

Abstract
Serotonin depletion during embryogenesis has been shown previously to retard the growth of the olfactory and accessory lobes of the lobster deutocerebrum (Benton et al., 1997). The present study was undertaken to determine whether morphological changes in the interneurons innervating these lobes contribute to this growth retardation. We examined the effects of in vivoserotonin depletion using 5,7-dihydroxytryptamine (5,7-DHT) on the morphology of the olfactory projection neurons, one of two major classes of interneurons that innervate both lobes. Intracellular dye fills of olfactory projection neurons in normal embryos showed that each neuron extensively innervates either the olfactory or accessory lobe before projecting to neuropil regions in the protocerebrum. In embryos injected with 5,7-DHT, however, the deutocerebral arbors of 13.5% of the olfactory projection neurons examined were either markedly reduced compared with normal neurons or absent. Affected neurons also exhibited a number of additional aberrant morphological features suggesting that these neurons represent cells that were affected during their initial morphogenesis. Olfactory projection neurons with aberrant morphologies were also encountered, although less frequently (7.5% of the neurons examined), in control (sham-injected) embryos indicating that the sham injections can affect the development of the brain. This observation provides insights into the nature of effects seen in control embryos in previous experiments (Benton et al., 1997). The results of the present study indicate that in vivo serotonin depletion inhibits the branching of olfactory projection neurons and suggest, therefore, that one of the functions of serotonin during normal development is to promote the ingrowth of these neurons into the deutocerebral neuropils.
Keywords: serotonin; development; lobster; Homarus americanus; olfaction; olfactory projection neuron; deutocerebrum; 5,7-dihydroxytryptamine
The developmental signals that control the growth and morphogenesis of individual neurons play crucial roles in shaping the nervous system. The early appearance of serotonin during the development of both vertebrates (Lauder and Bloom, 1974;Fujimiya et al., 1986) and invertebrates (Taghert and Goodman, 1984;Glover et al., 1987; Goldberg and Kater, 1989; Beltz et al., 1990) has led to the hypothesis that this neurotransmitter plays an important role in the regulation of neural development (for review, seeWhitaker-Azmitia et al., 1996; Levitt et al., 1997). This hypothesis has been supported by studies demonstrating that perturbations of serotonin levels can effect the development of neuropil areas that receive serotonergic innervation during development (Rhoades et al., 1993; Bennett-Clarke et al., 1994; Cases et al., 1996; Benton et al., 1997). Although studies at the cellular level have shown that developmental events such as neuronal proliferation (Lauder et al., 1981), synaptogenesis (Haydon et al., 1984; Chubakov et al., 1986), and neurite outgrowth (Mercer et al., 1996; Goldberg, 1998; Lieske et al., 1999; Lotto et al., 1999) are susceptible to modulation by serotonin levels, relatively few studies (for instance, Goldberg and Kater, 1989) have examined the effects of serotonin on the development of individual neurons by the use of in vivo models.
Dense serotonergic innervation is a characteristic feature of neuropil regions involved in the processing of primary olfactory input (Halasz and Shepherd, 1983; Kent et al., 1987; Sudlow et al., 1998; Beltz, 1999). Studies in embryonic lobsters have shown that the growth of olfactory neuropil areas in the midbrain, or deutocerebrum, is retarded by pharmacological depletion of serotonin by the use of 5,7-dihydroxytryptamine (5,7-DHT) (Benton et al., 1997) (J. L. Benton and B. S. Beltz, unpublished observations). These effects were found to be specific to those neuropil areas that normally receive serotonergic innervation (Benton et al., 1997).
In the lobster, primary sensory neurons from the olfactory organs (first antennae) terminate in the deutocerebrum within the highly structured glomerular neuropil of the paired olfactory lobes. Olfactory lobe glomeruli are sites of synaptic contact between the sensory neurons, olfactory projection neurons, and several classes of local interneurons. The latter two classes of neuron also innervate two adjacent neuropil regions, the accessory lobe and the olfactory-globular tract neuropil (OGTN), that are also involved in the processing of olfactory information (Fig.1) (Mellon and Alones, 1994; Sandeman and Sandeman, 1994; Wachowiak et al., 1996). All three of these neuropil regions receive dense serotonergic innervation and are affected by serotonin depletion during their embryonic development (Benton et al., 1997) (Benton and Beltz, unpublished observations). In contrast, the growth of the antenna II neuropil (Fig. 1), which is not innervated by serotonergic neurons during embryogenesis, is not affected by serotonin depletion (Benton et al., 1997).
Fig. 1.

Morphology of the lobster deutocerebrum during late embryonic development. A, Schematic diagram outlining the arrangement of the deutocerebral neuropils that have been examined in previous serotonin depletion experiments (other neuropil areas in the deutocerebrum not shown). In decapod crustaceans, the OL, AL, andOGTN are innervated by branches of the olfactory projection neurons whose cell bodies are located lateral to the neuropils in a dense cluster, known as cluster 10. The axons of the projection neurons form a large tract, theOGT, that projects bilaterally to the lateral protocerebrum in the eyestalks. The black outlined square indicates the region of the brain detailed inB. Anterior is at the top.B, Confocal microscope image of the deutocerebrum of an embryonic lobster brain (E75%) stained with an antibody raised againstDrosophila synapsin (SYNORF1; E. Buchner, Universität Würzburg, Würzburg, Germany). Tissues were processed by the use of standard procedures (see Harzsch et al., 1999). The primary neurites of the projection neurons can be seen entering the OL and AL fromcluster 10 (diverging arrows). On leaving the lobes, the axons of the neurons form the OGT(converging arrows) that passes through theOGTN (asterisk) en route to the lateral protocerebrum. AL, Accessory lobe; AnII, antenna II neuropil; cluster 10, cell body cluster of the olfactory projection neurons; OGT, olfactory-globular tract;OGTN, olfactory-globular tract neuropil;OL, olfactory lobe [terminology from Sandeman et al. (1992)]. Scale bars: A, 100 μm; B, 20 μm.
The predominant serotonergic input to the deutocerebral neuropils of the lobster is provided by a bilateral pair of dorsal giant neurons (DGNs). Morphological analyses of the DGNs in 5,7-DHT-treated embryos have shown that neither the morphology nor the size of these neurons is significantly altered by serotonin depletion (Benton and Beltz, unpublished observations). This result suggests that the major effects of serotonin depletion are not on the serotonergic cells but rather on the olfactory interneurons that innervate the deutocerebral neuropils. In the present study we have examined the effects of serotonin depletion in vivo on the development and morphology of the olfactory projection neurons, one of the two major groups of interneurons innervating the deutocerebral olfactory neuropils.
MATERIALS AND METHODS
Embryonic lobsters, Homarus americanus, were obtained from the New England Aquarium (Boston, MA) and maintained at 14°C in aquaria with circulating artificial seawater and a light/dark cycle of 12:12 hr. The developmental stage of the embryos was determined by the use of the percentage staging system of Helluy and Beltz (1991) in which egg extrusion is defined as 0% and hatching as 100%. Embryos at 55% of embryonic development (E55%) were assigned to one of three groups: experimental, control, or normal. Experimental animals were injected with 0.13–0.26 μl of 10−2m 5,7-DHT (Sigma, St. Louis, MO) in a vehicle solution (saline, 1% ascorbate, and 0.05% visual marker) by the use of the procedure described previously by Benton et al. (1997). Control animals were injected only with vehicle solution, whereas normal animals were handled but not injected. Embryos were injected or handled once a week for 3 weeks and then allowed to develop over a period of several weeks until they reached ∼E80%. HPLC studies in previous serotonin depletion experiments have shown that several days after a series of three 5,7-DHT injections beginning at E55%, serotonin levels in H. americanus embryos are reduced by ∼90% and that this depletion is long lasting (Benton et al., 1997).
During normal embryonic development, H. americanusexperiences a period of developmental arrest beginning at ∼E80% that lasts ∼2 weeks at 18°C (Helluy and Beltz, 1991). After embryos entered this period of developmental arrest, morphological analyses of the olfactory projection neurons were performed to assess the effects of serotonin depletion on the normal development of these neurons. The brain and ventral nerve cord of the embryo were dissected free and placed in a well of cold lobster saline on a poly-l-lysine (0.01% in dH2O; Sigma)-coated slide. Preparations were then viewed with a fixed-stage Nikon compound microscope equipped with Nomarski optics. The morphology of the projections neurons within the deutocerebrum was examined by intracellular staining of the cells with Lucifer yellow CH (Sigma). Neurons were penetrated primarily in the cell body and stained by iontophoretic injection of Lucifer yellow using hyperpolarizing current pulses of up to 6 nA (500 msec in duration; 1 Hz in frequency) for 4–7 min. The cell bodies of the olfactory projection neurons are situated lateral to the deutocerebral neuropils in a densely packed cluster, known as cluster 10 (Fig. 1). Numerous large neuroblasts arise in this region during early embryogenesis and continue to generate new projection neurons throughout the period of embryonic development examined in the present study (Harzsch et al., 1999). These neuroblasts are concentrated in the posterior half of the cluster where they form a large proliferative domain (Harzsch et al., 1999). To preferentially target projection neurons that were born during E55–E80% we only penetrated neurons whose cell bodies were located within the region of the proliferative domain.
After the injection of Lucifer yellow, preparations were fixed in 4% paraformaldehyde for 4 hr, dehydrated in an ethanol series (8 min in each of 50, 70, 80, 90, 95, and 100%), and cleared in methyl salicylate. Subsequently, the brains were mounted in DPX (Fluka, Buchs, Switzerland) and viewed by the use of a Leica scanning confocal microscope. To exclude both fills of neurons in the initial stages of axonogenesis and neurons that were not completely filled with dye, only those preparations in which the axons of the projection neurons were stained in the olfactory-globular tract (OGT; Fig. 1) well beyond the deutocerebral neuropil areas were included in the analyses. The results of the present study are based on the analysis of a total of 113 olfactory projection neurons (experimental, n = 37 neurons in 9 embryos; control, n = 53 neurons in 13 embryos; normal, n = 23 neurons in 8 embryos).
RESULTS
Olfactory projection neuron morphology
Intracellular dye fills of individual projection neurons in normal embryos show that the soma of each neuron gives rise to a primary neurite that projects ipsilaterally to either the olfactory or accessory lobe (Fig. 2). Typically, the primary neurite of each neuron courses into and out of the initial portion of a lobe with its dendritic tree arising from the side of the neurite facing the center of the lobe. Each of the stained projection neurons that innervated the olfactory lobe (OL) was multiglomerular and branched widely within the lobe (Fig. 2A). Projection neurons innervating the accessory lobe (AL) could be identified as belonging to one of two groups on the basis of their arborizations within the lobe. The primary neurites of the first group (type I) passed only into the initial portion of the AL before arborizing around the outside of the lobe (Fig. 2C). AL projection neurons of the second group (type II) were encountered less frequently than the first (Fig. 3) and possessed primary neurites that passed into the center of the lobe before branching outward (Fig. 2D). Projection neurons innervating both lobes, such as those that have been described in the crayfish (Mellon et al., 1992a; Mellon and Alones, 1993), were not observed in the present study in the lobster.
Fig. 2.
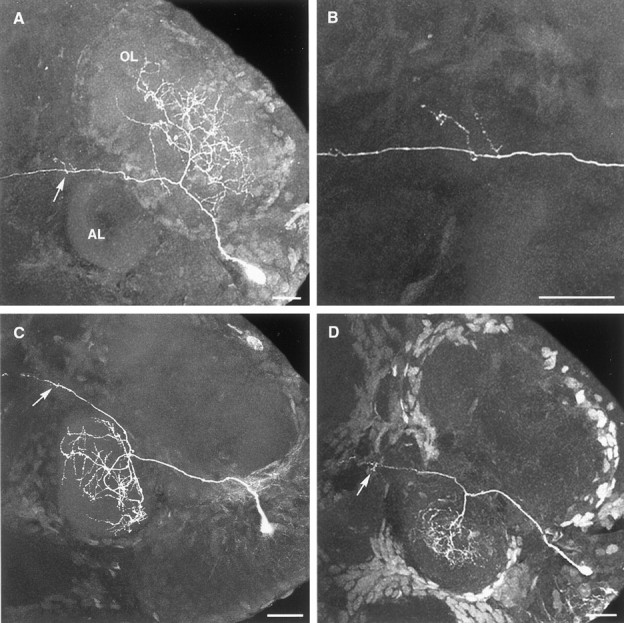
Stacked confocal images illustrating the morphologies of the three olfactory projection neuron types observed in the deutocerebrum of the embryonic lobster. The neurons were filled intracellularly with Lucifer yellow, and the arrowsindicate the branches of the neurons within the OGTN. Anterior is at the top. A, An olfactory lobe projection neuron. B, Detail of the neuron shown inA, showing the dendritic branches of the neuron within the OGTN. C, A type I accessory lobe projection neuron.D, A type II accessory lobe projection neuron.AL, Accessory lobe; OL, olfactory lobe. Scale bars: A–D, 20 μm.
Fig. 3.

Percentages with which the three olfactory projection neuron types observed in normal embryos (OL,AL-Type I, and AL-Type II) and the projection neurons with aberrant morphologies (Aberrant) were encountered in the three groups of embryos.AL, Accessory lobe; OL, olfactory lobe.
On leaving the olfactory or accessory lobe, the axons of the projection neurons form the OGT and pass through a small region of neuropil, known as the OGTN, where some of the neurons give off small axon collaterals (Figs. 1, 2). Both OL and AL projection neurons were observed to branch within the OGTN (Fig. 2). The amount of branching within the OGTN varied between neurons but, when present, rarely consisted of more than one or two fine branches. On passing through the OGTN, the OGT projects bilaterally to the eyestalks where it innervates the lateral protocerebrum (Fig. 1) (Helluy et al., 1995).
Effects of serotonin depletion on projection neuron morphology
Each of the projection neurons stained in normal embryos (n = 23) extensively innervated either the olfactory or the accessory lobe. The percentages with which the three projection neuron types (OL, AL-type I, and AL-type II) were encountered in normal embryos are presented in Figure 3. These three neuronal types were also encountered with similar frequencies in experimental and control embryos (Fig. 3). In the brains of experimental and control embryos, however, projection neurons were also encountered in which dendritic branching within the lobes was either markedly reduced compared with normal neurons or completely absent (Fig.4). The primary neurites of a number of these neurons neither coursed into nor branched within either lobe (e.g., Fig. 4B) but rather passed directly into the OGT, and in several preparations the axons of the neurons could be traced into the lateral protocerebrum (data not shown). Other neurons with markedly reduced branching also differed from normal projection neurons in possessing fine dendritic branches along both sides of the primary neurite rather than from the side of the neurite facing the lobe (e.g., Fig. 4D).
Fig. 4.
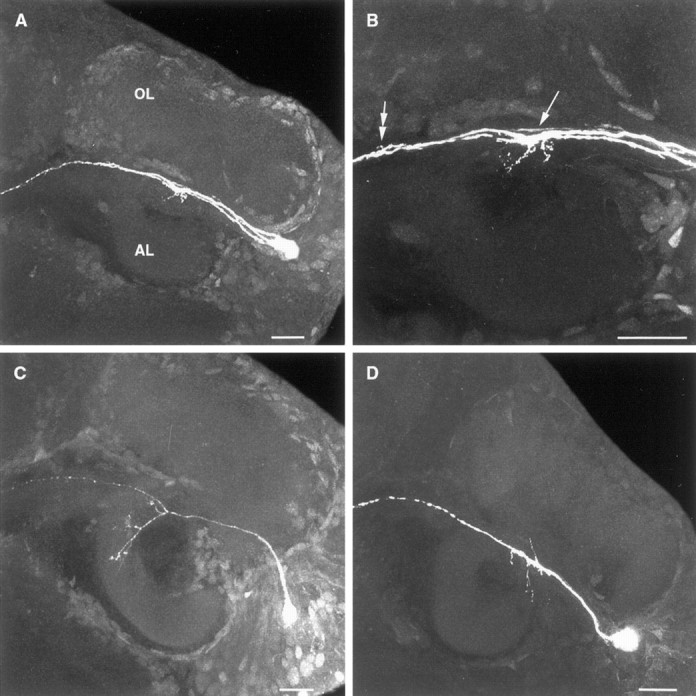
Stacked confocal images illustrating the morphologies of olfactory projection neurons affected by the experimental treatments. Neurons were stained with Lucifer yellow. Anterior is at the top. A, Three neurons stained sequentially in an experimental embryo. B, Detail of the preparation shown in A. The primary neurite of one of these cells (arrow) courses directly between the OL and AL and does not possess any dendritic branching. The remaining two neurons give rise to fine dendritic branches in the region between the lobes, whereas the posterior-most neuron also branches in the OGTN (double arrow). C, Projection neuron stained in an experimental embryo exhibiting markedly reduced branching within theAL. D, Projection neuron within the deutocerebrum of a control embryo. Fine dendritic branches arise from both sides of the primary neurite and project toward both the olfactory and accessory lobes. AL, Accessory lobe;OL, olfactory lobe. Scale bars: A–D, 20 μm.
Olfactory projection neurons possessing such aberrant morphological traits constituted 13.5% of the neurons stained in experimental embryos (Fig. 3; n = 5 of 37; cells observed in 2 of 9 experimental embryos). A two-sample Z test of proportions showed that aberrant neurons occurred significantly more frequently in experimental embryos than in normal embryos (p = 0.03). Projection neurons with altered morphologies were also encountered, although less frequently (Fig. 3), in control embryos in which they constituted 7.5% of the neurons stained (n= 4 of 53; cells observed in 4 of 13 control embryos). A two-sampleZ test of proportions showed that aberrant neurons did not occur significantly more frequently in control embryos than in normal embryos (p = 0.09). Although aberrant neurons were encountered more frequently in experimental than in control embryos, this difference was not statistically significant with our sample size (two-sample Z test; p = 0.17). In both experimental and control embryos, projection neurons with aberrant morphologies occurred within the same brains as projection neurons exhibiting normal morphologies, indicating that only a subset of the projection neurons within each brain were affected by the treatments. These observations also indicate that the effects of the experimental treatments were not restricted to particular animals.
DISCUSSION
In the present study, we examined whether the in vivodevelopment of olfactory projection neurons in the lobster deutocerebrum is affected by serotonin depletion using 5,7-DHT. Previous experiments in embryonic lobsters have shown that the growth of the three deutocerebral neuropils innervated by these neurons is significantly reduced by serotonin depletion (Benton et al., 1997) (Benton and Beltz, unpublished observations). Intracellular dye injections of projection neurons in normal embryos show that each neuron extensively innervates either the olfactory or accessory lobe, a result consistent with those of previous studies in adult decapod crustaceans (Tsvileneva and Titova, 1985; Mellon et al., 1992a; Mellon and Alones, 1993; Wachowiak and Ache, 1994; Wachowiak et al., 1996). The presence of projection neurons in experimental embryos in which branching within the deutocerebral neuropils is markedly reduced or absent demonstrates that the development of at least a portion of these neurons is affected by serotonin depletion. The results of this study indicate, therefore, that effects of serotonin depletion in vivo on the development of olfactory projection neurons contribute to the reduction in deutocerebral neuropil volumes that occurs after treatment with 5,7-DHT.
Neuroblasts within the cell body cluster (cluster 10) of the olfactory projection neurons commence their proliferative activity during early embryonic development (before E25%) and continue to generate new neurons until late embryonic development (Harzsch et al., 1999). During the period of serotonin depletion in the present study (E55–E80%), therefore, the deutocerebrum of the lobster is comprised of both developing neurons and projection neurons born at an earlier stage of embryogenesis whose dendritic trees were established within the lobes before the injection of 5,7-DHT. In the present study, we examined the morphology of olfactory projection neurons whose cell bodies were located within the posterior half of cluster 10. This region of the cluster was selected because the neuroblasts and, therefore, newly born and developing neurons are concentrated within this region (Harzsch et al., 1999). The aberrant morphological features of the projection neurons affected by the experimental treatments, such as the presence of branching from both sides of the primary neurites and primary neurites that do not course into the lobes, would be unlikely to arise in established neurons whose dendritic trees have atrophied. The affected neurons are, therefore, likely to be neurons that either were born or commenced axonogenesis during the period of serotonin depletion or a particular subset thereof. Consequently, the projection neurons with normal morphologies present in the brains of experimental animals may represent neurons whose development is not sensitive to modulation by serotonin levels and/or neurons whose dendritic trees were established within the lobes before the period of serotonin depletion. These results suggest, therefore, that one of the functions of serotonin during normal development in vivo is to promote the ingrowth of the neurites of at least a subset of developing olfactory projection neurons into the deutocerebral neuropils.
In contrast to the relatively small numbers of olfactory projection neurons in insects, which number in the hundreds (Homberg et al., 1988;Malun et al., 1988), the deutocerebral lobes of adult decapod crustaceans are innervated by tens of thousands of projection neurons (Mellon et al., 1992b; Schmidt, 1997; Sandeman et al., 1998). In lobster embryos, cluster 10 contains the cell bodies of ∼2500 projection neurons by midembryonic development, and in normal embryos this number increases to ∼4500 by E80% (J. M. Sullivan, unpublished observations). Neuronal proliferation in a range of species, however, has been shown to be susceptible to modulation by serotonin levels (Lauder et al., 1981; Brezun and Daszuta, 1999;Gould, 1999). Studies in the lobster have shown that neuronal proliferation rates in cluster 10 of embryos treated with 5,7-DHT are significantly reduced compared with the rates in both normal and control embryos (Benton and Beltz, unpublished observations). In the present study, therefore, significantly fewer neurons were born in experimental embryos during the period of serotonin depletion than in control and normal embryos during the corresponding periods of development. Consequently, the frequency with which aberrant neurons were encountered in embryos treated with 5,7-DHT is likely to represent an underestimate of the proportion of projection neurons affected by the experimental compared with control and normal treatments.
The presence of neurons with aberrant morphologies in both experimental and control embryos indicates that the sham injections involved in the control treatment can also affect the development of the deutocerebrum. This finding was also noted in the initial serotonin depletion experiments, in which deutocerebral neuropil volumes in control embryos were found to be significantly smaller than those of normal embryos (Benton et al., 1997). The reductions in neuropil volume seen in embryos treated with 5,7-DHT, however, were far more dramatic than were those seen in control embryos and were specific to those neuropils that normally receive serotonergic innervation during embryonic development (Benton et al., 1997). The sham injections therefore appear to effect the development of the deutocerebrum in a manner that is parallel to, but of lesser magnitude than, treatment with 5,7-DHT. One possible explanation for the results observed in control embryos is that stress resulting from the sham injections may alter serotonin levels within the animals. It has been well documented in mammals that extracellular serotonin levels are modulated by stressful stimuli (for review, seeChaouloff et al., 1999). In decapod crustaceans serotonin has been shown to mediate the release of stress-related hormones (Konishi and Kravitz, 1978; Mattson and Spaziani, 1985, 1986). It has yet to be determined, however, how serotonin levels are modulated during stress responses. An alternative explanation is that one or more of the stress-related hormones may directly influence the morphogenesis of the projection neurons via mechanisms parallel to those that mediate the effects of serotonin depletion.
Numerous studies have shown direct effects of serotonin on neurite outgrowth in vitro. The specific effects of the amine, however, vary according to the cell type examined. In the pond snailHelisoma trivolvis, for example, the principal effect of serotonin on developing neurons is to inhibit neurite outgrowth (for review, see Goldberg, 1998), an effect that has also been demonstrated in in vivo studies using 5,7-DHT (Goldberg and Kater, 1989). In contrast, the addition of serotonin to the cell medium surrounding cultured rodent thalamic neurons promotes neurite outgrowth and branching (Lieske et al., 1999; Lotto et al., 1999), an effect consistent with that observed in the present study. Similarly, Mercer et al. (1996) showed that in the sphinx moth Manduca sextaserotonin enhances the in vitro growth of antennal (olfactory) lobe projection neurons, structural and functional analogs of the projection neurons of decapod crustaceans (Ache, 1991;Strausfeld and Hildebrand, 1999). As in the lobster, in which the axons of projection neurons affected by serotonin depletion still project successfully into the OGT (Fig. 4), the in vitro effects of serotonin on the projection neurons of M. sexta were found to be restricted to the regions of the neurons that would normally innervate the glomeruli of the antennal lobe in situ (Mercer et al., 1996).
Although serotonin has been shown to have a variety of regulatory effects on neuronal outgrowth in vitro, few studies have examined how serotonin regulates the development of individual neuronsin vivo. Goldberg and Kater (1989) examined the effects ofin vivo serotonin depletion during embryogenesis inHelisoma trivolvis on the morphology of the identified neuron B19, a neuron whose development in vitro had been shown previously to be sensitive to perturbations in serotonin levels (McCobb et al., 1988). Neuron B19 was found to possess aberrant neuritic structures in 15% of the animals in which serotonin was depleted (Goldberg and Kater, 1989). Whereas Goldberg and Kater (1989)examined the effects of serotonin depletion on the morphology of a single identified neuron, the present study examined the morphologies of individual projection neurons sampled from a heterogeneous population of cells. The finding that 13.5% of the olfactory projection neurons stained in experimental embryos possessed aberrant morphologies suggests, therefore, that serotonin may exert a stronger regulatory effect on the development of the projection neurons than that observed on B19. The effects of serotonin on the development of individual projection neurons may perhaps be even more extensive if, as was observed in the in vitro study of projection neurons inM. sexta (Mercer et al., 1996), only a portion of developing neurons are sensitive to serotonin levels.
Although the present study focused on the embryonic development of olfactory projection neurons, neurogenic activity in the deutocerebrum of the lobster is not restricted to embryogenesis. Neurogenesis within cluster 10 ceases during late embryonic development (Harzsch et al., 1999) but resumes after hatching and thereafter continues throughout the lifetime of the animal (Harzsch et al., 1999;Schmidt and Harzsch, 1999). New projection neurons are therefore continually being born throughout the postembryonic life of the lobster. These neurons must then grow into the deutocerebral neuropils and intercalate into the existing neuronal circuits. The dense serotonergic innervation of the deutocerebral neuropils that is present during embryonic development also remains stable throughout the postembryonic development of the lobster (Beltz et al., 1990). Serotonin may therefore continue to play a role in regulating the morphogenesis of projection neurons throughout the lifetime of the animal.
Footnotes
This study was supported by National Science Foundation Grant IBN 9709514. We thank Dr. S. Kohler for invaluable assistance with confocal microscopy and Dr. E. Buchner for kindly providing the synapsin antibody. We are indebted to J. Goldstein from the Lobster Rearing and Research Facility at the New England Aquarium (Boston, MA) and to P. Carey and V. Quinan from the Wellesley College Animal Care Facility for maintenance of experimental animals.
Correspondence should be addressed to Dr. Jeremy M. Sullivan at the above address. E-mail: jsulliva@wellesley.edu.
REFERENCES
- 1.Ache BW. Phylogeny of smell and taste. In: Getchell TV, Doty RL, Bartoshuk LM, Snow JB, editors. Smell and taste in health and disease. Raven; New York: 1991. pp. 3–18. [Google Scholar]
- 2.Beltz B, Pontes M, Helluy S, Kravitz E. Patterns of appearance of serotonin and proctolin immunoreactivities in the nervous system of the lobster. J Neurobiol. 1990;21:521–542. doi: 10.1002/neu.480210402. [DOI] [PubMed] [Google Scholar]
- 3.Beltz BS. Distribution and functional anatomy of amine-containing neurons in decapod crustaceans. Microsc Res Tech. 1999;44:105–120. doi: 10.1002/(SICI)1097-0029(19990115/01)44:2/3<105::AID-JEMT5>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 4.Bennett-Clarke CA, Leslie MJ, Lane RD, Rhoades RW. Effect of serotonin depletion on vibrissa-related patterns of thalamic afferents in the rat's somatosensory cortex. J Neurosci. 1994;14:7594–7607. doi: 10.1523/JNEUROSCI.14-12-07594.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Benton J, Huber R, Ruchhoeft M, Helluy S, Beltz B. Serotonin depletion by 5,7-dihydroxytryptamine alters deutocerebral development in the lobster, Homarus americanus. J Neurobiol. 1997;33:357–373. doi: 10.1002/(sici)1097-4695(199710)33:4<357::aid-neu2>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- 6.Brezun JM, Daszuta A. Depletion in serotonin decreases neurogenesis in the dentate gyrus and the subventricular zone of adult rats. Neuroscience. 1999;89:999–1002. doi: 10.1016/s0306-4522(98)00693-9. [DOI] [PubMed] [Google Scholar]
- 7.Cases O, Vitalis T, Seif I, De Maeyer E, Sotelo C, Gaspar P. Lack of barrels in the somatosensory cortex of monoamine oxidase A deficient mice: role of serotonin excess during the critical period. Neuron. 1996;16:297–307. doi: 10.1016/s0896-6273(00)80048-3. [DOI] [PubMed] [Google Scholar]
- 8.Chaouloff F, Berton O, Mormède P. Serotonin and stress. Neuropsychopharmacology. 1999;21:28–32. doi: 10.1016/S0893-133X(99)00008-1. [DOI] [PubMed] [Google Scholar]
- 9.Chubakov AR, Gromova EA, Konovalov GV, Sarikisova EF, Chumasov EI. The effects of serotonin on the morpho-functional development of rat cerebral neocortex in tissue culture. Brain Res. 1986;369:285–297. doi: 10.1016/0006-8993(86)90537-8. [DOI] [PubMed] [Google Scholar]
- 10.Fujimiya M, Hosoda S, Kitahama K, Kimura H, Maeda T. Early development of serotonin neurons in the rat brain as studied by immunocytochemistry combined with tryptophan administration. Brain Dev. 1986;8:335–342. doi: 10.1016/s0387-7604(86)80053-5. [DOI] [PubMed] [Google Scholar]
- 11.Glover JC, Stuart DK, Cline HT, McCaman RE, Magill C, Stent GS. Development of neurotransmitter metabolism in leech embryos. J Neurosci. 1987;7:581–594. doi: 10.1523/JNEUROSCI.07-02-00581.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Goldberg JI. Serotonin regulation of neurite outgrowth in identified neurons from mature and embryonic Helisoma trivolvis. Perspect Dev Neurobiol. 1998;5:373–387. [PubMed] [Google Scholar]
- 13.Goldberg JI, Kater SB. Expression and function of the neurotransmitter serotonin during development of the Heliosoma nervous system. Dev Biol. 1989;131:483–495. doi: 10.1016/s0012-1606(89)80019-3. [DOI] [PubMed] [Google Scholar]
- 14.Gould E. Serotonin and hippocampal neurogenesis. Neuropsychopharmacology. 1999;21:46–51. doi: 10.1016/S0893-133X(99)00045-7. [DOI] [PubMed] [Google Scholar]
- 15.Halasz N, Shepherd GM. Neurochemistry of the vertebrate olfactory bulb. J Neurosci. 1983;3:579–619. doi: 10.1016/0306-4522(83)90206-3. [DOI] [PubMed] [Google Scholar]
- 16.Harzsch S, Miller J, Benton J, Beltz B. From embryo to adult: persistent neurogenesis and apoptotic cell death shape the lobster deutocerebrum. J Neurosci. 1999;19:3472–3485. doi: 10.1523/JNEUROSCI.19-09-03472.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Haydon PG, McCobb DP, Kater SB. Serotonin selectively inhibits growth cone motility and synaptogenesis of specific identified neurons. Science. 1984;226:561–564. doi: 10.1126/science.6093252. [DOI] [PubMed] [Google Scholar]
- 18.Helluy SM, Beltz BS. Embryonic development of the American lobster: quantitative staging and characterization of the embryonic molt cycle. Biol Bull. 1991;180:355–371. doi: 10.2307/1542337. [DOI] [PubMed] [Google Scholar]
- 19.Helluy SM, Ruchhoeft ML, Beltz BS. Development of the olfactory and accessory lobes in the American lobster: an allometric analysis and its implications for the deutocerebral structure of decapods. J Comp Neurol. 1995;357:433–445. doi: 10.1002/cne.903570308. [DOI] [PubMed] [Google Scholar]
- 20.Homberg U, Montague RA, Hildebrand JG. Anatomy of antenno-cerebral pathways in the brain of the sphinx moth Manduca sexta. Cell Tissue Res. 1988;254:255–281. doi: 10.1007/BF00225800. [DOI] [PubMed] [Google Scholar]
- 21.Kent KS, Hoskins SG, Hildebrand JG. A novel serotonin-immunoreactive neuron in the antennal lobe of the sphinx moth Manduca sexta persists throughout postembryonic life. J Neurobiol. 1987;18:451–465. doi: 10.1002/neu.480180506. [DOI] [PubMed] [Google Scholar]
- 22.Konishi S, Kravitz EA. The physiological properties of amine-containing neurons in the lobster nervous system. J Physiol (Lond) 1978;279:215–229. doi: 10.1113/jphysiol.1978.sp012341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lauder JM, Bloom FE. Ontogeny of monoamine neurons in the locus coeruleus, raphe nuclei and substantia nigra of the rat. I. Cell differentiation. J Comp Neurol. 1974;155:469–482. doi: 10.1002/cne.901550407. [DOI] [PubMed] [Google Scholar]
- 24.Lauder JM, Wallace KA, Krebs H. Roles of serotonin in neuroembryogenesis. Adv Exp Med Biol. 1981;133:477–506. doi: 10.1007/978-1-4684-3860-4_28. [DOI] [PubMed] [Google Scholar]
- 25.Levitt P, Harvey JA, Friedman E, Simansky K, Murphy EH. New evidence for neurotransmitter influences on brain development. Trends Neurosci. 1997;20:269–274. doi: 10.1016/s0166-2236(96)01028-4. [DOI] [PubMed] [Google Scholar]
- 26.Lieske V, Bennett-Clarke CA, Rhoades RW. Effects of serotonin on neurite outgrowth from thalamic neurons in vitro. Neuroscience. 1999;90:967–974. doi: 10.1016/s0306-4522(98)00501-6. [DOI] [PubMed] [Google Scholar]
- 27.Lotto B, Upton L, Price DJ, Gaspar P. Serotonin receptor activation enhances neurite outgrowth of thalamic neurones in rodents. Neurosci Lett. 1999;269:87–90. doi: 10.1016/s0304-3940(99)00422-x. [DOI] [PubMed] [Google Scholar]
- 28.Malun D, Waldow U, Kraus D, Boeckh J. Connections between the deutocerebrum and the protocerebrum, and neuroanatomy of several classes of deutocerebral projection neurons in the brain of male Periplaneta americana. J Comp Neurol. 1988;329:143–162. doi: 10.1002/cne.903290202. [DOI] [PubMed] [Google Scholar]
- 29.Mattson MP, Spaziani E. 5-Hydroxytryptamine mediates release of molt-inhibiting hormone activity from isolated crab eyestalk ganglia. Biol Bull. 1985;171:264–273. [Google Scholar]
- 30.Mattson MP, Spaziani E. Regulation of the stress-responsive X-organ–Y-organ axis by 5-hydroxytryptamine in the crab, Cancer antennarius. Gen Comp Endocrinol. 1986;62:419–427. doi: 10.1016/0016-6480(86)90052-3. [DOI] [PubMed] [Google Scholar]
- 31.McCobb DP, Haydon PG, Kater SB. Dopamine and serotonin inhibition of neurite elongation of different identified neurons. J Neurosci Res. 1988;19:19–26. doi: 10.1002/jnr.490190104. [DOI] [PubMed] [Google Scholar]
- 32.Mellon DeF, Alones V. Cellular organization and growth-related plasticity of the crayfish olfactory midbrain. Microsc Res Tech. 1993;24:231–259. doi: 10.1002/jemt.1070240304. [DOI] [PubMed] [Google Scholar]
- 33.Mellon DeF, Alones V. Identification of three classes of multiglomerular, broad-spectrum neurons in the crayfish olfactory midbrain by correlated patterns of electrical activity and dendritic arborization. J Comp Physiol [A] 1994;177:55–71. [Google Scholar]
- 34.Mellon DeF, Alones V, Lawrence MD. Anatomy and fine structure of neurons in the deutocerebral projection pathway of the crayfish olfactory system. J Comp Neurol. 1992a;321:93–111. doi: 10.1002/cne.903210109. [DOI] [PubMed] [Google Scholar]
- 35.Mellon DeF, Sandeman DC, Sandeman RE. Characterization of oscillatory olfactory interneurones in the protocerebrum of the crayfish. J Exp Biol. 1992b;167:15–38. [Google Scholar]
- 36.Mercer AR, Kirschof BS, Hildebrand JG. Enhancement by serotonin of the growth in vitro of antennal lobe neurons of the sphinx moth Manduca sexta. J Neurobiol. 1996;29:49–64. doi: 10.1002/(SICI)1097-4695(199601)29:1<49::AID-NEU4>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 37.Rhoades RW, Bennett-Clarke CA, Lane RD, Leslie MJ, Mooney RD. Increased serotonergic innervation of the hamster's superior colliculus alters retinotectal projections. J Comp Neurol. 1993;334:397–409. doi: 10.1002/cne.903340306. [DOI] [PubMed] [Google Scholar]
- 38.Sandeman DC, Sandeman RE. Electrical responses and synaptic connections of giant-immunoreactive neurons in the crayfish olfactory and accessory lobes. J Comp Neurol. 1994;341:130–144. doi: 10.1002/cne.903410111. [DOI] [PubMed] [Google Scholar]
- 39.Sandeman DC, Sandeman R, Derby C, Schmidt M. Morphology of the brain of crayfish, crabs, and spiny lobsters: a common nomenclature for homologous structures. Biol Bull. 1992;183:304–326. doi: 10.2307/1542217. [DOI] [PubMed] [Google Scholar]
- 40.Sandeman R, Clarke D, Sandeman D, Manly M. Growth-related and antennular amputation-induced changes in the olfactory centers of crayfish brain. J Neurosci. 1998;18:6195–6206. doi: 10.1523/JNEUROSCI.18-16-06195.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schmidt M. Continuous neurogenesis in the olfactory brain of adult shore crabs, Carcinus maenus. Brain Res. 1997;762:131–143. doi: 10.1016/s0006-8993(97)00376-4. [DOI] [PubMed] [Google Scholar]
- 42.Schmidt M, Harzsch S. Comparative analysis of neurogenesis in the central olfactory pathway of adult decapod crustaceans by in vivo BrdU-labeling. Biol Bull. 1999;196:127–136. doi: 10.2307/1542558. [DOI] [PubMed] [Google Scholar]
- 43.Strausfeld NJ, Hildebrand JG. Olfactory systems: common design, uncommon origin? Curr Opin Neurobiol. 1999;9:634–639. doi: 10.1016/S0959-4388(99)00019-7. [DOI] [PubMed] [Google Scholar]
- 44.Sudlow LC, Jing J, Moroz LL, Gilette R. Serotonin immunoreactivity in the central nervous system of the marine molluscs Pleurobranchaea california and Tritonia diomedea. J Comp Neurol. 1998;395:466–480. [PubMed] [Google Scholar]
- 45.Taghert PH, Goodman CS. Cell determination and differentiation of identified serotonin-immunoreactive neurons in the grasshopper embryo. J Neurosci. 1984;4:989–1000. doi: 10.1523/JNEUROSCI.04-04-00989.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tsvileneva VA, Titova VA. On the brain structures of decapods. Zool Jb Anat. 1985;113:217–266. [Google Scholar]
- 47.Wachowiak M, Ache BW. Morphology and physiology of multiglomerular olfactory projection neurons in the spiny lobster. J Comp Physiol [A] 1994;175:35–48. [Google Scholar]
- 48.Wachowiak M, Diebel CE, Ache BW. Functional organization of olfactory processing in the accessory lobe of the spiny lobster. J Comp Physiol [A] 1996;178:211–226. [Google Scholar]
- 49.Whitaker-Azmitia PM, Druse M, Walker P, Lauder JM. Serotonin as a developmental signal. Behav Brain Res. 1996;73:19–29. doi: 10.1016/0166-4328(96)00071-x. [DOI] [PubMed] [Google Scholar]