense hybridization by exhibiting no signals within the SCN (data not shown). The distribution or expression patterns of hamster Per mRNA inside and outside the SCN were consistent with those of already published findings in the mouse (Albrecht et al., 1997; Shearman et al., 1997; Sun et al., 1997; Tei et al., 1997; Takumi et al., 1998a,b; Zylka et al., 1998). Next, we examined the circadian pattern of Per gene expression in the hamster SCN. There were clear circadian rhythms of Per1,Per2, and Per3 expression in the hamster SCN with a peak at CT4 for Per1 and at CT8 for both Per2and Per3 (Fig. 1). In nocturnal rodents, it is well established that light pulses administered during the early subjective night cause phase delays of the circadian rhythm, whereas pulses delivered during the latter half of the subjective night cause phase advances. Recent reports indicated that mPer1 andmPer2 expression in the mouse SCN was increased rapidly and transiently after brief light exposure during both early and late subjective night, whereas mPer3 expression was not (Takumi et al., 1998b; Zylka et al., 1998). We also observed thatPer1 and Per2 expression in the hamster SCN was induced in response to brief light exposure at CT14 or CT20, andPer3 expression was not affected by light stimulation at CT20 (Fig. 2). In both Per1and Per2 cases, we found that signal intensities were more likely to be stronger in the ventrolateral part of the SCN than in the dorsomedial part. These results were consistent with previous data found in the mouse.
Fig. 1.
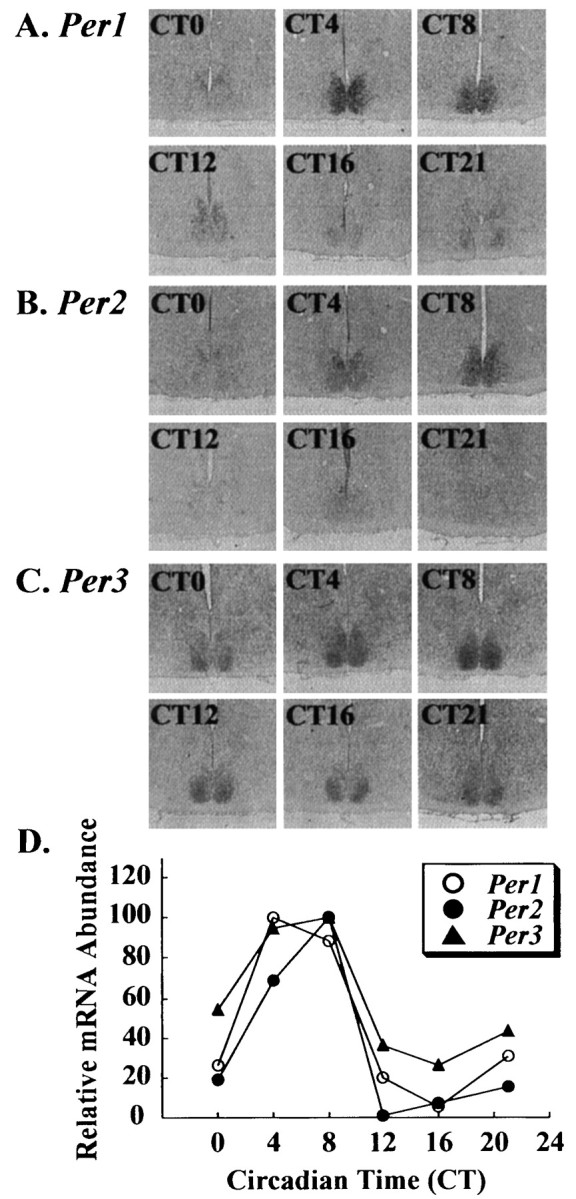
Circadian expressions of (A)Per1, (B) Per2, and (C) Per3 in the hamster suprachiasmatic nucleus. mRNA expression was demonstrated by DIGin situ hybridization and semiquantified by a Macintosh computer using the public domain NIH Image program (D). There are clear circadian rhythms ofPer1, Per2, and Per3expression in the hamster SCN with a peak at CT4 forPer1 and at CT8 for both Per2 andPer3.
Fig. 2.
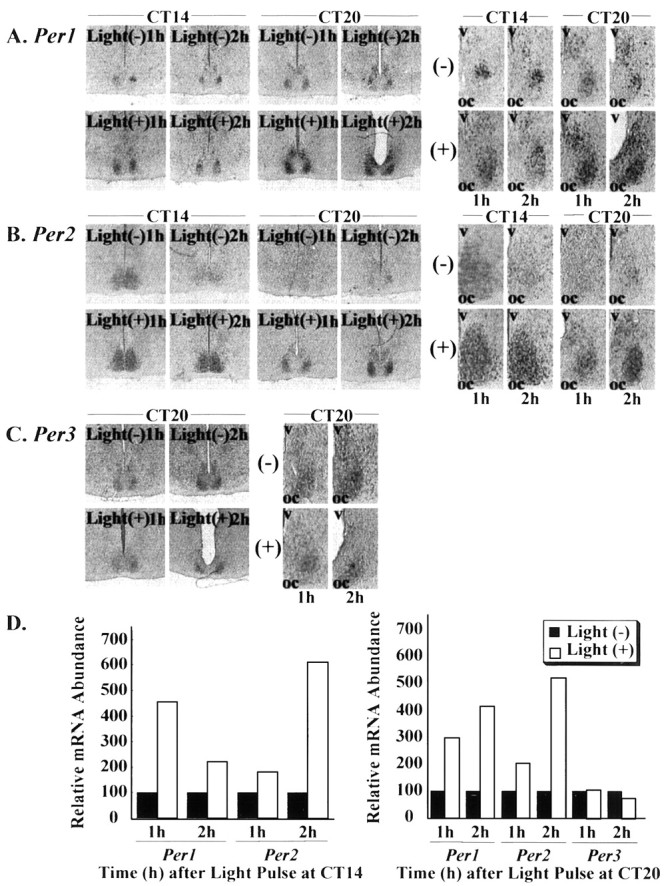
Effect of light exposure on the expression of (A) Per1, (B) Per2, and (C) Per3 in the hamster suprachiasmatic nucleus. mRNA expression was demonstrated by DIGin situ hybridization, and a more magnified picture is shown. Per mRNA expression was semiquantified by a Macintosh computer using the public domain NIH Image program (D). Hamster was exposed to light (60 lux, 15 min) at CT14 or CT20 and then killed 60 or 120 min after light pulse. Light at CT14 or CT20 induced Per1 andPer2 expression in the SCN; however, Per3expression was not affected by light exposure at CT20.oc, Optic chiasma; v, third ventricle;(+), light pulses were applied;(−), light pulses were not applied.
Wheel-running activity in response to 8-OH DPAT
Vehicle administration at CT6 did not show any change in phase (Fig. 3A); however, 8-OH DPAT (5.0 mg/kg) administration at CT6 produced a clear phase advance (Fig.3B). Administration of this compound at CT20 did not affect the wheel-running rhythm (Fig. 3C). Administration of 8-OH DPAT (5.0 mg/kg) at various CTs (CT1, CT6, CT8, CT14, CT20) was compared with vehicle administration (Fig. 3D). Significant phase advances were observed when this compound was administered at CT6 or CT8. There were no significant differences at other CTs. Administration of triazolam (20 mg/kg) at CT6 also caused a phase advance in hamster wheel-running rhythm (data not shown).
Fig. 3.
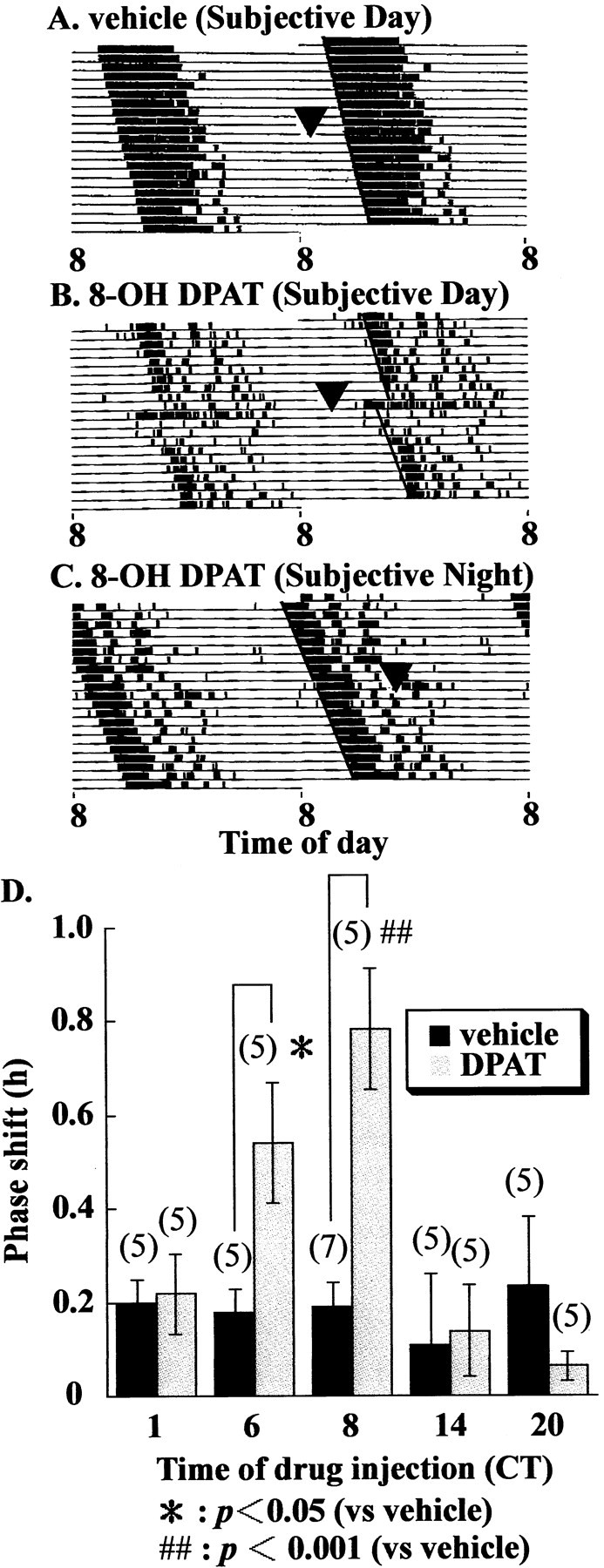
Effects of 8-OH DPAT administration on the hamster circadian wheel-running rhythm. Double-plotted actogram shows wheel-running activity records of (A) vehicle and (B) 8-OH DPAT (5.0 mg/kg, i.p.)-injected hamster at CT6, and (C) 8-OH DPAT (5.0 mg/kg, i.p.) injection at CT20. Each animal was injected at CT6 or CT20 (arrowheads) and returned to constant darkness.D, Mean phase advances induced by 8-OH DPAT (5.0 mg/kg, i.p.) administration at CT1, CT6, CT8, CT14, and CT20.Numbers in parentheses indicate the number of experiments. Injection of 8-OH DPAT at CT6 or CT8 induced a significant phase advance (*p < 0.05,##p < 0.001, Student'st test).
Effect of 8-OH DPAT on SCN Per mRNA
Administration of 8-OH DPAT (5.0 mg/kg) reduced the levels ofPer1 in the entire SCN and Per2 preferentially in the ventrolateral part of the SCN (Fig.4A,B). Four hours after drug injection, however, reduced Per1 andPer2 levels recovered to the control level of vehicle-treated animals. Injection of 8-OH DPAT at CT20 did not change the Per1 and Per2 levels in the SCN. (+) 8-OH DPAT (2.5 mg/kg)-induced reduction of Per1 andPer2 was similar to 8-OH DPAT (5.0 mg/kg)-induced reductions (Fig. 4A,B). Injection of 8-OH DPAT did not reduce the Per3 mRNA levels (Fig. 4C). For quantitative measurements of Per mRNA levels, in situ hybridizations using RI-labeled probes were performed. In Figure 5, the mean values forPer1, Per2, and Per3 in the hamster SCN of 8-OH DPAT and vehicle-treated groups at various CTs are shown. Vehicle-treated hamsters exhibit clear circadian rhythms ofPer1 (ANOVA, F(2,12) = 9.363, p = 0.0032), Per2 (ANOVA,F(2,13) = 62.375, p = 0.0001), and Per3 (ANOVA,F(2,13) = 39.892, p = 0.0001) expression in the SCN. High expression of these genes was seen at CT6. Two hours after the administration of 8-OH DPAT (5.0 mg/kg) at CT6, the amount of both Per1 and Per2 mRNA was significantly reduced in comparison with the group receiving vehicle treatment (Fig. 5A,B). On the other hand, injection of 8-OH DPAT at CT1 or CT20 did not affect the amount of Per1 and Per2 mRNA. Interestingly, this drug did not change the Per3 mRNA of the SCN at any CTs. In the next experiment, we examined whether 8-OH DPAT reduced Per1and Per2 mRNA in a dose-dependent manner (Fig.6). Administration of 8-OH DPAT caused a phase advance in hamster wheel-running rhythm in a dose-dependent manner (Fig. 6A), and this compound also reduced the amount of both Per1 and Per2 mRNA in a dose-related fashion (Fig. 6B,C). Thus, the effective dose for behavioral phase shift induction and reduction of Per1 and Per2 was very similar. Amplitude of Per1 and Per2 reduction by (+) 8-OH DPAT (5.0 mg/kg) was similar to that of (+) 8-OH DPAT (2.5 mg/kg) (Fig.6). Administration of triazolam (20 mg/kg), a central-type benzodiazepine receptor ligand, at CT 6 also reduced the amount of SCNPer1 and Per2 but not Per3 mRNA.
Fig. 4.
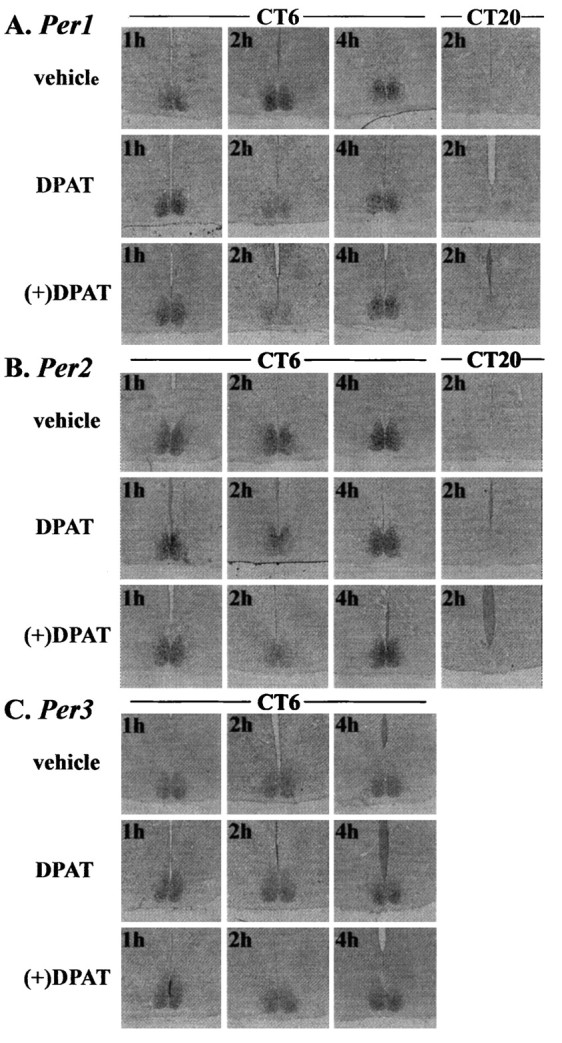
Effects of 8-OH DPAT (5.0 mg/kg, i.p.) or (+) 8-OH DPAT (2.5 mg/kg, i.p.) on the expression of (A)Per1, (B) Per2, and (C) Per3 in the hamster suprachiasmatic nucleus. mRNA levels were demonstrated by DIG in situ hybridization. Drug was administered at CT6 or CT20, andPer mRNA levels were examined 60–240 min after drug injection. 8-OH DPAT reduced Per1 andPer2 mRNA levels in the SCN 120 min after injection at CT6, but mRNA levels returned to the control level 240 min after injection. Injection of 8-OH DPAT at CT20 did not affectPer1 or Per2 mRNA levels. The amount ofPer3 mRNA was not affected by injection of 8-OH DPAT.
Fig. 5.

Effect of 8-OH DPAT at various CTs onPer1, Per2, and Per3expression in the hamster suprachiasmatic nucleus. RI in situ hybridization was performed for the quantitative analysis. Circadian expression of these genes was observed at (A) Per1 (ANOVA,F(2,12) = 9.363,p = 0.0032), (B)Per2 (ANOVA, F(2,13) = 62.375, p = 0.0001), and (C)Per3 (ANOVA,F(2,13) = 39.892,p = 0.0001). 8-OH DPAT (5.0 mg/kg, i.p.) significantly reduced Per1(#p < 0.005, Student'st test) and Per2(∫∫p < 0.0001, Student'st test) but not Per3 mRNA levels 2 hr after drug injection at CT6. Numbers inparentheses indicate the number of experiments.
Fig. 6.
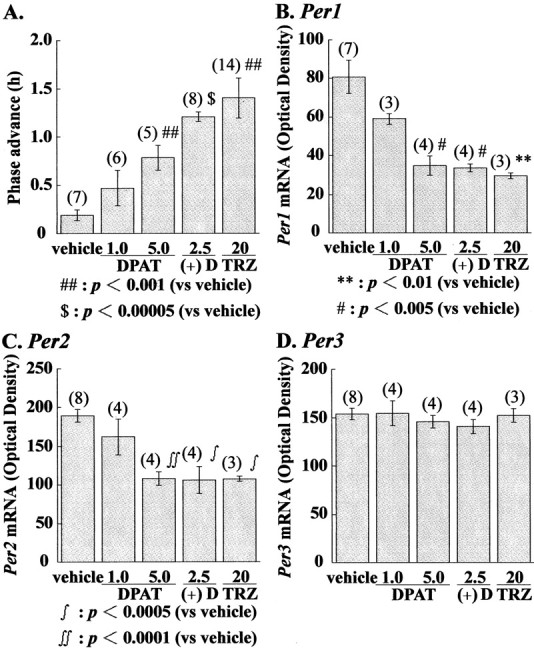
Dose-dependent advance of behavioral rhythms (A) and dose-dependent reduction ofPer1 (B), Per2(C), and Per3(D) mRNA levels by 8-OH DPAT (1.0 or 5.0 mg/kg, i.p.), (+) 8-OH DPAT (2.5 mg/kg, i.p.), or triazolam (20 mg/kg, i.p.). mRNA levels were quantified using RI in situhybridization. **p < 0.01,#p < 0.005,##p < 0.001,∫p < 0.0005,∫∫p < 0.0001,$p < 0.00005 versus vehicle alone, Student's t test. Numbersin parentheses indicate the number of experiments.DPAT, 8-OH DPAT; (+)D, (+) 8-OH DPAT;TRZ, triazolam.
DISCUSSION
In situ hybridizations using digoxigenin-labeled probes of hamster Per genes revealed clear circadian expressions of Per1, Per2, and Per3 in the hamster SCN. The pattern of expression of these genes is very similar to that observed in the mouse (Albrecht et al., 1997; Shearman et al., 1997; Sun et al., 1997; Tei et al., 1997; Takumi et al., 1998a,b; Zylka et al., 1998) and rat (Sakamoto et al., 1998; Yan et al., 1999) SCN, with peak expression of hamster Per1,Per2, and Per3 found at CT4, CT8, and CT8, respectively. Reportedly, light exposure during subjective night causes a rapid induction of mPer1 and mPer2 in the SCN (Albrecht et al., 1997; Shearman et al., 1997; Takumi et al., 1998a). In the hamster, we also observed that Per1 andPer2 have light-responsive characters similar to mouseperiod genes.
Anatomical subdivisions such as the ventrolateral and dorsomedial parts in the rat SCN have been well established (Moore, 1982). Recently, we reported that light exposure at CT16 induced the expression of ratPer1 and Per2 in SCN neurons of the ventrolateral part, although the circadian rat Per1 and Per2mRNA oscillations in light/dark and constant dark conditions occurred strongly in neurons in the dorsomedial part but weakly in neurons in the ventrolateral part of the SCN (Yan et al., 1999). In the hamster SCN, light-induced expression of Per1 and Per2was preferential to the ventrolateral part of the SCN. Although the basic compartment profile of Per gene expression may be preserved in the hamster SCN, however, anatomical subdivision of the hamster SCN is vague compared with the rat SCN. Therefore, further experiments are needed to elucidate the compartmentalization of circadian profiles in Per genes in the hamster in detail.
In the present experiment, we demonstrated that 8-OH DPAT reduces SCNPer1 and Per2 mRNA in a circadian time-dependent manner. Actually, 8-OH DPAT reduced these gene mRNAs when administered at CT6 but not at CT1 or CT20. This result relates to the behavioral result showing a large phase advance at CT6 and CT8 but not at CT1 or CT20. In addition, 8-OH DPAT reduced Per1 andPer2 in a dose-dependent fashion. The dose and threshold closely correlated with Per1 and Per2 reduction in the SCN. mPer1 and mPer2 transcription is rapidly induced by light in a time-of-day-dependent manner (Shigeyoshi et al., 1997; Takumi et al., 1998b). Gating is known to play a role in the light-induced phase shifts of behavioral rhythms. The responsiveness of mPer1 mRNA to light is closely related to behavioral phase delays induced by light (Shigeyoshi et al., 1997). In addition, we previously demonstrated that light-induced phase delays in locomotor activity at CT16 were significantly inhibited when the mice were pretreated with mPer1 antisense ODN before light exposure (Akiyama et al., 1999). Therefore, the gated induction ofmPer1 is a step necessary for producing behavioral phase shifts. These results along with our present results suggest that gated inhibition of Per1 and/or Per2 expression by nonphotic stimulation may facilitate Per gene reduction resulting in the onset of the next circadian induction ofPer gene expression.
(+) 8-OH DPAT exhibited a higher affinity for 5-HT7 receptors than for 5-HT1A receptors, and amplitude ofPer1 and Per2 reduction by 8-OH DPAT (5 mg/kg) was similar to that of (+) 8-OH DPAT (2.5 mg/kg). Thus, we estimate that the potential effect of (+) 8-OH DPAT is two times higher than that of 8-OH DPAT. The present results suggest that 5-HT7 receptors, rather than 5-HT1A receptors, have a more important role in phase shifting, as pointed out by Ying and Rusak (1997) during investigation of the inhibitory effect of 5-HT7receptors on light-sensitive SCN neurons. Challet et al. (1998)reported that bilateral 8-OH DPAT injections into either the SCN or the intergeniculate leaflet cause significant phase advances in hamster wheel-running activity. Additional studies proposed that perfusion of 8-OH DPAT at CT6–CT8 advances neuron activity rhythm of the SCNin vitro (Shibata et al., 1992; Prosser et al., 1993). These reports prefer the direct action of 8-OH DPAT on SCN Pergene expression. On the other hand, Schuhler et al. (1998) demonstrated that the 5-HT fibers connecting the median raphe to the SCN are essential for the phase-shifting action of peripheral 8-OH DPAT injections into the SCN using microinjections of 5-HT neurotoxin. The present results demonstrate that 8-OH DPAT reduces Per1 mRNA levels in the entire SCN and preferentially reduces Per2mRNA in the ventrolateral part of the SCN. Because serotonergic fibers from the median raphe nucleus innervate the ventrolateral part of the hamster SCN (Meyer-Bernstein and Morin, 1996; Leander et al., 1998), the reduction of this SCN serotonergic input may be one of the possible outcomes of 8-OH DPAT-induced reductions of Per mRNA. It is interesting that triazolam (20 mg/kg) causes not only a big phase advance but also a strong inhibition of Per1 andPer2 mRNA. Thus, reduction of Per1 andPer2 mRNA correlates well with phase advances induced by 8-OH DPAT as well as triazolam.
The SCN entrains to the environmental light/dark cycle via a retinal projection called the retinohypothalamic tract (RHT). Glutamate, which acts as an RHT transmitter (de Vries et al., 1993), and glutamate and NMDA application to the rat SCN in vitro reportedly causes phase delays in SCN firing rhythms when applied during early subjective night (Ding et al., 1994; Shibata et al., 1994; Shirakawa and Moore, 1994; Ding et al., 1997). Excitation of glutamate receptors is reported to facilitate the phosphorylation of cAMP response element binding protein (CREB) (Ding et al., 1997; McNulty et al., 1998). Furthermore, light exposure at night is reported to produce mitogen-activated protein kinase phosphorylation and CREB phosphorylation (Obrietan et al., 1998). Thus, the signal cascade of photic entrainment is well documented, whereas the signal cascade of nonphotic entrainment is obscure at present. Currently, we do not know the mechanism ofPer1 and Per2 reduction in the SCN by 8-OH DPAT or triazolam.
Treatments using pituitary adenylate cyclase-activating polypeptide or cAMP during subjective day are reported to induce the phase shift of circadian rhythm, apparently via activation of adenylate cyclase and PKA activity (Prosser and Gillette, 1989; Hannibal et al., 1997;Harrington et al., 1999). 5-HT7 receptors are positively coupled to adenylate cyclase (Lovenberg et al., 1993; Tsou AP et al., 1994), and activation of both PKA and K+ channels is necessary for 5-HT-induced phase advances of circadian rhythm (Prosser et al., 1994). Therefore, we speculate that activation of PKA may be involved in the 8-OH DPAT-induced phase advance and transient reduction of PermRNA levels.
In this experiment, we demonstrated that administration of triazolam and 8-OH DPAT during subjective day reduces Per1 andPer2 mRNA in the hamster SCN. Nonphotic resetting by benzodiazepine or novel wheel-running requires neuropeptide Y innervation of the SCN from the thalamus (Biello et al., 1994; Maywood et al., 1997). Recently, Maywood et al. (1999) also demonstrated the acute downregulation of SCN Per1 and Per2, whereas there was no significant change in SCN PER1 immunoreactivity by novel wheel-running during the daytime under a light/dark cycle. Our present results are highly consistent with the data of Maywood et al. (1997, 1999). Although we do not know whether the signal transduction mechanism of benzodiazepine-GABA, 8-OH DPAT, and novel wheel-running are identical, it is strongly suggested that nonphotic stimuli presented during subjective day cause a phase advance through the reduction of Per1 and Per2 mRNA in the hamster SCN. However, further experiments are needed to investigate the response of other clock elements such as Clock,Bmal1, Cry1, Cry2 (Gekakis et al., 1998; Thresher et al., 1998; Griffin et al., 1999; Jin et al., 1999;Kume et al., 1999; Miyamoto and Sancar, 1999; Okamura et al., 1999; van der Horst et al., 1999; Vitaterna et al., 1999), or otherPer proteins in the hamster SCN after nonphotic stimuli, because these assessments will better clarify the cascading effects of nonphotic stimuli on the clock loop.
The levels of mPer3 mRNA are not affected by light exposure (Takumi et al., 1998b; Zylka et al., 1998). In the present experiment,Per3 mRNA levels in the hamster SCN were unresponsive to light exposure at CT20 or to administration of 8-OH DPAT at various CTs. Triazolam also did not affect Per3 mRNA levels. We do not know why Per3 mRNA levels in the SCN are insensitive to both photic and nonphotic stimuli. It could be that acute changing of the expression of Per3 is not required for phase shifts in mice and hamster circadian rhythms.
In summary, we found that the acute and circadian time-dependent reduction of Per1 or Per2 mRNA, or both, in the hamster SCN by 8-OH DPAT strongly correlates with the phase resetting in response to 8-OH DPAT. Therefore, the present findings suggest that nonphotic shifts involve alteration in Per1 orPer2 mRNA levels, or both, in the SCN.
Footnotes
This study was partially supported by grants awarded to S.S. from the Research Project for the Future Program (RFTF96L00310), the Japanese Ministry of Education, Science, Sports, and Culture (11170248, 1123207, 11145240), and The Special Coordination Funds of the Japanese Science and Technology Agency.
Correspondence should be addressed to Shigenobu Shibata, Department of Pharmacology and Brain Science, School of Human Sciences, Waseda University, Tokorozawa, Saitama 359-1192, Japan. E-mail address:shibata@human.waseda.ac.jp.
REFERENCES
- 1.Akiyama M, Kouzu Y, Takahashi S, Wakamatsu H, Moriya T, Maetani M, Watanabe S, Tei H, Sakaki Y, Shibata S. Inhibition of light- or glutamate-induced mPer1 expression represses the phase shifts into the mouse circadian locomotor and suprachiasmatic firing rhythms. J Neurosci. 1999;19:1115–1121. doi: 10.1523/JNEUROSCI.19-03-01115.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Albrecht U, Sun ZS, Eichele G, Lee CC. A differential response of two putative mammalian circadian regulators, mPer1 and mPer2, to light. Cell. 1997;91:1055–1064. doi: 10.1016/s0092-8674(00)80495-x. [DOI] [PubMed] [Google Scholar]
- 3.Biello SM, Janik D, Mrosovsky N. Neuropeptide Y and behaviorally induced phase shifts. Neuroscience. 1994;62:273–279. doi: 10.1016/0306-4522(94)90331-x. [DOI] [PubMed] [Google Scholar]
- 4.Challet E, Scarbrough K, Penev PD, Turek FW. Roles of suprachiasmatic nuclei and intergeniculate leaflets in mediating the phase-shifting effects of a serotonergic agonist and their photic modulation during subjective day. J Biol Rhythms. 1998;13:410–421. doi: 10.1177/074873098129000237. [DOI] [PubMed] [Google Scholar]
- 5.Cutrera RA, Saboureau M, Pevet P. Phase-shifting effect of 8-OH DPAT, a 5-HT1A/5-HT7 receptor agonist, on locomotor activity in golden hamster in constant darkness. Neurosci Lett. 1996;210:1–4. doi: 10.1016/0304-3940(96)12655-0. [DOI] [PubMed] [Google Scholar]
- 6.Daan S, Pittendrigh C. A functional analysis of circadian pacemakers in nocturnal rodents II. The variability of phase response curves. J Comp Physiol [A] 1976;106:253–266. [Google Scholar]
- 7.de Vries MJ, Nunes Cardozo B, van der Want J, de Wolf A, Meijer JH. Glutamate immunoreactivity in terminals of the retinohypothalamic tract of the brown Norwegian rat. Brain Res. 1993;612:231–237. doi: 10.1016/0006-8993(93)91665-f. [DOI] [PubMed] [Google Scholar]
- 8.Ding JM, Chen D, Weber ET, Faiman LE, Rea MA, Gillette MU. Resetting the biological clock: mediation of nocturnal circadian shifts by glutamate and NO. Science. 1994;266:1713–1717. doi: 10.1126/science.7527589. [DOI] [PubMed] [Google Scholar]
- 9.Ding JM, Faiman LE, Hurst WJ, Kuriashkina LR, Gillette MU. Resetting the biological clock: mediation of nocturnal CREB phosphorylation via light, glutamate, and nitric oxide. J Neurosci. 1997;17:667–675. doi: 10.1523/JNEUROSCI.17-02-00667.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dunlap JC. Molecular bases for circadian clocks. Cell. 1999;96:271–290. doi: 10.1016/s0092-8674(00)80566-8. [DOI] [PubMed] [Google Scholar]
- 11.Edgar DM, Miller JD, Prosser R, Dean RR, Dement WC. Serotonin and the mammalian circadian system: II. Phase-shifting rat behavioral rhythms with serotonergic agonists. J Biol Rhythms. 1993;8:17–31. doi: 10.1177/074873049300800102. [DOI] [PubMed] [Google Scholar]
- 12.Eriksson H, Evrin K. An evaluation of the 5-HT7 receptor mediated signal transduction pathway in a transfected cell line by using the cytosensor microphysiometer. Soc Neurosci Abstr. 1996;22:1782. [Google Scholar]
- 13.Gekakis N, Staknis D, Nguyen HB, Davis FC, Wilsbacher LD, King DP, Takahashi JS, Weitz CJ. Role of the CLOCK protein in the mammalian circadian mechanism. Science. 1998;280:1564–1569. doi: 10.1126/science.280.5369.1564. [DOI] [PubMed] [Google Scholar]
- 14.Griffin EA, Jr, Staknis D, Weitz CJ. Light-independent role of CRY1 and CRY2 in the mammalian circadian clock. Science. 1999;286:768–771. doi: 10.1126/science.286.5440.768. [DOI] [PubMed] [Google Scholar]
- 15.Hannibal J, Ding JM, Chen D, Fahrenkrug J, Larsen PJ, Gillette MU, Mikkelsen JD. Pituitary adenylate cyclase-activating peptide (PACAP) in the retinohypothalamic tract: a potential daytime regulator of the biological clock. J Neurosci. 1997;17:2637–2644. doi: 10.1523/JNEUROSCI.17-07-02637.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Harrington ME, Hoque S, Hall A, Golombek D, Biello S. Pituitary adenylate cyclase activating peptide phase shifts circadian rhythms in a manner similar to light. J Neurosci. 1999;19:6637–6642. doi: 10.1523/JNEUROSCI.19-15-06637.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hastings MH. Central clocking. Trends Neurosci. 1997;20:459–464. doi: 10.1016/s0166-2236(97)01087-4. [DOI] [PubMed] [Google Scholar]
- 18.Jin X, Shearman LP, Weaver DR, Zylka MJ, de Vries GJ, Reppert SM. A molecular mechanism regulating rhythmic output from the suprachiasmatic circadian clock. Cell. 1999;96:57–68. doi: 10.1016/s0092-8674(00)80959-9. [DOI] [PubMed] [Google Scholar]
- 19.Kume K, Zylka MJ, Sriram S, Shearman LP, Weaver DR, Jin X, Maywood ES, Hastings MH, Reppert SM. mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell. 1999;98:193–205. doi: 10.1016/s0092-8674(00)81014-4. [DOI] [PubMed] [Google Scholar]
- 20.Leander P, Vrang N, Moller M. Neuronal projections from the mesencephalic raphe nuclear complex to the suprachiasmatic nucleus and the deep pineal gland of the golden hamster (Mesocricetus auratus). J Comp Neurol. 1998;14:73–93. [PubMed] [Google Scholar]
- 21.Lovenberg TW, Baron BM, de Lecea L, Miller JD, Prosser RA, Rea MA, Foye PE, Racke M, Slone AL, Siegel BW, Danielson PE, Sutcliffe JG, Erlander MG. A novel adenylyl cyclase-activating serotonin receptor (5-HT7) implicated in the regulation of mammalian circadian rhythms. Neuron. 1993;11:449–458. doi: 10.1016/0896-6273(93)90149-l. [DOI] [PubMed] [Google Scholar]
- 22.Maywood ES, Smith E, Hall SJ, Hastings MH. A thalamic contribution to arousal-induced, non-photic entrainment of the circadian clock of the Syrian hamster. Eur J Neurosci. 1997;9:1739–1747. doi: 10.1111/j.1460-9568.1997.tb01531.x. [DOI] [PubMed] [Google Scholar]
- 23.Maywood ES, Mrosovsky N, Field MD, Hastings MH. Rapid down-regulation of mammalian period genes during behavioral resetting of the circadian clock. Proc Natl Acad Sci USA. 1999;96:15211–15216. doi: 10.1073/pnas.96.26.15211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.McNulty S, Schurov IL, Sloper PJ, Hastings MH. Stimuli which entrain the circadian clock of the neonatal Syrian hamster in vivo regulate the phosphorylation of the transcription factor CREB in the suprachiasmatic nucleus in vitro. Eur J Neurosci. 1998;10:1063–1072. doi: 10.1046/j.1460-9568.1998.00114.x. [DOI] [PubMed] [Google Scholar]
- 25.Mead S, Ebling FJ, Maywood ES, Humby T, Herbert J, Hastings MH. A nonphotic stimulus causes instantaneous phase advances of the light-entrainable circadian oscillator of the Syrian hamster but does not induce the expression of c-fos in the suprachiasmatic nuclei. J Neurosci. 1992;12:2516–2522. doi: 10.1523/JNEUROSCI.12-07-02516.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Meyer-Bernstein EL, Morin LP. Differential serotonergic innervation of the suprachiasmatic nucleus and the intergeniculate leaflet and its role in circadian rhythm modulation. J Neurosci. 1996;16:2097–2111. doi: 10.1523/JNEUROSCI.16-06-02097.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Miller JD, Liu ZW, Tenner T. Activity of 8-OH-DPAT enantiomers at the 5-HT7 receptor in SCN and transfected CHO cells. Soc Neurosci Abstr. 1996;22:1142. [Google Scholar]
- 28.Mintz EM, Gillespie CF, Marvel CL, Huhman KL, Albers HE. Serotonergic regulation of circadian rhythms in syrian hamsters. Neuroscience. 1997;79:563–569. doi: 10.1016/s0306-4522(96)00696-3. [DOI] [PubMed] [Google Scholar]
- 29.Miyamoto Y, Sancar A. Circadian regulation of cryptochrome genes in the mouse. Brain Res Mol Brain Res. 1999;71:238–243. doi: 10.1016/s0169-328x(99)00192-8. [DOI] [PubMed] [Google Scholar]
- 30.Moore RY. The suprachiasmatic nucleus and the organization of a circadian system. Trends Neurosci. 1982;5:404–407. [Google Scholar]
- 31.Mrosovsky N. Phase response curves for social entrainment. J Comp Physiol [A] 1988;162:35–46. doi: 10.1007/BF01342701. [DOI] [PubMed] [Google Scholar]
- 32.Obrietan K, Impey S, Storm DR. Light and circadian rhythmicity regulate MAP kinase activation in the suprachiasmatic nuclei. Nat Neurosci. 1998;1:693–700. doi: 10.1038/3695. [DOI] [PubMed] [Google Scholar]
- 33.Okamura H, Miyake S, Sumi Y, Yamaguchi S, Yasui A, Muijtjens M, Hoeijmakers JH, van der Horst GT. Photic induction of mPer1 and mPer2 in cry-deficient mice lacking a biological clock. Science. 1999;286:2531–2534. doi: 10.1126/science.286.5449.2531. [DOI] [PubMed] [Google Scholar]
- 34.Prosser RA, Gillette MU. The mammalian circadian clock in the suprachiasmatic nuclei is reset in vitro by cAMP. J Neurosci. 1989;9:1073–1081. doi: 10.1523/JNEUROSCI.09-03-01073.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Prosser RA, Dean RR, Edgar DM, Heller HC, Miller JD. Serotonin and the mammalian circadian system: I. In vitro phase shifts by serotonergic agonists and antagonists. J Biol Rhythms. 1993;8:1–16. doi: 10.1177/074873049300800101. [DOI] [PubMed] [Google Scholar]
- 36.Prosser RA, Heller HC, Miller JD. Serotonergic phase advances of the mammalian circadian clock involve protein kinase A and K+ channel opening. Brain Res. 1994;644:67–73. doi: 10.1016/0006-8993(94)90348-4. [DOI] [PubMed] [Google Scholar]
- 37.Reebs SG, Mrosovsky N. Effects of induced wheel running on the circadian activity rhythm of Syrian hamsters: entrainment and phase response curve. J Biol Rhythms. 1989;4:39–48. doi: 10.1177/074873048900400103. [DOI] [PubMed] [Google Scholar]
- 38.Sakamoto K, Nagase T, Fukui H, Horikawa K, Okada T, Tanaka H, Sato K, Miyake Y, Ohara O, Kako K, Ishida N. Multitissue circadian expression of rat period homolog (rPer2) mRNA is governed by the mammalian circadian clock, the suprachiasmatic nucleus in the brain. J Biol Chem. 1998;273:27039–27042. doi: 10.1074/jbc.273.42.27039. [DOI] [PubMed] [Google Scholar]
- 39.Schuhler S, Saboureau M, Pitrosky B, Pevet P. In Syrian hamsters, 5-HT fibres within the suprachiasmatic nuclei are necessary for the expression of 8-OH-DPAT induced phase-advance of locomotor activity rhythm. Neurosci Lett. 1998;256:33–36. doi: 10.1016/s0304-3940(98)00749-6. [DOI] [PubMed] [Google Scholar]
- 40.Shearman LP, Zylka MJ, Weaver DR, Kolakowski LF, Jr, Reppert SM. Two period homologs: circadian expression and photic regulation in the suprachiasmatic nuclei. Neuron. 1997;19:1261–1269. doi: 10.1016/s0896-6273(00)80417-1. [DOI] [PubMed] [Google Scholar]
- 41.Shibata S, Tsuneyoshi A, Hamada T, Tominaga K, Watanabe S. Phase-resetting effect of 8-OH DPAT, a serotonin 1A receptor agonist, on the circadian rhythm of firing rate in the rat suprachiasmatic nuclei in vitro. Brain Res. 1992;582:353–356. doi: 10.1016/0006-8993(92)90156-4. [DOI] [PubMed] [Google Scholar]
- 42.Shibata S, Watanabe A, Hamada T, Ono M, Watanabe S. N-methyl-d-aspartate induces phase shifts in circadian rhythm of neuronal activity of rat SCN in vitro. Am J Physiol. 1994;267:R360–R364. doi: 10.1152/ajpregu.1994.267.2.R360. [DOI] [PubMed] [Google Scholar]
- 43.Shigeyoshi Y, Taguchi K, Yamamoto S, Takekida S, Yan L, Tei H, Moriya T, Shibata S, Loros JJ, Dunlap JC, Okamura H. Light-induced resetting of a mammalian circadian clock is associated with rapid induction of the mPer1 transcript. Cell. 1997;91:1043–1053. doi: 10.1016/s0092-8674(00)80494-8. [DOI] [PubMed] [Google Scholar]
- 44.Shirakawa T, Moore RY. Glutamate shifts the phase of the circadian neuronal firing rhythm in the rat suprachiasmatic nucleus in vitro. Neurosci Lett. 1994;178:47–50. doi: 10.1016/0304-3940(94)90286-0. [DOI] [PubMed] [Google Scholar]
- 45.Sun ZS, Albrecht U, Zhuchenko O, Bailey J, Eichele G, Lee CC. RIGUI, a putative mammalian ortholog of the Drosophila period gene. Cell. 1997;19:1003–1011. doi: 10.1016/s0092-8674(00)80366-9. [DOI] [PubMed] [Google Scholar]
- 46.Takumi T, Matsubara C, Shigeyoshi Y, Taguchi K, Yagita K, Maebayashi Y, Sakakida Y, Okumura K, Takashima N, Okamura H. A new mammalian period gene predominantly expressed in the suprachiasmatic nucleus. Genes Cells. 1998a;3:167–176. doi: 10.1046/j.1365-2443.1998.00178.x. [DOI] [PubMed] [Google Scholar]
- 47.Takumi T, Taguchi K, Miyake S, Sakakida Y, Takashima N, Matsubara C, Maebayashi Y, Okumura K, Takekida S, Yamamoto S, Yagita K, Yan L, Young MW, Okamura H. A light-independent oscillatory gene mPer3 in mouse SCN and OVLT. EMBO J. 1998b;17:4753–4759. doi: 10.1093/emboj/17.16.4753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Tei H, Okamura H, Shigeyoshi Y, Fukuhara C, Ozawa R, Hirose M, Sakaki Y. Circadian oscillation of a mammalian homologue of the Drosophila period gene. Nature. 1997;389:512–516. doi: 10.1038/39086. [DOI] [PubMed] [Google Scholar]
- 49.Thresher RJ, Vitaterna MH, Miyamoto Y, Kazantsev A, Hsu DS, Petit C, Selby CP, Dawut L, Smithies O, Takahashi JS, Sancar A. Role of mouse cryptochrome blue-light photoreceptor in circadian photoresponses. Science. 1998;282:1490–1494. doi: 10.1126/science.282.5393.1490. [DOI] [PubMed] [Google Scholar]
- 50.Tominaga K, Shibata S, Ueki S, Watanabe S. Effects of 5-HT1A receptor agonists on the circadian rhythm of wheel-running activity in hamsters. Eur J Pharmacol. 1992;214:79–84. doi: 10.1016/0014-2999(92)90099-p. [DOI] [PubMed] [Google Scholar]
- 51.Tsou AP, Kosaka A, Bach C, Zuppan P, Yee C, Tom L, Alvarez R, Ramsey S, Bonhaus DW, Stefanich E. Cloning and expression of a 5-hydroxytryptamine7 receptor positively coupled to adenylyl cyclase. J Neurochem. 1994;63:456–464. doi: 10.1046/j.1471-4159.1994.63020456.x. [DOI] [PubMed] [Google Scholar]
- 52.van der Horst GT, Muijtjens M, Kobayashi K, Takano R, Kanno S, Takao M, de Wit J, Verkerk A, Eker AP, van Leenen D, Buijs R, Bootsma D, Hoeijmakers JH, Yasui A. Mammalian Cry1 and Cry2 are essential for maintenance of circadian rhythms. Nature. 1999;398:627–630. doi: 10.1038/19323. [DOI] [PubMed] [Google Scholar]
- 53.Vitaterna MH, Selby CP, Todo T, Niwa H, Thompson C, Fruechte EM, Hitomi K, Thresher RJ, Ishikawa T, Miyazaki J, Takahashi JS, Sancar A. Differential regulation of mammalian period genes and circadian rhythmicity by cryptochromes 1 and 2. Proc Natl Acad Sci USA. 1999;96:12114–12119. doi: 10.1073/pnas.96.21.12114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Yan L, Takekida S, Shigeyoshi Y, Okamura H. Per1 and Per2 gene expression in the rat suprachiasmatic nucleus: circadian profile and the compartment-specific response to light. Neuroscience. 1999;94:141–150. doi: 10.1016/s0306-4522(99)00223-7. [DOI] [PubMed] [Google Scholar]
- 55.Ying SW, Rusak B. 5-HT7 receptors mediate serotonergic effects on light-sensitive suprachiasmatic nucleus neurons. Brain Res. 1997;755:246–254. doi: 10.1016/s0006-8993(97)00102-9. [DOI] [PubMed] [Google Scholar]
- 56.Zylka MJ, Shearman LP, Weaver DR, Reppert SM. Three period homologs in mammals: differential light responses in the suprachiasmatic circadian clock and oscillating transcripts outside of brain. Neuron. 1998;20:1103–1110. doi: 10.1016/s0896-6273(00)80492-4. [DOI] [PubMed] [Google Scholar]