

Abstract
Background
Melatonin is a pleiotropic signaling molecule that plays multifarious roles in plants stress tolerance. The polyamine (PAs) metabolic pathway has been suggested to eliminate the effects of environmental stresses. However, the underlying mechanism of how melatonin and PAs function together under heat stress largely remains unknown. In this study, we investigated the potential role of melatonin in regulating PAs and nitric oxide (NO) biosynthesis, and counterbalancing oxidative damage induced by heat stress in tomato seedlings.
Results
Heat stress enhanced the overproduction of reactive oxygen species (ROS) and damaged inherent defense system, thus reduced plant growth. However, pretreatment with 100 μM melatonin (7 days) followed by exposure to heat stress (24 h) effectively reduced the oxidative stress by controlling the overaccumulation of superoxide (O2•−) and hydrogen peroxide (H2O2), lowering the lipid peroxidation content (as inferred based on malondialdehyde content) and less membrane injury index (MII). This was associated with increased the enzymatic and non-enzymatic antioxidants activities by regulating their related gene expression and modulating the ascorbate–glutathione cycle. The presence of melatonin induced respiratory burst oxidase (RBOH), heat shock transcription factors A2 (HsfA2), heat shock protein 90 (HSP90), and delta 1-pyrroline-5-carboxylate synthetase (P5CS) gene expression, which helped detoxify excess ROS via the hydrogen peroxide-mediated signaling pathway. In addition, heat stress boosted the endogenous levels of putrescine, spermidine and spermine, and increased the PAs contents, indicating higher metabolic gene expression. Moreover, melatonin-pretreated seedlings had further increased PAs levels and upregulated transcript abundance, which coincided with suppression of catabolic-related genes expression. Under heat stress, exogenous melatonin increased endogenous NO content along with nitrate reductase- and NO synthase-related activities, and expression of their related genes were also elevated.
Conclusions
Melatonin pretreatment positively increased the heat tolerance of tomato seedlings by improving their antioxidant defense mechanism, inducing ascorbate–glutathione cycle, and reprogramming the PAs metabolic and NO biosynthesis pathways. These attributes facilitated the scavenging of excess ROS and increased stability of the cellular membrane, which mitigated heat-induced oxidative stress.
Electronic supplementary material
The online version of this article (10.1186/s12870-019-1992-7) contains supplementary material, which is available to authorized users.
Keywords: Melatonin, Heat stress, Polyamines, NO biosynthesis, Redox, AsA-GSH cycle, Tomato
Background
Global warming has led to climate change, including heat stress and these changes considerate as a major threat for worldwide crop production [1]. Heat stress can cause misfolds or disorganized cellular homeostasis because of excess reactive oxygen species (ROS) accumulation, distorted protein structure, impeded protein synthesis, and overall cell division and growth disruption from reduced water content [2–4]. Heat shock-induced oxidative damage occurs as a result of excess formation of singlet oxygen (1O2), superoxide radical (O2•−), hydrogen peroxide (H2O2), and hydroxyl radical (OH•) under heat stress [5]; this leads to overproduction of malondialdehyde (MDA) and thus reduces membrane stability, permeability, and mobility, and impairs protein membrane polymerization [6, 7]. As a sessile organism, in an unfavorable environment, plants develop an inherent antioxidative defense strategy to detoxify excess ROS, which helps to protect them from oxidative damage [8]. This efficient anti-oxidative defense mechanism consists of different enzymatic antioxidants, such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), ascorbate peroxidase (APX), glutathione reductase (GR), monodehydroascorbate reductase (MDHAR), and dehydroascorbate (DHAR), and non-enzymatic antioxidants, such as ascorbate (AsA), glutathione (GSH), carotenoids, and phenols [2, 9, 10]. Additionally, heat-shock proteins (HSPs) play vital roles in ROS scavenging [11], because heat stress induces APX and CAT production [12]. Moreover, heat shock transcription factors A2 (HsfA2) plays a key role in the regulation of expression of heat-shock proteins, ascorbate peroxidase 2 and galactinol synthase 1 and 2 under high-temperature challenged [13].
Melatonin (N-acetyl-5-methoxytryptamine) is a naturally occurring low-molecular-weight multi-regulatory molecule that exists in all living organisms, including plants and animals [14, 15]. Since its detection in plants, scientists’ curiosity regarding melatonin has increased, because of its diversified biological role as a plant master regulator and defensive roles in capricious environmental conditions, such as extreme temperatures, salinity, drought, heavy metals, UV radiation, and oxidative stress [15–18]. Melatonin also accelerates seed germination [19], influences root and plant architecture [20], enhances growth vitality, ameliorates leaf senescence [21], regulates nitrogen metabolism [21], and alters physiological processes by inducing differential gene expression [16]. The most important function of melatonin is ROS detoxification through the production of free radicle scavengers (H2O2, O2•−) and modulation of both antioxidant enzyme activity and concentration [22, 23]. Rodriguez et al. [24] reported that pre-treated melatonin protects oxidative damage in cucumber through melatonin-mediated redox signaling pathways. Under both dark and light conditions, melatonin increases APX and CAT activity, and elevates AsA and GSH content; the AsA–GSH cycle also helps to reduce the dark-induced senescence [12, 25]. There is lack of research on how melatonin protects seedlings from possible damage caused by thermal stress. Consequently, the aim of this research was to elucidate melatonin’s mode of action.
Polyamines (PAs) are essential stress response biomolecules; they are small molecular weight nitrogenous compounds that exist ubiquitously in plants, mostly as putrescine (Put), spermidine (Spd), and spermine (Spm) [26]. PAs possess a wide variety of functions, including plant morphogenesis, reproductive stimulation, and delayed leaf senescence, and they play key roles against abiotic stresses, such as extreme temperature (high and low), salt, drought, heavy metals, osmotic stress, ultraviolet radiation stress, and submerged stress [27, 28]. Additionally, PAs have a cationic charge that helps uphold membrane integrity, assists smooth enzyme function, and protects DNA, RNA, and protein structure. Therefore, plant physiological, biochemical, and molecular activities are enhanced through interactions with nucleic acids, proteins, and phospholipids [29]. Ke et al. [28] determined that supplemental melatonin alleviates salinity stress in wheat seedlings by regulating PAs metabolism. Melatonin increases iron-deficiency tolerance through increased accumulation of PA-mediated nitric oxide (NO) [30]. Shi et al. [31] noted that the PAs metabolic system also changed under oxidative stress conditions with melatonin pretreatment in Bermuda grass. Moreover, melatonin increased Put and Spd levels in carrot suspension cells, which helped reduce cold-induce apoptosis [32]. Additionally, melatonin pretreatment alleviated chilling stress in harvested peach fruits [33] and cucumber seedlings [34] which are closely related to PAs metabolism. These research findings revealed that melatonin may play critical roles in capricious environments by mediating PAs metabolism. Alternatively, the signaling molecule NO functions as a mediator of PAs metabolism and plant hormones, and also triggers NO biosynthesis [35]. However, until now, how melatonin regulates PAs metabolism have not been entirely understood. We hypothesize that melatonin may be associated with PAs via the NO biosynthesis pathway, thus helping plants cope with high-temperature challenges. Correlations between melatonin biosynthesis and PAs metabolism, and their underlying mechanisms could provide a novel insight that can help to promote plant production and protection.
Result
Melatonin improved morphological parameters in tomato seedling under heat stress
Fresh weight (FW) and dry weight (DW) of shoots and roots significantly decreased in high-temperature treatment seedlings, especially root FW and DW (Table 1). Shoot and root FW were reduced by around 30 and 12%, respectively, under heat stressed seedlings compared to normal growth conditions. Conversely, exogenous melatonin mitigated temperature-induced inhibition of growth components and facilitated better growth.
Table 1.
Effects of melatonin on the morphology of heat stress exposed tomato seedlings
| Treatments | Shoot fresh weight (g plant−1) | Shoot dry weight (g plant− 1) | Root fresh weight (g plant− 1) | Root dry weight (g plant− 1) |
|---|---|---|---|---|
| CK | 8.65 ± 0.17a | 0.88 ± 0.05a | 2.30 ± 0.26b | 0.16 ± 0.01ab |
| MT | 8.90 ± 0.15a | 0.91 ± 0.06a | 4.17 ± 0.13a | 0.21 ± 0.02a |
| HT | 6.03 ± 0.18b | 0.47 ± 0.02c | 2.04 ± 0.15b | 0.12 ± 0.05b |
| MT + HT | 8.03 ± 0.16a | 0.65 ± 0.03b | 4.04 ± 0.21a | 0.20 ± 0.02a |
Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
CK: Control, MT: 100 μM melatonin, HT: High temperature (42 °C), MT + HT: 100 μM melatonin + high temperature (42 °C)
Melatonin controlled the overaccumulation of ROS in heat stressed tomato seedlings
To investigate if melatonin alleviates heat stress-induced oxidative stress, we first detected H2O2 and O2•− generation in tomato leaves by histochemical staining. As shown in Fig. 1a, b, we observed deeper blue staining, which indicates O2•− production, and acute brown staining, which denotes H2O2 production, on the leaf surface of high temperature-stressed tomato seedlings. Conversely, the differences among the other treated seedlings were less in their leaf blades, which indicates that melatonin inhibits ROS overproduction under stress conditions.
Fig. 1.

Effects of Melatonin (100 μM) on accumulation of (A) superoxide ion (O2•−), (B) hydrogen peroxide (H2O2), (C) generation of superoxide ion (O2•−) and (D) content of hydrogen peroxide (H2O2) in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. CK: Control; MT: 100 μM melatonin; HT: High temperature (42 °C); MT + HT: 100 μM melatonin + high temperature (42 °C). Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
To evaluate the ROS accumulation trends under melatonin and/or heat stress, we further examined the generation rates of O2•− and H2O2 in tomato leaves (Fig. 1c, d). Relative to the control, sharp increases in O2•− and H2O2 production were observed in the leaf tissues upon subjected to heat stress. Under heat stress, H2O2 and O2•− contents increased by 32 and 137%, respectively, compared with the control seedlings. By contrast, application of melatonin increased the heat-stress tolerance of seedlings by reducing the formation rates of H2O2 and O2•− in leaf tissue by 15 and 36%, respectively, compared with the plants grown solely in the high-temperature environment.
Melatonin maintained cellular membrane integrity in tomato leaves under heat stress
High temperatures destroyed tomato seedling leaves cellular membranes as indicated by MII (80%) and the higher accumulation of MDA content (45%) compared with the control (Fig. 2). Application of 100 μM melatonin was more effective for overcoming harsh impacts of heat stress, as shown by a substantial reduction of MII (29%) and lower MDA concentration (16%) compared with untreated heat-stressed plants.
Fig. 2.
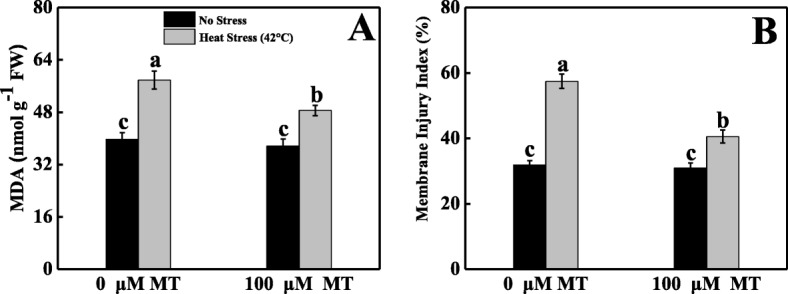
Effects of Melatonin (100 μM) on (A) malondialdehyde (MDA) content and (B) membrane injury index (MII) in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
Melatonin enhanced proline metabolism and RWC in tomato seedlings under heat stress
Heat-stressed plants induced elevated proline biosynthesis that was 158% greater compared with the corresponding control (Fig. 3a). The melatonin-pretreatment combined with heat-stressed seedlings showed a maximum proline content that was 212% greater than that of control. To verify this, we further quantified the gene expression of P5CS, which is responsible for proline biosynthesis. The P5CS expression pattern was substantially upregulated (1.62-fold) in heat-stressed seedlings compared with the control plants. Melatonin pretreatment in heat-stressed seedlings further markedly upregulated P5CS expression by 6-folds in contrast with seedlings that were grown only heat-stressed conditions (Fig. 3b).
Fig. 3.

Effects of Melatonin (100 μM) on (A) proline content and (B) relative transcript expression of P5CS and (C) relative water content (REL) in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
For RWC, in respect to normal plants, high-temperature challenge seedlings considerably decreased the RWC by 10%; supplemental melatonin application curtailed a significant amount of water loss from their tissues, and these plants contained 7% more RWC than the untreated heat-stressed seedlings (Fig. 3c).
Melatonin balanced the antioxidant defense system in tomato seedlings under heat stress
To evaluate the role of melatonin in oxidative stress remediation, we investigated the activities of antioxidant enzymes upon exposure to heat stress (Fig. 4). Under heat stress, SOD activity substantially declined and was 1.89-fold lower than in the control; alternatively, SOD activity significantly increased and was 1.29-fold higher in plants with melatonin pretreatment under high temperatures than untreated heat-stressed seedlings (Fig. 4a). Heat stress caused a marked decrease of 41% in CAT activity in leaves compared with normally grown leaves; melatonin-pretreated heat-stressed seedlings had upwards of 36% greater in CAT activity than untreated heat-stressed leaves (Fig. 4b). Melatonin-pretreated heat-stressed seedlings had dramatically increased POD activity (by 23%) in contrast with untreated heat-stressed seedlings; however, relative to control plants, POD activity sharply declined in untreated heat-stressed seedlings (29%, Fig. 4c). Under heat stress, APX activity decreased by 60% compared with the control, whereas, melatonin pretreatment in heat-stressed seedlings resulted in 230% greater APX activity than that of untreated heat-stressed seedlings (Fig. 4d).
Fig. 4.

Effects of Melatonin (100 μM) on antioxidant enzymes activities and their related transcript expression of (A, E) superoxide dismutase (SOD), (B, F) catalase (CAT), (C, G) peroxidase (POD) and (D, H) ascorbate peroxidase (APX) in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
Melatonin induced the AsA–GSH cycle and homeostasis in tomato seedlings under heat stress
As displayed in Fig. 5a, AsA content was markedly increased by 47% after heat stress in contrast to the control plants. Moreover, in response to normal seedlings, AsA content was further increased by 62% in melatonin-pretreated heat-stressed seedlings. Under exposure to heat stress, the GSH content was remarkably increased (168%) in contrast to the corresponding control seedlings (Fig. 5b). Alternatively, melatonin-pretreated heat-stressed seedlings had 28% greater GSH content with respect to the untreated heat-stressed seedlings.
Fig. 5.
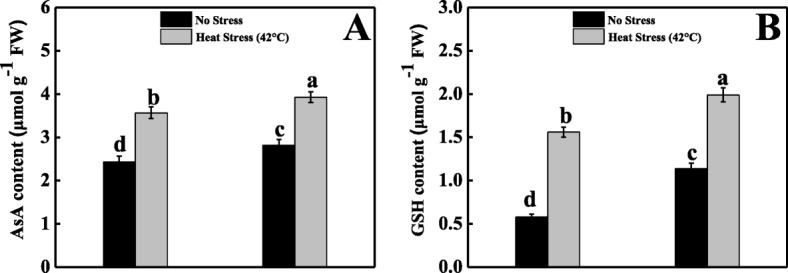
Effects of Melatonin (100 μM) on (A) ascorbic acid (AsA) and (B) glutathione (GSH) content in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
There were no significant differences in GR activity among the treatments except in the untreated heat-stressed seedlings (Fig. 6a). However, GR activity sharply decreased by 45% in thermal-stressed plants with respect to the control seedlings. By contrast, when plants treated with melatonin followed by high-temperature exposure showed upregulation of GR activity by 94% than seedlings subjected to heat-stressed alone.
Fig. 6.

Effects of Melatonin (100 μM) on antioxidant enzymes activities and their related transcript expression of (A, E) glutathione reductase (GR), (B, F) monodehydroascorbate reductase (MDHAR), (C, G) dehydroascorbate reductase (DHAR) and (D, H) glutathione S-transferase (GST) in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
However, marked decreases of MDHAR and DHAR activities were observed in heat-stressed seedlings compared to other seedlings. Additionally, in respect to only heat-stressed seedlings MDHAR and DHAR enzyme contents rose by 91 and 56%, respectively, in melatonin-pretreated heat-stressed seedlings (Fig. 6b, c).
In untreated heat-stressed seedlings, the GST activity slightly decreased by around 19% compared with the control groups. In contrast, melatonin pretreatment increased GST activity by 39% than untreated heat-stressed seedlings (Fig. 6d).
Melatonin modulated the transcription of enzymatic antioxidants under heat stress
To elucidate the molecular mechanism underlying how melatonin alleviates heat stress-induced oxidative damage, the transcript levels of some key genes that encode antioxidant enzymes were assayed. The results showed that SOD (Fig. 4e), CAT (Fig. 4f), APX (Fig. 4h), and GST (Fig. 6h) gene expression levels were upregulated by 1.6-, 1.2-, 3.2-, and 1.3-fold, respectively, in response to their corresponding control groups. Alternatively, the application of exogenous melatonin upregulated SOD, CAT, POD (Fig. 4g), APX, GR (Fig. 6e), MDHAR (Fig. 6f), DHAR (Fig. 6g), and GST expression by 2.1-, 1.3-, 1.7-, 3.0-, 5.6-, 2.4-, 1.8-, and 3.4-fold, respectively, compared with untreated heat-stressed seedlings.
Melatonin regulated the RBOH expression in tomato leaves under heat stress
As indicated in Fig. 7a, we analyzed the tomato RBOH expression level. The RBOH transcript levels were prominently elevated in heat-stressed seedlings than control ones. Conversely, exogenously applied melatonin with subsequent high-temperature exposure also increased RBOH expression by 1.89-fold compared with untreated heat-stressed plants.
Fig. 7.

Effects of Melatonin (100 μM) on relative transcript expression of (A) Respiratory burst oxidase (RBOH) and (B) heat shock factor A 2 (HsfA2) (C) heat shock protein 90 (HSP90) in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
Melatonin regulated the heat shock-related genes expression in tomato leaves under heat stress
As shown in Fig. 7b, c, HSP90 and HsfA2 expression levels were increased by 4.1- and 3.26-fold, respectively, in untreated heat-stressed seedlings compared with control seedlings. In contrast to untreated heat-stressed seedlings, the HSP90 and HsfA2 transcript levels were sharply upregulated by 1.29- and 1.55-fold, respectively, in melatonin-pretreated heat-stressed seedlings.
Melatonin modulated endogenous levels of PAs and their genes expression in tomato leaves under heat stress
We quantified endogenous free PAs accumulation to explicate how PAs and melatonin coordinate in order to eliminate the adverse effects of thermal stress. The Put, Spd, and Spm contents significantly increased in heat-stressed seedlings by 25.26, 48.24, and 24.43%, respectively, compared with the control group (Fig. 8). Melatonin supplementation further increased Put by 82.52%, Spd by 78.72%, and Spm by 247.80% relative to their corresponding control plants.
Fig. 8.

Effects of Melatonin (100 μM) on endogenous free polyamines content of (A) Putrescine (Put), (B) Spermidine (Spd), (C) Spermine (Spm) in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
To reveal the expression profile of PAs metabolism, we performed heatmap visualization and hierarchical cluster analysis (Fig. 9). In the presence of melatonin, the mRNA levels of PAs metabolic genes showed higher expression than control. High-temperature stress exposure with or without melatonin treatments caused upregulation of the transcript levels of the assayed genes. The ADC1, ADC2, and ODC1 mRNA levels were increased in stressed seedlings, with and without melatonin, whereas ODC2 expression was downregulated in all heat-stressed seedlings compared with control seedlings. Similarly, the SAMDC1, SAMDC2, and SPMS transcript abundance also increased in response to either all heat-stressed seedlings compared with control seedlings. Furthermore, in contrast to untreated heat-stressed seedlings, the SPDS1, SPDS2, SPDS3, SPDS4, and SPDS5 expression levels were also upregulated in melatonin-pretreated plants. Interestingly, the PAO1 and PAO2 transcript levels were downregulated in melatonin-pretreated seedlings than untreated heat-stressed plants, which indicates that melatonin might inhibit heat-induced damage via the PAs metabolic pathway and not a catabolic process.
Fig. 9.

Heat-map representing the relative transcript abundance of differentially expressed Polyamines genes in leaves of tomato seedlings under high temperature stress with or without melatonin pretreatment and also hierarchical cluster analysis were used. The gene expression intensity extended from blue color (low) to red color (high). CK: Control; MT: 100 μM melatonin; HT: High temperature (42 °C); MT + HT: 100 μM melatonin + high temperature (42 °C)
Melatonin regulated the NO biosynthesis pathway under heat stress
As displayed in Fig. 10, untreated heat-stressed plants released 38% more NO than control seedlings. In contrast, melatonin pretreatment further increased the NO content in thermal-stressed tomato seedlings by approximately 205% than control plants. NR activity increased upon exposure to high temperatures with or without melatonin applied to the seedlings. Melatonin-pretreated heat-stressed seedlings markedly increased NR activity in respect to control plants by around 218%.
Fig. 10.
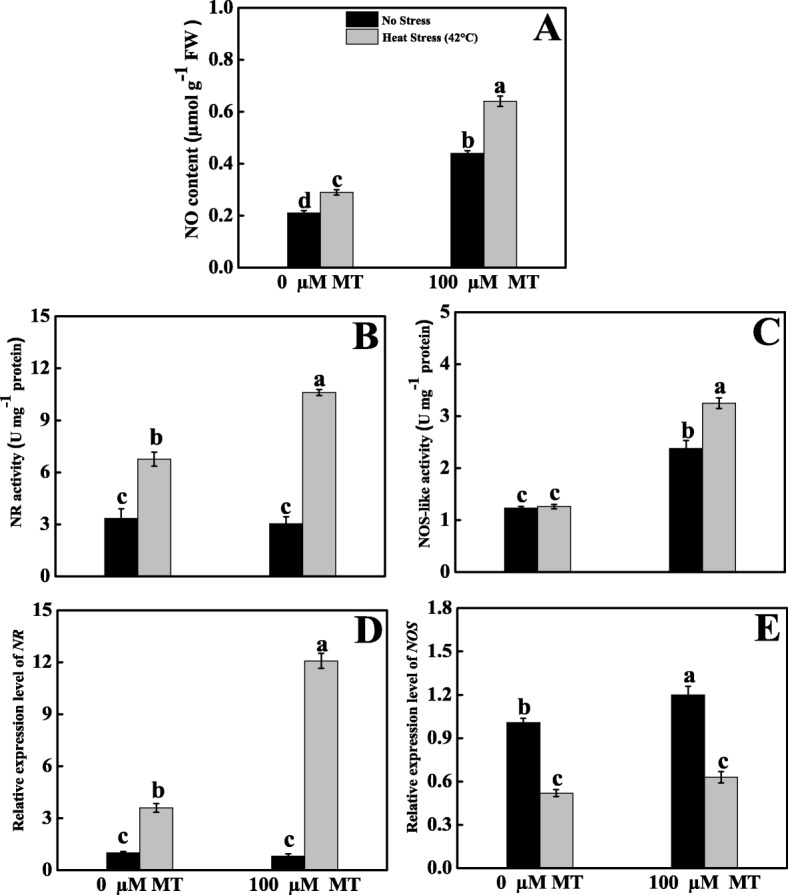
Effects of Melatonin (100 μM) on NO biosynthesis pathway in leaves of tomato seedlings in presence or absence of high temperature (42 °C) stress. A NO content, B, D NR activity and its transcript expression level, C, E NOS-like activity and its transcript level. Data represent as a mean of standard deviation (SD) of three replications. Different letters indicate significant differences according to Tukey’s HSD test at P ≤ 0.05
Alternatively, in the case of NOS-like activity, there was not significant variation between control and heat-stressed seedlings. The NOS-like activity was dramatically increased by 164% in melatonin-pretreated heat-stressed seedlings in contrast with control seedlings. The relative transcript level of NR was increased (2.6- fold) in untreated heat-stressed seedlings than control seedlings. However, NR expression was further exponentially upregulated (2.35- fold) in melatonin-pretreated heat-stressed seedlings in respect to untreated heat-stressed seedlings. However, in the case of NOS, expression was downregulated in all heat-stressed seedlings with or without melatonin pretreatment compared with the control seedlings.
Discussion
Melatonin is a pleiotropic molecule that is involved in diverse plant physiological functions, including seed morphogenesis, growth, and development; root architecture; photosynthesis; and chlorophyll pigment production; and it is a plant master regulator and defensive player in capricious environments [16]. Cellular oxidative damage is a stress marker of high-temperature stress, and H2O2 and O2•− generation, MDA content, and MII are the representative of these stress markers. Heat-stressed tomato leaves conspicuously displayed deep blue spots, which indicated that they produced more O2•−. The leaves also had dark brown patches, which indicated greater accumulation of H2O2 under the same stress conditions. These staining symptoms were supported by quantification of H2O2, and O2•−, which showed markedly higher generation under elevated temperatures compared with melatonin-pretreated heat-stressed seedlings (Fig. 1). These findings are consistent with prior findings that melatonin decreased H2O2 and O2•− accumulation in kiwifruit [36], watermelon [37], cucumber [38], and Malus hupehensis [39]. The probable mechanism underlying this decrease may be that melatonin acts as an electron donor [40]. MDA often represents an essential stress symptom that forms through an auto-oxidative chain reaction as a result of ROS-induced bio-membrane damage [41]. However, in a capricious environment, the MII and MDA concentration are predominantly associated with each other. Plants exposed to heat stress had sharply increased MDA levels that could potentially damage the plasma membrane integrity, which elevated MII in tomato seedlings (Fig. 2). Melatonin application decreased both MDA and MII, which is consistent with the findings of previous studies on kiwifruit [36, 42], Bermuda grass [31], and tomato [43] under various abiotic stresses. These results indicate that melatonin may be able to repair the disrupted cellular membrane and reduce heat-induced oxidative damage by balancing ROS in a high-temperature environment. HsfA2 and HSP90 are the key regulators that stimulate ROS detoxification through the H2O2-mediated signaling pathway and, therefore, increase plant thermo-tolerance. In this study, melatonin-pretreated tomato seedlings had upregulated HsfA2 and HSP90 expression compared with untreated heat-stressed seedlings (Fig. 7), which indicates that melatonin ameliorated the heat stress-induced oxidative damage caused by HsfA2 and HSP90 activation. A recent report also revealed that HsfA2 plays roles in H2O2 signaling and increases heat stress memory subsistence, whereas HSP90 coordinates DNA-binding enhancement process and HSF balanced in plants exposed to heat stress, and this whole mechanism might be related to melatonin-mediated heat tolerance [22, 44–46].
Melatonin is a dynamic antioxidant [47, 48] that extensively stimulates cellular redox homeostasis by enhancing the activity of enzymatic antioxidants, including SOD, CAT, POD, APX, GR, MDAR, and DHAR, and non-enzymatic antioxidants, including AsA and GSH [49–52]. Therefore, melatonin helps detoxify excess ROS, which helps plants survive under stressful conditions. We assayed enzyme activity and conducted expression analysis of antioxidant-related genes and observed that all the antioxidant-related enzymes activities were reduced under thermal stress. Conversely, melatonin-pretreated heat-stressed plants showed higher SOD, CAT, and POD activity relative to untreated heat-stressed plants (Fig. 4). The first mechanism of defense against ROS in plants is through SOD, which eliminates O2•− by converting it into O2 and H2O2 [53]. In addition, CAT and POD also actively participate in scavenging H2O2, which they convert into H2O and O2 [54], which indicates that these enzymes have a vital role in scavenging more H2O2, and these and the MDA results were consistent with those of melatonin-treated kiwifruit [42], wheat [55], and tea [17].
Moreover, we speculated that heat stress elevated the proline content, upregulated the proline biosynthesis gene (P5CS), and lowered the water content in leaves, whereas melatonin pretreatment further increased the proline level, P5CS expression, and leaf water content. This finding indicates that melatonin has the potential to help leaves maintain a higher water level and lower cellular osmotic potential (Fig. 3) by the proline biosynthesis pathway, which enhanced the plants’ ability to cope with heat stress [56]; this was also supported by previous experimental results [57].
Alternatively, ascorbate is a vital antioxidant enzyme that substantially detoxifies ROS; APX and GR are also crucial enzymes in the AsA–GSH cycle. The activities of APX, MDHAR, DHAR, and GR enzymes were only decreased in heat stress-exposed seedlings, whereas melatonin pretreatment elevated the AsA content as a result of increased the activities of APX, MDHAR, DHAR, and GR enzymes under heat stress. All of these enzymes actively contributed to the AsA–GSH cycle, which converts the tiny non-enzymatic molecules AsA and GSH [58]. The AsA values depend upon metabolizing (APX) and recycling (MDHAR, DHAR) enzyme activity. Moreover, at the time of ROS detoxification, DHAR oxidizes GSH to GSSG. Simultaneously, GR recycles GSH. Therefore, we concluded that melatonin pretreatment might have the potential to reduce oxidative damage by inducing the AsA–GSH cycle [59, 60]. To elucidate the inherent mechanisms, we quantified the expression of several related genes. Under heat stress, RBOH expression was upregulated and the expression level was further magnified in melatonin-pretreated heat-stressed plants (Fig. 7a). In melatonin-pretreated heat-stressed seedlings, the relative transcript abundance of enzymatic (SOD, CAT, POD) and non-enzymatic (APX, GR, MDHAR, DHAR, GST) antioxidant genes were upregulated (Figs. 4 and 6), which indicates that plants were more stable under heat stress because of excess ROS scavenging; these findings are consistent with those of previous research done on kiwifruit [42], watermelon [52], apple [61], and Arabidopsis [62] under various abiotic stress conditions.
PAs play a critical role in plant signaling transduction that is beneficial for counteracting the effects of different capricious environments [27]. Some previous studies determined that melatonin has a positive regulatory effects on plant development and abiotic stress (alkaline stress, cold, thermal, oxidative, and iron deficiency tolerance) management by interacting with the PAs signaling pathway [28, 63]. Melatonin might ameliorate the thermal oxidative stress by interacting with the PAs and NO biosynthesis pathways. The exogenous application of melatonin elevated the endogenous free PAs level. Similarly, expression levels of different PAs biosynthesis genes were also upregulated in melatonin-pretreated heat-stressed seedlings. The transcript abundances of ADC1/2, SAMDC1/2, SPMS, and SPDS1/2/3/5/6 were upregulated (Fig. 9), and that of PAO1/2 was downregulated in melatonin-pretreated heat-stressed seedlings, and these genes are associated with Put, Spd, and Spm biosynthesis. These findings indicate that melatonin and PAs metabolism have close interactions. This finding is also similar to that of previous research performed on various crops under different stresses [30, 32–34, 64]. Alam et al. [65] concluded that long-term heat-stressed seedlings treated with melatonin adjusted through the modulation of PAs metabolism.
Melatonin along with NO has the potential to combat different stress conditions through the L-arginine and PAs metabolic pathways [66]. However, the NOS and NR pathways are also regulated via PAs [67, 68]. Our current data also highlight that the NO content, NR activity, and NOS-like activity along with the expression of their related genes were elevated in melatonin-pretreated heat-stressed tomato seedlings (Fig. 10), which indicates that melatonin triggered the NO activity [30]. Overall, melatonin enhanced mitigation of heat-induced damage through coordination with PA- and NO-mediated signaling pathways.
Conclusions
To determine how melatonin mitigated heat stress-induced adverse effects in tomato seedlings, we described a probable mechanism (Fig. 11). We observed that 100 μM exogenous melatonin treatment improved the thermal tolerance of tomato seedlings by lowering ROS (H2O2, O2•−, MDA) production, enhanced antioxidant enzyme activity, AsA–GSH cycle modulation, and upregulation of antioxidant-related gene expression. Additionally, melatonin elevates endogenous PAs via upregulation of PAs biosynthesis genes. NO content along with NR and NOS activity were also increased with melatonin supplementation. Therefore, we concluded that heat stress-induced damage was suppressed by melatonin, which coordinates with the PAs and NO biosynthesis pathways, which helps to detoxify the overaccumulated ROS. These findings provide novel insight into the cross-talk that exists among melatonin, PAs, and NO to inhibit thermal stress. To better understand this phenomenon, further investigation is needed to determine how these three molecules collectively function to alleviate the heat-stress induced damage.
Fig. 11.
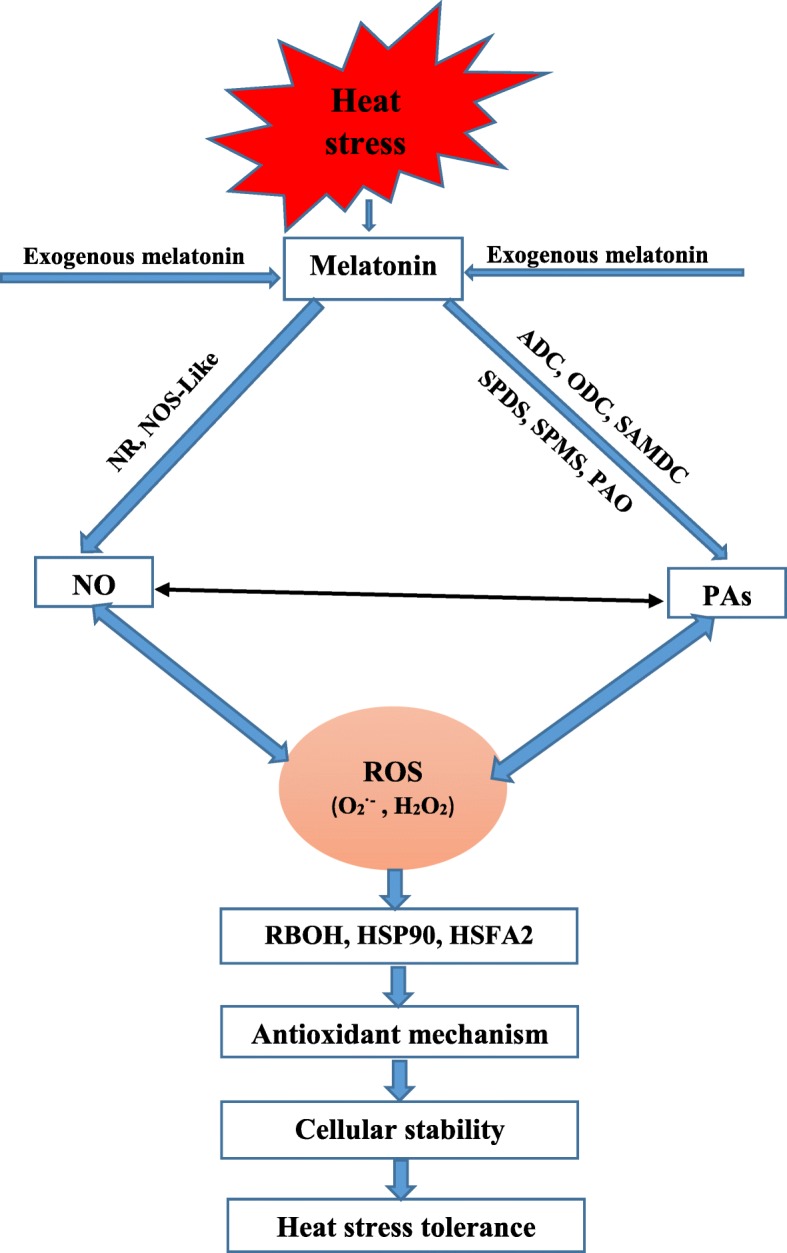
The schematic representation of possible mechanism of melatonin mediated high temperature stress tolerance in tomato seedlings via the PAs and NO biosynthesis pathway. PAs: polyamines; ADC: arginine decarboxylase; ODC: ornithine decarboxylase; SPDS: spermidine synthase; SPMS: spermine synthase; SAMDC: s-adenosyl methionine decarboxylase; PAO: polyamine oxidase; NO: Nitric oxide; NR: nitrate reductase; NOS: nitric oxide synthase; ROS: reactive oxygen species; H2O2: hydrogen peroxide; O2•−: superoxide anion; RBOH: Respiratory burst oxidase; HSFA2: heat shock transcription factors A2; HSP90: heat shock protein 90
Methods
Plant material and growth conditions
Tomato (Solanum lycopersicum L. Cv. Hezuo 903) seeds (Shanghai Tomato Research Institute, Shanghai, China) were sorted by uniform size and then sterilized by 0.1% sodium hypochloride (NaOCl) for 5 min, followed by washing several times with deionized water; then, they were placed in dark conditions for 36 h at 28 ± 1 °C for germination. Germinated seeds were then sown in plastic trays (41 × 41 × 5 cm) that contained a peat and vermiculite (2:1, v:v) mixture and cultured in a growth chamber at Nanjing Agriculture University, where the environmental conditions were maintained at 28 ± 1 °C (day) and 19 ± 1 °C (night), relative humidity from 65 to 75%, and 12 h photoperiods (PAR 300 μmol m− 2 s− 1). After the second true leaf was fully expanded, the uniformly grown seedlings were selected and transferred into containers filled with a peat and vermiculite (2,1, v, v) mixture and watered on alternate days with full-strength Hoagland solution.
Treatment and sampling
When the fourth true leaves were fully developed, the seedlings were divided into two sub-groups for challenge under different treatments. Melatonin was applied as described in previous experiment performed by Martinez et al. [69]. In the first sub-group, 80 mL of 100 μM melatonin was sprayed on each tomato seedling leaves each day and for 7 days; in the second sub-group, each tomato seedling leaves were sprayed with the same volume of water. Melatonin stock solution was prepared by dissolving melatonin in ddH2O with 0.01% v/v Tween-20 used as a surfactant. One week after treatment, half of the melatonin-treated seedlings and half of the water-sprayed seedlings were separated and exposed to a high temperature (42 °C) for 24 h [10]. After 24 h of heat treatment, leaves were harvested for subsequent analysis and immediately stored at − 80 °C.
Measurement of growth indicators
To assess the combined effects of melatonin and heat-stressed in tomato seedlings, we measured different growth indicators such as fresh and dry weight of leaves and roots. Fresh weight of leaves and roots were measured by electric balance. For dry weight records plants were oven dried (80 °C for 72 h).
Histochemical detection of H2O2 and O2•−
H2O2 and O2•− generation rate were detected using 3,3- diamino benzidine (DAB) and nitro blue tetrazolium (NBT), respectively, using a previously described method [70] with minor modification. For H2O2 localization, stained leaves were placed in vacuum along with 0.5 mg·mL− 1 fresh DAB solution prepared by 25 mM Tris-HCl (pH 3.8) and kept for 12 h at room temperature. Brown spots appeared on the surface of leaves because of the reaction between DAB and H2O2. For O2•− detection, the other leaf samples were immersed with 1 mg·mL− 1 NBT solution, which was made with 10 mM phosphate buffer (pH 7.8), and incubated at room temperature in the dark for 12 h. Blue spots were also present on leaves because of the reaction of NBT and O2•−. Both of the stained leaf samples were bleached by boiling in 95% ethanol for 20 min to remove chlorophyll. Then, the samples were placed into absolute ethanol for several hours before taking photos with a digital camera.
Determination of H2O2 production level
The H2O2 concentration in leaves was estimated by slightly modifying a method described by Velikova et al. [71]. First, 0.2 g leaves were homogenized with 1.6 mL 0.1% trichloroacetic acid (TCA) in an ice bath for 30 min and centrifuged at 12000×g for 20 min at 4 °C. Then, 0.5 mL 0.1 M potassium phosphate buffer (pH 7.8) and 1 mL 1 M KI (Potassium Iodine) were added to 0.5 mL supernatant and kept in a dark place for 1 h. The absorbance was measured at 390 nm. Finally, the H2O2 content was quantified with a standard curve and expressed as μmol g− 1 FW.
Determination of O2•− production rate
The O2•− generation rate was determined following the procedure reported by Nahar et al. [72] with some alterations. Briefly, 0.2 g leaves were homogenized with 2 mL 50 mM phosphate buffer (pH 7.8) and centrifuged at 12000×g for 20 min at 4 °C. Then, 0.5 mL 50 mM phosphate buffer (pH 7.8) and 0.1 mL 10 mM hydroxylamine hydrochloride were mixed in 0.5 mL supernatant and incubated at room temperature for 30 min. After incubation, 1 mL 17 mM sulfanilamide and 1 mL of 7 mM naphthylamine were added to the mixture and incubated for 30 min. The absorbance reading of the mixture was measured at 530 nm. O2•− production was then calculated with a standard curve of NaNO2 and expressed as nmol g− 1 min− 1 FW.
Membrane injury index (MII) measurement
Membrane injury index (MII) of leaves was computed to the method outlined by Jahan et al. [10] with few modifications. Briefly, 0.5 g fresh leaves were thoroughly washed with deionized water, cut into small pieces, put into tubes filled with 20 mL deionized water, and placed at room temperature for 4–5 h under dark conditions in a shaker; then, the initial electrical conductivity (EC1) in the bathing solution was determined by a portable conductivity meter (DDS-307, Shanghai Precision and Scientific Instrument LTD., Shanghai, China). Subsequently, the samples were boiled at 95 °C for 20 min and cooled to room temperature, and the final electrical conductivity (EC2) was measured in the bathing solution. Simultaneously, we determined the deionized water conductivity (EC0). The MII was calculated as follows:
Lipid peroxidation measurement
Lipid peroxidation was inferred based on MDA content in leaves, which was measured as described by Alexieva et al. [73] with slight adjustments. First, 0.2 g leaf samples were homogenized in a 1.6 mL 0.1% (w/v) TCA solution and centrifuged at 4 °C for 20 min at 12000×g. From the supernatant, a 1.0-mL aliquot was added to 1.0 mL TCA containing 0.67% TBA; then, the sample was boiled at 95 °C for 15 min and kept on ice for cooling. Subsequently, the mixture was centrifuged at 4400×g for 10 min. Then, MDA content was measured at 532 nm and 600 nm by a spectrophotometer (Evolution 300, Thermo Fisher Scientific, Waltham, MA, USA).
Proline content determination
The proline content was evaluated following the method described by Bates et al. [74]. Fresh leaf samples (0.2 g) were digested in 3% sulphosalicylic acid followed by centrifugation at 12000×g for 20 min at 4 °C. The same amount of glacial acetic acid and ninhydrin solutions were incorporated in the supernatant and incubated for 30 min. Consequently, the sample was heated at 100 °C for 1 h, and 5 mL toluene was added after cooling. Toluene absorbance was read at 520 nm by a spectrophotometer (Spectronic 20D, Milton Roy, Philadelphia, PA, USA).
Relative water content measurement
The relative water content (RWC) was calculated using the method established by Barrs and Weatherley [75] with some changes. Fully developed leaves were arbitrarily detached from treated plants and immediately weighed as FW, followed by soaking in distilled water and incubation for 6 h at room temperature. Then, the excess surface water was removed with a paper towel, and the turgid weight (TW) was recorded. Leaf samples were then oven dried at 80 °C for 72 h to obtain the dry weight (DW). RWC was calculated using the following equation:
Leaf enzymes activity assays
Fresh leaf samples (0.2 g) were digested with a chilled pestle and mortar in 1.6 mL 50 mM pre-cooled phosphate buffer (pH 7.8), and supernatants were obtained by centrifugation of the homogenate at 12000×g for 20 min at 4 °C. The supernatants were then used to estimate the antioxidant enzymes activities.
SOD activity (EC 1.15.1.1) was calculated using a modified version of the protocol described by Giannopolitis and Ries [76], and Maresca et al. [77] described the procedure that was used to estimate POD (EC 1.11.1.7) activity. Briefly, a 40-μL enzyme extract was added to a 3-mL reaction mixture that contained 14.5 mM Met, 30 μM EDTA–Na2 solution, 50 mM phosphate buffer (pH 7.8), 2.25 mM NBT solution, and 60 μM riboflavin solution. The SOD content was monitored at 560 nm. For POD activity assay, 40 μL enzyme solution was mixed with a 3-mL reaction volume that included 0.2 M phosphate buffer (pH 6.0), 50 mM guaiacol, and 2% H2O2 solution, and absorbance was quantified at 470 nm.
To determine catalase (CAT, EC 1.11.1.6) activity, the protocol described by Dhindsa et al. [78] was used. Briefly, a 0.1 mL enzyme solution was added followed by a 3-mL reaction mixture that contained 0.15 M phosphate buffer (pH 7.0) and 0.3% H2O2 solution. The level of activity was calculated at 240 nm.
To examine APX (EC 1.11.1.11) activity, the method described by Nakano and Asada [79] was used. Briefly, a 1.6-mL assay mixture that consisted of 50 mM phosphate buffer (pH 7.0), 0.1 mM EDTA–Na2, 5 mM AsA, and 20 mM H2O2 were added to 0.1 mL enzyme solution. The activity was calculated at 290 nm.
AsA content was quantified as previously reported by Logan et al. [80]. Briefly, 0.1 g leaf samples were homogenized in 1.5 mL 6% pre-chilled HClO4. After grinding, the sample was centrifuged at 12000×g for 15 min at 4 °C, and supernatant was collected for further analysis. For neutralization, 200 mM sodium acetate buffer (pH 5.6) was added to the supernatant and AsA was assayed at 265 nm; the absorbance reading was recorded before and after incubation of the supernatant in 1.5 units of AsA oxidase for 15 min.
GSH was assayed with the protocol priorly described by Griffith [81]. Briefly, the leaf sample (0.1 g) was ground in 1.5 mL 5% sulfosalicylic acid, and the homogenized sample was centrifuged at 4 °C for 20 min at 12000×g. The supernatant was neutralized with 200 mM sodium acetate buffer (pH 5.6). Then, 5, 5-dithiobis-(2-nitrobenzoic acid) was incorporated for enzymatic recycling of GSH. The GSH content was calculated by recording the absorbance at 412 nm with a spectrometer.
MDHAR (EC 1.6.5.4) activity was determined based on the change (due to NADPH oxidation) in absorbance at 340 nm, as previously described by Hossain et al. [82].
DHAR (EC 1.8.5.1) activity was assayed based on the change in absorbance at 265 nm, as described by Nakano and Asada [79].
Glutathione S-transferase (GST) activity was assayed with GST detection kit (Solarbio Life Science, Beijing, China) following the manufacturer’s instructions. First, fresh leaves (0.1 g) were ground with extraction buffer (1 mL) in an ice bath and homogenized by centrifugation at 4 °C for 10 min at 8000×g with the supernatant used for testing GST. GST activity was calculated using the molar extinction coefficient 9.6 × 103 Lmol− 1 cm− 1.
Glutathione reductase (GR) activity was determined with GR detection kit (Solarbio Life Science, Beijing, China) following the manufacturer’s instructions. Briefly, 0.1 g leaf tissue was taken and homogenized in 1 mL extraction solution in an ice bath and centrifuged at 10000×g for 10 min at 4 °C, and the supernatant was used to determine GR activity. To calculate GR, 6.22 × 103 L mol− 1 cm− 1 was used as the extinction coefficient.
Nitrate reductase (NR) activity was measured with NR detection kit (Solarbio Life Science, Beijing, China) following the manufacturer’s instructions. First, 0.1 g fresh leaf samples were gently washed and the water was removed from the leaf surface. Samples were incubated for 2 h in work solution under dark condition in room temperature and then kept at − 20 °C for 30 min. Then, samples were taken, ground in an ice bath with 1 mL extract solution, and centrifuged at 4000×g for 10 min; the supernatant was used to determine NR activity.
Nitric oxide synthase (NOS) activity was assayed by NOS detection kit (Solarbio Life Science, Beijing, China) following the manufacturer’s instructions.
NO content was quantified using NO detection kit (Solarbio Life Science, Beijing, China) according to the manufacturer’s protocol.
Protein extraction
The protein content was determined using Bovine serum albumin (BSA) as the standard following the method described by Bradford [83].
Determination of endogenous free polyamines
Endogenous free polyamines content were assayed as the approaches reported by Shen et al. [84] with minor modifications. Briefly, 0.5 g leaf tissue was homogenized in 5% (v/v) cold perchloric acid and incubated on ice for 1 h. Then, homogenates were centrifuged for 20 min at 12000×g and the upper supernatant was used to determine the free PAs. A 0.7 mL aliquot was reacted with 1.4 mL NaOH (2 N) and 15 μL benzoyl chloride, and then gently vortexed the mixer and incubated for 30 min at 37 °C. Later, to stop the reaction, 2 mL saturated NaCl was added to the solution. To extract benzoyl PAs, 2 mL cold diethyl ether was mixed into the solution, which was then centrifuged at 3000×g for 5 min. The extracted benzoyl PAs were evaporated to dryness and then re-dissolved in 1 mL of 64% (v/v) methanol. To separate and analyze the PAs content, we used UHPLC (Ultimate 3000, Thermo Scientific, San Jose, CA, USA) with a C18 reversed-phase column at a flow rate of 0.8 mL min− 1.
Total RNA extraction and quantitative real-time PCR analysis
Total RNA was extracted from 0.1 g tomato leaves tissues using the RNAsimple Total RNA Kit (TIANGEN, Beijing, China) according to the manufacturer’s instructions. One microgram of total RNA was reverse-transcribed into cDNA using a SuperScript First-strand Synthesis System for quantitative real-time PCR based on the manufacturer’s instructions (Takara, Tokyo, Japan). The gene-specific primers were designed using DNA sequences from the NCBI database (https://www.ncbi.nlm.nih.gov/), and Sol Genomics Network (solgenomics.net) and the primer pair sequences are listed in Additional file 1: Table S1. Real-time PCR was performed on a StepOnePlus™ Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) with ChamQ Universal SYBR qPCR Master Mix (Vazyme Biotech Co., Ltd., Nanjing, China). The total reaction system volume was 20 μL, which consisted of 10 μL ChamQ SYBR qPCR Master Mix (2×), 0.4 μL ROX reference dye 1 (50×), 2 μL template cDNA (10×), 0.8 μL each specific primer (10 μM), and 6 μL sterilized ddH2O. Three biological replicates were performed for each reaction and the cycling conditions were as follows: 95 °C for 5 min, followed by 40 cycles of denaturation at 95 °C for 15 s and annealing at 60 °C for 1 min, and a final extension at 95 °C for 15 s. Relative expression was calculated using the 2−ΔΔCt formula [85], and the mRNA expression level was normalized against actin (used as an internal control) and compared.
Statistical analysis
At least five independent biological replicates were performed for each treatment, and three replicates were performed for the whole experiment. All of the data were statistically analyzed with SPSS 20.0 (SPSS Inc., Chicago, IL, USA). One way analysis of variance was performed, and statistically significant differences among the treatments were determined using Tukey’s honest significant difference test at P < 0.05. A transcript expression heatmap was created using the TBtools statistic package. Origin Pro 8.0 was used to make graphs.
Additional files
Table S1. List of primers used for qRT-PCR assays. (DOCX 29 kb)
Acknowledgments
We thanks to Dr.Mallory Eckstut for critical reading and editing of this manuscript.
Abbreviations
- 1O2
Singlet oxygen
- ADC
Arginine decarboxylase
- APX
Ascorbate peroxidase
- AsA
Ascorbate
- BSA
Bovine serum albumin
- CAT
Catalase
- DHAR
Dehydroascorbate
- DNA
Deoxy ribo nucleic acid
- EC
Electrical conductivity
- GR
Glutathione reductase
- GSH
Glutathione
- GST
Glutathione S-transferase
- H2O2
Hydrogen peroxide
- HsfA2
Heat shock transcription factors A2
- HSP90
Heat shock protein 90
- HSPs
Heat shock proteins
- MDA
Malondialdehyde
- MDHAR
Monodehydroascorbate reductase
- MII
Membrane injury index
- NO
Nitric oxide
- NOS
Nitric oxide synthase
- NR
Nitrate reductase
- O2•−
Superoxide anion
- ODC
Ornithine decarboxylase
- OH•
Hydroxyl radical
- P5CS
Delta 1-pyrroline-5-carboxylate synthetase
- PAO
Polyamine oxidase
- PAs
Polyamines
- POD
Peroxidase
- PUT
Putrescine
- RBOH
Respiratory burst oxidase
- RNA
Ribo nucleic acid
- ROS
Reactive oxygen species
- RWC
Relative water content
- SAMDC
S-adenosyl methionine decarboxylase
- SOD
Superoxide dismutase
- SPD
Spermidine
- SPDS
Spermidine synthase
- SPM
Spermine
- SPMS
Spermine synthase
Authors’ contributions
SRG conceived and designed the experiments. MSJ conducted the experiments and wrote the manuscript. ZC, MH, MT contributed supplied materials, data collection, analysis tools and analyzed the data. YW, SS and JS supervised and revised the manuscript. All authors reviewed and approved the manuscript.
Funding
This work was financially supported by the National Natural Science Foundation of China (31672199 and 31801902), the China Earmarked Fund for Modern Agro-industry Technology Research System (CARS-23-B12), and Jiangsu Province Scientific and Technological Achievements into Special Fund (BE2017701). The supporters did not play any role in the design, collection, analysis, interpretation of the relevant data, or in writing the manuscript.
Availability of data and materials
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing of interest.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Mohammad Shah Jahan and Sheng Shu contributed equally to this work.
Contributor Information
Mohammad Shah Jahan, Email: shahjahansau@gmail.com.
Sheng Shu, Email: shusheng@njau.edu.cn.
Yu Wang, Email: ywang@njau.edu.cn.
Zheng Chen, Email: 1310235651@qq.com.
Mingming He, Email: 2017204050@njau.edu.cn.
Meiqi Tao, Email: 810998804@qq.com.
Jin Sun, Email: jinsun@njau.edu.cn.
Shirong Guo, Email: srguo@njau.edu.cn.
References
- 1.Ohama N, Sato H, Shinozaki K, Yamaguchi-Shinozaki K. Transcriptional regulatory network of plant heat stress response. Trends Plant Sci. 2017;22(1):53–65. doi: 10.1016/j.tplants.2016.08.015. [DOI] [PubMed] [Google Scholar]
- 2.Hasanuzzaman M, Nahar K, Alam M, Roychowdhury R, Fujita M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int J Mol Sci. 2013;14(5):9643–9684. doi: 10.3390/ijms14059643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lazar D, Murch SJ, Beilby MJ, Al Khazaaly S. Exogenous melatonin affects photosynthesis in characeae Chara australis. Plant Signal Behav. 2013;8(3):e23279. doi: 10.4161/psb.23279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Salvucci ME, Crafts-Brandner SJ. Relationship between the heat tolerance of photosynthesis and the thermal stability of rubisco activase in plants from contrasting thermal environments. Plant Physiol. 2004;134(4):1460–1470. doi: 10.1104/pp.103.038323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vasseur F, Pantin F, Vile D. Changes in light intensity reveal a major role for carbon balance in Arabidopsis responses to high temperature. Plant Cell Environ. 2011;34(9):1563–1576. doi: 10.1111/j.1365-3040.2011.02353.x. [DOI] [PubMed] [Google Scholar]
- 6.Mishkind M, Vermeer JE, Darwish E, Munnik T. Heat stress activates phospholipase D and triggers PIP2 accumulation at the plasma membrane and nucleus. Plant J. 2009;60(1):10–21. doi: 10.1111/j.1365-313X.2009.03933.x. [DOI] [PubMed] [Google Scholar]
- 7.Narayanan S, Tamura PJ, Roth MR, Prasad PV, Welti R. Wheat leaf lipids during heat stress: I. high day and night temperatures result in major lipid alterations. Plant Cell Environ. 2016;39(4):787–803. doi: 10.1111/pce.12649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mittler R, Vanderauwera S, Suzuki N, Miller G, Tognetti VB, Vandepoele K, Gollery M, Shulaev V, Van Breusegem F. ROS signaling: the new wave? Trends Plant Sci. 2011;16(6):300–309. doi: 10.1016/j.tplants.2011.03.007. [DOI] [PubMed] [Google Scholar]
- 9.Allakhverdiev SI, Kreslavski VD, Klimov VV, Los DA, Carpentier R, Mohanty P. Heat stress: an overview of molecular responses in photosynthesis. Photosyn Res. 2008;98(1–3):541. doi: 10.1007/s11120-008-9331-0. [DOI] [PubMed] [Google Scholar]
- 10.Jahan MS, Wang Y, Shu S, Zhong M, Chen Z, Wu J, Sun J, Guo S. Exogenous salicylic acid increases the heat tolerance in tomato (Solanum lycopersicum L) by enhancing photosynthesis efficiency and improving antioxidant defense system through scavenging of reactive oxygen species. Sci Hortic. 2019;247:421–429. doi: 10.1016/j.scienta.2018.12.047. [DOI] [Google Scholar]
- 11.Li H, LIU SS, Yi CY, Wang F, Zhou J, Xia XJ, Shi K, Zhou YH, Yu JQ. Hydrogen peroxide mediates abscisic acid-induced HSP 70 accumulation and heat tolerance in grafted cucumber plants. Plant Cell Environ. 2014;37(12):2768–2780. doi: 10.1111/pce.12360. [DOI] [PubMed] [Google Scholar]
- 12.Ahammed Golam Jalal, Xu Wen, Liu Airong, Chen Shuangchen. Endogenous melatonin deficiency aggravates high temperature-induced oxidative stress in Solanum lycopersicum L. Environmental and Experimental Botany. 2019;161:303–311. doi: 10.1016/j.envexpbot.2018.06.006. [DOI] [Google Scholar]
- 13.Nishizawa A, Yabuta Y, Yoshida E, Maruta T, Yoshimura K, Shigeoka S. Arabidopsis heat shock transcription factor A2 as a key regulator in response to several types of environmental stress. Plant J. 2006;48(4):535–547. doi: 10.1111/j.1365-313X.2006.02889.x. [DOI] [PubMed] [Google Scholar]
- 14.Arnao MB, Hernández-Ruiz J. Melatonin and its relation-ship to plant hormones. Ann Bot. 2018;121(2):195–207. doi: 10.1093/aob/mcx114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang Y, Reiter RJ, Chan Z. Phytomelatonin: a universal abiotic stress regulator. J Exp Bot. 2017;69(5):963–974. doi: 10.1093/jxb/erx473. [DOI] [PubMed] [Google Scholar]
- 16.Arnao Marino B., Hernández-Ruiz Josefa. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends in Plant Science. 2019;24(1):38–48. doi: 10.1016/j.tplants.2018.10.010. [DOI] [PubMed] [Google Scholar]
- 17.Li X, Wei J-P, Scott E, Liu J-W, Guo S, Li Y, Zhang L, Han W-Y. Exogenous melatonin alleviates cold stress by promoting antioxidant defense and redox homeostasis in Camellia sinensis L. Molecules. 2018;23(1):165. doi: 10.3390/molecules23010165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wei Y, Chang Y, Zeng H, Liu G, He C, Shi H. RAV transcription factors are essential for disease resistance against cassava bacterial blight via activation of melatonin biosynthesis genes. J Pineal Res. 2018;64(1):e12454. doi: 10.1111/jpi.12454. [DOI] [PubMed] [Google Scholar]
- 19.Tiryaki I, Keles H. Reversal of the inhibitory effect of light and high temperature on germination of Phacelia tanacetifolia seeds by melatonin. J Pineal Res. 2012;52(3):332–339. doi: 10.1111/j.1600-079X.2011.00947.x. [DOI] [PubMed] [Google Scholar]
- 20.Arnao MB, Hernández-Ruiz J. Melatonin promotes adventitious-and lateral root regeneration in etiolated hypocotyls of Lupinus albus L. J Pineal Res. 2007;42(2):147–152. doi: 10.1111/j.1600-079X.2006.00396.x. [DOI] [PubMed] [Google Scholar]
- 21.Zhang R, Sun Y, Liu Z, Jin W, Sun Y. Effects of melatonin on seedling growth, mineral nutrition, and nitrogen metabolism in cucumber under nitrate stress. J Pineal Res. 2017;62(4):e12403. doi: 10.1111/jpi.12403. [DOI] [PubMed] [Google Scholar]
- 22.Shi H, Tan DX, Reiter RJ, Ye T, Yang F, Chan Z. Melatonin induces class A1 heat-shock factors (HSFA 1s) and their possible involvement of thermotolerance in Arabidopsis. J Pineal Res. 2015;58(3):335–342. doi: 10.1111/jpi.12219. [DOI] [PubMed] [Google Scholar]
- 23.Rodriguez C, Mayo JC, Sainz RM, Antolín I, Herrera F, Martín V, Reiter RJ. Regulation of antioxidant enzymes: a significant role for melatonin. J Pineal Res. 2004;36(1):1–9. doi: 10.1046/j.1600-079X.2003.00092.x. [DOI] [PubMed] [Google Scholar]
- 24.Li H, He J, Yang X, Li X, Luo D, Wei C, Ma J, Zhang Y, Yang J, Zhang X. Glutathione-dependent induction of local and systemic defense against oxidative stress by exogenous melatonin in cucumber (Cucumis sativus L.) J Pineal Res. 2016;60(2):206–216. doi: 10.1111/jpi.12304. [DOI] [PubMed] [Google Scholar]
- 25.Wang P, Yin L, Liang D, Li C, Ma F, Yue Z. Delayed senescence of apple leaves by exogenous melatonin treatment: toward regulating the ascorbate–glutathione cycle. J Pineal Res. 2012;53(1):11–20. doi: 10.1111/j.1600-079X.2011.00966.x. [DOI] [PubMed] [Google Scholar]
- 26.Wu J, Shu S, Li C, Sun J, Guo S. Spermidine-mediated hydrogen peroxide signaling enhances the antioxidant capacity of salt-stressed cucumber roots. Plant Physiol Biochem. 2018;128:152–162. doi: 10.1016/j.plaphy.2018.05.002. [DOI] [PubMed] [Google Scholar]
- 27.Gill SS, Tuteja N. Polyamines and abiotic stress tolerance in plants. Plant Signal Behav. 2010;5(1):26–33. doi: 10.4161/psb.5.1.10291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ke Q, Ye J, Wang B, Ren J, Yin L, Deng X, Wang S. Melatonin mitigates salt stress in wheat seedlings by modulating polyamine metabolism. Front Plant Sci. 2018;9:914. doi: 10.3389/fpls.2018.00914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Groppa M, Benavides M. Polyamines and abiotic stress: recent advances. Amino Acids. 2008;34(1):35. doi: 10.1007/s00726-007-0501-8. [DOI] [PubMed] [Google Scholar]
- 30.Zhou C, Liu Z, Zhu L, Ma Z, Wang J, Zhu J. Exogenous melatonin improves plant iron deficiency tolerance via increased accumulation of polyamine-mediated nitric oxide. Int J Mol Sci. 2016;17(11):1777. doi: 10.3390/ijms17111777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Shi H, Jiang C, Ye T, Tan D-X, Reiter RJ, Zhang H, Liu R, Chan Z. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass [Cynodon dactylon (L). Pers.] by exogenous melatonin. J Exp Bot. 2014;66(3):681–694. doi: 10.1093/jxb/eru373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lei XY, Zhu RY, Zhang GY, Dai YR. Attenuation of cold-induced apoptosis by exogenous melatonin in carrot suspension cells: the possible involvement of polyamines. J Pineal Res. 2004;36(2):126–131. doi: 10.1046/j.1600-079X.2003.00106.x. [DOI] [PubMed] [Google Scholar]
- 33.Cao S, Song C, Shao J, Bian K, Chen W, Yang Z. Exogenous melatonin treatment increases chilling tolerance and induces defense response in harvested peach fruit during cold storage. J Agric Food Chem. 2016;64(25):5215–5222. doi: 10.1021/acs.jafc.6b01118. [DOI] [PubMed] [Google Scholar]
- 34.Zhao H, Zhang K, Zhou X, Xi L, Wang Y, Xu H, Pan T, Zou Z. Melatonin alleviates chilling stress in cucumber seedlings by up-regulation of CsZat12 and modulation of polyamine and abscisic acid metabolism. Sci Rep. 2017;7(1):4998. doi: 10.1038/s41598-017-05267-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pál M, Szalai G, Janda T. Speculation: polyamines are important in abiotic stress signaling. Plant Sci. 2015;237:16–23. doi: 10.1016/j.plantsci.2015.05.003. [DOI] [PubMed] [Google Scholar]
- 36.Liang D, Gao F, Ni Z, Lin L, Deng Q, Tang Y, Wang X, Luo X, Xia H. Melatonin improves heat tolerance in kiwifruit seedlings through promoting antioxidant enzymatic activity and glutathione S-transferase transcription. Molecules. 2018;23(3):584. doi: 10.3390/molecules23030584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Li H, Chang J, Chen H, Wang Z, Gu X, Wei C, Zhang Y, Ma J, Yang J, Zhang X. Exogenous melatonin confers salt stress tolerance to watermelon by improving photosynthesis and redox homeostasis. Front Plant Sci. 2017;8:295. doi: 10.3389/fpls.2017.00295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhao H, Ye L, Wang Y, Zhou X, Yang J, Wang J, Cao K, Zou Z. Melatonin increases the chilling tolerance of chloroplast in cucumber seedlings by regulating photosynthetic electron flux and the ascorbate-glutathione cycle. Front Plant Sci. 2016;7:1814. doi: 10.3389/fpls.2016.01814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Li C, Wang P, Wei Z, Liang D, Liu C, Yin L, Jia D, Fu M, Ma F. The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J Pineal Res. 2012;53(3):298–306. doi: 10.1111/j.1600-079X.2012.00999.x. [DOI] [PubMed] [Google Scholar]
- 40.Pieri C, Marra M, Moroni F, Recchioni R, Marcheselli F. Melatonin: a peroxyl radical scavenger more effective than vitamin E. Life Sci. 1994;55(15):PL271–PL276. doi: 10.1016/0024-3205(94)00666-0. [DOI] [PubMed] [Google Scholar]
- 41.García JJ, López-Pingarrón L, Almeida-Souza P, Tres A, Escudero P, García-Gil FA, Tan DX, Reiter RJ, Ramírez JM, Bernal-Pérez M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: a review. J Pineal Res. 2014;56(3):225–237. doi: 10.1111/jpi.12128. [DOI] [PubMed] [Google Scholar]
- 42.Liang D, Ni Z, Xia H, Xie Y, Lv X, Wang J, Lin L, Deng Q, Luo X. Exogenous melatonin promotes biomass accumulation and photosynthesis of kiwifruit seedlings under drought stress. Sci Hortic. 2019;246:34–43. doi: 10.1016/j.scienta.2018.10.058. [DOI] [Google Scholar]
- 43.Hasan M, Ahammed GJ, Yin L, Shi K, Xia X, Zhou Y, Yu J, Zhou J. Melatonin mitigates cadmium phytotoxicity through modulation of phytochelatins biosynthesis, vacuolar sequestration, and antioxidant potential in Solanum lycopersicum L. Front Plant Sci. 2015;6:601. doi: 10.3389/fpls.2015.00601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Miller G, Mittler R. Could heat shock transcription factors function as hydrogen peroxide sensors in plants? Ann Bot. 2006;98(2):279–288. doi: 10.1093/aob/mcl107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Katano K, Honda K, Suzuki N. Integration between ROS regulatory systems and other signals in the regulation of various types of heat responses in plants. Int J Mol Sci. 2018;19(11):3370. doi: 10.3390/ijms19113370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Driedonks N, Xu J, Peters JL, Park S, Rieu I. Multi-level interactions between heat shock factors, heat shock proteins, and the redox system regulate acclimation to heat. Front Plant Sci. 2015;6:999. doi: 10.3389/fpls.2015.00999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Manchester LC, Coto-Montes A, Boga JA, Andersen LPH, Zhou Z, Galano A, Vriend J, Tan DX, Reiter RJ. Melatonin: an ancient molecule that makes oxygen metabolically tolerable. J Pineal Res. 2015;59(4):403–419. doi: 10.1111/jpi.12267. [DOI] [PubMed] [Google Scholar]
- 48.Reiter RJ, Mayo JC, Tan DX, Sainz RM, Alatorre-Jimenez M, Qin L. Melatonin as an antioxidant: under promises but over delivers. J Pineal Res. 2016;61(3):253–278. doi: 10.1111/jpi.12360. [DOI] [PubMed] [Google Scholar]
- 49.Arnao MB, Hernández-Ruiz J. Functions of melatonin in plants: a review. J Pineal Res. 2015;59(2):133–150. doi: 10.1111/jpi.12253. [DOI] [PubMed] [Google Scholar]
- 50.Reiter R, Tan D-X, Zhou Z, Cruz M, Fuentes-Broto L, Galano A. Phytomelatonin: assisting plants to survive and thrive. Molecules. 2015;20(4):7396–7437. doi: 10.3390/molecules20047396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hardeland R. Melatonin in plants and other phototrophs: advances and gaps concerning the diversity of functions. J Exp Bot. 2014;66(3):627–646. doi: 10.1093/jxb/eru386. [DOI] [PubMed] [Google Scholar]
- 52.Nawaz MA, Jiao Y, Chen C, Shireen F, Zheng Z, Imtiaz M, Bie Z, Huang Y. Melatonin pretreatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating melatonin biosynthesis and antioxidant-related gene expression. J Plant Physiol. 2018;220:115–127. doi: 10.1016/j.jplph.2017.11.003. [DOI] [PubMed] [Google Scholar]
- 53.Alscher RG, Erturk N, Heath LS. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J Exp Bot. 2002;53(372):1331–1341. doi: 10.1093/jexbot/53.372.1331. [DOI] [PubMed] [Google Scholar]
- 54.Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002;7(9):405–410. doi: 10.1016/S1360-1385(02)02312-9. [DOI] [PubMed] [Google Scholar]
- 55.Li X, Brestic M, Tan DX, Zivcak M, Zhu X, Liu S, Song F, Reiter RJ, Liu F. Melatonin alleviates low PS I-limited carbon assimilation under elevated CO 2 and enhances the cold tolerance of offspring in chlorophyll b-deficient mutant wheat. J Pineal Res. 2018;64(1):e12453. doi: 10.1111/jpi.12453. [DOI] [PubMed] [Google Scholar]
- 56.Meng JF, Xu TF, Wang ZZ, Fang YL, Xi ZM, Zhang ZW. The ameliorative effects of exogenous melatonin on grape cuttings under water-deficient stress: antioxidant metabolites, leaf anatomy, and chloroplast morphology. J Pineal Res. 2014;57(2):200–212. doi: 10.1111/jpi.12159. [DOI] [PubMed] [Google Scholar]
- 57.Ding F, Liu B, Zhang S. Exogenous melatonin ameliorates cold-induced damage in tomato plants. Sci Hortic. 2017;219:264–271. doi: 10.1016/j.scienta.2017.03.029. [DOI] [Google Scholar]
- 58.Noctor G, Foyer CH. Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Biol. 1998;49(1):249–279. doi: 10.1146/annurev.arplant.49.1.249. [DOI] [PubMed] [Google Scholar]
- 59.Bonnefont-Rousselot D, Collin F, Jore D, Gardès-Albert M. Reaction mechanism of melatonin oxidation by reactive oxygen species in vitro. J Pineal Res. 2011;50(3):328–335. doi: 10.1111/j.1600-079X.2010.00847.x. [DOI] [PubMed] [Google Scholar]
- 60.Maresca V, Sorbo S, Keramat B, Basile A. Effects of triacontanol on ascorbate-glutathione cycle in Brassica napus L. exposed to cadmium-induced oxidative stress. Ecotoxicol Environ Saf. 2017;144:268–274. doi: 10.1016/j.ecoenv.2017.06.035. [DOI] [PubMed] [Google Scholar]
- 61.Wang P, Sun X, Li C, Wei Z, Liang D, Ma F. Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J Pineal Res. 2013;54(3):292–302. doi: 10.1111/jpi.12017. [DOI] [PubMed] [Google Scholar]
- 62.Zheng X, Tan DX, Allan AC, Zuo B, Zhao Y, Reiter RJ, Wang L, Wang Z, Guo Y, Zhou J. Chloroplastic biosynthesis of melatonin and its involvement in protection of plants from salt stress. Sci Rep. 2017;7:41236. doi: 10.1038/srep41236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zhan H, Nie X, Zhang T, Li S, Wang X, Du X, Tong W, Song W. Melatonin: a small molecule but important for salt stress tolerance in plants. Int J Mol Sci. 2019;20(3):709. doi: 10.3390/ijms20030709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Zhang Q, Liu X, Zhang Z, Liu N, Li D, Hu L. Melatonin improved waterlogging tolerance in alfalfa (Medicago sativa) by reprogramming polyamine and ethylene metabolism. Front Plant Sci. 2019;10:44. [DOI] [PMC free article] [PubMed]
- 65.Alam MN, Zhang L, Yang L, Islam MR, Liu Y, Luo H, Yang P, Wang Q, Chan Z. Transcriptomic profiling of tall fescue in response to heat stress and improved thermotolerance by melatonin and 24-epibrassinolide. BMC Genomics. 2018;19(1):224. doi: 10.1186/s12864-018-4588-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Gupta KJ, Fernie AR, Kaiser WM, van Dongen JT. On the origins of nitric oxide. Trends Plant Sci. 2011;16(3):160–168. doi: 10.1016/j.tplants.2010.11.007. [DOI] [PubMed] [Google Scholar]
- 67.Rosales EP, Iannone MF, Groppa MD, Benavides MP. Polyamines modulate nitrate reductase activity in wheat leaves: involvement of nitric oxide. Amino Acids. 2012;42(2–3):857–865. doi: 10.1007/s00726-011-1001-4. [DOI] [PubMed] [Google Scholar]
- 68.Shi H, Chan Z. Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J Integr Plant Biol. 2014;56(2):114–121. doi: 10.1111/jipb.12128. [DOI] [PubMed] [Google Scholar]
- 69.Martinez V, Nieves-Cordones M, Lopez-Delacalle M, Rodenas R, Mestre T, Garcia-Sanchez F, Rubio F, Nortes P, Mittler R, Rivero R. Tolerance to stress combination in tomato plants: new insights in the protective role of melatonin. Molecules. 2018;23(3):535. doi: 10.3390/molecules23030535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Shi J, Fu X-Z, Peng T, Huang X-S, Fan Q-J, Liu J-H. Spermine pretreatment confers dehydration tolerance of citrus in vitro plants via modulation of antioxidative capacity and stomatal response. Tree Physiol. 2010;30(7):914–922. doi: 10.1093/treephys/tpq030. [DOI] [PubMed] [Google Scholar]
- 71.Velikova V, Yordanov I, Edreva A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines. Plant Sci. 2000;151(1):59–66. doi: 10.1016/S0168-9452(99)00197-1. [DOI] [Google Scholar]
- 72.Nahar K, Hasanuzzaman M, Alam MM, Fujita M. Exogenous glutathione confers high temperature stress tolerance in mung bean (Vigna radiata L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environ Exp Bot. 2015;112:44–54. doi: 10.1016/j.envexpbot.2014.12.001. [DOI] [Google Scholar]
- 73.Alexieva V, Sergiev I, Mapelli S, Karanov E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001;24(12):1337–1344. doi: 10.1046/j.1365-3040.2001.00778.x. [DOI] [Google Scholar]
- 74.Bates LS, Waldren RP, Teare I. Rapid determination of free proline for water-stress studies. Plant Soil. 1973;39(1):205–207. doi: 10.1007/BF00018060. [DOI] [Google Scholar]
- 75.Barrs H, Weatherley P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust J Biol Sci. 1962;15(3):413–428. doi: 10.1071/BI9620413. [DOI] [Google Scholar]
- 76.Giannopolitis CN, Ries SK. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977;59(2):309–314. doi: 10.1104/pp.59.2.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Nickel KS, Cunningham B. Improved peroxidase assay method using leuco 2, 3′, 6-trichloroindophenol and application to comparative measurements of peroxidatic catalysis. Anal Biochem. 1969;27(2):292–299. doi: 10.1016/0003-2697(69)90035-9. [DOI] [PubMed] [Google Scholar]
- 78.Dhindsa RS, Plumb-Dhindsa PL, Reid DM. Leaf senescence and lipid peroxidation: effects of some phytohormones, and scavengers of free radicals and singlet oxygen. Physiol Plant. 1982;56(4):453–457. doi: 10.1111/j.1399-3054.1982.tb04539.x. [DOI] [Google Scholar]
- 79.Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981;22(5):867–880. [Google Scholar]
- 80.Logan BA, Demmig-Adam B, Adams WW., III Antioxidants and xanthophyll cycle-dependent energy dissipation in Cucurbita pepo L. and Vinca major L. upon a sudden increase in growth PPFD in the field. J Exp Bot. 1998;49(328):1881–1888. doi: 10.1093/jxb/49.328.1881. [DOI] [Google Scholar]
- 81.Griffith OW. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal Biochem. 1980;106(1):207–212. doi: 10.1016/0003-2697(80)90139-6. [DOI] [PubMed] [Google Scholar]
- 82.Hossain MA, Nakano Y, Asada K. Monodehydroascorbate reductase in spinach chloroplasts and its participation in regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984;25(3):385–395. [Google Scholar]
- 83.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72(1–2):248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 84.Shen J-l, Wang Y, Shu S, Jahan MS, Zhong M, Wu J-q, Sun J, Guo S-r. Exogenous putrescine regulates leaf starch overaccumulation in cucumber under salt stress. Sci Hortic. 2019;253:99–110. doi: 10.1016/j.scienta.2019.04.010. [DOI] [Google Scholar]
- 85.Wang Y, Cai SY, Yin LL, Shi K, Xia XJ, Zhou YH, Yu JQ, Zhou J. Tomato HsfA1a plays a critical role in plant drought tolerance by activating ATG genes and inducing autophagy. Autophagy. 2015;11(11):2033–2047. doi: 10.1080/15548627.2015.1098798. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. List of primers used for qRT-PCR assays. (DOCX 29 kb)
Data Availability Statement
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.