

Abstract
The functional interactions between the endogenous cannabinoid and opioid systems were evaluated in pre-proenkephalin-deficient mice. Antinociception induced in the tail-immersion test by acute Δ9-tetrahydrocannabinol was reduced in mutant mice, whereas no difference between genotypes was observed in the effects induced on body temperature, locomotion, or ring catalepsy. During a chronic treatment with Δ9-tetrahydrocannabinol, the development of tolerance to the analgesic responses induced by this compound was slower in mice lacking enkephalin. In addition, cannabinoid withdrawal syndrome, precipitated in Δ9-tetrahydrocannabinol-dependent mice by the injection of SR141716A, was significantly attenuated in mutant mice. These results indicate that the endogenous enkephalinergic system is involved in the antinociceptive responses of Δ9-tetrahydrocannabinol and participates in the expression of cannabinoid abstinence.
Keywords: cannabinoid, opioid, mice, mutation, withdrawal, addiction, tolerance
Recent studies have suggested a functional link between the endogenous opioid and cannabinoid systems (Ayhan et al., 1979; Welch and Stevens, 1992; Smith et al., 1994; Vela et al., 1995; Rodriguez de Fonseca et al., 1997; Tanda et al., 1997;Ledent et al., 1999). Both systems display significant anatomical and functional overlap. Indeed, the administration of cannabinoids and opioids produces similar responses, including antinociception, hypothermia, and reduced locomotion, as well as the development of tolerance and physical dependence after a chronic treatment (Maldonado et al., 1997; Rodriguez de Fonseca et al., 1997; Hutcheson et al., 1998; Ledent et al., 1999). The possible relationships between these two drug dependence processes has recently become a subject of intense investigation (Rodriguez de Fonseca et al., 1997; Tanda et al., 1997;Manzanares et al., 1999; Valverde et al., 2000).
Most of the effects of cannabinoid drugs on the CNS are thought to be mediated by CB1 cannabinoid receptors (Ledent et al., 1999; Zimmer et al., 1999), whereas the opioids activate μ-, δ-, and κ-opioid receptors. Cannabinoid and opioid receptors are members of the G-protein-coupled receptor superfamily and are associated with the inhibition of adenylyl cyclase (Howlett and Fleming, 1984; Felder et al., 1995). Both classes of receptors are abundantly expressed in the central and peripheral nervous systems. Different hypotheses have been put forward to explain the interaction between opioid and cannabinoid systems (Manzanares et al., 1999), such as signal-transduction interactions between both systems, the release of opioid peptides by cannabinoids (Manzanares et al., 1999), or an interaction via the stimulation of the dopaminergic system (Tanda et al., 1997).
In this study, we have evaluated the role of endogenous enkephalins in the behavioral responses induced by acute and chronic stimulation of the CB1 cannabinoid receptors, including the development of cannabinoid tolerance and withdrawal syndrome. For this purpose, we have used pre-proenkephalin-deficient mice that have been generated in our lab (Konig et al., 1996).These mice are hyperalgesic and exhibit a striking set of behavioral abnormalities, including alterations in aggressivity and anxiety (Konig et al., 1996). These behavioral abnormalities support a role for enkephalins in the regulation of emotional behaviors previously suggested by the distribution of enkephalinergic neurons in many areas involved in the control of nociception and mood (Rosenfeld, 1994; Mansour et al., 1995).
MATERIALS AND METHODS
Animals. Enkephalin-deficient mice are maintained in our laboratory as a congenic C57BL/6J-Penk2tmlzim strain. Homozygous mutant and control wild-type C57BL/6J mice were raised from separate colonies from in-house breedings and were kept under identical conditions. Animals were housed in groups of four or five animals per cage. Before the experiment, animals were separated into individual cages. The light cycle was 6:00 A.M. lights on, 6:00 P.M. lights off. Food and water was provided ad libitum. Animals were 8–12 weeks old. Both sexes were equally represented. All animal procedures met the guidelines of the National Institutes of Health detailed in the “Guide for the Care and Use of Laboratory Animals”, the European Communities directive 86/609/EEC regulating animal research, and the Local Ethical Committees.
Receptor-binding studies. Receptor-binding studies were essentially performed as described (Herkenham et al., 1991). Briefly, saggital brain sections (12-μm-thick) were cryostat cut and mounted on gelatin-coated slides. Sections were stored at −35°C. Binding of [3H]CP55,940 (8 nm) was performed in cytomailers (3 hr at 37°C) in 50 mm Tris-HCl, pH 7.4, containing 5% BSA. Nonspecific binding was determined in the presence of 10 μm CP55,244. Slides were washed (4 hr at 0°C) in 50 mm Tris-HCl, pH 7.4, containing 1% BSA, fixed at 25°C in 0.5% formalin in 50 mm Tris-HCL, pH 7.4, and blown dry. Sections were apposed to 3H-sensitive film (Hyperfilm, Amersham Pharmacia) together with 3H standards for 10 d.
Drugs. Δ9-tetrahydrocannabinol (THC) was provided by the National Institute of Drug Abuse (NIDA; Bethesda, MD) as a stock solution (THC, 5 mg/ml) or bought from Sigma (St. Louis, MO) (THC, 100 mg/ml). THC from NIDA was used in all experiments to determine acute effects, whereas THC from Sigma was used in all chronic studies. THC was diluted to a working solution in drug/5% emulphor/0.9% saline solution or in 10% ethanol/10% cremophor EL/80% distilled water. SR141716A was generously provided by Sanofi Recherche (Centre de Labege France) and dissolved in 10% ethanol/10% cremophor EL/80% distilled water. The volume of injection was 1 ml/100 gm for THC, and 1 ml/50 gm for SR141716A.
Acute THC treatment. The acute behavioral and physiological effects of THC were evaluated by determining immobility, catalepsy, analgesia, and hypothermia essentially as described (Martin et al., 1991) using a randomized series of dose treatments. Immediately before the injection of THC or vehicle, animals were weighed, and their body temperatures were determined using a rectal probe (BAT-12; Physitemp Instruments, Clifton, NJ). Twenty minutes after the injection, animals were placed in a well lit open-field apparatus (400 lux; MedAssociates, St. Albans, VT) for 5 min. Subsequently, mice were analyzed in the ring catalepsy test, which consisted of a vertical open tube (5.5 cm diameter) on which the mice were placed. The immobility index (determined from videotaped recordings) was calculated as a percentage of time that the animal spends motionless during the 4 min test session. If an animal fell down or jumped off the ring, it was immediately placed on the tube again. After a maximum of five such escapes the test was terminated. The immobility index (%) was calculated as the time spent motionless × 100/duration of the test session. After the ring test, mice were briefly returned to their home cage. The body temperature was recorded 50 min after the injection.
THC tolerance studies. THC was administered twice a day for 6 d (days 1–5, 9:00 A.M. and 7:00 P.M.; day 6, 9:00 A.M.) at the dose of 20 mg/kg by intraperitoneal route. The responses induced by THC on body weight, rectal temperature, nociception, and spontaneous locomotor activity were evaluated during this chronic treatment. Body weights were recorded for each animal using an electronic balance (Mettler PM 4800, sensitive to 0.01 gm), twice a day before morning and evening injections. The changes in body weight were calculated by subtracting each morning its body weight from the value of the preceding morning. Rectal temperature was measured in each mouse using an electronic thermocouple flexible probe (Panlab, Barcelona, Spain). The probe was placed 3 cm into the rectum of the mice for 30 sec before the temperature was recorded. On days 1 and 2, measures were taken immediately before, and 20 min after, each injection. On days 3, 4, 5, and 6 rectal temperature was evaluated before and 20 min after the morning injection only.
Antinociceptive responses were measured using the tail immersion assay, as described previously (Simonin et al., 1998). The water used for immersing the tail was maintained at a constant temperature of 50 ± 0.5°C. The time (seconds) to withdraw the tail from the bath was measured, with a cutoff latency of 15 sec to prevent tissue damage. Nociceptive measurements were taken just after rectal temperature measurement, 20 min after morning (every day) and evening (days 1 and 2) injections. Spontaneous locomotor activity was measured using activity boxes consisting of an individual plastic rectangular area (30 × 30 cm) isolated in a soundproof room with slight illumination (<20 lux). Animals' activity was recorded during a period of 10 min by a video camera connected to a computer provided with the SMART program (Panlab). The measurements were performed every day 25 min after THC morning injection.
Somatic expression of THC withdrawal. Four hours after the last THC or vehicle injection, mice were placed in a circular clear plastic observation area for a 15 min period of habituation. Immediately after habituation, animals were observed for a further period of 15 min, followed by the administration of SR 14 17 16A (10 mg/kg, i.p.). The mice were observed for an additional 45 min period after SR 141716A injection. Observation of the somatic signs was performed as previously described (Hutcheson et al., 1998; Ledent et al., 1999). The number of bouts of writhing, wet dog shakes, front paw tremor, and sniffing was counted. Penile licking or erection, ataxia, hunched posture, tremor, mastication, and ptosis were scored 1 for appearance or 0 for nonappearance within each 5 min time. A global withdrawal score was calculated for each animal by giving each individual sign a relative weight: 0.9 for the presence of each of the following signs in each period of 5 min: ptosis, body tremor, ataxia, piloerection, hunched posture, mastication, and penile licking; 0.4 for each episode of sniffing, writhing, paw tremor, and wet dog shakes. Values for the global score were from 0 to 100.
Statistical analysis. At least 10 animals were used for each experimental group. Acute effects and global withdrawal scores were compared by using two-way ANOVA (genotype and treatment) between subjects followed by one-way ANOVA and post hoc comparisons (Scheffe's F test). Data from tolerance studies were compared by using the following two-way ANOVA with repeated measurements: day (within subjects) and genotype (between subjects); day and treatment (between subjects); genotype and treatment. One-way ANOVA post hoc comparisons (Scheffe's F test) were subsequently performed when required.
RESULTS
CB1 receptor-binding site in enkephalin-deficient mice
We have previously shown that μ- and δ-opioid receptor-binding sites are upregulated in the brains of enkephalin-deficient mice. To determine whether the lack of enkephalin also led to compensatory changes in central cannabinoid receptors, we analyzed serial brain sections from knock-out and wild-type animals using receptor-binding autoradiography (Fig. 1). There was no apparent difference in the density or distribution of [3H]CP55,940 binding sites between the two genotypes, demonstrating that CB1 receptor expression was not affected by the enkephalin mutation.
Fig. 1.

Comparison of the specific binding of [3H]CP55,940 to cannabinoid receptors in saggital brain sections of wild-type and enkephalin-deficient mice. Specific binding is shown in the top panel. Unspecific binding, determined in the presence of 10 μm CP55,244, is shown in the bottom panel.
Acute responses to THC administration
The acute behavioral and physiological effects of THC were evaluated by measuring open-field activity, ring-catalepsy, and body temperature (Martin et al., 1991) (Fig. 2). We found that locomotor activity in the open field was similarly reduced after THC administration in a dose-dependent manner in wild-type and pre-proenkephalin knock-out mice (treatment effect:F(3,73) = 15.18, p < 0.0001; genotype effect: F(1,73) = 2.07, NS; Fig. 1). For horizontal movements, the reduction was significant with a THC dose of 20 mg/kg (p < 0.0001), whereas vertical movements were significantly reduced at 10 mg/kg (p < 0.0001). THC also produced a profound catalepsy in mice of both genotypes in the ring test (treatment effect: F(3,72) = 26.92,p < 0.0001; genotype effect:F(1,72) = 0.09, NS). This catalepsy was significant at 10 mg/kg THC (p < 0.0001) and was only marginally increased with higher doses. THC also produced a dose-dependent hypothermia in both genotypes (treatment effect:F(3,71) = 45.10, p < 0.0001; genotype effect: F(1,71) = 0.12, NS) that was significant at 10 mg/kg (p < 0.0001).
Fig. 2.
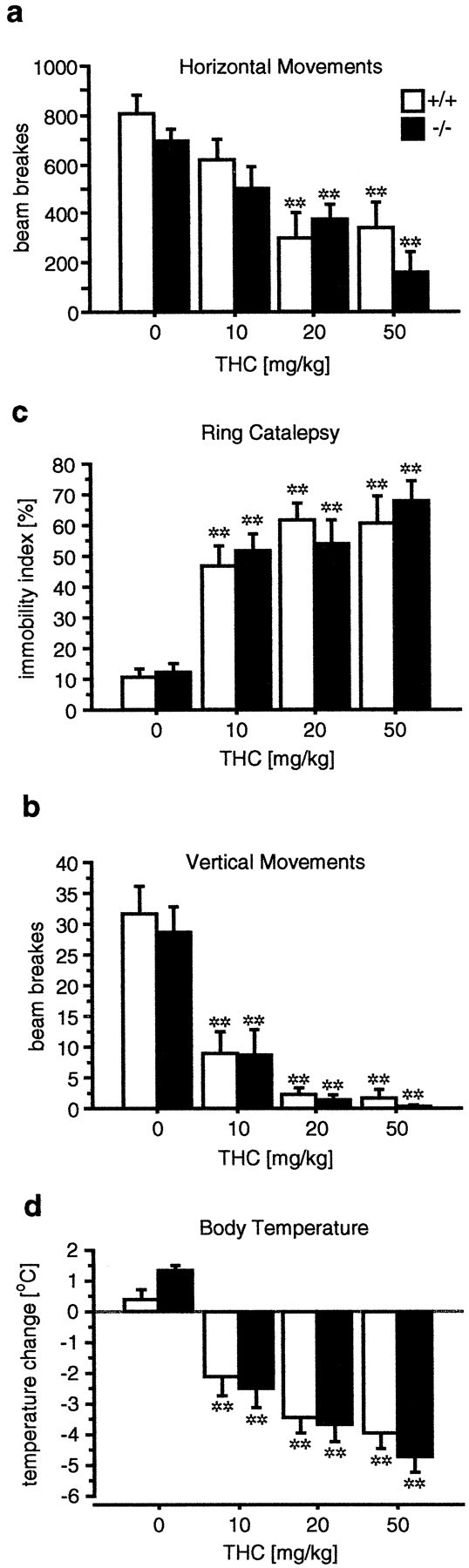
Acute effects of THC are normal in enkephalin-deficient mice. Horizontal (a) and vertical (b) movements in the open field are similarly reduced in a dose-dependent manner in wild-type and knock-out mice. THC increases ring catalepsy (c) and reduces body temperature (d). Significant treatment effects, but no significant genotype effects, were detected using two-way ANOVA in all paradigms. Data are expressed as mean ± SE. *p ≤ 0.05; **p ≤ 0.01 determined by Scheffe's F test.
Tolerance to the behavioral effects of THC
The development of tolerance to the effects induced by THC on rectal temperature, body weight, locomotor activity, and antinociception was evaluated in pre-proenkephalin knock-out and wild-type mice (Fig.3).
Fig. 3.

Development of THC tolerance. a,Tolerance to temperature changes developed similarly in both genotypes.b, Body weights were determined each morning before the THC injection. Changes were calculated by subtracting the weight from the measurement on the previous day. Note that wild-type animals lost significantly more weight than enkephalin knock-out mice.c, No significant tolerance for the effects of THC on spontaneous activity was observed in knock-out or wild-type mice during the course of this experiment. d, THC analgesia was significantly reduced in enkephalin-deficient mice. e,Tolerance to the antinociceptive effects of THC developed slower in mutant mice. Significant differences between the two genotypes were found on days 1 and 2 of testing. Data are expressed as mean ± SE. *p ≤ 0.05 versus vehicle, **p ≤ 0.01 versus vehicle; ★p ≤ 0.05 +/+ versus −/−, ★★p ≤ 0.01 +/+ versus −/−.
Rectal temperature
As in the previous experiment, THC (20 mg/kg) induced a significant decrease in the rectal temperature on day 1 (morning and afternoon measurements) and day 2 (morning measurement) in both genotypes (p < 0.01 for each measurement). Tolerance to the hypothermic effects of 20 mg/kg THC was reached after the fourth injection (day 2) in both genotypes because no significant response was observed at this time point. Tolerance remained for all subsequent measurements (Fig. 3).
Changes in body weight
A significant decrease in body weight was observed in animals treated with THC (20 mg/kg) from day 1 to day 2 in wild-type mice (p < 0.01). The reduction in body weight was less pronounced in knock-out mice than in wild-type animals. Thus, the decrease in the body weight on day 2 was significantly less marked in knock-outs (p < 0.01). At the subsequent time points, the reduction in the body weight was maintained in both genotypes without reaching the control baseline (Fig. 3).
Locomotor activity
As in the previous experiment, spontaneous locomotion was strongly decreased in mutant and wild-type animals after acute injection of THC (20 mg/kg, i.p.). This decrease was maintained during the total period of chronic THC treatment. Thus, no significant tolerance to the hypolocomotion induced by THC was developed in any group (Fig. 3).
Antinociception
THC (20 mg/kg, i.p.) induced an antinociceptive response in the tail-immersion test (p < 0.01 for both genotypes). However, this effect was significantly less intense in mutant mice (p < 0.01). The chronic treatment with THC induced a rapid tolerance to the antinociceptive effect in wild-type mice. In this group, the acute THC response was decreased to 50% in the morning of day 2. The development of tolerance to THC antinociceptive effects was slower in mutant mice, and significant differences between genotypes (p < 0.05) were observed on day 1 (afternoon) and day 2 (morning and afternoon). However, a similar degree of tolerance was observed in both genotypes at the end of THC chronic treatment (Fig. 3).
Somatic expression of THC withdrawal
During the behavioral observation performed before the administration of the CB1 antagonist, SR 141716A, no somatic signs of withdrawal were observed. After the injection of the SR 141716A, the cannabinoid withdrawal syndrome was manifested by the presence of a variety of somatic signs, as previously reported (Hutcheson et al., 1998; Ledent et al., 1999). The intensity of the withdrawal was strongly decreased in pre-proenkephalin knock-out mice. Indeed, ptosis (two-way ANOVA: genotype:F(1,1) = 14.050, p < 0.01; treatment: F(1,1) = 166.265,p < 0.001; g × t:F(1,1) = 14.050,p < 0.01), piloerection (two-way ANOVA: genotype:F(1,1) = 10.989, p < 0.01; treatment: F(1,1) = 33.653,p < 0.001; g × t:F(1,1) = 10.989, p < 0.01), mastication (two-way ANOVA: genotype:F(1,1) = 17.500, p < 0.001; treatment: F(1,1) = 115.759,p < 0.0001; g × t:F(1,1) = 19.901, p < 0.001), body tremor (two-way ANOVA: genotype:F(1,1) = 6.743, p < 0.05; treatment: F(1,1) = 20.040,p < 0.001; g × t:F(1,1) = 21.994, p < 0.001), paw tremor (two-way ANOVA: genotype:F(1,1) = 6.271, p < 0.05; treatment: F(1,1) = 55.045,p < 0.001; g × t:F(1,1) = 9.241, p < 0.01), ataxia (two-way ANOVA: genotype:F(1,1) = 11.830, p < 0.01; treatment: F(1,1) = 4.406,p < 0.05; g × t:F(1,1) = 6.521, p < 0.01), and hunched posture (two-way ANOVA: genotype:F(1,1) = 0.853, NS; treatment:F(1,1) = 12.673, p < 0.01; g × t:F(1,1) = 4.979, p < 0.05) were significantly decreased in knock-out animals (Fig. 4). Only 2 of 9 signs, wet dog shakes and sniffing, were not significantly attenuated in knock-out mice.
Fig. 4.

Severity of THC withdrawal syndrome is reduced in enkephalin-deficient mice. Abstinence was precipitated by the administration of the CB1 antagonist SR141716A (10 mg/kg, i.p.) after chronic THC or vehicle treatment. Seven of 10 abstinence signs were decreased in the knock-out mice. Counted (wet dog shakes, front paw tremor, and sniffing) and checked (tremor, ptosis, ataxia, mastication, hunched posture, piloerection, and penile lick) somatic signs of withdrawal were observed for 45 min immediately after SR 141716A administration. A global withdrawal score was calculated for each animal by giving each individual sign a relative weight. Values for the global score were from 0 to 100. Data are expressed as mean ± SE. *p ≤ 0.05 versus vehicle, **p ≤ 0.01 versus vehicle; ★p ≤ 0.05 +/+ versus −/−, ★★p ≤ 0.01 +/+ versus −/− (Scheffe'sF test).
When the global withdrawal score was calculated and analyzed, a significant decrease in the severity of THC withdrawal was observed in mutant mice (two-way ANOVA: genotype:F(1,1) = 24.051, p < 0.001; treatment: F(1,1) = 163.267,p < 0.001; g × t:F(1,1) = 28.424, p < 0.001). This attenuation of the abstinence was ∼35% compared with wild-type THC-dependent mice.
DISCUSSION
Mouse strains with genetic deletions in the opioid and cannabinoid systems were recently used to study the role of opioids and cannabinoids in the control of nociceptive responses (Kieffer, 1999;Ledent et al., 1999; Steiner et al., 1999; Zimmer et al., 1999; Zimmer and Usdin, 2000). In this manuscript we have analyzed THC responses in enkephalin-deficient mice. We show that the distribution of CB1 receptor-binding sites was similar in wild-type and mutant mice. Whereas most acute effects of THC were largely unaffected by the enkephalin mutation, THC analgesia was significantly decreased in the tail-immersion test in pre-proenkephalin knock-out mice. Chronic THC treatment led to the development of similar levels of tolerance in mice of both genotypes. Strikingly, however, THC withdrawal symptoms, induced by the administration of the CB1 receptor antagonist SR141716A after chronic THC treatment, were largely reduced in enkephalin-deficient mice. These results suggest that enkephalins mediate some of the antinociceptive effects of THC and strongly support a functional interaction between the opioid and cannabinoid systems.
A complementary role in drug addiction has been proposed for tolerance and withdrawal in the opponent process theory (Koob et al., 1989). Although the tolerance to THC antinociceptive effects reached a similar degree at the end of chronic treatment in both genotypes, the development of this tolerance was slower in the mutant group. Thus, as suggested by the opponent process theory, a simultaneous attenuation of THC tolerance and somatic withdrawal was observed in mutant mice
Chronic THC-induced weight loss was significantly smaller in enkephalin knock-outs than in wild-type mice. The acute stimulation of opioid receptors, as well as cannabinoid receptors stimulates food intake, probably through a variety of mechanisms including the control of appetite, meal maintenance, and orosensory reward. The differential effects of chronic THC treatment now indicate that opioid and cannabinoid systems may also interact in the regulation of feeding behavior.
Opioid and cannabinoid receptor distribution overlaps in several neural regions involved in the control of pain and both neurotransmitter systems modulate similar analgesia circuits. Previous studies using selective opioid receptor antagonists suggested that μ and particularly κ-, but not δ-receptors participate in THC analgesia. The molecular and cellular mechanisms of these interactions are not known, but they may involve the release of opioid peptides by THC. Such a release has been demonstrated in the spinal cord for dynorphin (Pugh et al., 1997; Houser et al., 2000).
An interaction between the opioid and cannabinoid systems has also been proposed in drug reward and addiction. Thus, opioid receptor antagonists blocked some effects of THC on the reward system (Gardner and Lowinson, 1991) and produced withdrawal syndrome in THC-dependent rodents (Kaymakcalan et al., 1977). Conversely, cannabinoid agonists alleviated some of the manifestations of naloxone-precipitated withdrawal syndrome (Vela et al., 1995), whereas CB1 antagonists precipitated withdrawal symptoms in rats that were chronically treated with morphine (Navarro et al., 1998). Consistent with this hypothesis, we demonstrate that the somatic expression of the cannabinoid withdrawal was dramatically attenuated in pre-proenkephalin knock-out mice. This result suggests that the endogenous enkephalinergic system also plays a critical role in the development of physical THC dependence. As previously reported (Hutcheson et al., 1998) the behavioral expression of THC withdrawal syndrome in mice was different from the opiate abstinence. Opiate withdrawal is associated in rodents with motoric behaviors such as jumping and hyperactivity, and autonomic signs including lacrimation, rhinorrhea, diarrhea, hypothermia, and hyperalgesia (Marshall and Weinstock, 1971). THC withdrawal induces different effects including wet dog shakes, hunched posture, ptosis, tremor, piloerection, and mastication. As in the case of opiate withdrawal, the large range of behaviors observed during THC abstinence seems to be caused by adaptive neural mechanisms that manifest themselves after the cessation of cannabinoid receptor activation. Indeed, an upregulation of the adenylyl cyclase activity was observed in the cerebellum during THC abstinence (Hutcheson et al., 1998), which resembles the upregulation of this intracellular messenger cascade observed during opioid withdrawal in some brain regions, such as the locus coeruleus (Nestler, 1992). Therefore, common cellular mechanisms may underlie the adaptive responses to chronic exposure to opioids and THC, but the effects produced in vivo by the chronic administration of these agents are different.
Although the possible addictive properties of cannabinoids in humans are still a matter of controversy, there is increasing evidence that cannabinoids, like other drugs of addiction, activate ascending mesolimbic dopaminergic projections (Koob et al., 1992; Rodriguez de Fonseca et al., 1997; Hutcheson et al., 1998) and produce neurochemical changes on the dopaminergic mesolimbic system (Chen et al., 1990; Tanda et al., 1997; Gardner and Vorel, 1998; Gessa et al., 1998). Chronic cannabinoid exposure can induce tolerance in animals and in humans as well as behavioral and physiological withdrawal symptoms (Jones et al., 1981; Mendelson et al., 1984; Beardsley et al., 1986; Duffy and Milin, 1996; Wiesbeck et al., 1996; Diana et al., 1998; Hutcheson et al., 1998). Here, we clearly demonstrate the involvement of the endogenous enkephalinergic system on these adaptive responses to THC.
Ledent et al. (1999) recently studied the effects of opioids in CB1 receptor knock-out mice (Ledent et al., 1999). Tolerance to the behavioral effects of opioids was not altered in these animals, whereas morphine withdrawal was attenuated. Together, these results indicate that the enkephalinergic system is not simply a downstream mediator of cannabinoid effects and vice versa. Rather, it seems that both systems must act in concert to develop all symptoms of drug dependence. A possible substrate for these interactions may be found in the mesolimbic dopaminergic system because both THC and opioid withdrawal induced an increase in corticotrophin release factor in the central nucleus of the amygdala during abstinence (Rodriguez de Fonseca et al., 1997). Indeed, the anandamide/CB1receptor system has been proposed to act as a brake of dopamine D2 receptor family-mediated responses (Giuffrida et al., 1999).
In summary, we demonstrate here that the endogenous enkephalinergic system contributes to THC analgesia and is required for the expression of cannabinoid dependence. These results strongly support the hypothesis that the endogenous opioid and cannabinoid systems are interdependent. Future analysis of the underlying neurobiological processes should help to evaluate whether interactions between cannabinoid and opioid drugs of abuse contribute to their dependence liability, and they should aid in the development of more rational therapeutic treatment of drug dependence.
Footnotes
This work was supported by European Commission Grant BIOMED-2/98–2227 and Fondo de Investigaciones Sanitaria Grant 99/0624 (R.M.), Dr. Esteve S.A. Laboratories, and La Fondation des Treilles, and by a grant from the Land Nordrhein-Westfalen (Innovationsprogramm Forschung) (A.Z.). We thank Eileen Briley and Miles Herkenham for their help with the receptor-binding studies.
Correspondence should be addressed to Andreas Zimmer, Department of Molecular Neurobiology, Clinic of Psychiatry, University of Bonn, Sigmund Freud Strasse 25, 53105 Bonn, Germany, E-mail:neuro@uni-bonn.de, or to Raphael Maldonado, Facultat de Ciéncies de la Salut i de la Vida, Universidat Pompeu Fabra, C/Dr Aiguader 80, 08003 Barcelona, Spain, E-mail: rafael.maldonado@cexs.upf.es.
REFERENCES
- 1.Ayhan IH, Kaymakcalan S, Tulunay FC. Interaction between delta 9-tetrahydrocannabinol and morphine on the motor activity of mice. Psychopharmacology (Berl) 1979;63:169–172. doi: 10.1007/BF00429697. [DOI] [PubMed] [Google Scholar]
- 2.Beardsley PM, Balster RL, Harris LS. Dependence on tetrahydrocannabinol in rhesus monkeys. J Pharmacol Exp Ther. 1986;239:311–319. [PubMed] [Google Scholar]
- 3.Chen JP, Paredes W, Li J, Smith D, Lowinson J, Gardner EL. Delta 9-tetrahydrocannabinol produces naloxone-blockable enhancement of presynaptic basal dopamine efflux in nucleus accumbens of conscious, freely-moving rats as measured by intracerebral microdialysis. Psychopharmacology. 1990;102:156–162. doi: 10.1007/BF02245916. [DOI] [PubMed] [Google Scholar]
- 4.Diana M, Melis M, Muntoni AL, Gessa GL. Mesolimbic dopaminergic decline after cannabinoid withdrawal. Proc Natl Acad Sci USA. 1998;95:10269–73. doi: 10.1073/pnas.95.17.10269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Duffy A, Milin R. Case study: withdrawal syndrome in adolescent chronic cannabis users. J Am Acad Child Adolesc Psychiatry. 1996;35:1618–1621. doi: 10.1097/00004583-199612000-00013. [DOI] [PubMed] [Google Scholar]
- 6.Felder CC, Joyce KE, Briley EM, Mansouri J, Mackie K, Blond O, Lai Y, Ma AL, Mitchell RL. Comparison of the pharmacology and signal transduction of the human cannabinoid CB1 and CB2 receptors. Mol Pharmacol. 1995;48:443–450. [PubMed] [Google Scholar]
- 7.Gardner EL, Lowinson JH. Marijuana's interaction with brain reward systems: update 1991. Pharmacol Biochem Behav. 1991;40:571–580. doi: 10.1016/0091-3057(91)90365-9. [DOI] [PubMed] [Google Scholar]
- 8.Gardner EL, Vorel SR. Cannabinoid transmission and reward-related events. Neurobiol Dis. 1998;5:502–533. doi: 10.1006/nbdi.1998.0219. [DOI] [PubMed] [Google Scholar]
- 9.Gessa GL, Melis M, Muntoni AL, Diana M. Cannabinoids activate mesolimbic dopamine neurons by an action on cannabinoid CB1 receptors. Eur J Pharmacol. 1998;341:39–44. doi: 10.1016/s0014-2999(97)01442-8. [DOI] [PubMed] [Google Scholar]
- 10.Giuffrida A, Parsons LH, Kerr TM, Rodriguez de Fonseca F, Navarro M, Piomelli D. Dopamine activation of endogenous cannabinoid signaling in dorsal striatum. Nat Neurosci. 1999;2:358–363. doi: 10.1038/7268. [DOI] [PubMed] [Google Scholar]
- 11.Herkenham M, Lynn AB, Johnson MR, Melvin LS, de Costa BR, Rice KC. Characterization and localization of cannabinoid receptors in rat brain: a quantitative in vitro autoradiographic study. J Neurosci. 1991;11:563–583. doi: 10.1523/JNEUROSCI.11-02-00563.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Houser SJ, Eads M, Embrey JP, Welch SP. Dynorphin B and spinal analgesia: induction of antinociception by the cannabinoids CP55,940, Delta(9)-THC and anadamide. Brain Res. 2000;857:337–342. doi: 10.1016/s0006-8993(00)01981-8. [DOI] [PubMed] [Google Scholar]
- 13.Howlett AC, Fleming RM. Cannabinoid inhibition of adenylate cyclase. Pharmacology of the response in neuroblastoma cell membranes. Mol Pharmacol. 1984;26:532–538. [PubMed] [Google Scholar]
- 14.Hutcheson DM, Tzavara ET, Smadja C, Valjent E, Roques BP, Hanoune J, Maldonado R. Behavioural and biochemical evidence for signs of abstinence in mice chronically treated with delta-9-tetrahydrocannabinol. Br J Pharmacol. 1998;125:1567–1577. doi: 10.1038/sj.bjp.0702228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jones RT, Benowitz NL, Herning RI. Clinical relevance of cannabis tolerance and dependence. J Clin Pharmacol. 1981;21:143S–152S. doi: 10.1002/j.1552-4604.1981.tb02589.x. [DOI] [PubMed] [Google Scholar]
- 16.Kaymakcalan S, Ayhan IH, Tulunay FC. Naloxone-induced or postwithdrawal abstinence signs in delta9-tetrahydrocannabinol-tolerant rats. Psychopharmacology (Berl) 1977;55:243–249. doi: 10.1007/BF00497855. [DOI] [PubMed] [Google Scholar]
- 17.Kieffer BL. Opioids: first lessons from knockout mice. Trends Pharmacol Sci. 1999;20:19–26. doi: 10.1016/s0165-6147(98)01279-6. [DOI] [PubMed] [Google Scholar]
- 18.Konig M, Zimmer AM, Steiner H, Holmes PV, Crawley JN, Brownstein MJ, Zimmer A. Pain responses, anxiety and aggression in mice deficient in pre-proenkephalin. Nature. 1996;383:535–538. doi: 10.1038/383535a0. [DOI] [PubMed] [Google Scholar]
- 19.Koob GF, Maldonado R, Stinus L. Neural substrates of opiate withdrawal. Trends Neurosci. 1992;15:186–191. doi: 10.1016/0166-2236(92)90171-4. [DOI] [PubMed] [Google Scholar]
- 20.Koob GF, Stinus L, LeMoal M, Bloom FE. Opponent process theory of motivation: neurobiological evidence from studies of opiate dependence. Neurosci Biobehav Rev. 1989;13:135–140. doi: 10.1016/s0149-7634(89)80022-3. [DOI] [PubMed] [Google Scholar]
- 21.Ledent C, Valverde O, Cossu G, Petitet F, Aubert JF, Beslot F, Bohme GA, Imperato A, Pedrazzini T, Roques BP, Vassart G, Fratta W, Parmentier M. Unresponsiveness to cannabinoids and reduced addictive effects of opiates in CB1 receptor knockout mice. Science. 1999;283:401–404. doi: 10.1126/science.283.5400.401. [DOI] [PubMed] [Google Scholar]
- 22.Maldonado R, Saiardi A, Valverde O, Samad TA, Roques BP, Borrelli E. Absence of opiate rewarding effects in mice lacking dopamine D2 receptors. Nature. 1997;388:586–589. doi: 10.1038/41567. [DOI] [PubMed] [Google Scholar]
- 23.Mansour A, Fox CA, Akil H, Watson SJ. Opioid-receptor mRNA expression in the rat CNS: anatomical and functional implications. Trends Neurosci. 1995;18:22–29. doi: 10.1016/0166-2236(95)93946-u. [DOI] [PubMed] [Google Scholar]
- 24.Manzanares J, Corchero J, Romero J, Fernandez-Ruiz JJ, Ramos JA, Fuentes JA. Pharmacological and biochemical interactions between opioids and cannabinoids. Trends Pharmacol Sci. 1999;20:287–294. doi: 10.1016/s0165-6147(99)01339-5. [DOI] [PubMed] [Google Scholar]
- 25.Marshall I, Weinstock M. Quantitative method for assessing one symptom of the withdrawal syndrome in mice after chronic morphine administration. Nature. 1971;234:223–224. doi: 10.1038/234223a0. [DOI] [PubMed] [Google Scholar]
- 26.Martin BR, Compton DR, Thomas BF, Prescott WR, Little PJ, Razdan RK, Johnson MR, Melvin LS, Mechoulam R, Ward SJ. Behavioral, biochemical, and molecular modeling evaluations of cannabinoid analogs. Pharmacol Biochem Behav. 1991;40:471–478. doi: 10.1016/0091-3057(91)90349-7. [DOI] [PubMed] [Google Scholar]
- 27.Mendelson JH, Mello NK, Lex BW, Bavli S. Marijuana withdrawal syndrome in a woman. Am J Psychiatry. 1984;141:1289–1290. doi: 10.1176/ajp.141.10.1289. [DOI] [PubMed] [Google Scholar]
- 28.Navarro M, Chowen J, Rocio ACM, del Arco I, Villanua MA, Martin Y, Roberts AJ, Koob GF, de Fonseca FR. CB1 cannabinoid receptor antagonist-induced opiate withdrawal in morphine-dependent rats. NeuroReport. 1998;9:3397–402. doi: 10.1097/00001756-199810260-00012. [DOI] [PubMed] [Google Scholar]
- 29.Nestler EJ. Molecular mechanisms of drug addiction. J Neurosci. 1992;12:2439–2450. doi: 10.1523/JNEUROSCI.12-07-02439.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pugh G, Jr, Mason DJ, Jr, Combs V, Welch SP. Involvement of dynorphin B in the antinociceptive effects of the cannabinoid CP55,940 in the spinal cord. J Pharmacol Exp Ther. 1997;281:730–737. [PubMed] [Google Scholar]
- 31.Rodriguez de Fonseca F, Carrera MRA, Navarro M, Koob GF, Weiss F. Activation of corticotropin-releasing factor in the limbic system during cannabinoid withdrawal. Science. 1997;276:2050–2054. doi: 10.1126/science.276.5321.2050. [DOI] [PubMed] [Google Scholar]
- 32.Rosenfeld JP. Interacting brain stem components of opiate-activated, descending, pain-inhibitory systems. Neurosci Biobehav Rev. 1994;18:403–409. doi: 10.1016/0149-7634(94)90053-1. [DOI] [PubMed] [Google Scholar]
- 33.Simonin F, Valverde O, Smadja C, Slowe S, Kitchen I, Dierich A, Le Meur M, Roques BP, Maldonado R, Kieffer BL. Disruption of the kappa-opioid receptor gene in mice enhances sensitivity to chemical visceral pain, impairs pharmacological actions of the selective kappa-agonist U-50,488H and attenuates morphine withdrawal. EMBO J. 1998;17:886–897. doi: 10.1093/emboj/17.4.886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Smith PB, Welch SP, Martin BR. Interactions between delta 9-tetrahydrocannabinol and kappa opioids in mice. J Pharmacol Exp Ther. 1994;268:1381–1387. [PubMed] [Google Scholar]
- 35.Steiner H, Bonner TI, Zimmer AM, Kitai ST, Zimmer A. Altered gene expression in striatal projection neurons in CB1 cannabinoid receptor knockout mice. Proc Natl Acad Sci USA. 1999;96:5786–5790. doi: 10.1073/pnas.96.10.5786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tanda G, Pontieri FE, Di Chiara G. Cannabinoid and heroin activation of mesolimbic dopamine transmission by a common mu1 opioid receptor mechanism. Science. 1997;276:2048–2050. doi: 10.1126/science.276.5321.2048. [DOI] [PubMed] [Google Scholar]
- 37.Valverde O, Ledent C, Beslot F, Parmentier M, Roques BP. Reduction of stress-induced analgesia but not of exogenous opioid effects in mice lacking CB1 receptors. Eur J Neurosci. 2000;12:1–7. doi: 10.1046/j.1460-9568.2000.00929.x. [DOI] [PubMed] [Google Scholar]
- 38.Vela G, Ruiz-Gayo M, Fuentes JA. Anandamide decreases naloxone-precipitated withdrawal signs in mice chronically treated with morphine. Neuropharmacology. 1995;34:665–668. doi: 10.1016/0028-3908(95)00032-2. [DOI] [PubMed] [Google Scholar]
- 39.Welch SP, Stevens DL. Antinociceptive activity of intrathecally administered cannabinoids alone, and in combination with morphine, in mice. J Pharmacol Exp Ther. 1992;262:10–18. [PubMed] [Google Scholar]
- 40.Wiesbeck GA, Schuckit MA, Kalmijn JA, Tipp JE, Bucholz KK, Smith TL. An evaluation of the history of a marijuana withdrawal syndrome in a large population. Addiction. 1996;91:1469–1478. [PubMed] [Google Scholar]
- 41.Zimmer A, Usdin T. Examination of the opioid system using targeted gene deletions. In: Wood JN, editor. Molecular basis of pain Induction. Wiley-Liss; New York: 2000. pp. 193–207. [Google Scholar]
- 42.Zimmer A, Zimmer AM, Hohmann AG, Herkenham M, Bonner TI. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice. Proc Natl Acad Sci USA. 1999;96:5780–5785. doi: 10.1073/pnas.96.10.5780. [DOI] [PMC free article] [PubMed] [Google Scholar]